
Immunity against Lagovirus europaeus and the Impact of the Immunological Studies on Vaccination


 , , , and
, , , and
Abstract
:1. Introduction
- (a)
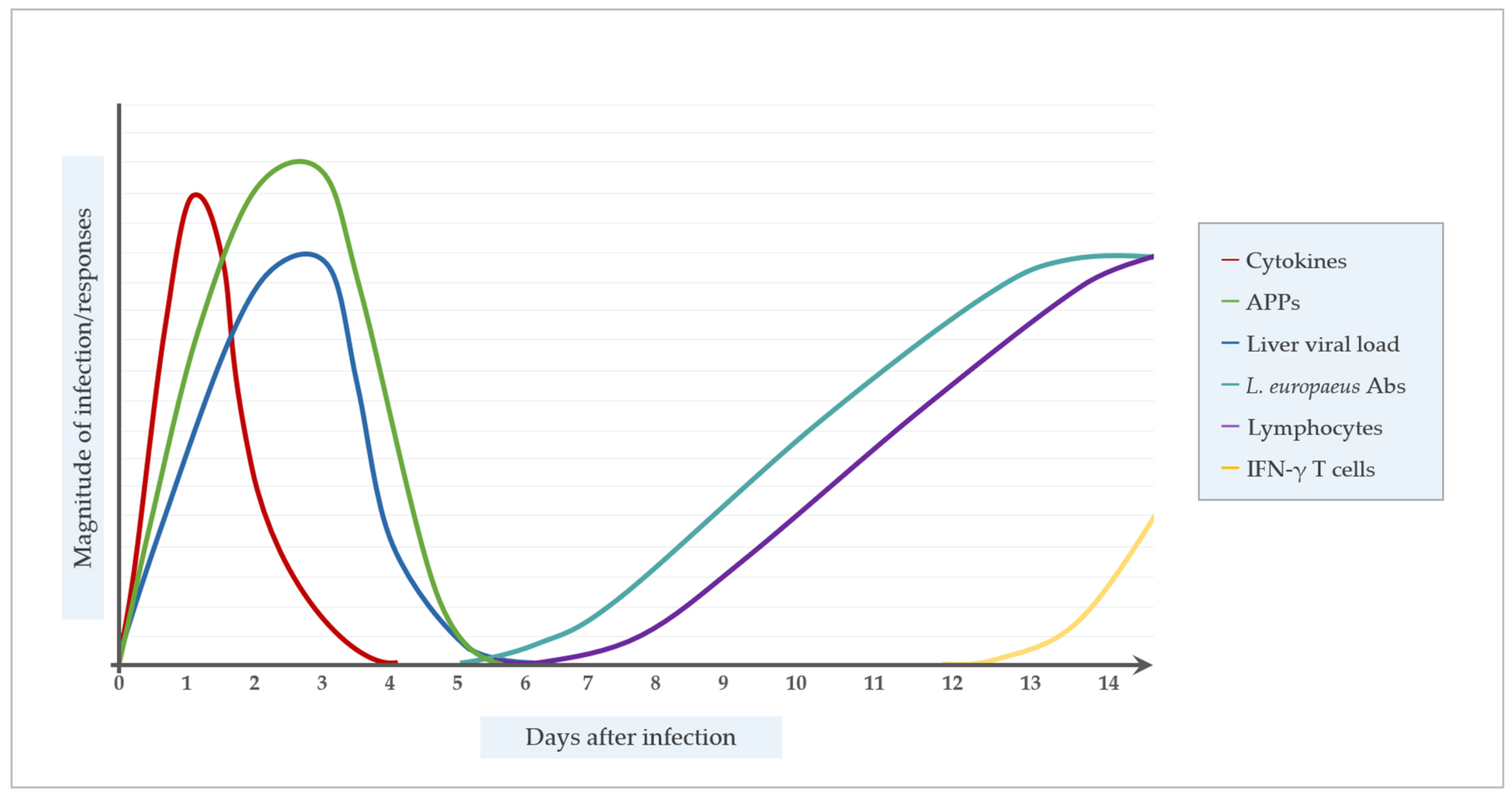
- Disturbed physiology with the severe destruction of the target organs (liver, spleen, kidney) along with an extremely rapid RHDV replication within 36–72 h post infection, with up to 214 HU related to 108 copies of viral RNA per g of liver tissue [42]. Immunohistological examinations and in-situ hybridization (ISH) studies of infected adult rabbits have demonstrated the presence of RHDV in periportal hepatocytes, and macrophages of the liver, lungs, and spleen [43,44]. The disease progression is correlated to an increased apoptosis of hepatocytes and liver endothelial cells [43]. The disturbed coagulation by increased prothrombin time, increased fibrin degradation, and decreased antithrombin III activity results in micro-thrombi, DIC, and a hypervariable condition in blood coagulation [45]. Liver enzymes significantly increase, whereas antioxidant enzymes decrease within the first 36 h post infection [46]. Respiratory acidosis, hypoglycemia, and increased creatinine kinase activity indicate multi organ dysfunction, which finally results in death.
- (b)
- Disturbed immune response with an induction of systemic apoptosis of lymphocytes, and especially T-cells, leading to a nearly complete loss of the immune effectors within 48–72 h after infection. In adult rabbits, the apoptosis of T- and B-cells in the liver and peripheral blood [47,48] with the infiltration of neutrophils [49] ends in a decrease of regulatory T-cells [50] and severe leukopenia before death [51]. After RHDV infection, an increase of pro-inflammatory cytokines in the serum, liver, and spleen of young and adult rabbits [50,52], and also anti-inflammatory cytokines in adult rabbits has been reported [53].
2. Lagovirus Europaeus—Epidemiological History of the RHD Causative Agent
3. Host-Pathogen Interaction in Rabbits after Infection with RHDV (Lagovirus europaeus/GI)
- (a)
- Missing virus receptor in peripheral mucosal surfaces.Juvenile rabbits do not express, or only express in incomplete formation, one identified attachment factor for RHDV, the so called HBGA (histo-blood group antigens) in the upper respiratory and digestive tract until the age of 9 weeks [122]. However, entry into hepatocytes is not provided by HBGA because they are not expressed at any age on hepatocytes [122]. Moreover, even in juvenile rabbits, RHDV was detected after infection in the liver [123,124]. Finally, immunosuppressed juvenile rabbits develop a full disease state as adults, indicating that immune mechanisms are involved in the juvenile resistance against RHDV [125].
- (b)
- Different replication kinetics in peripheral mucosal surfaces.Flies or direct contact transfer RHDV via the oral/oculo-nasal aerogenic route through the upper respiratory tract with an extremely low dose of possibly less than 100 viral particles [112]. However, between 24 and 48 h post-infection, RHDV replicates to very high titers in the liver of adult rabbits, whereas the amount of RHDV in the liver of juveniles remains low [124]. Whether this is due to limited virus replication in peripheral mucosa in juveniles or due to more effective type I or III IFN responses in juveniles compared to adult rabbits is not clear.
- (c)
- Different effective innate immune responses.The pathological processes (see above) leading finally to a DIC are described for both juvenile and adult rabbits [124,126]. Protective immune mechanisms preventing severe clinical processes are mostly developed in juvenile rabbits, but rarely in adults, The more robust innate immune response in juvenile rabbits was characterized as one reason for the RHDV resistance in a transcriptome analysis-based study [127]. The involvement of cellular immune mechanisms, and especially of CD8+ T-cells in adult rabbits in protective responses were elucidated [111,128].
- (d)
- Different regulation of the Th1/Th2 balance in juvenile and adult rabbits.In surviving adult rabbits, an early and strong increase of IFNγ as well as a limited loss of T-cells was determined. The course of the disease is less severe with a reduction of the viral replication in the liver, an increase of CD8+ T-cells, and an increase of IFNγ as well as type I IFNs [53]. Similar response patterns are described for juvenile rabbits where macrophages, T- and B-cells increased in the liver and spleen [52,123]. The reasons for the ‘decision’ towards Th1-based immune responses are different in adults and in juveniles. In juveniles, it might be based on their immature B-cell development [117], as the different outcome after RHDV infection is related to B1 cells contributing to innate immune responses, and not B2 cells as part of the adaptive immune response [129].
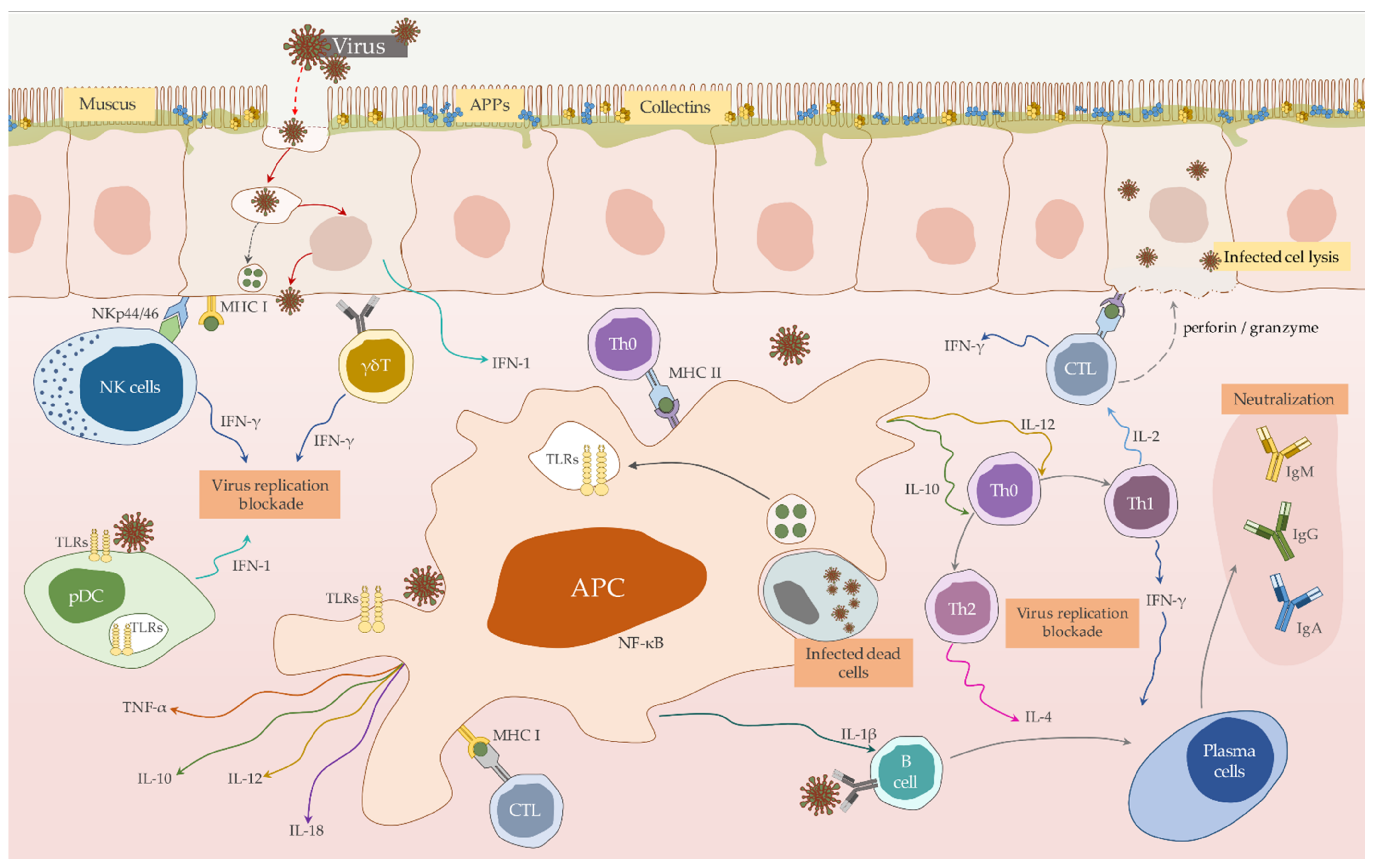
3.1. Innate Immune Mechanisms Involved in the Anti-RHDV Immune Response of Juvenile and Adult Rabbits
3.1.1. Pathogen Sensing
3.1.2. Apoptosis and Autophagy in RHDV Infection
3.1.3. Apoptosis and Autophagy in RHDV Infection
3.1.4. Innate Effector Component and Functions
3.2. Antigen Presentation
3.2.1. DC’s Interaction with RHDV
3.2.2. Antigen Processing
3.3. Adaptive Immunity
3.3.1. Humoral Immunity to RHDV
3.3.2. Cellular Response
3.4. Immune Response Pattern Protecting Naïve Rabbits—Lessons to Learn for Vaccination
4. Vaccines—From Inactivated RHDV-Infected Liver Preparation to Recombinant Vaccines
4.1. Conventional Vaccines against RHDV
4.2. Recombinant Vaccines against RHDV
4.3. New Approaches in Vaccine Design
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Di Giallonardo, F.; Holmes, EC. Viral biocontrol: Grand experiments in disease emergence and evolution. Trends Microbiol. 2015, 23, 83–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elsworth, P.G.; Kovaliski, J.; Cooke, B. Rabbit Haemorrhagic Disease: Are Australian rabbits (Oryctolagus cuniculus) evolving resistance to infection with Czech CAPM 351 RHDV? Epidemiol. Infect. 2012, 140, 1972–1981. [Google Scholar] [CrossRef]
- Elsworth, P.; Cooke, B.D.; Kovaliski, J.; Sinclair, R.; Holmes, E.C.; Strive, T. Increased virulence of rabbit haemorrhagic disease virus associated with genetic resistance in wild Australian rabbits (Oryctolagus cuniculus). Virology 2014, 464–465, 415–423. [Google Scholar] [CrossRef] [Green Version]
- Strive, T.; Elsworth, P.; Liu, J.; Wright, J.D.; Kovaliski, J.; Capucci, L. The non-pathogenic Australian rabbit calicivirus RCV-A1 provides temporal and partial cross protection to lethal Rabbit Haemorrhagic Disease Virus infection which is not dependent on antibody titres. Vet. Res. 2013, 44, 51. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.J.; Xue, H.P.; Pu, B.Q.; Qian, N.H. A New Viral Disease in Rabbits. Anim. Hus. Vet. Med. 1984, 16, 253–255. [Google Scholar]
- Abrantes, J.; Van der Loo, W.; Le Pendu, J.; Esteves, P.J. Rabbit haemorrhagic disease (RHD) and rabbit haemorrhagic disease virus (RHDV): A review. Vet. Res. 2012, 43, 12. [Google Scholar] [CrossRef] [Green Version]
- Górski, J.; Mizak, B.; Mizak, Z.; Komorowski, A. Obraz kliniczny oraz zmiany anatomopatologiczne w przebiegu pomoru królików (wirusowej krwiotocznej bronchopneumonii królików). Życie Wet. 1988, 63, 266–269. [Google Scholar]
- Löliger, H.C.; Eskens, U. Incidence, epizootiology and control of viral haemorrhagic disease of rabbits and the European brown hare syndrome in Germany. Rev. Sci. Tech. 1991, 10, 423–434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morisse, J.P.; le Gall, G.; Boilletot, E. Hepatitis of Viral Origin in Leporidae: Introduction and Aetiological Hypotheses. Rev. Sci. Tech. 1991, 10, 269–310. [Google Scholar] [CrossRef] [Green Version]
- Le Gall, G.; Arnauld, C.; Boilletot, E.; Morisse, J.P.; Rasschaert, D. Molecular Epidemiology of Rabbit Haemorrhagic Disease Virus Outbreaks in France during 1988 to 1995. J. Gen. Virol. 1998, 79, 11–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooke, B.D. Rabbit Haemorrhagic Disease: Field Epidemiology and the Management of Wild Rabbit Populations. OIE Rev. Sci. Tech. 2002, 21, 347–358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alda, F.; Gaitero, T.; Suárez, M.; Merchán, T.; Rocha, G.; Doadrio, I. Evolutionary History and Molecular Epidemiology of Rabbit Haemorrhagic Disease Virus in the Iberian Peninsula and Western Europe. BMC Evol. Biol. 2010, 10, 347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinnear, M.; Linde, C.C. Capsid Gene Divergence in Rabbit Hemorrhagic Disease Virus. J. Gen. Virol. 2010, 91, 174–181. [Google Scholar] [CrossRef]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Melo, P.; Correia, J.J.; Ramada, M.; Alves, P.C.; Parra, F.; Esteves, P.J. New Variant of Rabbit Hemorrhagic Disease Virus, Portugal, 2012–2013. Emerg. Infect. Dis. 2013, 19, 1900–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abrantes, J.; Lopes, A.M.; Dalton, K.P.; Parra, F.; Esteves, P.J. Detection of RHDVa on the Iberian Peninsula: Isolation of an RHDVa Strain from a Spanish Rabbitry. Arch. Virol. 2014, 159, 321–326. [Google Scholar] [CrossRef]
- Westcott, D.G.; Frossard, J.P.; Everest, D.; Dastjerdi, A.; Duff, J.P.; Steinbach, F.; Choudhury, B. Incursion of RHDV2-like Variant in Great Britain. Vet. Rec. 2014, 174, 333. [Google Scholar] [CrossRef]
- Bárcena, J.; Guerra, B.; Angulo, I.; González, J.; Valcárcel, F.; Mata, C.P.; Castón, J.R.; Blanco, E.; Alejo, A. Comparative Analysis of Rabbit Hemorrhagic Disease Virus (RHDV) and New RHDV2 Virus Antigenicity, Using Specific Virus-like Particles. Vet. Res. 2015, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monterroso, P.; Garrote, G.; Serronha, A.; Santos, E.; Delibes-Mateos, M.; Abrantes, J.; de Ayala, R.P.; Silvestre, F.; Carvalho, J.; Vasco, I.; et al. Disease-Mediated Bottom-up Regulation: An Emergent Virus Affects a Keystone Prey, and Alters the Dynamics of Trophic Webs. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Rouco, C.; Abrantes, J.; Serronha, A.; Lopes, A.M.; Maio, E.; Magalhães, M.J.; Blanco, E.; Bárcena, J.; Esteves, P.J.; Santos, N.; et al. Epidemiology of RHDV2 (Lagovirus Europaeus/GI.2) in Free-Living Wild European Rabbits in Portugal. Transbound. Emerg. Dis. 2018, 65, e373–e382. [Google Scholar] [CrossRef] [PubMed]
- Gibb, J.A.; Williams, J.M. The rabbit in New Zealand. In The European Rabbit: History and Biology of a Successful Colonizer; Thompson, H.V., King, C.M., Eds.; Oxford University Press: Oxford, UK, 1994; pp. 158–200. [Google Scholar]
- Mutze, G.; Cooke, B.; Alexander, P. The Initial Impact of Rabbit Hemorrhagic Disease on European Rabbit Populations in South Australia. J. Wildl. Dis. 1998, 34, 221–227. [Google Scholar] [CrossRef] [Green Version]
- Fenner, F. Deliberate Introduction of the European Rabbit, Oryctolagus Cuniculus, into Australia. OIE Rev. Sci. Tech. 2010, 29, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Granzow, H.; Schirrmeier, H.; Tews, G. Hämorrhagische Septikämie Der Kaninchen—Erregernachweis Und Erste Elektronenmikroskopische Charakterisierung. Mh. Vet. Med. 1989, 44, 379–380. [Google Scholar]
- Ohlinger, V.F.; Haas, B.; Ahl, R.; Weiland, F. Rabbit haemorrhagic disease—A contagious disease caused by Calicivirus. Tierärztliche Umschau. 1989, 44, 284–294. [Google Scholar]
- Ohlinger, V.F.; Haas, B.; Meyers, G.; Weiland, F.; Thiel, H.J. Identification and Characterization of the Virus Causing Rabbit Hemorrhagic Disease. J. Virol. 1990, 64, 3331–3336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parra, F.; Prieto, M. Purification and Characterization of a Calicivirus as the Causative Agent of a Lethal Hemorrhagic Disease in Rabbits. J. Virol. 1990, 64, 4013–4015. [Google Scholar] [CrossRef] [Green Version]
- Meyers, G.; Wirblich, C.; Thiel, H.J. Rabbit Hemorrhagic Disease Virus-Molecular Cloning and Nucleotide Sequencing of a Calicivirus Genome. Virology 1991, 184, 664–676. [Google Scholar] [CrossRef]
- Moussa, A.; Chasey, D.; Lavazza, A.; Capucci, L.; Šmid, B.; Meyers, G.; Rossi, C.; Thiel, H.J.; Vlásak, R.; Rønsholt, L.; et al. Haemorrhagic Disease of Lagomorphs: Evidence for a Calicivirus. Vet. Microbiol. 1992, 33, 375–381. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lavazza, A.; Marchandeau, S.; Bertagnoli, S.; Zwingelstein, F.; Cavadini, P.; Martinelli, N.; Lombardi, G.; Guérin, J.L.; Lemaitre, E.; et al. Emergence of a New Lagovirus Related to Rabbit Haemorrhagic Disease Virus. Vet. Res. 2013, 44, 81. [Google Scholar] [CrossRef] [Green Version]
- Puggioni, G.; Cavadini, P.; Maestrale, C.; Scivoli, R.; Botti, G.; Ligios, C.; le Gall-Reculé, G.; Lavazza, A.; Capucci, L. The New French 2010 Rabbit Hemorrhagic Disease Virus Causes an RHD-like Disease in the Sardinian Cape Hare (Lepus Capensis Mediterraneus). Vet. Res. 2013, 44, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Camarda, A.; Pugliese, N.; Cavadini, P.; Circella, E.; Capucci, L.; Caroli, A.; Legretto, M.; Mallia, E.; Lavazza, A. Detection of the New Emerging Rabbit Haemorrhagic Disease Type 2 Virus (RHDV2) in Sicily from Rabbit (Oryctolagus Cuniculus) and Italian Hare (Lepus Corsicanus). Res. Vet. Sci. 2014, 97, 642–645. [Google Scholar] [CrossRef] [PubMed]
- Rouco, C.; Aguayo-Adán, J.A.; Santoro, S.; Abrantes, J.; Delibes-Mateos, M. Worldwide Rapid Spread of the Novel Rabbit Haemorrhagic Disease Virus (GI.2/RHDV2/b). Transbound. Emerg. Dis. 2019, 66, 1762–1764. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Lemaitre, E.; Bertagnoli, S.; Hubert, C.; Top, S.; Decors, A.; Marchandeau, S.; Guitton, J.S. Large-Scale Lagovirus Disease Outbreaks in European Brown Hares (Lepus Europaeus) in France Caused by RHDV2 Strains Spatially Shared with Rabbits (Oryctolagus Cuniculus). Vet. Res. 2017, 48, 70. [Google Scholar] [CrossRef] [Green Version]
- Neimanis, A.S.; Ahola, H.; Larsson Pettersson, U.; Lopes, A.M.; Abrantes, J.; Zohari, S.; Esteves, P.J.; Gavier-Widén, D. Overcoming Species Barriers: An Outbreak of Lagovirus Europaeus GI.2/RHDV2 in an Isolated Population of Mountain Hares (Lepus Timidus). BMC Vet. Res. 2018, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Lopes, A.M.; Marques, S.; Silva, E.; Magalhães, M.J.; Pinheiro, A.; Alves, P.C.; le Pendu, J.; Esteves, P.J.; Thompson, G.; Abrantes, J. Detection of RHDV Strains in the Iberian Hare (Lepus Granatensis): Earliest Evidence of Rabbit Lagovirus Cross-Species Infection. Vet. Res. 2014, 45, 94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- House Rabbit Society. Rabbit Hemorrhagic Disease Virus (RHDV). Available online: https://rabbit.org/rhdv/#southwest (accessed on 22 February 2021).
- USGS Science for a Changing World. An Update on NWHC Support to Wildlife Managers and Response to RHDV2. Available online: https://www.usgs.gov/media/files/update-nwhc-support-wildlife-managers-and-response-rhdv2 (accessed on 22 February 2021).
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. The Importance of Liver Lesions and Changes to Biochemical and Coagulation Factors in the Pathogenesis of RHD. Acta Biochim. Pol. 2015, 62, 169–171. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, N.; Fuchs, A.; Schilcher, F.; Loupal, G. Zum Auftreten der Rabbit Haemorrhagic Disease (RHD) in Österreich: I Pathomorphologische und virologische Untersuchungen. Wien. Tierärztl. Mschr. 1990, 77, 19–23. [Google Scholar]
- Chasey, D.; Lucas, M.; Westcott, D.; Williams, M. European Brown Hare Syndrome in the U.K.; a Calicivirus Related to but Distinct from That of Viral Haemorrhagic Disease in Rabbits. Arch. Virol. 1992, 124, 363–370. [Google Scholar] [CrossRef] [PubMed]
- Gregg, D.A.; House, C.; Meyer, R.; Berninger, M. Viral Haemorrhagic Disease of Rabbits in Mexico: Epidemiology and Viral Characterization. Rev. Sci. Tech. 1991, 10, 435–451. [Google Scholar] [CrossRef]
- Gall, A.; Hoffmann, B.; Teifke, J.P.; Lange, B.; Schirrmeier, H. Persistence of Viral RNA in Rabbits Which Overcome an Experimental RHDV Infection Detected by a Highly Sensitive Multiplex Real-Time RT-PCR. Vet. Microbiol. 2007, 120, 17–32. [Google Scholar] [CrossRef]
- Alonso, C.; Oviedo, J.M.; Martín-Alonso, J.M.; Díaz, E.; Boga, J.A.; Parra, F. Programmed Cell Death in the Pathogenesis of Rabbit Hemorrhagic Disease. Arch. Virol. 1998, 143, 321–332. [Google Scholar] [CrossRef] [PubMed]
- Teifke, J.P.; Reimann, I.; Schirrmeier, H. Subacute Liver Necrosis after Experimental Infection with Rabbit Haemorrhagic Disease Virus (RHDV). J. Comp. Pathol. 2002, 126, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Ueda, K.; Park, J.H.; Ochiai, K.; Itakura, C. Disseminated intravascular coagulation (DIC) in rabbit haemorrhagic disease. Jpn. J. Vet. Res. 1992, 40, 133–141. [Google Scholar]
- Sanchez-Campos, S.; Alvarez, M.; Culebras, J.M.; Gonzalez-Gallego, J.; Tunon, M.J. Pathogenic Molecular Mechanisms in an Animal Model of Fulminant Hepatic Failure: Rabbit Hemorrhagic Viral Disease. J. Lab. Clin. Med. 2004, 144, 215–222. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Deptuła, W. Apoptosis of Peripheral Blood Leukocytes from Rabbits Infected with Non-Haemagglutinating Strains of Rabbit Haemorrhagic Disease Virus (RHDV). Vet. Immunol. Immunopathol. 2012, 149, 54–57. [Google Scholar] [CrossRef] [PubMed]
- Niedźwiedzka-Rystwej, P.; Hukowska-Szematowicz, B.; Tokarz-Deptuła, B.; Trzeciak-Ryczek, A.; Działo, J.; Deptuła, W. Apoptosis of peripheral blood leucocytes in rabbits infected with different strains of rabbit haemorrhagic disease virus. Acta Biochim. Pol. 2013, 60, 65–69. [Google Scholar] [CrossRef] [Green Version]
- Marques, R.M.; Costa-E-Silva, A.; Águas, A.P.; Teixeira, L.; Ferreira, P.G. Early Acute Depletion of Lymphocytes in Calicivirus-Infected Adult Rabbits. Vet. Res. Commun. 2010, 34, 659–668. [Google Scholar] [CrossRef]
- Teixeira, L.; Marques, R.M.; Águas, A.P.; Ferreira, P.G. Regulatory T Cells Are Decreased in Acute RHDV Lethal Infection of Adult Rabbits. Vet. Immunol. Immunopathol. 2012, 148, 343–347. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.G.; Costa-e-Silva, A.; Oliveira, M.J.R.; Monteiro, E.; Cunha, E.M.; Águas, A.P. Severe Leukopenia and Liver Biochemistry Changes in Adult Rabbits after Calicivirus Infection. Res. Vet. Sci. 2006, 80, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Marques, R.M.; Costa-E-Silva, A.; Águas, A.P.; Teixeira, L.; Ferreira, P.G. Early Inflammatory Response of Young Rabbits Attending Natural Resistance to Calicivirus (RHDV) Infection. Vet. Immunol. Immunopathol. 2012, 150, 181–188. [Google Scholar] [CrossRef]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. Expression of IL-1Ra, IL-6, IL-8, IL-18, TNF-α and IFN-γ Genes in Peripheral Blood Leukocytes of Rabbits Infected with RHDV (Rabbit Haemorrhagic Disease Virus). Dev. Comp. Immunol. 2017, 76, 310–315. [Google Scholar] [CrossRef] [PubMed]
- Argüello Villares, J.L. Viral Haemorrhagic Disease of Rabbits: Vaccination and Immune Response. Rev. Sci. Tech. 1991, 10, 459–480. [Google Scholar] [CrossRef]
- Huang, H.B. Vaccination against and Immune Response to Viral Haemorrhagic Disease of Rabbits: A Review of Research in the People’s Republic of China. Rev. Sci. Tech. Off. Int. Epiz. 1991, 10, 481–498. [Google Scholar]
- Šmíd, B.; Valíček, L.; Rodák, L.; Štěpánek, J.; Jurák, E. Rabbit Haemorrhagic Disease: An Investigation of Some Properties of the Virus and Evaluation of an Inactivated Vaccine. Vet. Microbiol. 1991, 26, 77–85. [Google Scholar] [CrossRef]
- Vautherot, J.F.; Laurent, S.; le Gall, G.; Morisse, J.P.; Rasschaert, D. Recombinant vaccine against rabbit hemorrhagic disease virus. Vet. Res. 1995, 26, 206–207. [Google Scholar]
- Soledad Marín, M.; Martín Alonso, J.M.; Pérez Ordoyo García, L.I.; Antonio Boga, J.; Argüello-Villares, J.L.; Casais, R.; Venugopal, K.; Jiang, W.; Gould, E.A.; Parra, F. Immunogenic Properties of Rabbit Haemorrhagic Disease Virus Structural Protein VP60 Expressed by a Recombinant Baculovirus: An Efficient Vaccine. Virus Res. 1995, 39, 119–128. [Google Scholar] [CrossRef]
- Plana-Duran, J.; Bastons, M.; Rodriguez, M.J.; Climent, I.; Cortés, E.; Vela, C.; Casal, I. Oral Immunization of Rabbits with VP60 Particles Confers Protection against Rabbit Hemorrhagic Disease. Arch. Virol. 1996, 141, 1423–1436. [Google Scholar] [CrossRef] [PubMed]
- Bertagnoli, S.; Gelfi, J.; le Gall, G.; Boilletot, E.; Vautherot, J.F.; Rasschaert, D.; Laurent, S.; Petit, F.; Boucraut-Baralon, C.; Milon, A. Protection against Myxomatosis and Rabbit Viral Hemorrhagic Disease with Recombinant Myxoma Viruses Expressing Rabbit Hemorrhagic Disease Virus Capsid Protein. J. Virol. 1996, 70, 5061–5066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertagnoli, S.; Gelfi, J.; Petit, F.; Vautherot, J.F.; Rasschaert, D.; Laurent, S.; le Gall, G.; Boilletot, E.; Chantal, J.; Boucraut-Baralon, C. Protection of Rabbits against Rabbit Viral Haemorrhagic Disease with a Vaccinia-RHDV Recombinant Virus. Vaccine 1996, 14, 506–510. [Google Scholar] [CrossRef]
- Fischer, L.; le Gros, F.X.; Mason, P.W.; Paoletti, E. A Recombinant Canarypox Virus Protects Rabbits against a Lethal Rabbit Hemorrhagic Disease Virus (RHDV) Challenge. Vaccine 1997, 15, 90–96. [Google Scholar] [CrossRef]
- Castañón, S.; Marín, M.S.; Martín-Alonso, J.M.; Boga, J.A.; Casais, R.; Humara, J.M.; Ordás, R.J.; Parra, F. Immunization with Potato Plants Expressing VP60 Protein Protects against Rabbit Hemorrhagic Disease Virus. J. Virol. 1999, 73, 4452–4455. [Google Scholar] [CrossRef] [Green Version]
- Abrantes, J.; Droillard, C.; Lopes, A.M.; Lemaitre, E.; Lucas, P.; Blanchard, Y.; Marchandeau, S.; Esteves, P.J.; Le Gall-Reculé, G. Recombination at the Emergence of the Pathogenic Rabbit Haemorrhagic Disease Virus Lagovirus Europaeus/GI.2. Sci. Rep. 2020, 10, 14502. [Google Scholar] [CrossRef]
- Abrantes, J.; Esteves, P.J.; van der Loo, W. Evidence for Recombination in the Major Capsid Gene VP60 of the Rabbit Haemorrhagic Disease Virus (RHDV). Arch. Virol. 2008, 153, 329–335. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Trout, R.C.; Gould, E.A. Benign Circulation of Rabbit Haemorrhagic Disease Virus on Lambay Island, Eire. Virology 2007, 358, 18–22. [Google Scholar] [CrossRef]
- Hu, B.; Wang, F.; Fan, Z.; Song, Y.; Abrantes, J.; Zuo, Y.; Esteves, P.J. Recombination between G2 and G6 Strains of Rabbit Hemorrhagic Disease Virus (RHDV) in China. Arch. Virol. 2017, 162, 269–272. [Google Scholar] [CrossRef]
- Lopes, A.M.; Dalton, K.P.; Magalhães, M.J.; Parra, F.; Esteves, P.J.; Holmes, E.C.; Abrantes, J. Full Genomic Analysis of New Variant Rabbit Hemorrhagic Disease Virus Revealed Multiple Recombination Events. J. Gen. Virol. 2015, 96, 1309–1319. [Google Scholar] [CrossRef]
- White, P.J.; Trout, R.C.; Moss, S.R.; Desai, A.; Armesto, M.; Forrester, N.L.; Gould, E.A.; Hudson, P.J. Epidemiology of Rabbit Haemorrhagic Disease Virus in the United Kingdom: Evidence for Seasonal Transmission by Both Virulent and Avirulent Modes of Infection. Epidemiol. Infect. 2004, 132, 555–567. [Google Scholar] [CrossRef]
- Capucci, L.; Fusi, P.; Lavazza, A.; Pacciarini, M.L.; Rossi, C. Detection and Preliminary Characterization of a New Rabbit Calicivirus Related to Rabbit Hemorrhagic Disease Virus but Nonpathogenic. J. Virol. 1996, 70, 8614–8623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moss, S.R.; Turner, S.L.; Trout, R.C.; White, P.J.; Hudson, P.J.; Desai, A.; Armesto, M.; Forrester, N.L.; Gould, E.A. Molecular Epidemiology of Rabbit Haemorrhagic Disease Virus. J. Gen. Virol. 2002, 83, 2461–2467. [Google Scholar] [CrossRef]
- Forrester, N.L.; Abubakr, M.I.; Abu Elzein, E.M.E.; Al-Afaleq, A.I.; Housawi, F.M.T.; Moss, S.R.; Turner, S.L.; Gould, E.A. Phylogenetic Analysis of Rabbit Haemorrhagic Disease Virus Strains from the Arabian Peninsula: Did RHDV Emerge Simultaneously in Europe and Asia? Virology 2006, 344, 277–282. [Google Scholar] [CrossRef]
- Strive, T.; Wright, J.; Kovaliski, J.; Botti, G.; Capucci, L. The Non-Pathogenic Australian Lagovirus RCV-A1 Causes a Prolonged Infection and Elicits Partial Cross-Protection to Rabbit Haemorrhagic Disease Virus. Virology 2010, 398, 125–134. [Google Scholar] [CrossRef] [Green Version]
- Kerr, P.J.; Kitchen, A.; Holmes, E.C. Origin and Phylodynamics of Rabbit Hemorrhagic Disease Virus. J. Virol. 2009, 83, 12129–12138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esteves, P.J.; Abrantes, J.; Bertagnoli, S.; Cavadini, P.; Gavier-Widén, D.; Guitton, J.-S.; Lavazza, A.; Lemaitre, E.; Letty, J.; Lopes, A.M.; et al. Emergence of Pathogenicity in Lagoviruses: Evolution from Pre-Existing Nonpathogenic Strains or through a Species Jump? PLoS Pathog. 2015, 11, e1005087. [Google Scholar] [CrossRef]
- Merchán, T.; Rocha, G.; Alda, F.; Silva, E.; Thompson, G.; de Trucios, S.H.; Pagés, A. Detection of Rabbit Haemorrhagic Disease Virus (RHDV) in Nonspecific Vertebrate Hosts Sympatric to the European Wild Rabbit (Oryctolagus Cuniculus). Infect. Genet. Evol. 2011, 11, 1469–1474. [Google Scholar] [CrossRef] [Green Version]
- Gavier-Widén, D.; Mörner, T. Epidemiology and Diagnosis of the European Brown Hare Syndrome in Scandinavian Countries: A Review. Rev. Sci. Tech. 1991, 10, 453–458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wirblich, C.; Meyers, G.; Ohlinger, V.F.; Capucci, L.; Eskens, U.; Haas, B.; Thiel, H.J. European Brown Hare Syndrome Virus: Relationship to Rabbit Hemorrhagic Disease Virus and Other Caliciviruses. J. Virol. 1994, 68, 5164–5173. [Google Scholar] [CrossRef] [Green Version]
- Nowotny, N.; Ros Bascuñana, C.; Ballagi-Pordány, A.; Gavier-Widén, D.; Uhlén, M.; Belák, S. Phylogenetic Analysis of Rabbit Haemorrhagic Disease and European Brown Hare Syndrome Viruses by Comparison of Sequences from the Capsid Protein Gene. Arch. Virol. 1997, 142, 657–673. [Google Scholar] [CrossRef]
- Lavazza, A.; Scicluna, M.T.; Capucci, L. Susceptibility of Hares and Rabbits to the European Brown Hare Syndrome Virus (EBHSV) and Rabbit Haemorrhagic Disease Virus (RHDV) under Experimental Conditions. Zentralbl. Veterinarmed. B 1996, 43, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Cancellotti, F.M.; Renzi, M. Epidemiology and Current Situation of Viral Haemorrhagic Disease of Rabbits and the European Brown Hare Syndrome in Italy. Rev. Sci. Tech. 1991, 10, 409–422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, N.Y.; Chong, C.Y.; Kim, J.H.; Cho, S.M.; Cha, Y.H.; Jung, B.T.; Kim, D.S.; Yoon, J.B. An Outbreak of Viral Haemorrhagic Pneumonia (Tentative Name) of Rabbits in Korea. J. Korean Vet. Med. Assoc. 1987, 23, 603–610. [Google Scholar]
- Xu, W.Y. Viral Haemorrhagic Disease of Rabbits in the People’s Republic of China: Epidemiology and Virus Characterisation. Rev. Sci. Tech. 1991, 10, 393–408. [Google Scholar] [PubMed]
- Argüello Villares, J.L.; Llanos Pellitero, A.; Pérez-Ordoyo Garcia, L.I. Enfermedad vírica hemorragica del canejo. Rev. Med. Vet. 1988, 5, 645–650. [Google Scholar]
- Villafuerte, R.; Calvete, C.; Blanco, J.C.; Lucientes, J. Incidence of Viral Hemorrhagic Disease in Wild Rabbit Populations in Spain. Mammalia 1995, 59, 651–660. [Google Scholar] [CrossRef]
- Delibes-Mateos, M.; Redpath, S.M.; Angulo, E.; Ferreras, P.; Villafuerte, R. Rabbits as a Keystone Species in Southern Europe. Biol. Conserv. 2007, 137, 149–156. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Laurent, S.; de Boisséson, C.; Portejoie, Y.; Rasschaert, D. Phylogenetic Analysis of Rabbit Haemorrhagic Disease Virus in France between 1993 and 2000, and the Characterisation of RHDV Antigenic Variants. Arch. Virol. 2003, 148, 65–81. [Google Scholar] [CrossRef] [PubMed]
- Farnós, O.; Rodríguez, D.; Valdés, O.; Chiong, M.; Parra, F.; Toledo, J.R.; Fernández, E.; Lleonart, R.; Suárez, M. Molecular and Antigenic Characterization of Rabbit Hemorrhagic Disease Virus Isolated in Cuba Indicates a Distinct Antigenic Subtype. Arch. Virol. 2007, 152, 1215–1221. [Google Scholar] [CrossRef]
- Cooke, B.D.; Fenner, F. Rabbit Haemorrhagic Disease and the Biological Control of Wild Rabbits, Oryctolagus Cuniculus, in Australia and New Zealand. Wildl. Res. 2002, 29, 689–706. [Google Scholar] [CrossRef]
- Thompson, J.; Clark, G. Rabbit Calicivirus Disease Now Established in New Zealand. Surveillance 1997, 24, 5–6. [Google Scholar]
- O’Keefe, J.S.; Tempero, J.; Atkinson, P.H.; Pacciarini, L.; Fallacara, F.; Horner, G.W.; Motha, J. Typing of Rabbit Haemorrhagic Disease Virus from New Zealand Wild Rabbits. N. Z. Vet. J. 1998, 46, 42–43. [Google Scholar] [CrossRef]
- Vinjé, J.; Estes, M.K.; Esteves, P.; Green, K.Y.; Katayama, K.; Knowles, N.J.; L’Homme, Y.; Martella, V.; Vennema, H.; White, P.A.; et al. ICTV Virus Taxonomy Profile: Caliciviridae. J. Gen. Virol. 2019, 100, 1469–1470. [Google Scholar] [CrossRef] [PubMed]
- Le Pendu, J.; Abrantes, J.; Bertagnoli, S.; Guitton, J.S.; le Gall-Reculé, G.; Lopes, A.M.; Marchandeau, S.; Alda, F.; Almeida, T.; Célio, A.P.; et al. Proposal for a Unified Classification System and Nomenclature of Lagoviruses. J. Gen. Virol. 2017, 98, 1658–1666. [Google Scholar] [CrossRef]
- Velarde, R.; Cavadini, P.; Neimanis, A.; Cabezón, O.; Chiari, M.; Gaffuri, A.; Lavín, S.; Grilli, G.; Gavier-Widén, D.; Lavazza, A.; et al. Spillover Events of Infection of Brown Hares (Lepus Europaeus) with Rabbit Haemorrhagic Disease Type 2 Virus (RHDV2) Caused Sporadic Cases of an European Brown Hare Syndrome-Like Disease in Italy and Spain. Transbound. Emerg. Dis. 2017, 64, 1750–1761. [Google Scholar] [CrossRef]
- Hall, R.N.; Peacock, D.E.; Kovaliski, J.; Mahar, J.E.; Mourant, R.; Piper, M.; Strive, T. Detection of RHDV2 in European Brown Hares (Lepus Europaeus) in Australia. Vet. Rec. 2017, 180, 121. [Google Scholar] [CrossRef]
- Capucci, L.; Scicluna, M.T.; Lavazza, A. Diagnosis of Viral Haemorrhagic Disease of Rabbits and the European Brown Hare Syndrome. Rev. Sci. Tech. 1991, 10, 347–370. [Google Scholar] [CrossRef]
- Marcato, P.S.; Benazzi, C.; Vecchi, G.; Galeotti, M.; della Salda, L.; Sarli, G.; Lucidi, P. Clinical and Pathological Features of Viral Haemorrhagic Disease of Rabbits and the European Brown Hare Syndrome. Rev. Sci. Tech. 1991, 10, 371–392. [Google Scholar] [CrossRef]
- Fuchs, A.; Weissenböck, H. Comparative histopathological study of rabbit haemorrhagic disease (RHD) and European brown hare syndrome (EBHS). Comp. Study J. Comp. Pathol. 1992, 107, 103–113. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Deptuła, W. Cytometric analysis of lymphocytes T and B in rabbits infected with non-haemagglutinogenic strains of RHD virus (rabbit haemorrhagic disease)—Rainham, Frankfurt and Asturias. Pol. J. Vet. Sci. 2010, 13, 157–162. [Google Scholar]
- Bębnowska, D.; Niedźwiedzka-Rystwej, P. Characteristics of a New Variant of Rabbit Haemorrhagic Disease Virus—RHDV2. Acta Biolog. 2019, 26, 83–97. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Tokarz-Deptuła, B.; Hukowska-Szematowicz, B.; Działo, J.; Deptuła, W. Indices of Non-Specific Immunity: An Element of Natural Immunity in Rabbits Infected with RHD (Rabbit Haemorrhagic Disease) Virus. Cent. Eur. J. Immunol. 2013, 38, 231–236. [Google Scholar] [CrossRef] [Green Version]
- Niedźwiedzka-Rystwej, P.; Tokarz-Deptuła, B.; Deptuła, W. Lymphocytes T and B in rabbits infected with RHD virus. Pol. J. Vet. Sci. 2013, 16, 563–565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopes, A.M.; Capucci, L.; Gavier-Widén, D.; le Gall-Reculé, G.; Brocchi, E.; Barbieri, I.; Quéméner, A.; le Pendu, J.; Geoghegan, J.L.; Holmes, E.C.; et al. Molecular Evolution and Antigenic Variation of European Brown Hare Syndrome Virus (EBHSV). Virology 2014, 468–470, 104–112. [Google Scholar] [CrossRef] [Green Version]
- International Committee on Taxonomy of Viruses (ICTV). Genus: Lagovirus. Available online: https://talk.ictvonline.org/ictv-reports/ictv_online_report/positive-sense-rna-viruses/w/caliciviridae/1163/genus-lagovirus (accessed on 10 January 2021).
- Capucci, L.; Frigoli, G.; Rønshold, L.; Lavazza, A.; Brocchi, E.; Rossi, C. Antigenicity of the Rabbit Hemorrhagic Disease Virus Studied by Its Reactivity with Monoclonal Antibodies. Virus Res. 1995, 37, 221–238. [Google Scholar] [CrossRef]
- Capucci, L.; Fallacara, F.; Grazioli, S.; Lavazza, A.; Pacciarini, M.L.; Brocchi, E. A Further Step in the Evolution of Rabbit Hemorrhagic Disease Virus: The Appearance of the First Consistent Antigenic Variant. Virus Res. 1998, 58, 115–126. [Google Scholar] [CrossRef]
- Schirrmeier, H.; Reimann, I.; Köllner, B.; Granzow, H. Pathogenic, Antigenic and Molecular Properties of Rabbit Haemorrhagic Disease Virus (RHDV) Isolated from Vaccinated Rabbits: Detection and Characterization of Antigenic Variants. Arch. Virol. 1999, 144, 719–735. [Google Scholar] [CrossRef] [PubMed]
- Dalton, K.P.; Nicieza, I.; Abrantes, J.; Esteves, P.J.; Parra, F. Spread of New Variant RHDV in Domestic Rabbits on the Iberian Peninsula. Vet. Microbiol. 2014, 169, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Calvete, C.; Mendoza, M.; Alcaraz, A.; Sarto, M.P.; Jiménez-de-Bagüéss, M.P.; Calvo, A.J.; Monroy, F.; Calvo, J.H. Rabbit Haemorrhagic Disease: Cross-Protection and Comparative Pathogenicity of GI.2/RHDV2/b and GI.1b/RHDV Lagoviruses in a Challenge Trial. Vet. Microbiol. 2018, 219, 87–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahali, N.; Sghaier, S.; Kbaier, H.; Zanati, A.; Bahloul, C. Genetic Characterization and Phylogenetic Analysis of Rabbit Hemorrhagic Disease Virus Isolated in Tunisia from 2015 to 2018. Arch. Virol. 2019, 164, 2327–2332. [Google Scholar] [CrossRef]
- Müller, C.; Ulrich, R.; Schinköthe, J.; Müller, M.; Köllner, B. Characterization of Protective Humoral and Cellular Immune Responses against RHDV2 Induced by a New Vaccine Based on Recombinant Baculovirus. Vaccine 2019, 37, 4195–4203. [Google Scholar] [CrossRef] [PubMed]
- Gehrmann, B.; Kretzschmar, C. Ein experimenteller Beitrag zur Epizootiologie der Viralen Hämorrhagischen Septikämie der Kaninchen (Rabbit Haemorrhagic Disease, RHD) Übertragung durch Fliegen. Berl. Munch. Tierarztl. Wschr. 1991, 104, 192–194. [Google Scholar]
- Xu, Z.J.; Chen, W.X. Viral Haemorrhagic Disease in Rabbits: A Review. Vet. Res. Commun. 1989, 13, 205–212. [Google Scholar] [CrossRef]
- Ohlinger, V.F.; Haas, B.; Thiel, H.J. Rabbit hemorrhagic disease (RHD): Characterization of the causative calicivirus. Vet. Res. 1993, 24, 103–116. [Google Scholar] [PubMed]
- Asgari, S.; Hardy, J.R.E.; Sinclair, R.G.; Cooke, B.D. Field Evidence for Mechanical Transmission of Rabbit Haemorrhagic Disease Virus (RHDV) by Flies (Diptera: Calliphoridae) among Wild Rabbits in Australia. Virus Res. 1998, 54, 123–132. [Google Scholar] [CrossRef]
- Dalton, K.P.; Nicieza, I.; Balseiro, A.; Muguerza, M.A.; Rosell, J.M.; Casais, R.; Álvarez, Á.L.; Parra, F. Variant Rabbit Hemorrhagic Disease Virus in Young Rabbits, Spain. Emerg. Infect. Dis. 2012, 18, 2009–2012. [Google Scholar] [CrossRef]
- Knight, K.L.; Winstead, C.R. Generation of Antibody Diversity in Rabbits. Curr. Opin. Immunol. 1997, 9, 228–232. [Google Scholar] [CrossRef]
- Crisci, E.; Almanza, H.; Mena, I.; Córdoba, L.; Gómez-Casado, E.; Castón, J.R.; Fraile, L.; Bárcena, J.; Montoya, M. Chimeric Calicivirus-like Particles Elicit Protective Anti-Viral Cytotoxic Responses without Adjuvant. Virology 2009, 387, 303–312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.N. Studies on interferon anti RHDV activities in rabbits. J. Anim. Quar. 1990, 3, 19–22. [Google Scholar]
- Cooke, B.D.; Robinson, A.J.; Merchant, J.C.; Nardin, A.; Capucci, L. Use of ELISAs in Field Studies of Rabbit Haemorrhagic Disease (RHD) in Australia. Epidemiol. Infect. 2000, 124, 563–576. [Google Scholar] [CrossRef]
- Ferreira, P.G.; Dinís, M.; Costa-e-Silva, A.; Águas, A.P. Adult Rabbits Acquire Resistance to Lethal Calicivirus Infection by Adoptive Transfer of Sera from Infected Young Rabbits. Vet. Immunol. Immunopathol. 2008, 121, 364–369. [Google Scholar] [CrossRef]
- Ruvoën-Clouet, N.; Ganière, J.P.; André-Fontaine, G.; Blanchard, D.; le Pendu, J. Binding of Rabbit Hemorrhagic Disease Virus to Antigens of the ABH Histo-Blood Group Family. J. Virol. 2000, 74, 11950–11954. [Google Scholar] [CrossRef] [Green Version]
- Mikami, O.; Park, J.H.; Kimura, T.; Ochiai, K.; Itakura, C. Hepatic Lesions in Young Rabbits Experimentally Infected with Rabbit Haemorrhagic Disease Virus. Res. Vet. Sci. 1999, 66, 237–242. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, P.G.; Costa-e-Silva, A.; Monteiro, E.; Oliveira, M.J.R.; Águas, A.P. Transient Decrease in Blood Heterophils and Sustained Liver Damage Caused by Calicivirus Infection of Young Rabbits That Are Naturally Resistant to Rabbit Haemorrhagic Disease. Res. Vet. Sci. 2004, 76, 83–94. [Google Scholar] [CrossRef]
- Marques, R.M.; Teixeira, L.; Águas, A.P.; Ribeiro, J.C.; Costa-E-Silva, A.; Ferreira, P.G. Immunosuppression Abrogates Resistance of Young Rabbits to Rabbit Haemorrhagic Disease (RHD). Vet. Res. 2014, 45, 14. [Google Scholar] [CrossRef] [Green Version]
- Vallejo, D.; Crespo, I.; San-Miguel, B.; Álvarez, M.; Prieto, J.; Tuñón, M.J.; González-Gallego, J. Autophagic Response in the Rabbit Hemorrhagic Disease, an Animal Model of Virally-Induced Fulminant Hepatic Failure. Vet. Res. 2014, 45. [Google Scholar] [CrossRef] [Green Version]
- Neave, M.J.; Hall, R.N.; Huang, N.; McColl, K.A.; Kerr, P.; Hoehn, M.; Taylor, J.; Strive, T. Robust Innate Immunity of Young Rabbits Mediates Resistance to Rabbit Hemorrhagic Disease Caused by Lagovirus Europaeus GI.1 but Not GI.2. Viruses 2018, 10, 512. [Google Scholar] [CrossRef] [Green Version]
- Müller, C.; Ulrich, R.; Franzke, K.; Müller, M.; Köllner, B. Crude Extracts of Recombinant Baculovirus Expressing Rabbit Hemorrhagic Disease Virus 2 VLPs from Both Insect and Rabbit Cells Protect Rabbits from Rabbit Hemorrhagic Disease Caused by RHDV2. Arch. Virol. 2019, 164, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Prieto, J.M.B.; Felippe, M.J.B. Development, Phenotype, and Function of Non-Conventional B Cells. Comp. Immunol. Microbiol. Infect. Dis. 2017, 54, 38–44. [Google Scholar] [CrossRef]
- Wilkins, C.; Gale, M. Recognition of Viruses by Cytoplasmic Sensors. Curr. Opin. Immunol. 2010, 22, 41–47. [Google Scholar] [CrossRef] [Green Version]
- Braciale, T.J.; Hahn, Y.S. Immunity to Viruses. Immunol. Rev. 2013, 255, 5–12. [Google Scholar] [CrossRef] [Green Version]
- Makhluf, H.; Shresta, S. Innate Antiviral Immunity against Dengue Virus. Crit. Rev. Immunol. 2015, 35, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak-Ryczek, A.; Tokarz-Deptuła, B.; Deptuła, W. Expression of IL-1β, IL-2, IL-10, TNF-β and GM-CSF in Peripheral Blood Leukocytes of Rabbits Experimentally Infected with Rabbit Haemorrhagic Disease Virus. Vet. Microbiol. 2016, 186, 71–81. [Google Scholar] [CrossRef] [PubMed]
- Elfeil, W.M.K.; Algammal, A.M.; Abouelmaatti, R.R.; GErdouh, A.; Abdel-Daim, M.M. Molecular Characterization and Analysis of TLR-1 in Rabbit Tissues. Cent. Eur. J. Immunol. 2016, 41, 236–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, A.; Pergolizzi, S.; Cimino, F.; Lauriano, E.R.; Speciale, A.; D’Angelo, V.; Sicurella, M.; Argnani, R.; Manservigi, R.; Marconi, P. Role of Herpes Simplex Envelope Glycoprotein B and Toll-Like Receptor 2 in Ocular Inflammation: An Ex Vivo Organotypic Rabbit Corneal Model. Viruses 2019, 11, 819. [Google Scholar] [CrossRef] [Green Version]
- Astakhova, N.M.; Perelygin, A.A.; Zharkikh, A.A.; Lear, T.L.; Coleman, S.J.; MacLeod, J.N.; Brinton, M.A. Characterization of Equine and Other Vertebrate TLR3, TLR7, and TLR8 Genes. Immunogenetics 2009, 61, 529–539. [Google Scholar] [CrossRef]
- Kajikawa, O.; Frevert, C.W.; Lin, S.M.; Goodman, R.B.; Mongovin, S.M.; Wong, V.; Ballman, K.; Daubeuf, B.; Elson, G.; Martin, T.R. Gene Expression of Toll-like Receptor-2, Toll-like Receptor-4, and MD2 Is Differentially Regulated in Rabbits with Escherichia Coli Pneumonia. Gene 2005, 344, 193–202. [Google Scholar] [CrossRef]
- Lai, C.Y.; Liu, Y.L.; Yu, G.Y.; Maa, M.C.; Leu, T.H.; Xu, C.; Luo, Y.; Xiang, R.; Chuang, T.H. TLR7/8 Agonists Activate a Mild Immune Response in Rabbits through TLR8 but Not TLR7. Vaccine 2014, 32, 5593–5599. [Google Scholar] [CrossRef]
- Liu, J.; Xu, C.; Liu, Y.L.; Matsuo, H.; Pe-feng Hsieh, R.; Lo, J.F.; Tseng, P.H.; Yuan, C.J.; Luo, Y.; Xiang, R.; et al. Activation of Rabbit TLR9 by Different CpG-ODN Optimized for Mouse and Human TLR9. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 443–451. [Google Scholar] [CrossRef]
- Abrantes, J.; Areal, H.; Esteves, P.J. Insights into the European Rabbit (Oryctolagus Cuniculus) Innate Immune System: Genetic Diversity of the Toll-like Receptor 3 (TLR3) in Wild Populations and Domestic Breeds. BMC Genet. 2013, 14. [Google Scholar] [CrossRef] [Green Version]
- Laliena, A.; Miguel, B.S.; Crespo, I.; Alvarez, M.; González-Gallego, J.; Tuñón, M.J. Melatonin Attenuates Inflammation and Promotes Regeneration in Rabbits with Fulminant Hepatitis of Viral Origin. J. Pineal. Res. 2012, 53, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Guo, M.; Zhang, C.; Zhang, C.; Zhang, X.; Wu, Y. Functional Characterization of NLRX1 in Rabbit during Enterohemorrhagic Escherichia Coli Infection. Dev. Comp. Immunol. 2020, 106. [Google Scholar] [CrossRef] [PubMed]
- Lemos De Matos, A.; McFadden, G.; Esteves, P.J. Evolution of Viral Sensing RIG-I-like Receptor Genes in Leporidae Genera Oryctolagus, Sylvilagus, and Lepus. Immunogenetics 2014, 66, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Niedźwiedzka-Rystwej, P.; Tokarz-Deptuła, B.; Deptuła, W. Apoptosis of Granulocytes and Lymphocytes in Peripheral Blood in Rabbits Infected with Haemagglutinating and Non-Haemagglutinating Antigenic Variants of the RHD (Rabbit Haemorrhagic Disease) Virus. Pol. J. Vet. Sci. 2013, 16, 223–229. [Google Scholar] [CrossRef] [Green Version]
- Niedzwiedzka-Rystwej, P.; Tokarz-Deptula, B.; Hukowska-Szematowicz, B.; Dzialo, J.; Trzeciak-Ryczek, A.; Deptula, W. Peripheral Blood Lymphocytes B and T and Their Subpopulations in Rabbits Infected with Rabbit Haemorrhagic Disease Virus (RHDV). Med. Weter. 2013, 69, 484–487. [Google Scholar]
- Semerjyan, A.B.; Sargsyan, M.A.; Arzumanyan, H.H.; Hakobyan, L.H.; Abroyan, L.O.; Semerjyan, Z.B.; Avetisyan, A.S.; Karalova, E.M.; Manukyan, D.M.; Matevosyan, H.S.; et al. Immune Cell Pathology in Rabbit Hemorrhagic Disease. Vet. World 2019, 12, 1332–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Lastra, R.; San-Miguel, B.; Crespo, I.; Jorquera, F.; Alvarez, M.; González-Gallego, J.; Tuñón, M.J. Signaling Pathways Involved in Liver Injury and Regeneration in Rabbit Hemorrhagic Disease, an Animal Model of Virally-Induced Fulminant Hepatic Failure. Vet. Res. 2010, 41. [Google Scholar] [CrossRef] [Green Version]
- Tuñon, M.J.; San Miguel, B.; Crespo, I.; Riezu-Boj, J.I.; Larrea, E.; Alvarez, M.; Gonzalez, I.; Bustos, M.; Gonzalez-Gallego, J.; Prieto, J. Cardiotrophin-1 Promotes a High Survival Rate in Rabbits with Lethal Fulminant Hepatitis of Viral Origin. J. Virol. 2011, 85, 13124–13132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokarz-Deptuła, B. Immunity phenomena in rabbits infected with the RHD (rabbit haemorrhagic disease) virus. Pol. J. Environ. Stud. 2009, 7, 1–81. [Google Scholar]
- Bergin, I.L.; Wise, A.G.; Bolin, S.R.; Mullaney, T.P.; Kiupel, M.; Maes, R.K. Novel Calicivirus Identified in Rabbits, Michigan, USA. Emerg. Inf. Dis. 2009, 15, 1955–1962. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowska, E.; Kubes, P. Neutrophil Recruitment and Function in Health and Inflammation. Nat. Rev. Immunol. 2013, 13, 159–175. [Google Scholar] [CrossRef] [PubMed]
- Hrynkiewicz, R.; Bębnowska, D.; Niedźwiedzka-Rystwej, P. Myeloperoxidase and Lysozymes as a Pivotal Hallmark of Immunity Status in Rabbits. Animals 2020, 10, 1581. [Google Scholar] [CrossRef]
- Niedźwiedzka-Rystwej, P.; Deptuła, W. Non-Specific Immunity in Rabbits Infected with 10 Strains of the Rabbit Haemorrhagic Disease Virus with Different Biological Properties. Cent. Eur. J. Biol. 2010, 5, 613–632. [Google Scholar] [CrossRef]
- Zepeda-Cervantes, J.; Ramírez-Jarquín, J.O.; Vaca, L. Interaction Between Virus-Like Particles (VLPs) and Pattern Recognition Receptors (PRRs) From Dendritic Cells (DCs): Toward Better Engineering of VLPs. Front. Immunol. 2020, 11, 1100. [Google Scholar] [CrossRef]
- Ulsenheimer, A.; Gerlach, J.T.; Jung, M.C.; Gruener, N.; Wächtler, M.; Backmund, M.; Santantonio, T.; Schraut, W.; Heeg, M.H.J.; Schirren, C.A.; et al. Plasmacytoid Dendritic Cells in Acute and Chronic Hepatitis C Virus Infection. Hepatology 2005, 41, 643–651. [Google Scholar] [CrossRef]
- Bousarghin, L.; Hubert, P.; Franzen, E.; Jacobs, N.; Boniver, J.; Delvenne, P. Human Papillomavirus 16 Virus-like Particles Use Heparan Sulfates to Bind Dendritic Cells and Colocalize with Langerin in Langerhans Cells. J. Gen. Virol. 2005, 86, 1297–1305. [Google Scholar] [CrossRef] [PubMed]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major Findings and Recent Advances in Virus–like Particle (VLP)-Based Vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef] [PubMed]
- De Sousa-Pereira, P.; Abrantes, J.; Baldauf, H.-M.; Keppler, O.T.; Esteves, P.J. Evolutionary study of leporid CD4 reveals a hotspot of a genetic variability within the D2 domain. Immunogenetics 2016, 68, 477–482. [Google Scholar] [CrossRef]
- Bachmann, M.F.; Jennings, G.T. Vaccine Delivery: A Matter of Size, Geometry, Kinetics and Molecular Patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Serradell, M.C.; Rupil, L.L.; Martino, R.A.; Prucca, C.G.; Carranza, P.G.; Saura, A.; Fernández, E.A.; Gargantini, P.R.; Tenaglia, A.H.; Petiti, J.P.; et al. Efficient Oral Vaccination by Bioengineering Virus-like Particles with Protozoan Surface Proteins. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Peñaflor-Téllez, Y.; Trujillo-Uscanga, A.; Escobar-Almazán, J.A.; Gutiérrez-Escolano, A.L. Immune Response Modulation by Caliciviruses. Front. Immunol. 2019, 10, 2334. [Google Scholar] [CrossRef] [PubMed]
- Ramiro-Ibáñez, F.; Martín-Alonso, J.M.; García Palencia, P.; Parra, F.; Alonso, C. Macrophage Tropism of Rabbit Hemorrhagic Disease Virus Is Associated with Vascular Pathology. Virus Res. 1999, 60, 21–28. [Google Scholar] [CrossRef]
- Prieto, J.M.; Fernandez, F.; Alvarez, V.; Espi, A.; García Marín, J.F.; Alvarez, M.; Martín, J.M.; Parra, F. Immunohistochemical Localisation of Rabbit Haemorrhagic Disease Virus VP-60 Antigen in Early Infection of Young and Adult Rabbits. Res. Vet. Sci. 2000, 68, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Kimura, T.; Mitsui, I.; Okada, Y.; Furuya, T.; Ochiai, K.; Umemura, T.; Itakura, C. Distribution of Rabbit Haemorrhagic Disease Virus RNA in Experimentally Infected Rabbits. J. Comp. Pathol. 2001, 124, 134–141. [Google Scholar] [CrossRef]
- Boehm, T.; McCurley, N.; Sutoh, Y.; Schorpp, M.; Kasahara, M.; Cooper, M.D. VLR-Based Adaptive Immunity. Annu. Rev. Immunol. 2012, 30, 203–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Danilova, N. The Evolution of Adaptive Immunity. Adv. Exp. Med. Biol. 2012, 738, 218–235. [Google Scholar] [CrossRef]
- Farber, D.L.; Netea, M.G.; Radbruch, A.; Rajewsky, K.; Zinkernagel, R.M. Immunological Memory: Lessons from the Past and a Look to the Future. Nat. Rev. Immunol. 2016, 16, 124–128. [Google Scholar] [CrossRef] [PubMed]
- Chachu, K.A.; Strong, D.W.; LoBue, A.D.; Wobus, C.E.; Baric, R.S.; Virgin, H.W. Antibody Is Critical for the Clearance of Murine Norovirus Infection. J. Virol. 2008, 82, 6610–6617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindesmith, L.C.; McDaniel, J.R.; Changela, A.; Verardi, R.; Kerr, S.A.; Costantini, V.; Brewer-Jensen, P.D.; Mallory, M.L.; Voss, W.N.; Boutz, D.R.; et al. Sera Antibody Repertoire Analyses Reveal Mechanisms of Broad and Pandemic Strain Neutralizing Responses after Human Norovirus Vaccination. Immunity 2019, 50, 1530–1541.e8. [Google Scholar] [CrossRef] [Green Version]
- Valíček, L.; Šmíd, B.; Rodák, L.; Kudrna, J. Electron and Immunoelectron Microscopy of Rabbit Haemorrhagic Disease Virus (RHDV). Arch. Virol. 1990, 112, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Hu, Z.; Tian, X.; Zhai, Y.; Xu, W.; Zheng, D.; Sun, F. Cryo-electron microscopy reconstructions of two types of wild rabbit hemorrhagic disease viruses characterized the structural features of Lagovirus. Protein Cell 2010, 1, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Xu, F.; Liu, J.; Gao, B.; Liu, Y.; Zhai, Y.; Ma, J.; Zhang, K.; Baker, T.S.; Schulten, K.; et al. Atomic Model of Rabbit Hemorrhagic Disease Virus by Cryo-Electron Microscopy and Crystallography. PLoS Pathog. 2013, 9, e1003132. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.J.; White, J.R.; Lenghaus, C.; Boyd, V.; Westbury, H.A. A Competition ELISA for the Detection of Antibodies to Rabbit Haemorrhagic Disease Virus. Vet. Microbiol. 1995, 43, 85–96. [Google Scholar] [CrossRef]
- Jennewein, M.F.; Abu-Raya, B.; Jiang, Y.; Alter, G.; Marchant, A. Transfer of Maternal Immunity and Programming of the Newborn Immune System. Semin. Immunopathol. 2017, 39, 605–613. [Google Scholar] [CrossRef]
- Furukawa, S.; Kuroda, Y.; Sugiyama, A. A Comparison of the Histological Structure of the Placenta in Experimental Animals. J. Toxicol. Pathol. 2014, 27, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Marchandeau, S.; Chaval, Y.; Goff, E. le Prolonged Decline in the Abundance of Wild European Rabbits Oryctolagus Cuniculus and High Immunity Level over Three Years Following the Arrival of Rabbit Haemorrhagic Disease. Wildlife Biol. 2000, 6, 141–147. [Google Scholar] [CrossRef]
- Baratelli, M.; Molist-Badiola, J.; Puigredon-Fontanet, A.; Pascual, M.; Boix, O.; Mora-Igual, F.X.; Woodward, M.; Lavazza, A.; Capucci, L. Characterization of the Maternally Derived Antibody Immunity against Rhdv-2 after Administration in Breeding Does of an Inactivated Vaccine. Vaccines 2020, 8, 484. [Google Scholar] [CrossRef]
- Martens, C.L.; Currier, S.J.; Knight, K.L. Molecular genetic analysis of genes encoding the heavy chains of rabbit IgG. J. Immunol. 1984, 133, 1022–1027. [Google Scholar] [PubMed]
- Lavinder, J.J.; Hoi, K.H.; Reddy, S.T.; Wine, Y.; Georgiou, G. Systematic Characterization and Comparative Analysis of the Rabbit Immunoglobulin Repertoire. PLoS ONE 2014, 9, e101322. [Google Scholar] [CrossRef]
- Hayward, A.R.; Simons, M.A.; Lawton, A.R.; Mage, R.G.; Cooper, M.D. Pre-B and B Cells in Rabbits: Ontogeny and Allelic Exclusion of Kappa Light Chain Genes. J. Exp. Med. 1978, 148, 1367–1377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McElroy, P.J.; Willcox, N.; Catty, D. Early Precursors of B Lymphocytes I. Rabbit/Mouse Species Differences in the Physical Properties and Surface Phenotype of Pre-B Cells, and in the Maturation Sequence of Early B Cells. Eur. J. Immunol. 1981, 11, 76–85. [Google Scholar] [CrossRef] [PubMed]
- Jasper, P.J.; Zhai, S.-K.; Kalis, S.L.; Kingzette, M.; Knight, K.L. B Lymphocyte Development in Rabbit: Progenitor B Cells and Waning of B Lymphopoiesis. J. Immunol. 2003, 171, 6372–6380. [Google Scholar] [CrossRef] [Green Version]
- Neimanis, A.S.; Ahola, H.; Zohari, S.; Larsson Pettersson, U.; Bröjer, C.; Capucci, L.; Gavier-Widén, D. Arrival of Rabbit Haemorrhagic Disease Virus 2 to Northern Europe: Emergence and Outbreaks in Wild and Domestic Rabbits (Oryctolagus Cuniculus) in Sweden. Transbound. Emerg. Dis. 2018, 65, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.Y.; Lee, B.J.; Tai, J.H.; Park, J.H.; Lee, Y.S. Apoptosis in Rabbit Haemorrhagic Disease. J. Comp. Pathol. 2000, 123, 135–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, P.G.; Costa-E-Silva, A.; Oliveira, M.J.R.; Monteiro, E.; Águas, A.P. Leukocyte-Hepatocyte Interaction in Calicivirus Infection: Differences between Rabbits That Are Resistant or Susceptible to Rabbit Haemorrhagic Disease (RHD). Vet. Immunol. Immunopathol. 2005, 103, 217–221. [Google Scholar] [CrossRef]
- Schwensow, N.I.; Detering, H.; Pederson, S.; Mazzoni, C.; Sinclair, R.; Peacock, D.; Kovaliski, J.; Cooke, B.; Fickel, J.; Sommer, S. Resistance to RHD Virus in Wild Australian Rabbits: Comparison of Susceptible and Resistant Individuals Using a Genomewide Approach. Mol. Ecol. 2017, 26, 4551–4561. [Google Scholar] [CrossRef]
- Fitzner, A.; Niedbalski, W. Serological Survey for RHD Antibodies in Rabbits from Two Types of Rabbit Breeding Farms. Pol. J. Vet. Sci. 2016, 19, 597–607. [Google Scholar] [CrossRef] [Green Version]
- Cox, T.E.; Liu, J.; van de Ven, R.; Strive, T. Different Serological Profiles to Co-Occurring Pathogenic and Nonpathogenic Caliciviruses in Wild European Rabbits (Oryctolagus Cuniculus) across Australia. J. Wildl. Dis. 2017, 53, 472–481. [Google Scholar] [CrossRef]
- Forrester, N.L.; Boag, B.; Buckley, A.; Moureau, G.; Gould, E.A. Co-Circulation of Widely Disparate Strains of Rabbit Haemorrhagic Disease Virus Could Explain Localised Epidemicity in the United Kingdom. Virology 2009, 393, 42–48. [Google Scholar] [CrossRef]
- Hall, R.N.; Capucci, L.; Matthaei, M.; Esposito, S.; Kerr, P.J.; Frese, M.; Strive, T. An in Vivo System for Directed Experimental Evolution of Rabbit Haemorrhagic Disease Virus. PLoS ONE 2017, 12, e0173727. [Google Scholar] [CrossRef] [Green Version]
- Abade dos Santos, F.A.; Magro, C.; Carvalho, C.L.; Ruivo, P.; Duarte, M.D.; Peleteiro, M.C. A Potential Atypical Case of Rabbit Haemorrhagic Disease in a Dwarf Rabbit. Animals 2021, 11, 40. [Google Scholar] [CrossRef]
- Le Gall-Reculé, G.; Zwingelstein, F.; Fages, M.P.; Bertagnoli, S.; Gelfi, J.; Aubineau, J.; Roobrouck, A.; Botti, G.; Lavazza, A.; Marchandeau, S. Characterisation of a Non-Pathogenic and Non-Protective Infectious Rabbit Lagovirus Related to RHDV. Virology 2011, 410, 395–402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Gall-Reculé, G.; Zwingelstein, F.; Boucher, S.; le Normand, B.; Plassiart, G.; Portejoie, Y.; Decors, A.; Bertagnoli, S.; Guérin, J.L.; Marchandeau, S. Virology: Detection of a New Variant of Rabbit Haemorrhagic Disease Virus in France. Vet. Rec. 2011, 168, 137–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagesha, H.S.; Wang, L.F.; Hyatt, A.D.; Morrissy, C.J.; Lenghaus, C.; Westbury, H.A. Self-Assembly, Antigenicity, and Immunogenicity of the Rabbit Haemorrhagic Disease Virus (Czechoslovakian Strain V-351) Capsid Protein Expressed in Baculovirus. Arch. Virol. 1995, 140, 1095–1108. [Google Scholar] [CrossRef]
- Fernández, E.; Toledo, J.R.; Chiong, M.; Parra, F.; Rodríguez, E.; Montero, C.; Méndez, L.; Capucci, L.; Farnós, O. Single Dose Adenovirus Vectored Vaccine Induces a Potent and Long-Lasting Immune Response against Rabbit Hemorrhagic Disease Virus after Parenteral or Mucosal Administration. Vet. Immunol. Immunopathol. 2011, 142, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Fernández, E.; Toledo, J.R.; Méndez, L.; González, N.; Parra, F.; Martín-Alonso, J.M.; Limonta, M.; Sánchez, K.; Cabrales, A.; Estrada, M.P.; et al. Conformational and Thermal Stability Improvements for the Large-Scale Production of Yeast-Derived Rabbit Hemorrhagic Disease Virus-Like Particles as Multipurpose Vaccine. PLoS ONE 2013, 8, e56417. [Google Scholar] [CrossRef] [Green Version]
- Guo, H.; Zhu, J.; Tan, Y.; Li, C.; Chen, Z.; Sun, S.; Liu, G. Self-Assembly of Virus-like Particles of Rabbit Hemorrhagic Disease Virus Capsid Protein Expressed in Escherichia Coli and Their Immunogenicity in Rabbits. Antiviral Res. 2016, 131, 85–91. [Google Scholar] [CrossRef]
- Mikschofsky, H.; Schirrmeier, H.; Keil, G.M.; Lange, B.; Polowick, P.L.; Keller, W.; Broer, I. Pea-Derived Vaccines Demonstrate High Immunogenicity and Protection in Rabbits against Rabbit Haemorrhagic Disease Virus. Plant Biotechnol. J. 2009, 7, 537–549. [Google Scholar] [CrossRef]
- Grgacic, E.V.L.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef]
- Chen, Q.; Lai, H. Plant-Derived Virus-like Particles as Vaccines. Hum. Vaccin. Immunother. 2013, 9, 26–49. [Google Scholar] [CrossRef] [PubMed]
- Metz, S.W.; Martina, B.E.; van den Doel, P.; Geertsema, C.; Osterhaus, A.D.; Vlak, J.M.; Pijlman, G.P. Chikungunya Virus-like Particles Are More Immunogenic in a Lethal AG129 Mouse Model Compared to Glycoprotein E1 or E2 Subunits. Vaccine 2013, 31, 6092–6096. [Google Scholar] [CrossRef]
- Kushnir, N.; Streatfield, S.J.; Yusibov, V. Virus-like Particles as a Highly Efficient Vaccine Platform: Diversity of Targets and Production Systems and Advances in Clinical Development. Vaccine 2012, 31, 58–83. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wang, S.; Zhang, W.; Liu, X.; Yi, Y.; Yang, S.; Xia, X.; Li, Y.; Zhang, Z. Development of a VLP-Based Vaccine in Silkworm Pupae against Rabbit Hemorrhagic Disease Virus. Int. Immunopharmacol. 2016, 40, 164–169. [Google Scholar] [CrossRef] [PubMed]
- Raeven, R.H.M.; van Riet, E.; Meiring, H.D.; Metz, B.; Kersten, G.F.A. Systems Vaccinology and Big Data in the Vaccine Development Chain. Immunology 2019, 156, 33–46. [Google Scholar] [CrossRef] [Green Version]
- Rappuoli, R. Reverse Vaccinology. Curr. Opin. Microbiol. 2000, 3, 445–450. [Google Scholar] [CrossRef]
- Fiuza, T.S.; Lima, J.P.M.S.; de Souza, G.A. EpitoCore: Mining Conserved Epitope Vaccine Candidates in the Core Proteome of Multiple Bacteria Strains. Front. Immunol. 2020, 11, 816. [Google Scholar] [CrossRef] [PubMed]
- Jain, S.; Baranwal, M. Conserved Peptide Vaccine Candidates Containing Multiple Ebola Nucleoprotein Epitopes Display Interactions with Diverse HLA Molecules. Med. Microbiol. Immunol. 2019, 208, 227–238. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Vaccine | Containing RHDV Strains | Manufacturer | Date of Admission | Admission Number |
|---|---|---|---|---|
| RIKA-VACC RHD | RHDV GI.1 strain ‘Eisenhüttenstadt’ inactivated | Ecuphar NV | 04.09.2003 | 200a/91 |
| RIKA-VACC Duo | myxoma virus strain ‘CAMP V-219’ attenuated RHDV GI.1 strain ‘CAMP V-351’ inactivated | Ecuphar NV | 12.06.2008 | PEI.V.03071.01.1 |
| CUNIVAK RHD | RHDV GI.1 strain ‘Eisenhüttenstadt’ inactivated | Ceva Tiergesundheit GmbH | 11.05.2004 | 206a/92 |
| CUNIVAK COMBO | myxoma virus strain ‘CAMP V-219’ attenuated | Ceva Tiergesundheit GmbH | 05.08.2009 | PEI.V.07962.01.1 |
| RHDV GI.1 strain ‘CAMP V-351’ inactivated | ||||
| Eravac | RHDV GI.2 strain ‘V-1037’ inactivated | Laboratorios Hipra S.A. | 26.09.2016 | EU/2/16/199 |
| Filavac VHD K C+V | RHDV GI.1 strain ‘IM507.SC.2011’ inactivated | FILAVIE, Roussay | 13.03.2017 | PEI.V.11900.01.1 |
| RHDV GI.2 strain ‘LP.SV.2012’ inactivated |
| Genotype | GI.1; RHDV1 | GI.1a; RHDV1a | GI.2; RHDV2 |
|---|---|---|---|
| Place/year of detection | China, 1984 | Italy, 1997 | France, 2010 |
| Infected species | O. cuniculus | O. cuniculus | O. cuniculus several Lepus species |
| Average mortality rate | >90% | ≤100% | >90% |
| Disease symptoms | Typical for RHD | Typical for RHD | Typical for RHD |
| Juvenile resistance | until 9 weeks | until 4 weeks | |
| Cross protective in surviving animals | limited for RHDV2 | No | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Müller, C.; Hrynkiewicz, R.; Bębnowska, D.; Maldonado, J.; Baratelli, M.; Köllner, B.; Niedźwiedzka-Rystwej, P. Immunity against Lagovirus europaeus and the Impact of the Immunological Studies on Vaccination. Vaccines 2021, 9, 255. https://doi.org/10.3390/vaccines9030255
Müller C, Hrynkiewicz R, Bębnowska D, Maldonado J, Baratelli M, Köllner B, Niedźwiedzka-Rystwej P. Immunity against Lagovirus europaeus and the Impact of the Immunological Studies on Vaccination. Vaccines. 2021; 9(3):255. https://doi.org/10.3390/vaccines9030255
Chicago/Turabian StyleMüller, Claudia, Rafał Hrynkiewicz, Dominika Bębnowska, Jaime Maldonado, Massimiliano Baratelli, Bernd Köllner, and Paulina Niedźwiedzka-Rystwej. 2021. "Immunity against Lagovirus europaeus and the Impact of the Immunological Studies on Vaccination" Vaccines 9, no. 3: 255. https://doi.org/10.3390/vaccines9030255