
Immune Complex Vaccine Strategies to Combat HIV-1 and Other Infectious Diseases


Abstract
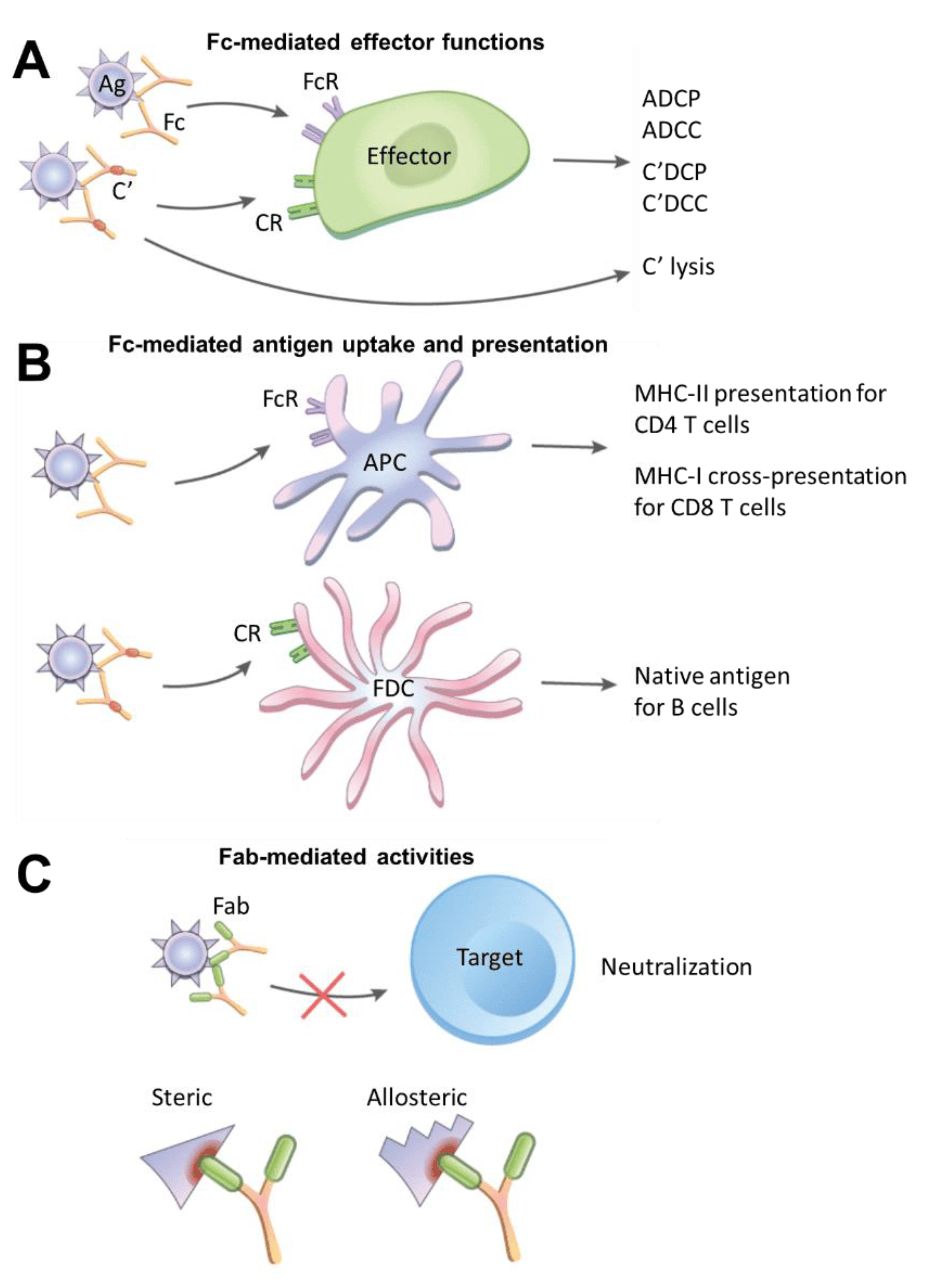
:1. Fc- and Fab-Mediated Activities of Immune Complexes
2. Immune Complex Vaccines against Human and Animal Pathogens

{kind=link}
{kind=link}
{kind=link}
| Pathogens | Immune Complex Components | Dosing Regimen and Study Design | Conferred Effects | Additional Notes | References |
|---|---|---|---|---|---|
| Hepatitis B Virus (HBV) | Yeast-derived hepatitis B surface antigen (HBsAg) complexed to human anti-HBs immunoglobulin (HBIG) | Phase IIb trial; 6 IM injections of 30 µg, 60 µg, or placebo at 4-week intervals, follow-up for 24 weeks after the last injection | No significant difference of group effects between 60 µg group vs. placebo in 1° endpoints (loss of HBeAg or induction of anti-HBe antibody or suppression of HBV DNA) A significant difference in HBe seroconversion (loss of HBeAg and anti-HBe seropositive) between 60 µg group (21.8%) vs. placebo (9%) at end of follow-up | Follow-up phase III trial utilizing 12 IM injections of 60 µg IC yielded lower HBe seroconversion compared to 6 IM injections in phase IIb (14.0% vs. 21.8%) | [27,36] |
| Infectious Bursal Disease Virus (IBDV) | Bursaplex® vaccine containing a live strain of IBDV of chicken embryo origin and IBD antiserum | 1-day old commercial broiler chicks immunized SC with IC vaccine; compared with recombinant turkey herpesvirus vector vaccine (HVT-IBD, Vaxxitek®), live attenuated vaccine, and killed vaccine; challenged with IBDV at week 4 | Bursaplex IC vaccine protected 100% of animals from clinical symptoms and mortality after IBDV challenge; still resulted in bursal atrophy and bursal lesions | IC vaccine resulted in low anti-IBDV antibody titers; viral vaccine component neutralized by maternal antibodies Vector HVT-IBD vaccine deemed superior, rendered 100% protection while avoiding bursal atrophy and lesions | [32] |
| Equine Herpesvirus-1 (EHV-1) | Solid matrix-antibody-antigen complex, consisting of MAb bound via Fc to Staphylococcus aureus and via Fab to EHV-1 glycoproteins C or D | BALB/c or C3H mice immunized IP on days 0, 28, and 43, followed by IN EHV-1 challenge on day 53 | Both neutralizing and lysis-mediating antibodies elicited against respective glycoproteins, along with priming of EHV-1-specific local and systemic T cell proliferative responses 35% reduction of virus load in turbinates of mice immunized with glycoprotein D, but not of mice immunized with glycoprotein C | Formation of CD8+ T suppressor cells in the cervical lymph nodes of glycoprotein C-immunized mice In the lungs, no significant difference in virus load of mice immunized with glycoproteins D and C compared to control | [34] |
| Porcine Parvovirus (PPV) | Immuno-precipitate of <1000 kDa PPV viral particles and pig or rabbit anti-PPV polyclonal immune sera. IC consisted mostly of PPV VP2 protein and IgG | Gilts immunized IM six weeks before mating and boosted two weeks later; chinchilla rabbits immunized IM once | Generation of anti-PPV antibodies in gilts and rabbits starting from 2 weeks post-vaccination. IC-induced antibody titers comparable to titers induced by Parvokal®, a commercial inactivated PPV vaccine | IC vaccine was dose-sparing, using half the amount of virus in Parvokal | [33] |
| Tick-Borne Encephalitis Virus (TBEV) | TBEV sE protein complexed with IgG against one of three distinct epitopes | C57BL/6 mice immunized IP twice with an interval of 14 days, boosted 8 weeks later | IC-immunized mice generated comparable titers of serum anti-sE antibodies as mice immunized with sE only at 22 weeks post-immunization. | For two of the three IC vaccines, fine specificities of antibody responses differed from the sE-only vaccinated group. Evidence of antibody-induced conformational changes and epitope shielding in the two IC vaccines | [37] |
| Francisella tularensis | Inactivated bacteria complexed with mouse IgG2a anti-F. tularensis mAb | Mice immunized IN and boosted on day 21, followed by IN bacterial challenge on day 35 | Significant enhancement in the survival of mice vaccinated with IC vaccine vs. non-complexed bacteria IC vaccination led to a 10-fold decrease in bacterial burden, tissue inflammation, and cytokine production; enhanced production of protective mucosal IgA | Immunomodulatory benefits of IC vaccine attributed to FcR targeting on mucosal antigen-presenting cells | [35] |
3. Immune Complex Vaccine Strategies against HIV-1
3.1. IC Vaccines to Elicit Antibody Responses against V3
3.2. IC Vaccines to Elicit Antibody Responses against V1V2
3.3. IC Vaccines to Elicit bNAb Responses
| Strategy | IC Components | Species Used in Study | Immunomodulatory Effects | Additional Notes | References |
|---|---|---|---|---|---|
| Targeting V3 | gp120 JRFL or LAI (both clade B) + anti-CD4bs mAb 654 | BALB/c mice | Both ICs elicited greater titers of antibody responses to V3 Anti-V3 antibodies generated by JRFL IC had limited neutralizing activity but were more cross-reactive than those by LAI IC | Enhanced V3 immunogenicity of ICs correlated with greater antigenicity | [44,49] |
| Targeting V3 | gp120 JRFL (clade B) + anti-CD4bs mAb 654, anti-V2i mAb 2158, or anti-C2 mAb 1006-30 | BALB/c mice | ICs with CD4bs mAb 654 or V2i mAb 2158 (to a lesser extent) enhanced V3 antibody responses IC with C2 mAb 1006-30 reduced V3 antibody responses | ICs with F(ab)2 654 were sufficient in enhancing V3 antibody responses, highlighting the key role of Fab | [48] |
| Targeting V3 Masking V3 Targeting V1V2 | gp120 JRFL (clade B) + anti-CD4bs mAb 654, anti-V2i mAb 2158 gp120 A244 (CRF_01.AE) + anti-V2i mAb 2158 | BALB/c mice | JRFL ICs elicited V3 antibody responses of greater titers and breadth, and with more tier 1 virus neutralizing activity A244 ICs induced higher levels of V1V2 antibodies with some cross-reactivity | JRFL ICs with C2 or V3 mAbs reduced V3 antibody response JRFL and A244 ICs modulated antibody responses to V1V2 and V3 without affecting the overall antibody responses to HIV-1 Env | [43] |
| Stabilizing CD4i (chemokine receptor binding site) | gp120 BaL or 89.6 (clade B) + mAb A32 | Outbred Harley guinea pigs | IC enhanced exposure and antigenicity of CD4i in vitro IC caused no change in CD4i immunogenicity and neutralizing antibody responses in vivo | Enhanced CD4i antigenicity in vitro did not translate to enhanced immunogenicity in vivo | [67] |
| Masking CD4i Targeting CD4bs bNAb (VRC01 lineage) | gp120 core + anti-CD4i mAb 17b | New Zealand White rabbits | IC suppressed antibodies against the CD4i bridging sheet, elicited tier 1 neutralization, transiently induced antibody response with similar binding profile to VRC01-class CD4bs bnAbs No enhanced and long-term induction of VRC01-like Abs | 17b mAb blocked CD4i bridging sheet and non-neutralizing CD4bs while exposing CD4bs for VRC01 approach from an alternate angle | [70] |
| Masking glycan hole | BG505 SOSIP.664 gp140 trimer (clade A) + mAbs that target strain-specific glycan hole | New Zealand White rabbits | ICs elicited lower levels of strain-specific antibody responses, indicating successful blockage of immunodominant glycan-hole region ICs stimulated binding antibodies with a lower rate of decay | Diversion away from glycan hole did not improve antibody responses against cross-reactive neutralizing epitopes | [68] |
3.4. Fc Functions in IC Vaccines
3.5. Lessons from Antibody Passive Transfer Studies
4. Challenges in the IC Vaccine Development
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vorup-Jensen, T.; Jensen, R.K. Structural Immunology of Complement Receptors 3 and 4. Front. Immunol. 2018, 9, 2716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournazos, S.; Ravetch, J.V. Fcgamma Receptor Function and the Design of Vaccination Strategies. Immunity 2017, 47, 224–233. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Ravetch, J.V. Diversification of IgG effector functions. Int. Immunol. 2017, 29, 303–310. [Google Scholar] [PubMed] [Green Version]
- Sondermann, P.; Pincetic, A.; Maamary, J.; Lammens, K.; Ravetch, J.V. General mechanism for modulating immunoglobulin effector function. Proc. Natl. Acad. Sci. USA 2013, 110, 9868–9872. [Google Scholar] [CrossRef] [Green Version]
- Boune, S.; Hu, P.; Epstein, A.L.; Khawli, L.A. Principles of N-Linked Glycosylation Variations of IgG-Based Therapeutics: Pharmacokinetic and Functional Considerations. Antibodies 2020, 9, 22. [Google Scholar] [CrossRef]
- Hayes, J.M.; Wormald, M.R.; Rudd, P.M.; Davey, G.P. Fc gamma receptors: Glycobiology and therapeutic prospects. J. Inflamm. Res. 2016, 9, 209–219. [Google Scholar] [CrossRef] [Green Version]
- Temming, A.R.; de Taeye, S.W.; de Graaf, E.L.; de Neef, L.A.; Dekkers, G.; Bruggeman, C.W.; Koers, J.; Ligthart, P.; Nagelkerke, S.Q.; Zimring, J.C.; et al. Functional Attributes of Antibodies, Effector Cells, and Target Cells Affecting NK Cell-Mediated Antibody-Dependent Cellular Cytotoxicity. J. Immunol. 2019, 203, 3126–3135. [Google Scholar] [CrossRef]
- Lewis, G.K.; Ackerman, M.E.; Scarlatti, G.; Moog, C.; Robert-Guroff, M.; Kent, S.J.; Overbaugh, J.; Reeves, R.K.; Ferrari, G.; Thyagarajan, B. Knowns and Unknowns of Assaying Antibody-Dependent Cell-Mediated Cytotoxicity Against HIV-1. Front. Immunol. 2019, 10, 1025. [Google Scholar] [CrossRef] [Green Version]
- Pincetic, A.; Bournazos, S.; DiLillo, D.J.; Maamary, J.; Wang, T.T.; Dahan, R.; Fiebiger, B.-M.; Ravetch, J.V. Type I and type II Fc receptors regulate innate and adaptive immunity. Nat. Immunol. 2014, 15, 707–716. [Google Scholar] [CrossRef]
- Liu, C.; Richard, K.; Wiggins, M.; Zhu, X.; Conrad, D.H.; Song, W. CD23 can negatively regulate B-cell receptor signaling. Sci. Rep. 2016, 6, 25629. [Google Scholar] [CrossRef] [Green Version]
- Heyman, B. The immune complex: Possible ways of regulating the antibody response. Immunol. Today 1990, 11, 310–313. [Google Scholar] [CrossRef]
- Regnault, A.; Lankar, D.; Lacabanne, V.; Rodriguez, A.; Thery, C.; Rescigno, M.; Saito, T.; Verbeek, S.; Bonnerot, C.; Ricciardi-Castagnoli, P.; et al. Fcgamma receptor-mediated induction of dendritic cell maturation and major histocompatibility complex class I-restricted antigen presentation after immune complex internalization. J. Exp. Med. 1999, 189, 371–380. [Google Scholar] [CrossRef] [PubMed]
- Manca, F.; Fenoglio, D.; Kunkl, A.; Cambiaggi, C.; Sasso, M.; Celada, F. Differential activation of T cell clones stimulated by macrophages exposed to antigen complexed with monoclonal antibodies. A possible influence of paratope specificity on the mode of antigen processing. J. Immunol. 1988, 140, 2893–2898. [Google Scholar] [PubMed]
- Cornacoff, J.B.; Hebert, L.A.; Smead, W.L.; VanAman, M.E.; Birmingham, D.J.; Waxman, F.J. Primate erythrocyte-immune complex-clearing mechanism. J. Clin. Investig. 1983, 71, 236–247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellner, C.; Otte, A.; Cappuzzello, E.; Klausz, K.; Peipp, M. Modulating Cytotoxic Effector Functions by Fc Engineering to Improve Cancer Therapy. Transfus Med. Hemother 2017, 44, 327–336. [Google Scholar] [CrossRef] [Green Version]
- Lee, C.H.; Romain, G.; Yan, W.; Watanabe, M.; Charab, W.; Todorova, B.; Lee, J.; Triplett, K.; Donkor, M.; Lungu, O.I.; et al. IgG Fc domains that bind C1q but not effector Fcgamma receptors delineate the importance of complement-mediated effector functions. Nat. Immunol. 2017, 18, 889–898. [Google Scholar] [CrossRef]
- Kranich, J.; Krautler, N.J. How Follicular Dendritic Cells Shape the B-Cell Antigenome. Front. Immunol. 2016, 7, 225. [Google Scholar] [CrossRef] [Green Version]
- Heesters, B.A.; Chatterjee, P.; Kim, Y.A.; Gonzalez, S.F.; Kuligowski, M.P.; Kirchhausen, T.; Carroll, M.C. Endocytosis and recycling of immune complexes by follicular dendritic cells enhances B cell antigen binding and activation. Immunity 2013, 38, 1164–1175. [Google Scholar] [CrossRef] [Green Version]
- Kosco, M.H.; Monfalcone, A.P.; Szakal, A.K.; Tew, J.G. Germinal center B cells present antigen obtained in vivo to T cells in vitro and stimulate mixed lymphocyte reactions. Adv. Exp. Med. Biol. 1988, 237, 883–888. [Google Scholar] [CrossRef]
- Suzuki, K.; Grigorova, I.; Phan, T.G.; Kelly, L.M.; Cyster, J.G. Visualizing B cell capture of cognate antigen from follicular dendritic cells. J. Exp. Med. 2009, 206, 1485–1493. [Google Scholar] [CrossRef] [Green Version]
- Manca, F.; Fenoglio, D.; Li Pira, G.; Kunkl, A.; Celada, F. Effect of antigen/antibody ratio on macrophage uptake, processing, and presentation to T cells of antigen complexed with polyclonal antibodies. J. Exp. Med. 1991, 173, 37–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mok, D.Z.L.; Chan, K.R. The Effects of Pre-Existing Antibodies on Live-Attenuated Viral Vaccines. Viruses 2020, 12, 1520. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.M. Antigen-antibody immunogenic complex: Promising novel vaccines for microbial persistent infections. Expert Opin. Biol. 2009, 9, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.Z.; Huang, K.L.; Zhao, K.; Xu, L.F.; Shi, N.; Yuan, Z.H.; Wen, Y.M. Vaccination with recombinant HBsAg-HBIG complex in healthy adults. Vaccine 2005, 23, 2658–2664. [Google Scholar] [CrossRef]
- Zhou, C.; Li, C.; Gong, G.Z.; Wang, S.; Zhang, J.M.; Xu, D.Z.; Guo, L.M.; Ren, H.; Xu, M.; Xie, Q.; et al. Analysis of immunological mechanisms exerted by HBsAg-HBIG therapeutic vaccine combined with Adefovir in chronic hepatitis B patients. Hum. Vaccin. Immunother. 2017, 13, 1989–1996. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.Z.; Zhao, K.; Guo, L.M.; Li, L.J.; Xie, Q.; Ren, H.; Zhang, J.M.; Xu, M.; Wang, H.F.; Huang, W.X.; et al. A randomized controlled phase IIb trial of antigen-antibody immunogenic complex therapeutic vaccine in chronic hepatitis B patients. PLoS ONE 2008, 3, e2565. [Google Scholar] [CrossRef]
- Li, J.; Bao, M.; Ge, J.; Ren, S.; Zhou, T.; Qi, F.; Pu, X.; Dou, J. Research progress of therapeutic vaccines for treating chronic hepatitis B. Hum. Vaccin. Immunother. 2017, 13, 986–997. [Google Scholar] [CrossRef] [Green Version]
- Han, J.; Gong, G.Z.; Lei, J.H.; Qin, W.J.; Qin, R.H.; Wang, X.Y.; Gu, J.X.; Ren, S.F.; Wen, Y.M. Response to immune complex vaccine in chronic hepatitis B patients is associated with lower baseline level of serum IgG galactosylation. Medicine 2019, 98, e16208. [Google Scholar] [CrossRef]
- Peschke, B.; Keller, C.W.; Weber, P.; Quast, I.; Lunemann, J.D. Fc-Galactosylation of Human Immunoglobulin Gamma Isotypes Improves C1q Binding and Enhances Complement-Dependent Cytotoxicity. Front. Immunol. 2017, 8, 646. [Google Scholar] [CrossRef]
- Thomann, M.; Reckermann, K.; Reusch, D.; Prasser, J.; Tejada, M.L. Fc-galactosylation modulates antibody-dependent cellular cytotoxicity of therapeutic antibodies. Mol. Immunol. 2016, 73, 69–75. [Google Scholar] [CrossRef]
- Sedeik, M.E.; El-Shall, N.A.; Awad, A.M.; Abd El-Hack, M.E.; Alowaimer, A.N.; Swelum, A.A. Comparative Evaluation of HVT-IBD Vector, Immune Complex, and Live IBD Vaccines against vvIBDV in Commercial Broiler Chickens with High Maternally Derived Antibodies. Animals 2019, 9, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roic, B.; Cajavec, S.; Ergotic, N.; Lipej, Z.; Madic, J.; Lojkic, M.; Pokric, B. Immune complex-based vaccine for pig protection against parvovirus. J. Vet. Med. B Infect. Dis. Vet. Public Health 2006, 53, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Alber, D.G.; Killington, R.A.; Stokes, A. Solid matrix-antibody-antigen complexes incorporating equine herpesvirus 1 glycoproteins C and D elicit anti-viral immune responses in BALB/c (H-2K(d)) and C3H (H-2K(k)) mice. Vaccine 2000, 19, 895–901. [Google Scholar] [CrossRef]
- Rawool, D.B.; Bitsaktsis, C.; Li, Y.; Gosselin, D.R.; Lin, Y.; Kurkure, N.V.; Metzger, D.W.; Gosselin, E.J. Utilization of Fc Receptors as a Mucosal Vaccine Strategy against an Intracellular Bacterium, Francisella tularensis. J. Immunol. 2008, 180, 5548–5557. [Google Scholar] [CrossRef] [PubMed]
- Tsouchnikas, G.; Zlatkovic, J.; Jarmer, J.; Strauss, J.; Vratskikh, O.; Kundi, M.; Stiasny, K.; Heinz, F.X. Immunization with Immune Complexes Modulates the Fine Specificity of Antibody Responses to a Flavivirus Antigen. J. Virol. 2015, 89, 7970–7978. [Google Scholar] [CrossRef] [Green Version]
- UNAIDS. UN AIDS DATA 2020. Available online: https://www.unaids.org/sites/default/files/media_asset/2020_aids-data-book_en.pdf (accessed on 22 January 2021).
- Xu, D.Z.; Wang, X.Y.; Shen, X.L.; Gong, G.Z.; Ren, H.; Guo, L.M.; Sun, A.M.; Xu, M.; Li, L.J.; Guo, X.H.; et al. Results of a phase III clinical trial with an HBsAg-HBIG immunogenic complex therapeutic vaccine for chronic hepatitis B patients: Experiences and findings. J. Hepatol. 2013, 59, 450–456. [Google Scholar] [CrossRef]
- Rerks-Ngarm, S.; Pitisuttithum, P.; Nitayaphan, S.; Kaewkungwal, J.; Chiu, J.; Paris, R.; Premsri, N.; Namwat, C.; de Souza, M.; Adams, E.; et al. Vaccination with ALVAC and AIDSVAX to prevent HIV-1 infection in Thailand. N. Engl. J. Med. 2009, 361, 2209–2220. [Google Scholar] [CrossRef]
- Haynes, B.F.; Gilbert, P.B.; McElrath, M.J.; Zolla-Pazner, S.; Tomaras, G.D.; Alam, S.M.; Evans, D.T.; Montefiori, D.C.; Karnasuta, C.; Sutthent, R.; et al. Immune-correlates analysis of an HIV-1 vaccine efficacy trial. N. Engl. J. Med. 2012, 366, 1275–1286. [Google Scholar] [CrossRef] [Green Version]
- Chung, A.W.; Ghebremichael, M.; Robinson, H.; Brown, E.; Choi, I.; Lane, S.; Dugast, A.S.; Schoen, M.K.; Rolland, M.; Suscovich, T.J.; et al. Polyfunctional Fc-effector profiles mediated by IgG subclass selection distinguish RV144 and VAX003 vaccines. Sci. Transl. Med. 2014, 6, 228ra238. [Google Scholar] [CrossRef]
- Yates, N.L.; Liao, H.X.; Fong, Y.; deCamp, A.; Vandergrift, N.A.; Williams, W.T.; Alam, S.M.; Ferrari, G.; Yang, Z.Y.; Seaton, K.E.; et al. Vaccine-induced Env V1-V2 IgG3 correlates with lower HIV-1 infection risk and declines soon after vaccination. Sci. Transl. Med. 2014, 6, 228ra239. [Google Scholar] [CrossRef] [Green Version]
- Hioe, C.E.; Kumar, R.; Upadhyay, C.; Jan, M.; Fox, A.; Itri, V.; Peachman, K.K.; Rao, M.; Liu, L.; Lo, N.C.; et al. Modulation of Antibody Responses to the V1V2 and V3 Regions of HIV-1 Envelope by Immune Complex Vaccines. Front. Immunol. 2018, 9, 2441. [Google Scholar] [CrossRef] [PubMed]
- Hioe, C.E.; Visciano, M.L.; Kumar, R.; Liu, J.; Mack, E.A.; Simon, R.E.; Levy, D.N.; Tuen, M. The use of immune complex vaccines to enhance antibody responses against neutralizing epitopes on HIV-1 envelope gp120. Vaccine 2009, 28, 352–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, R.; Tuen, M.; Li, H.; Tse, D.B.; Hioe, C.E. Improving immunogenicity of HIV-1 envelope gp120 by glycan removal and immune complex formation. Vaccine 2011, 29, 9064–9074. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Tuen, M.; Liu, J.; Nadas, A.; Pan, R.; Kong, X.; Hioe, C.E. Elicitation of broadly reactive antibodies against glycan-modulated neutralizing V3 epitopes of HIV-1 by immune complex vaccines. Vaccine 2013, 31, 5413–5421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visciano, M.L.; Tuen, M.; Chen, P.D.; Hioe, C.E. Antibodies to the CD4-binding site of HIV-1 gp120 suppress gp120-specific CD4 T cell response while enhancing antibody response. Infect. Agent Cancer 2008, 3, 11. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Visciano, M.L.; Li, H.; Hioe, C. Targeting a Neutralizing Epitope of HIV Envelope Gp120 by Immune Complex Vaccine. J. Aids Clin. Res. 2012, S8, 5512. [Google Scholar] [CrossRef] [Green Version]
- Visciano, M.L.; Tuen, M.; Gorny, M.K.; Hioe, C.E. In vivo alteration of humoral responses to HIV-1 envelope glycoprotein gp120 by antibodies to the CD4-binding site of gp120. Virology 2008, 372, 409–420. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Huber, M.; Doores, K.J.; Falkowska, E.; Pejchal, R.; Julien, J.P.; Wang, S.K.; Ramos, A.; Chan-Hui, P.Y.; Moyle, M.; et al. Broad neutralization coverage of HIV by multiple highly potent antibodies. Nature 2011, 477, 466–470. [Google Scholar] [CrossRef] [Green Version]
- Doores, K.J.; Kong, L.; Krumm, S.A.; Le, K.M.; Sok, D.; Laserson, U.; Garces, F.; Poignard, P.; Wilson, I.A.; Burton, D.R. Two classes of broadly neutralizing antibodies within a single lineage directed to the high-mannose patch of HIV envelope. J. Virol. 2015, 89, 1105–1118. [Google Scholar] [CrossRef] [Green Version]
- Julien, J.P.; Sok, D.; Khayat, R.; Lee, J.H.; Doores, K.J.; Walker, L.M.; Ramos, A.; Diwanji, D.C.; Pejchal, R.; Cupo, A.; et al. Broadly neutralizing antibody PGT121 allosterically modulates CD4 binding via recognition of the HIV-1 gp120 V3 base and multiple surrounding glycans. PLoS Pathog. 2013, 9, e1003342. [Google Scholar] [CrossRef]
- Tuen, M.; Visciano, M.L.; Chien, P.C., Jr.; Cohen, S.; Chen, P.D.; Robinson, J.; He, Y.; Pinter, A.; Gorny, M.K.; Hioe, C.E. Characterization of antibodies that inhibit HIV gp120 antigen processing and presentation. Eur. J. Immunol. 2005, 35, 2541–2551. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Hioe, C.E.; Swetnam, J.; Zolla-Pazner, S.; Cardozo, T. Quantitative assessment of masking of neutralization epitopes in HIV-1. Vaccine 2011, 29, 6736–6741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Upadhyay, C.; Feyznezhad, R.; Yang, W.; Zhang, H.; Zolla-Pazner, S.; Hioe, C.E. Alterations of HIV-1 envelope phenotype and antibody-mediated neutralization by signal peptide mutations. PLoS Pathog. 2018, 14, e1006812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zolla-Pazner, S.; Cohen, S.S.; Boyd, D.; Kong, X.P.; Seaman, M.; Nussenzweig, M.; Klein, F.; Overbaugh, J.; Totrov, M. Structure/Function Studies Involving the V3 Region of the HIV-1 Envelope Delineate Multiple Factors That Affect Neutralization Sensitivity. J. Virol. 2016, 90, 636–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balasubramanian, P.; Kumar, R.; Williams, C.; Itri, V.; Wang, S.; Lu, S.; Hessell, A.J.; Haigwood, N.L.; Sinangil, F.; Higgins, K.W.; et al. Differential induction of anti-V3 crown antibodies with cradle- and ladle-binding modes in response to HIV-1 envelope vaccination. Vaccine 2017, 35, 1464–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hessell, A.J.; Powell, R.; Jiang, X.; Luo, C.; Weiss, S.; Dussupt, V.; Itri, V.; Fox, A.; Shapiro, M.B.; Pandey, S.; et al. Multimeric Epitope-Scaffold HIV Vaccines Target V1V2 and Differentially Tune Polyfunctional Antibody Responses. Cell Rep. 2019, 28, 877–895.e6. [Google Scholar] [CrossRef] [Green Version]
- Lyumkis, D.; Julien, J.P.; de Val, N.; Cupo, A.; Potter, C.S.; Klasse, P.J.; Burton, D.R.; Sanders, R.W.; Moore, J.P.; Carragher, B.; et al. Cryo-EM structure of a fully glycosylated soluble cleaved HIV-1 envelope trimer. Science 2013, 342, 1484–1490. [Google Scholar] [CrossRef] [Green Version]
- McLellan, J.S.; Pancera, M.; Carrico, C.; Gorman, J.; Julien, J.P.; Khayat, R.; Louder, R.; Pejchal, R.; Sastry, M.; Dai, K.; et al. Structure of HIV-1 gp120 V1/V2 domain with broadly neutralizing antibody PG9. Nature 2011, 480, 336–343. [Google Scholar] [CrossRef]
- Pan, R.; Gorny, M.K.; Zolla-Pazner, S.; Kong, X.P. The V1V2 Region of HIV-1 gp120 Forms a Five-Stranded Beta Barrel. J. Virol. 2015, 89, 8003–8010. [Google Scholar] [CrossRef] [Green Version]
- Pancera, M.; Shahzad-Ul-Hussan, S.; Doria-Rose, N.A.; McLellan, J.S.; Bailer, R.T.; Dai, K.; Loesgen, S.; Louder, M.K.; Staupe, R.P.; Yang, Y.; et al. Structural basis for diverse N-glycan recognition by HIV-1-neutralizing V1-V2-directed antibody PG16. Nat. Struct. Mol. Biol. 2013, 20, 804–813. [Google Scholar] [CrossRef] [Green Version]
- Van Eeden, C.; Wibmer, C.K.; Scheepers, C.; Richardson, S.I.; Nonyane, M.; Lambson, B.; Mkhize, N.N.; Vijayakumar, B.; Sheng, Z.; Stanfield-Oakley, S.; et al. V2-Directed Vaccine-like Antibodies from HIV-1 Infection Identify an Additional K169-Binding Light Chain Motif with Broad ADCC Activity. Cell Rep. 2018, 25, 3123–3135.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.X.; Bonsignori, M.; Alam, S.M.; McLellan, J.S.; Tomaras, G.D.; Moody, M.A.; Kozink, D.M.; Hwang, K.K.; Chen, X.; Tsao, C.Y.; et al. Vaccine induction of antibodies against a structurally heterogeneous site of immune pressure within HIV-1 envelope protein variable regions 1 and 2. Immunity 2013, 38, 176–186. [Google Scholar] [CrossRef] [Green Version]
- Jia, M.; Liberatore, R.A.; Guo, Y.; Chan, K.W.; Pan, R.; Lu, H.; Waltari, E.; Mittler, E.; Chandran, K.; Finzi, A.; et al. VSV-Displayed HIV-1 Envelope Identifies Broadly Neutralizing Antibodies Class-Switched to IgG and IgA. Cell Host Microbe 2020, 27, 963–975.e5. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Wilson, R.; O’Dell, S.; Guenaga, J.; Feng, Y.; Tran, K.; Chiang, C.I.; Arendt, H.E.; DeStefano, J.; Mascola, J.R.; et al. An HIV-1 Env-Antibody Complex Focuses Antibody Responses to Conserved Neutralizing Epitopes. J. Immunol. 2016, 197, 3982–3998. [Google Scholar] [CrossRef]
- Tolbert, W.D.; Sherburn, R.T.; Van, V.; Pazgier, M. Structural Basis for Epitopes in the gp120 Cluster A Region that Invokes Potent Effector Cell Activity. Viruses 2019, 11, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, H.X.; Alam, S.M.; Mascola, J.R.; Robinson, J.; Ma, B.; Montefiori, D.C.; Rhein, M.; Sutherland, L.L.; Scearce, R.; Haynes, B.F. Immunogenicity of constrained monoclonal antibody A32-human immunodeficiency virus (HIV) Env gp120 complexes compared to that of recombinant HIV type 1 gp120 envelope glycoproteins. J. Virol. 2004, 78, 5270–5278. [Google Scholar] [CrossRef] [Green Version]
- Gach, J.S.; Mara, K.J.V.; LaBranche, C.C.; van Gils, M.J.; McCoy, L.E.; Klasse, P.J.; Montefiori, D.C.; Sanders, R.W.; Moore, J.P.; Forthal, D.N. Antibody Responses Elicited by Immunization with BG505 Trimer Immune Complexes. J. Virol. 2019, 93. [Google Scholar] [CrossRef] [Green Version]
- Malherbe, D.C.; Wibmer, C.K.; Nonyane, M.; Reed, J.; Sather, D.N.; Spencer, D.A.; Schuman, J.T.; Guo, B.; Pandey, S.; Robins, H.; et al. Rapid Induction of Multifunctional Antibodies in Rabbits and Macaques by Clade C HIV-1 CAP257 Envelopes Circulating During Epitope-Specific Neutralization Breadth Development. Front. Immunol. 2020, 11, 984. [Google Scholar] [CrossRef]
- Doria-Rose, N.A.; Bhiman, J.N.; Roark, R.S.; Schramm, C.A.; Gorman, J.; Chuang, G.Y.; Pancera, M.; Cale, E.M.; Ernandes, M.J.; Louder, M.K.; et al. New Member of the V1V2-Directed CAP256-VRC26 Lineage That Shows Increased Breadth and Exceptional Potency. J. Virol. 2016, 90, 76–91. [Google Scholar] [CrossRef] [Green Version]
- Chien, P.C., Jr.; Cohen, S.; Tuen, M.; Arthos, J.; Chen, P.D.; Patel, S.; Hioe, C.E. Human immunodeficiency virus type 1 evades T-helper responses by exploiting antibodies that suppress antigen processing. J. Virol. 2004, 78, 7645–7652. [Google Scholar] [CrossRef] [Green Version]
- Hioe, C.E.; Jones, G.J.; Rees, A.D.; Ratto-Kim, S.; Birx, D.; Munz, C.; Gorny, M.K.; Tuen, M.; Zolla-Pazner, S. Anti-CD4-binding domain antibodies complexed with HIV type 1 glycoprotein 120 inhibit CD4+ T cell-proliferative responses to glycoprotein 120. AIDS Res. Hum. Retrovir. 2000, 16, 893–905. [Google Scholar] [CrossRef] [PubMed]
- Rosskopf, S.; Eichholz, K.M.; Winterberg, D.; Diemer, K.J.; Lutz, S.; Munnich, I.A.; Klausz, K.; Rosner, T.; Valerius, T.; Schewe, D.M.; et al. Enhancing CDC and ADCC of CD19 Antibodies by Combining Fc Protein-Engineering with Fc Glyco-Engineering. Antibodies 2020, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Bournazos, S.; Ravetch, J.V. Fcgamma receptor pathways during active and passive immunization. Immunol. Rev. 2015, 268, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Haigwood, N.L.; Montefiori, D.C.; Sutton, W.F.; McClure, J.; Watson, A.J.; Voss, G.; Hirsch, V.M.; Richardson, B.A.; Letvin, N.L.; Hu, S.L.; et al. Passive immunotherapy in simian immunodeficiency virus-infected macaques accelerates the development of neutralizing antibodies. J. Virol. 2004, 78, 5983–5995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ng, C.T.; Jaworski, J.P.; Jayaraman, P.; Sutton, W.F.; Delio, P.; Kuller, L.; Anderson, D.; Landucci, G.; Richardson, B.A.; Burton, D.R.; et al. Passive neutralizing antibody controls SHIV viremia and enhances B cell responses in infant macaques. Nat. Med. 2010, 16, 1117–1119. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, Y.; Gautam, R.; Chun, T.W.; Sadjadpour, R.; Foulds, K.E.; Shingai, M.; Klein, F.; Gazumyan, A.; Golijanin, J.; Donaldson, M.; et al. Early antibody therapy can induce long-lasting immunity to SHIV. Nature 2017, 543, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Shah, R.R.; Hassett, K.J.; Brito, L.A. Overview of Vaccine Adjuvants: Introduction, History, and Current Status. Methods Mol. Biol. 2017, 1494, 1–13. [Google Scholar] [PubMed]
- Bastola, R.; Noh, G.; Keum, T.; Bashyal, S.; Seo, J.E.; Choi, J.; Oh, Y.; Cho, Y.; Lee, S. Vaccine adjuvants: Smart components to boost the immune system. Arch. Pharm. Res. 2017, 40, 1238–1248. [Google Scholar] [CrossRef]
- Brito, L.A.; Malyala, P.; O’Hagan, D.T. Vaccine adjuvant formulations: A pharmaceutical perspective. Semin. Immunol. 2013, 25, 130–145. [Google Scholar] [CrossRef]
- Wang, T.T.; Ravetch, J.V. Immune complexes: Not just an innocent bystander in chronic viral infection. Immunity 2015, 42, 213–215. [Google Scholar] [CrossRef] [Green Version]
- Wieland, A.; Shashidharamurthy, R.; Kamphorst, A.O.; Han, J.H.; Aubert, R.D.; Choudhury, B.P.; Stowell, S.R.; Lee, J.; Punkosdy, G.A.; Shlomchik, M.J.; et al. Antibody effector functions mediated by Fcgamma-receptors are compromised during persistent viral infection. Immunity 2015, 42, 367–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, D.H.; Elsaesser, H.; Lux, A.; Timmerman, J.M.; Morrison, S.L.; de la Torre, J.C.; Nimmerjahn, F.; Brooks, D.G. Suppression of Fcgamma-receptor-mediated antibody effector function during persistent viral infection. Immunity 2015, 42, 379–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tang, A.F.; Enyindah-Asonye, G.; Hioe, C.E. Immune Complex Vaccine Strategies to Combat HIV-1 and Other Infectious Diseases. Vaccines 2021, 9, 112. https://doi.org/10.3390/vaccines9020112
Tang AF, Enyindah-Asonye G, Hioe CE. Immune Complex Vaccine Strategies to Combat HIV-1 and Other Infectious Diseases. Vaccines. 2021; 9(2):112. https://doi.org/10.3390/vaccines9020112
Chicago/Turabian StyleTang, Alex F., Gospel Enyindah-Asonye, and Catarina E. Hioe. 2021. "Immune Complex Vaccine Strategies to Combat HIV-1 and Other Infectious Diseases" Vaccines 9, no. 2: 112. https://doi.org/10.3390/vaccines9020112