
K48-Linked Ubiquitination Contributes to Nicotine-Augmented Bone Marrow-Derived Dendritic-Cell-Mediated Adaptive Immunity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mice
2.2. Reagents and Antibodies
2.3. Murine Bone Marrow Precursor Cell-Derived Dendritic Cell Preparation
2.4. Cell Culture and Cell Line
2.5. Ubiquitin siRNA Transfection
2.6. Murine Bone Marrow Precursor Cell-Derived Dendritic Cell Treatment
2.7. Murine Bone Marrow-Derived Dendritic Cell Adoptive Transfer
2.8. Antigen-Specific Interferon-γ Enzyme-Linked Immunospot Assay
2.9. Labeling Cells with CellTrace Carboxyfluorescein Diacetate Succinimidyl Ester
2.10. Cytotoxic T Lymphocyte Cytolytic Assay
2.11. Co-Immunoprecipitation and Immunoprecipitation
2.12. Western Blots
2.13. Confocal Immunofluorescent Microscope
2.14. Statistical Analysis
3. Results
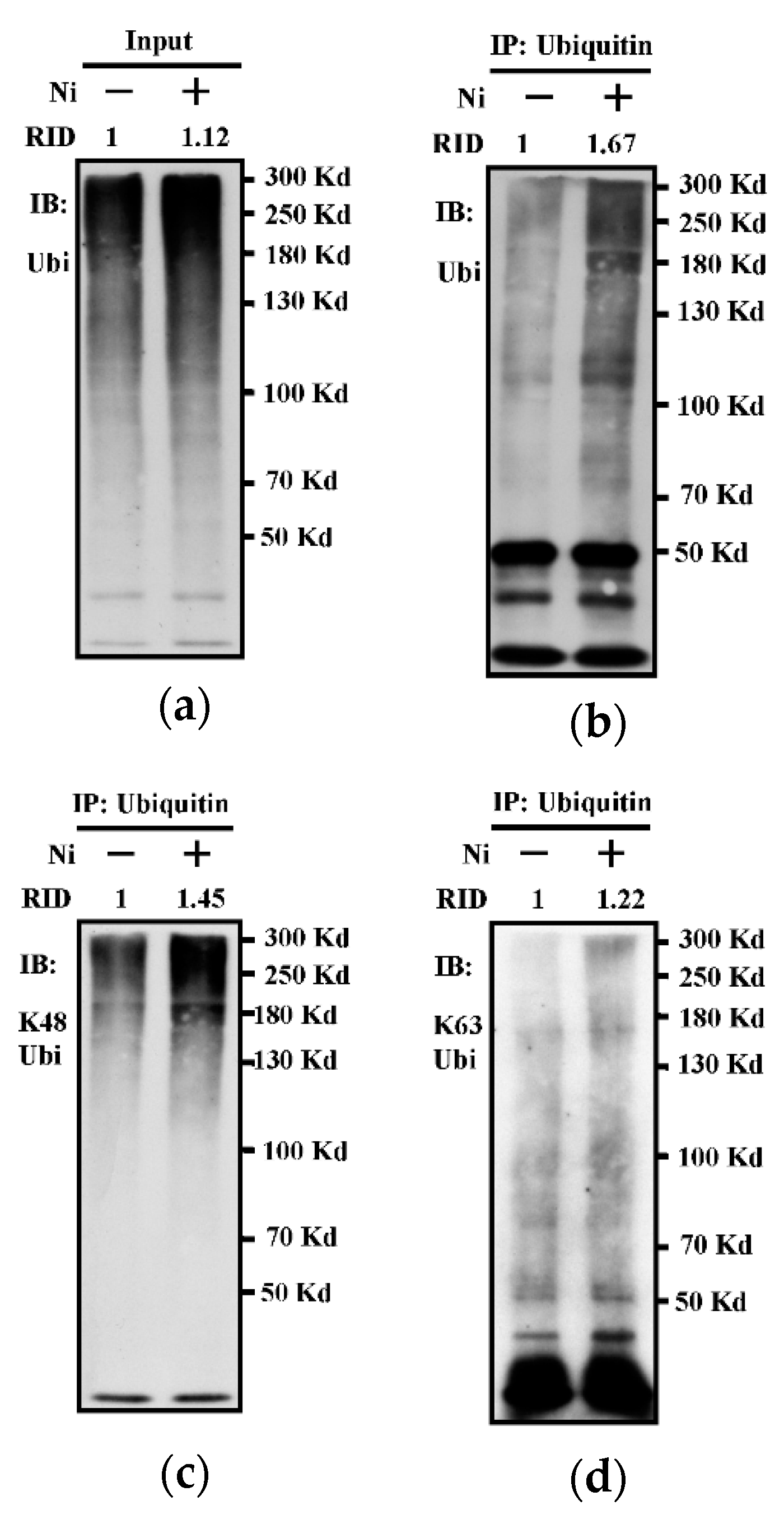
3.1. The Treatment with Nicotine Increases K48-linked Ubiquitination in Bone Marrow-Derived Dendritic Cells
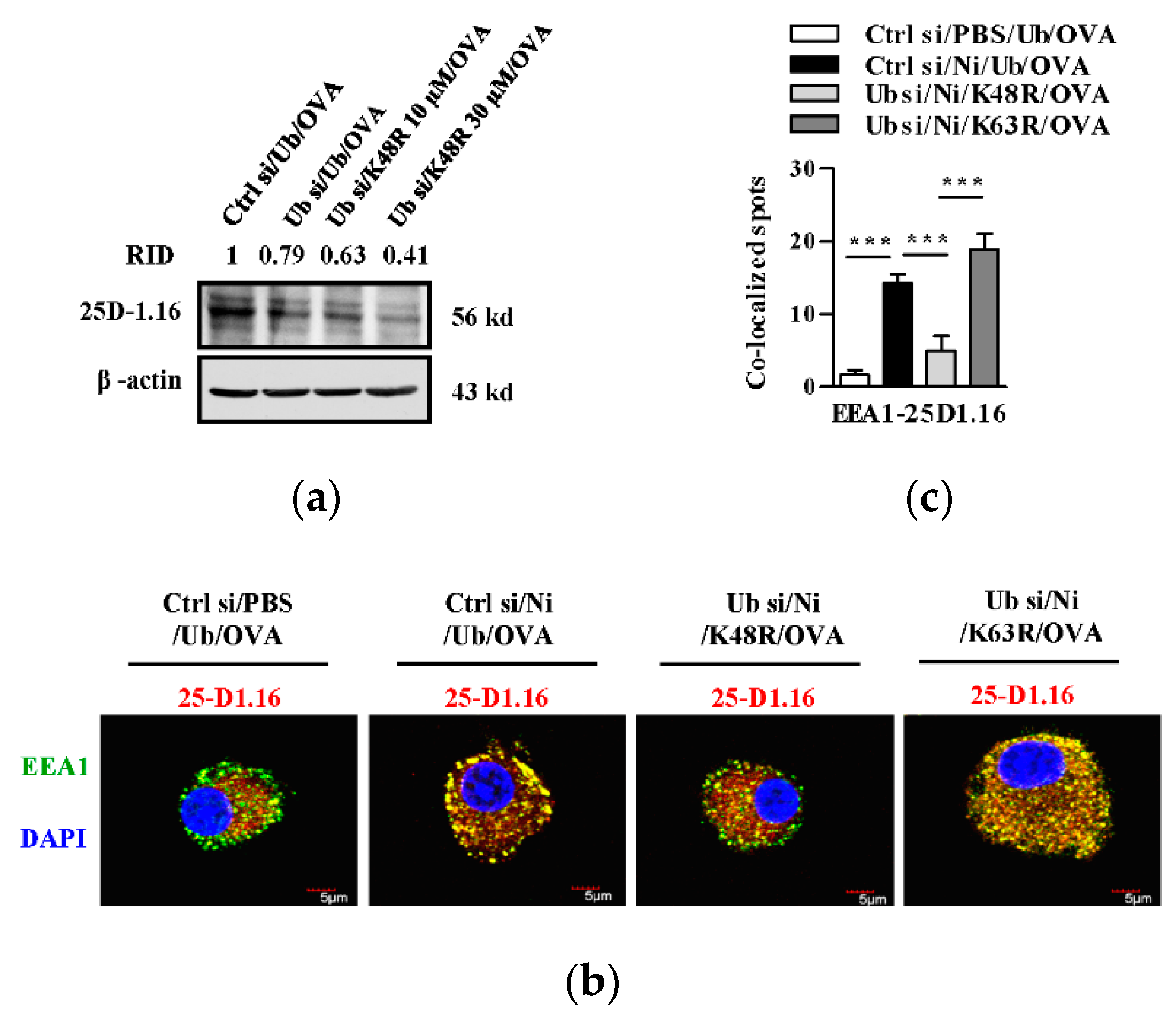
3.2. K48-linked Ubiquitination Contributes to Nicotine-Increased Cross-Presentation in Bone Marrow-Derived Dendritic Cells
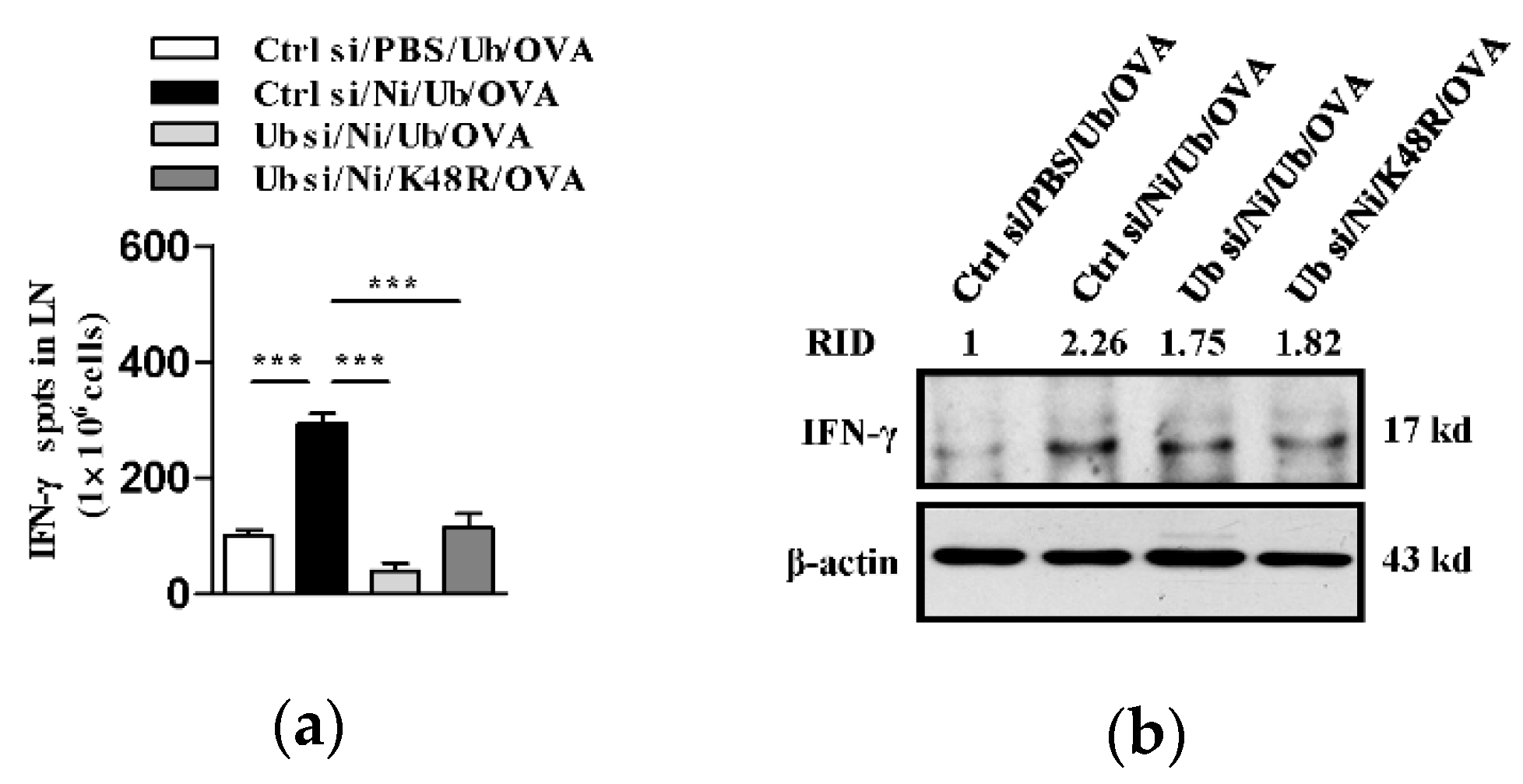
3.3. K48-Linked Ubiquitination Is Essential For Nicotine-Augmented Dendritic Cell-Based CTL Priming
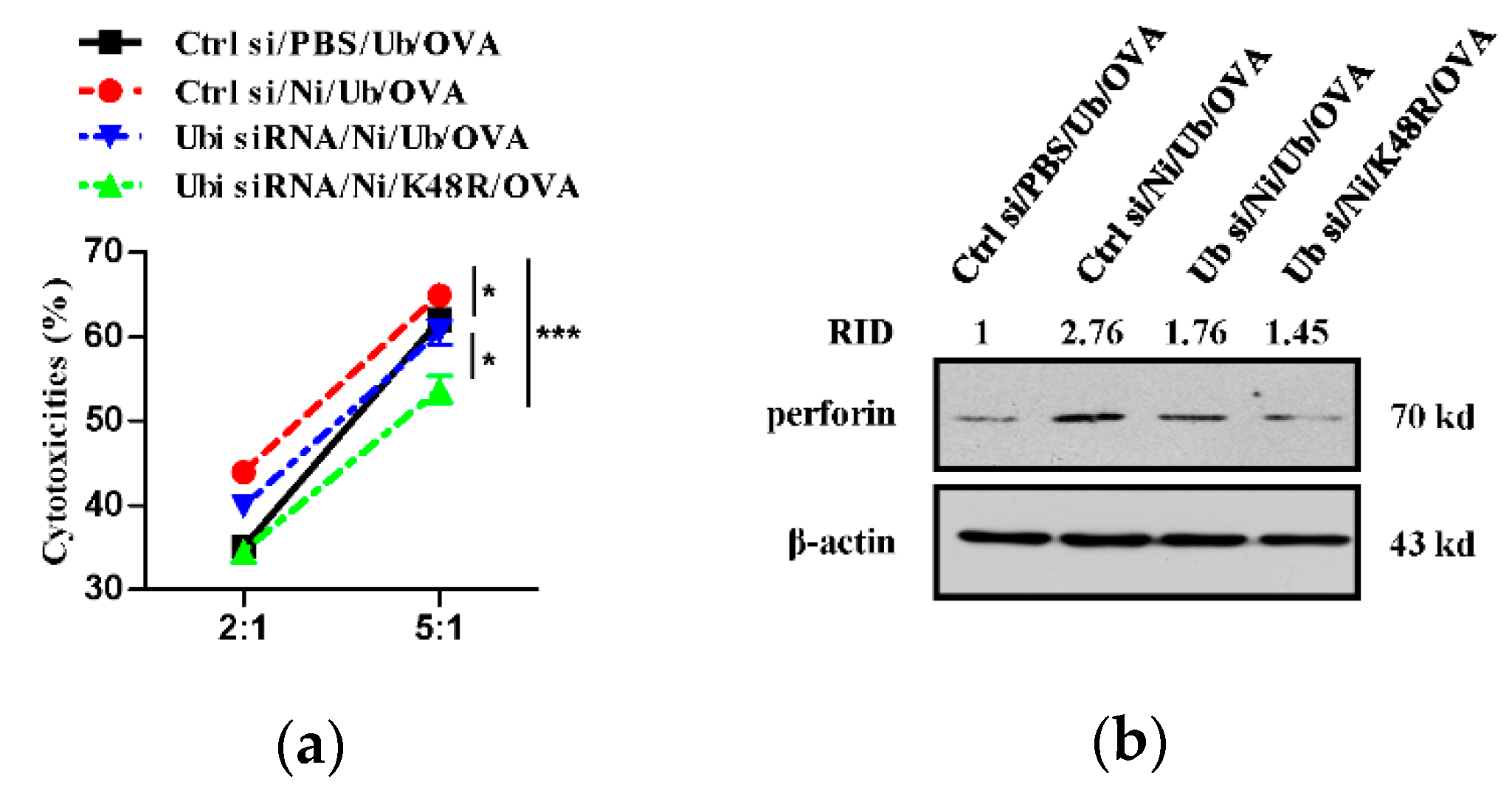
3.4. K48-Linked Ubiquitination Contributes to Nicotine-Increased Superior Cytolytic Capacity of Dendritic Cell-Activated T Cells
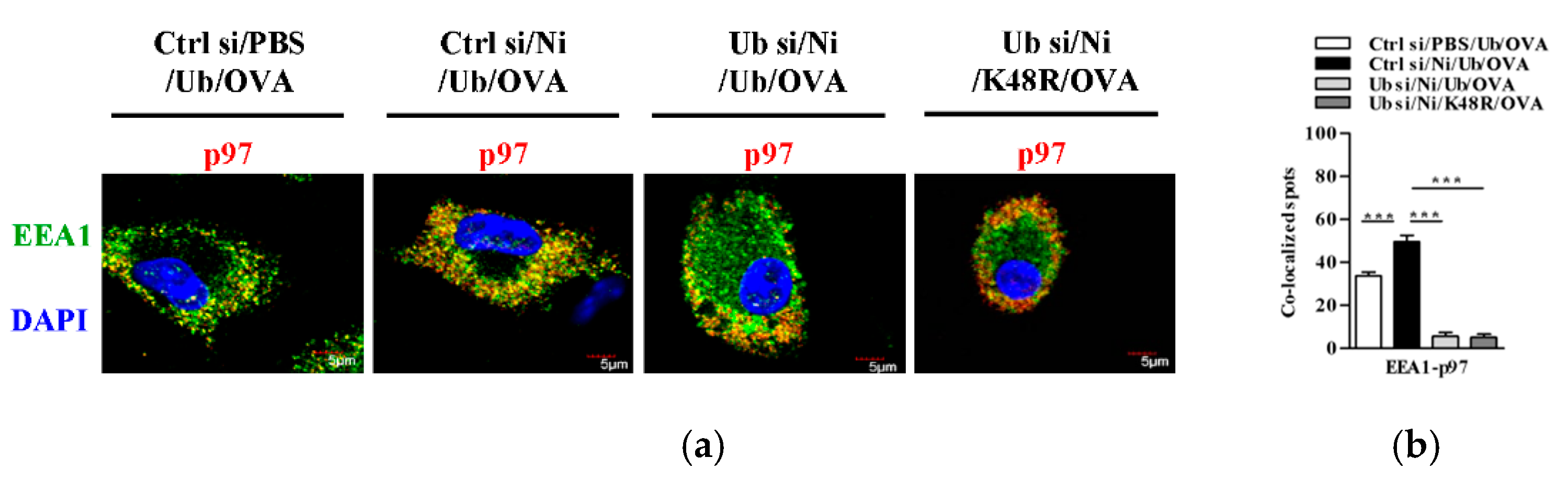
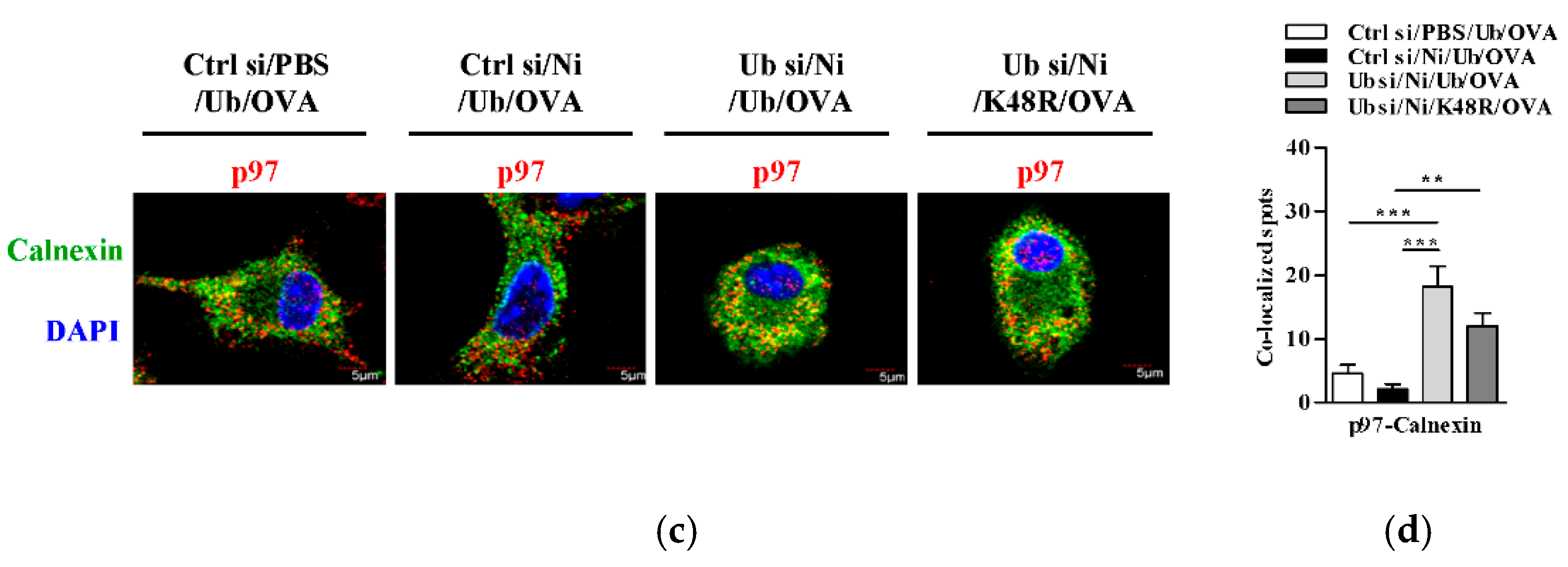
3.5. K48-Linked Ubiquitination Facilitetes Nicotine-Increased Endosomal Relocation of p97 in Dendritic Cells
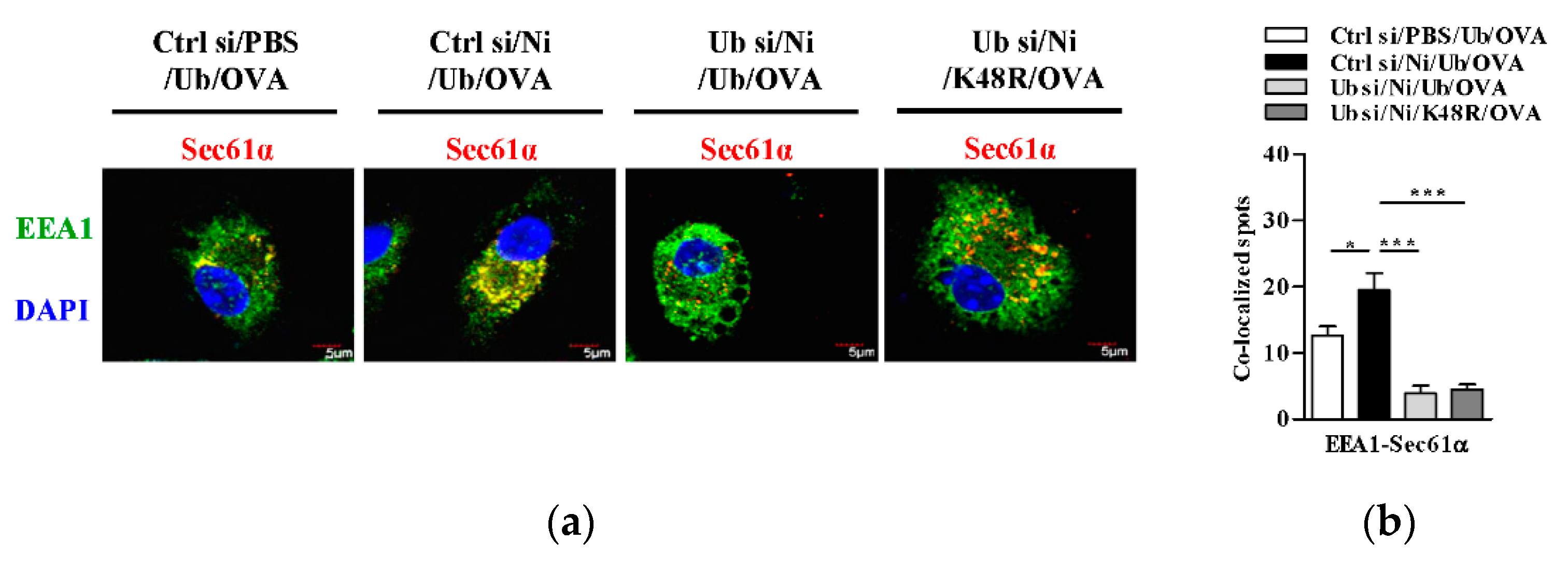
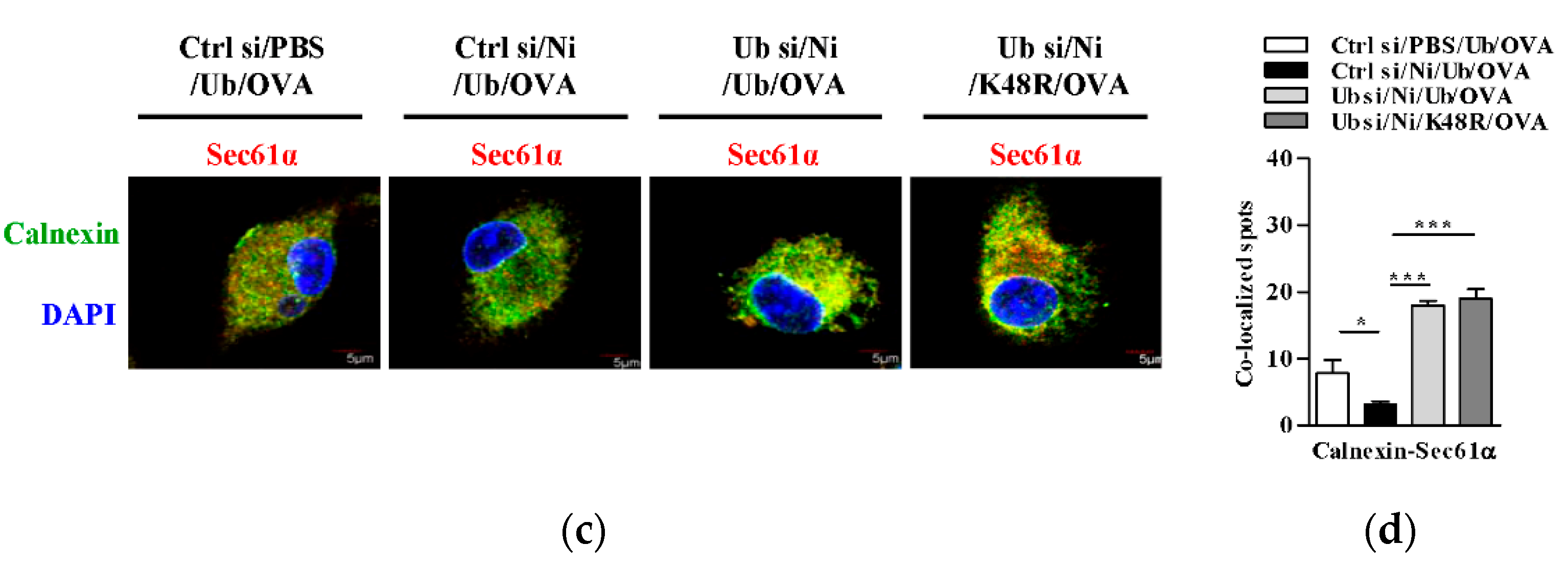
3.6. K48-Linked Ubiquitination Contributes to Nicotine-Increased Endosomal Relocation of Sec61 in Dendritic Cells
3.7. Nicotine-Increased K48-Linked Ubiquitination of Mannose Receptor in Dendritic Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Villadangos, J.A.; Heath, W.R.; Carbon, F.R. Outside looking in: The inner workings of the cross-presentation pathway within dendritic cells. Trends Immunol. 2007, 28, 45–47. [Google Scholar] [CrossRef]
- Wessler, I.K.; Kirkpatrick, C.J. The non-neuronal cholinergic system: An emerging drug target in the airways. Pulm. Pharmacol. Ther. 2001, 14, 423–434. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Lei, B.; Yuan, Y.; Zhang, L.; Hu, L.; Jin, S.; Kang, B.; Liao, X.; Sun, W.; Xu, F.; et al. Brain control of humoral immune responses amenable to behavioural modulation. Nature 2020, 581, 204–208. [Google Scholar] [CrossRef]
- Gao, F.G.; Wan, D.F.; Gu, J.R. Ex vivo nicotine stimulation augments the efficacy of therapeutic bone marrow-derived dendritic cell vaccination. Clin. Cancer Res. 2007, 13, 3706–3712. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.G.; Li, H.T.; Li, Z.J.; Gu, J.R. Nicotine stimulated dendritic cells could achieve anti-tumor effects in mouse lung and liver cancer. J. Clin. Immunol. 2011, 31, 80–88. [Google Scholar] [CrossRef]
- Jin, H.J.; Li, H.T.; Sui, H.X.; Xue, M.Q.; Wang, Y.N.; Wang, J.X.; Gao, F.G. Nicotine stimulated bone marrow-derived dendritic cells could augment HBV specific CTL priming by activating PI3K-Akt pathway. Immunol. Lett. 2012, 146, 40–49. [Google Scholar] [CrossRef]
- Jin, H.J.; Sui, H.X.; Wang, Y.N.; Gao, F.G. Nicotine up-regulated 4-1BBL expression by activating Mek-PI3K pathway augments the efficacy of bone marrow-derived dendritic cell vaccination. J. Clin. Immunol. 2013, 33, 246–254. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Yang, Y.W.; You, X.; Deng, X.Q.; Hu, C.F.; Zhu, C.; Wang, J.Y.; Gu, J.J.; Wang, Y.N.; Li, Q.; et al. Ex vivo nicotine stimulation augments the efficacy of human peripheral blood mononuclear cell-derived dendritic cell vaccination via activating Akt-S6 pathway. Anal. Cell Pathol. 2015, 2015, 741487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, N.; Shabek, N. Ubiquitin ligases: Structure, function, and regulation. Annu. Rev. Biochem. 2017, 86, 129–157. [Google Scholar] [CrossRef]
- Zhang, M.X.; Cai, Z.; Zhang, M.; Wang, X.M.; Wang, Y.; Zhao, F.; Zhou, J.; Luo, M.H.; Zhu, Q.; Xu, Z.; et al. USP20 promotes cellular antiviral responses via deconjugating K48-linked ubiquitination of MITA. J. Immunol. 2019, 202, 2397–2406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goldszmid, R.S.; Coppens, I.; Lev, A.; Caspar, P.; Mellman, I.; Sher, A. Host ER-parasitophorous vacuole interaction provides a route of entry for antigen cross-presentation in Toxoplasma gondii-infected dendritic cells. J. Exp. Med. 2009, 206, 399–410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rezvani, K.; Teng, Y.; Shim, D.; de Biasi, M. Nicotine regulates multiple synaptic proteins by inhibiting proteasomal activity. J. Neurosci. 2007, 27, 10508–10519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, X.; Han, C.; Wen, J.; Guo, X.; Zhang, K. Nicotine increases apoptosis in HUVECs cultured in high glucose/high fat via Akt ubiquitination and degradation. Clin. Exp. Pharmacol. Physiol. 2018, 45, 198–204. [Google Scholar] [CrossRef]
- Joffre, O.P.; Segura, E.; Savina, A.; Amigorena, S. Cross-presentation by dendritic cells. Nat. Rev. Immunol. 2012, 12, 557–569. [Google Scholar] [CrossRef]
- Embgenbroich, M.; Burgdorf, S. Current Concepts of Antigen Cross-Presentation. Front. Immunol. 2018, 9, 1643. [Google Scholar] [CrossRef] [Green Version]
- Zehner, M.; Burgdorf, S. Regulation of antigen transport into the cytosol for cross-presentation by ubiquitination of the mannose receptor. Mol. Immunol. 2013, 55, 146–148. [Google Scholar] [CrossRef] [PubMed]
- Grotzke, J.E.; Kozik, P.; Morel, J.D.; Impens, F.; Pietrosemoli, N.; Cresswell, P.; Amigorena, S.; Demangel, C. Sec61 blockade by mycolactone inhibits antigen cross-presentation independently of endosome-to-cytosol export. Proc. Natl. Acad. Sci. USA 2017, 114, E5910–E5919. [Google Scholar] [CrossRef] [Green Version]
- Zehner, M.; Marschall, A.L.; Bos, E.; Schloetel, J.G.; Kreer, C.; Fehrenschild, D.; Limmer, A.; Ossendorp, F.; Lang, T.; Koster, A.J.; et al. The translocon protein Sec61 mediates antigen transport from endosomes in the cytosol for cross-presentation to CD8(+) T cells. Immunity 2015, 42, 850–863. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.D.; Hu, C.F.; You, X.; Lu, N.N.; Gao, F.G. Akt+ IKKα/β+ Rab5+ signalosome mediate the endosomal recruitment of Sec61 and contribute to cross-presentation in bone marrow precursor cells. Vaccines 2020, 8, 539. [Google Scholar] [CrossRef]
- Weck, M.M.; Grünebach, F.; Werth, D.; Sinzger, C.; Bringmann, A.; Brossart, P. TLR ligands differentially affect uptake and presentation of cellular antigens. Blood 2007, 109, 3890–3894. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.Y.; Hu, C.F.; Li, J.; You, X.; Gao, F.G. Increased translocation of antigens to endosomes and TLR4 mediated endosomal recruitment of TAP contribute to nicotine augmented cross-presentation. Oncotarget 2016, 7, 38451–38466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, X.; Xu, D.D.; Zhang, D.; Chen, J.; Gao, F.G. PYR-41 and Thalidomide Impair Dendritic Cell Cross-Presentation by Inhibiting Myddosome Formation and Attenuating the Endosomal Recruitments of p97 and Sec61 via NF-κB Inactivation. J. Immunol. Res. 2018, 2018, 5070573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.X.; Chen, R.L.; Liao, X.Y.; You, X.; Gao, F.G. Ex vivo IL-15 replenishment augments bone marrow precursor cell-mediated adaptive immunity via PI3K-Akt pathway. J. Leukoc. Bio. 2020, 108, 177–188. [Google Scholar] [CrossRef]
- Wang, F.; Wang, Y.Y.; Li, J.; You, X.; Qiu, X.H.; Wang, Y.N.; Gao, F.G. Increased antigen presentation but impaired T cells priming after upregulation of interferon-beta induced by lipopolysaccharides is mediated by upregulation of B7H1 and GITRL. PLoS ONE 2014, 9, e105636. [Google Scholar] [CrossRef]
- Rudd-Schmidt, J.A.; Trapani, J.A.; Voskoboinik, I. Distinguishing perforin-mediated lysis and granzyme-dependent apoptosis. Methods Enzymol. 2019, 629, 291–306. [Google Scholar]
- Burgdorf, S.; Kautz, A.; Böhnert, V.; Knolle, P.A.; Kurts, C. Distinct pathways of antigen uptake and intracellular routing in CD4 and CD8 T cell activation. Science 2007, 316, 612–616. [Google Scholar] [CrossRef]
- Wan, C.; Wu, M.; Zhang, S.; Chen, Y.; Lu, C. α7nAChR-mediated recruitment of PP1γ promotes TRAF6/NF-κB cascade to facilitate the progression of Hepatocellular Carcinoma. Mol. Carcinog. 2018, 57, 1626–1639. [Google Scholar] [CrossRef] [PubMed]
- Grumati, P.; Dikic, I. Ubiquitin signaling and autophagy. J. Biol. Chem. 2018, 293, 5404–5413. [Google Scholar] [CrossRef] [Green Version]
- Lin, M.; Zhao, Z.; Yang, Z.; Meng, Q.; Tan, P.; Xie, W.; Qin, Y.; Wang, R.F.; Cui, J. USP38 inhibits type I interferon signaling by editing TBK1 ubiquitination through NLRP4 signalosome. Mol. Cell 2016, 64, 267–281. [Google Scholar] [CrossRef] [Green Version]
- Massaly, N.; Francès, B.; Moulédous, L. Roles of the ubiquitin proteasome system in the effects of drugs of abuse. Front. Mol. Neurosci. 2015, 7, 99. [Google Scholar] [CrossRef] [Green Version]
- Jang, H.D.; Hwang, H.Z.; Kim, H.S.; Lee, S.Y. C-Cbl negatively regulates TRAF6-mediated NF-kappaB activation by promoting K48-linked polyubiquitination of TRAF6. Cell Mol. Biol. Lett. 2019, 24, 29. [Google Scholar] [CrossRef]
- Teng, Y.; Rezvani, K.; de Biasi, M. UBXN2A regulates nicotinic receptor degradation by modulating the E3 ligase activity of CHIP. Biochem. Pharmacol. 2015, 97, 518–530. [Google Scholar] [CrossRef] [PubMed]
- Figueroa, C.; Gálvez-Cancino, F.; Oyarce, C.; Contreras, F.; Prado, C.; Valeria, C.; Cruz, S.; Lladser, A.; Pacheco, R. Inhibition of dopamine receptor D3 signaling in dendritic cells increases antigen cross-presentation to CD8(+) T-cells favoring anti-tumor immunity. J. Neuroimmunol. 2017, 303, 99–107. [Google Scholar] [CrossRef]
- Di Chiara, G. Role of dopamine in the behavioural actions of nicotine related to addiction. Eur. J. Pharmacol. 2000, 393, 295–314. [Google Scholar] [CrossRef]
- Zhang, M.; Harrison, E.; Biswas, L.; Tran, T.; Liu, X. Menthol facilitates dopamine-releasing effect of nicotine in rat nucleus accumbens. Pharmacol. Biochem. Behav. 2018, 175, 47–52. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, C.F.; Liao, X.Y.; Xu, D.D.; Ruan, Y.B.; Gao, F.G. K48-Linked Ubiquitination Contributes to Nicotine-Augmented Bone Marrow-Derived Dendritic-Cell-Mediated Adaptive Immunity. Vaccines 2021, 9, 278. https://doi.org/10.3390/vaccines9030278
Hu CF, Liao XY, Xu DD, Ruan YB, Gao FG. K48-Linked Ubiquitination Contributes to Nicotine-Augmented Bone Marrow-Derived Dendritic-Cell-Mediated Adaptive Immunity. Vaccines. 2021; 9(3):278. https://doi.org/10.3390/vaccines9030278
Chicago/Turabian StyleHu, Chun Fang, Xiao Yan Liao, Dan Dan Xu, Yi Bin Ruan, and Feng Guang Gao. 2021. "K48-Linked Ubiquitination Contributes to Nicotine-Augmented Bone Marrow-Derived Dendritic-Cell-Mediated Adaptive Immunity" Vaccines 9, no. 3: 278. https://doi.org/10.3390/vaccines9030278