Immunization with Brucella abortus S19Δper Conferred Protection in Water Buffaloes against Virulent Challenge with B. abortus Strain S544

 , , , ,
, , , ,
Abstract
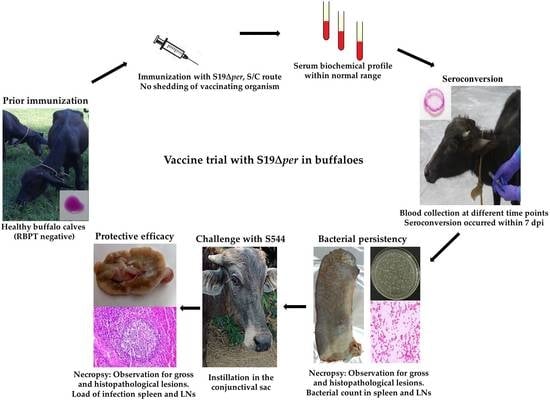
:
1. Introduction
2. Materials and Methods
2.1. Biosafety and Compliance with Animal Ethics
2.2. Bacterial Strains, Media and Growth Conditions
2.3. Safety Assay of S19∆per in Guinea Pigs
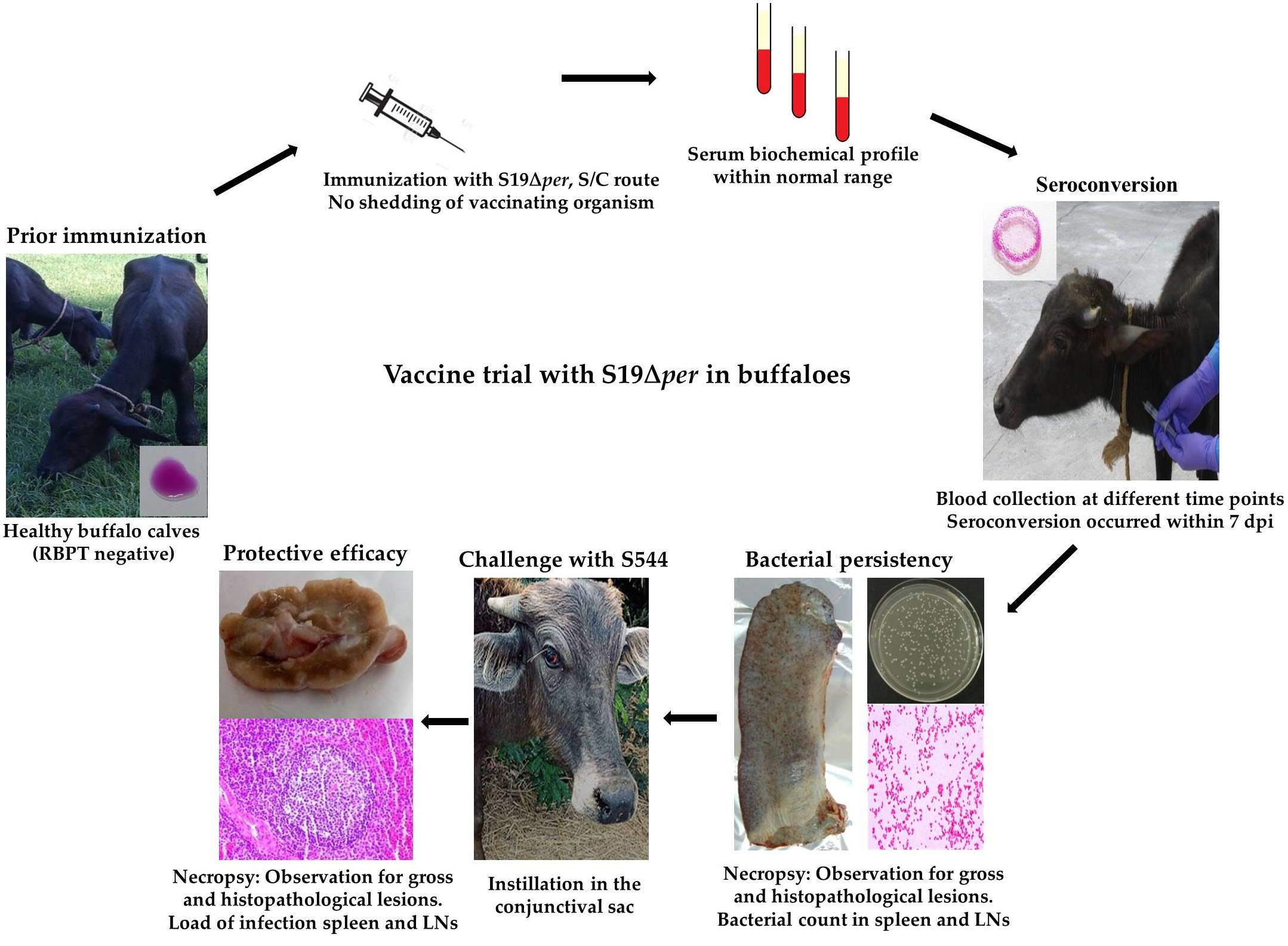
2.4. Experimental Vaccine Preparation, Immunization and Challenge Infection in Buffaloes
2.5. Indirect ELISA for the Detection of Anti-Brucella Antibodies
2.6. Serum Biochemical Profiles
2.7. Complement Sensitivity Assay
2.8. RBPT and DIVA Assessment
2.9. Bacterial Persistency
2.10. Necropsy Examination
2.11. Histopathological Studies
2.12. Statistical Analysis
3. Results
3.1. Gross and Histopathological Lesions of Vaccine Safety Assay in Guinea Pigs
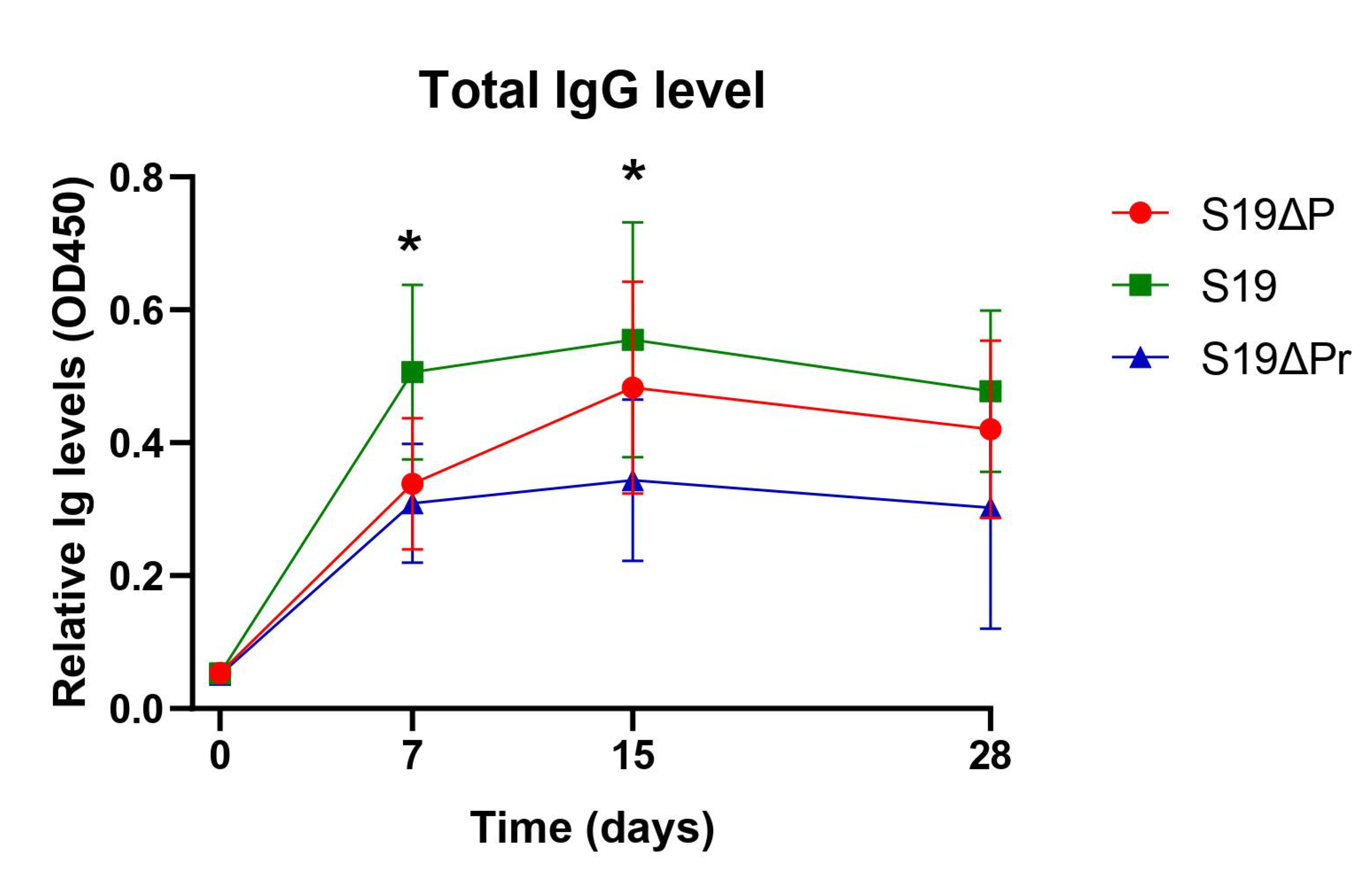
3.2. Post Immunization Humoral Immune Response and DIVA Assessment
3.3. Serum Biochemical Profiles of Vaccinated Animals
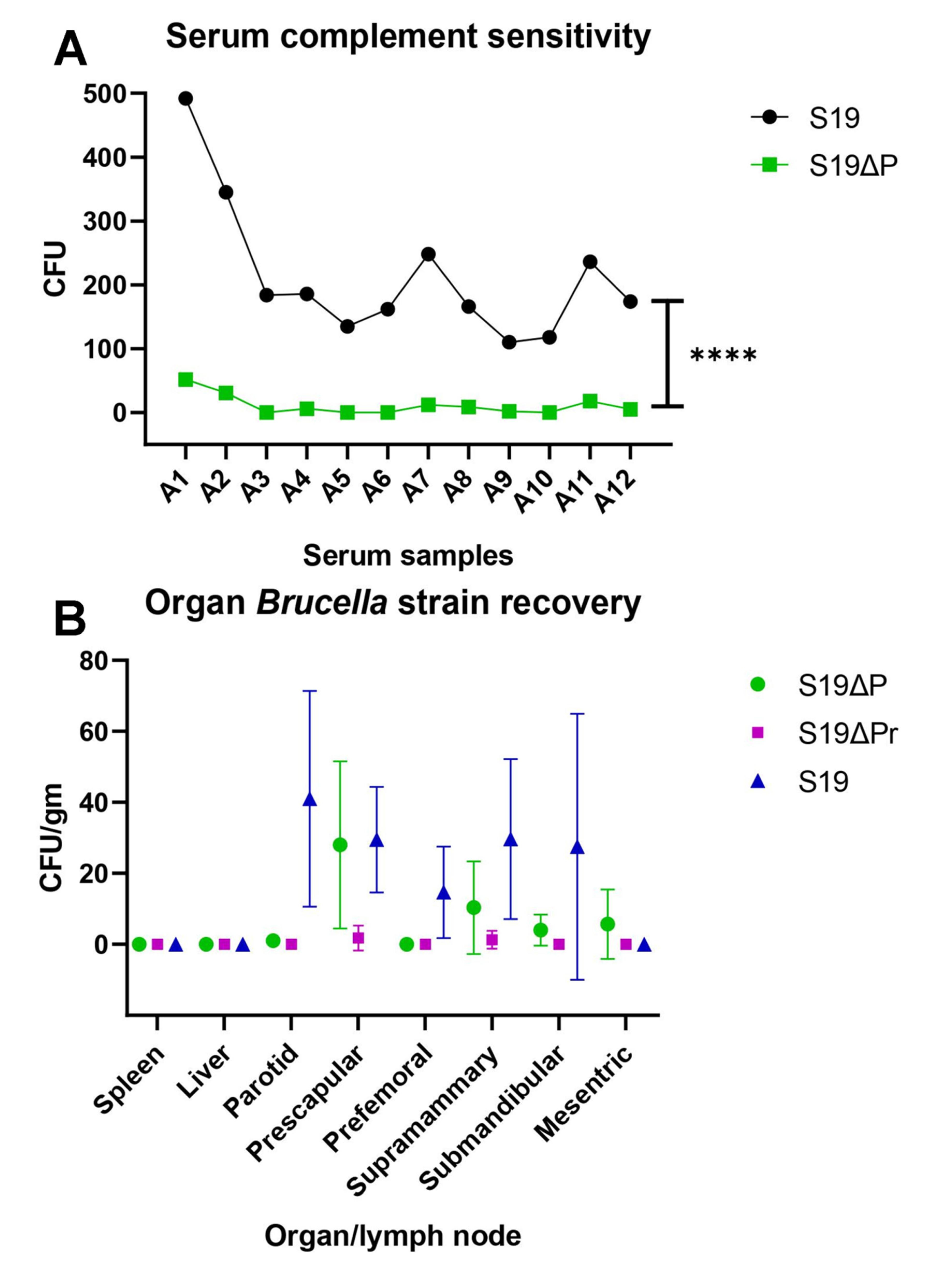
3.4. In Vitro and In Vivo Persistency of Brucella Vaccine Strains
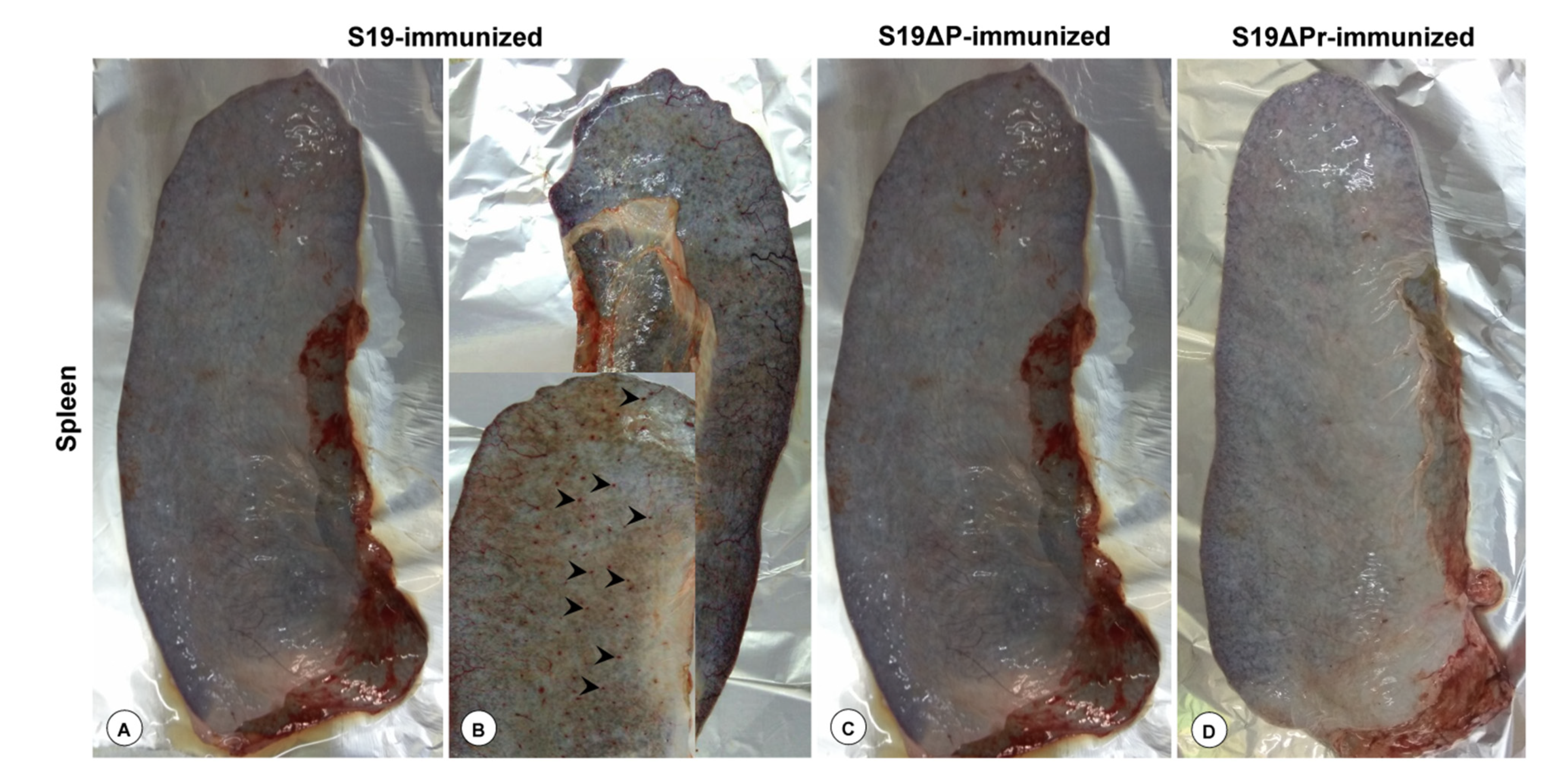
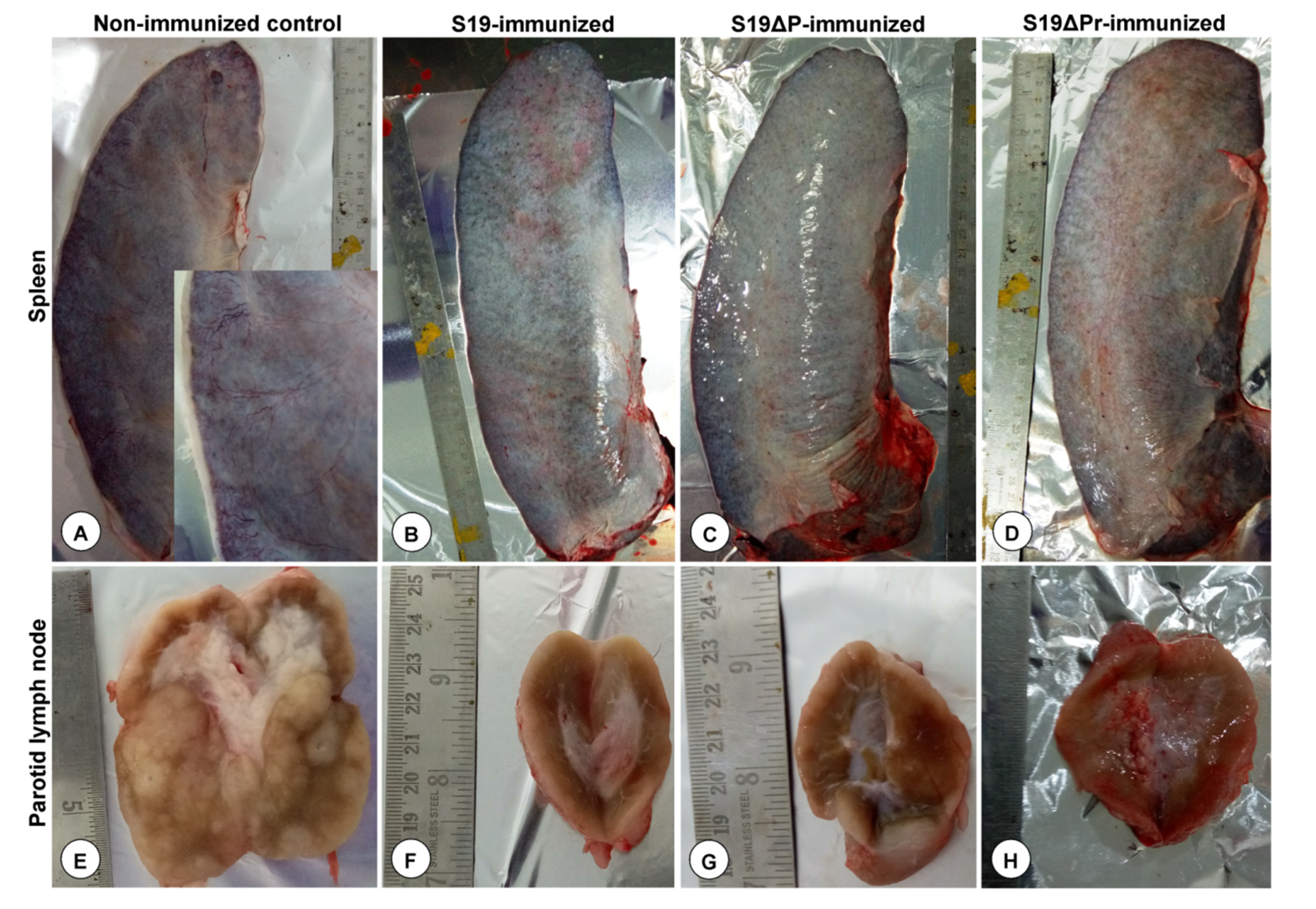
3.5. Gross and Histopathological Lesions
3.6. Assessment of Protective Efficacy of S19Δper Vaccine in Buffaloes
3.6.1. Postmortem Examination
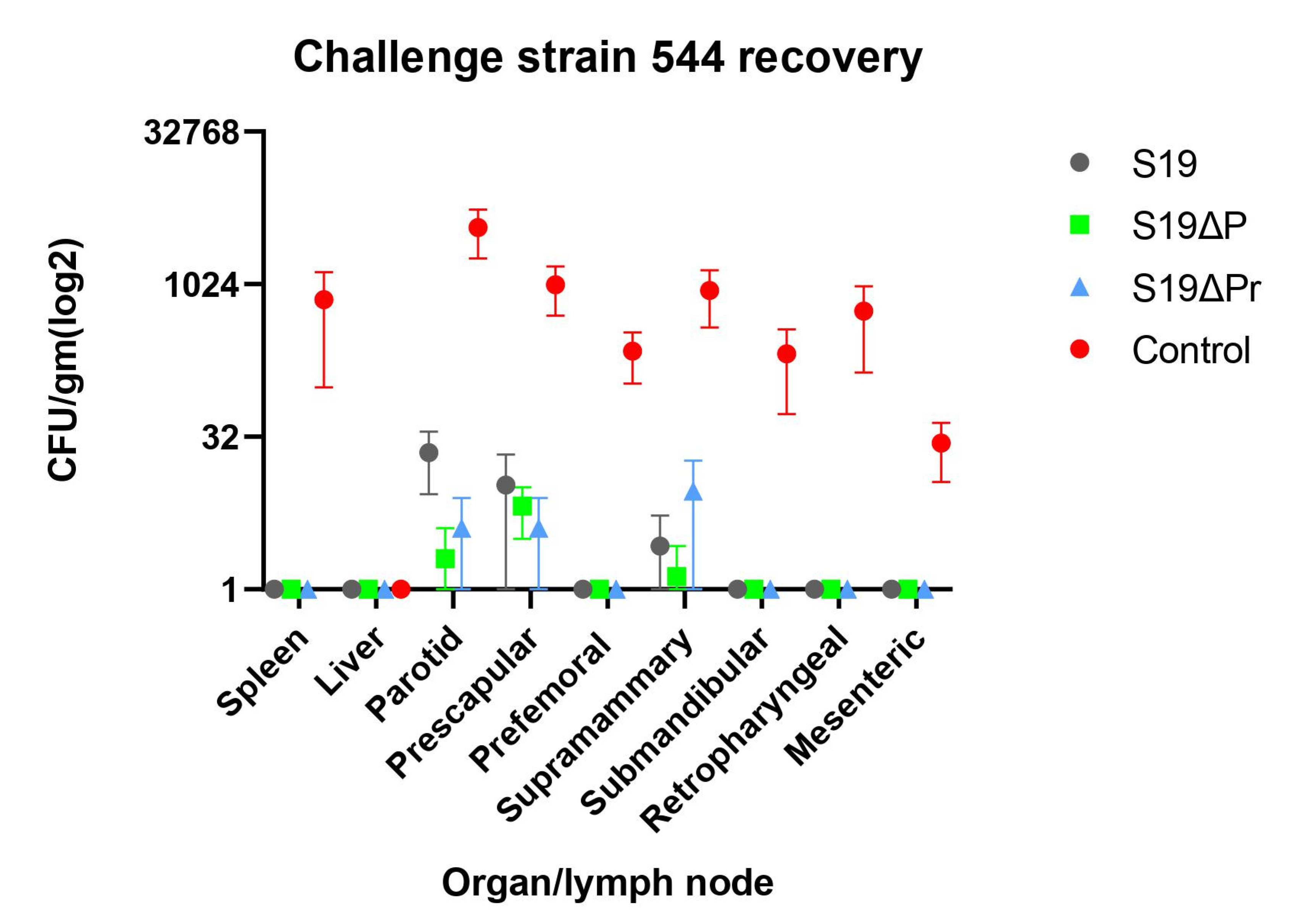
3.6.2. Clearance of Challenge Infection
3.6.3. Histopathological Lesions in Post Challenge Animals
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Khan, M.Z.; Zahoor, M. An overview of brucellosis in cattle and humans, and its serological and molecular diagnosis in control strategies. Trop. Med. Infect. Dis. 2018, 3, 65. [Google Scholar] [CrossRef] [Green Version]
- Lokamar, P.N.; Kutwah, M.A.; Atieli, H.; Gumo, S.; Ouma, C. Socio-economic impacts of brucellosis on livestock production and reproduction performance in Koibatek and Marigat regions, Baringo County, Kenya. BMC Vet. Res. 2020, 16, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhand, N.K.; Gumber, S.; Singh, B.B.; Aradhana, M.S.B.; Kumar, H.; Sharma, D.R.; Singh, J.; Sandhu, K.S. A study on the epidemiology of brucellosis in Punjab (India) using Survey Toolbox. Rev. Sci. Tech. 2005, 24, 879–885. [Google Scholar] [CrossRef] [PubMed]
- Renukaradhya, G.J.; Isloor, S.; Rajasekhar, M. Epidemiology, zoonotic aspects, vaccination and control/eradication of brucellosis in India. Vet. Microbiol. 2002, 90, 183–195. [Google Scholar] [CrossRef]
- Singh, B.B.; Dhand, N.K.; Gill, J.P. Economic losses occurring due to brucellosis in Indian livestock populations. Prev. Vet. Med. 2015, 119, 211–215. [Google Scholar] [CrossRef] [Green Version]
- Meltzer, E.; Sidi, Y.; Smolen, G.; Banai, M.; Bardenstein, S.; Schwartz, E. Sexually transmitted brucellosis in humans. Clin. Infect. Dis. 2010, 51, 12–15. [Google Scholar] [CrossRef] [PubMed]
- Wyatt, H.V. Surgeon Captain Sheldon F. Dudley and the person to person spread of brucellosis by inhalation. J. R. Nav. Med. Serv. 2010, 96, 185–187. [Google Scholar] [CrossRef] [PubMed]
- Shumilov, K.V.; Sklyarov, O.; Klimanov, A. Designing vaccines against cattle brucellosis. Vaccine 2010, 28, F31–F34. [Google Scholar] [CrossRef]
- Saminathan, M.; Rana, R.; Ramakrishnan, M.A.; Karthik, K.; Malik, Y.S.; Dhama, K. Prevalence, diagnosis, management and control of important diseases of ruminants with special reference to Indian scenario. J. Exp. Biol. Agric. Sci. 2016, 4, 338–367. [Google Scholar]
- Bjorkman, G.; Bengtson, H. Eradication of bovine brucellosis in Sweden. J. Am. Vet. Med. Assoc. 1962, 140, 1192–1195. [Google Scholar]
- van Waveren, G.M. The value of the complement fixation test for the eradication of bovine brucellosis in the Netherlands. Bull. Off. Int. Epiz. 1965, 63, 1015–1024. [Google Scholar]
- Godfroid, J.; Käsbohrer, A. Brucellosis in the European Union and Norway at the turn of the twenty-first century. Vet. Microbol. 2002, 90, 135–145. [Google Scholar] [CrossRef]
- Nicoletti, P. Vaccination. In Animal Brucellosis; Nielsen, K., Dunca, J.R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 284–299. [Google Scholar]
- Lalsiamthara, J.; Gogia, N.; Goswami, T.K.; Singh, R.K.; Chaudhuri, P. Intermediate rough Brucella abortus S19Δper mutant is DIVA enable, safe to pregnant guinea pigs and confers protection to mice. Vaccine 2015, 33, 2577–2583. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, P.; Lalsiamthara, J.; Kaur, G.; Vishnu, U.S.; Sankarasubramanian, J.; Gunasekaran, P.; Rajendhran, J. Draft genome sequence of the intermediate rough vaccine strain Brucella abortus S19Δper mutant. Genome Announc. 2015, 3, e01336-15. [Google Scholar] [CrossRef] [Green Version]
- Lalsiamthara, J.; Lee, J.H. Development and trial of vaccines against Brucella. J. Vet. Sci. 2017, 18, 281–290. [Google Scholar] [CrossRef]
- Solanki, K.S.; Qureshi, S.; Kumar, P.; Sahoo, M.; Saminathan, M.; De, U.K.; Chaudhuri, P. Patho-physiological response of LPS defective Brucella abortus S19Δper in experimentally infected mice. J. Anim. Res. 2019, 9, 79–85. [Google Scholar]
- Radwan, A.I.; Bekairi, S.I.; Al-Bokmy, A.M.; Prasad, P.V.; Mohamed, O.M.; Hussain, S.T. Successful therapeutic regimens for treating Brucella melitensis and Brucella abortus infections in cows. Rev. Off. Int. Epizoot. 1993, 12, 909–922. [Google Scholar] [CrossRef] [Green Version]
- Nicoletti, P.; Lenk, R.P.; Popescu, M.C.; Swenson, C.E. Efficacy of various treatment regimens, using liposomal streptomycin in cows with brucellosis. Am. J. Vet. Res. 1989, 50, 1004–1007. [Google Scholar]
- Milward, F.W.; Nicoletti, P.; Hoffmann, E. Effectiveness of various therapeutic regimens for bovine brucellosis. Am. J. Vet. Res. 1984, 45, 1825–1828. [Google Scholar]
- Hall, W.H. Modern chemotherapy for brucellosis in humans. Rev. Infect. Dis. 1990, 12, 1060–1099. [Google Scholar] [CrossRef]
- Gopalakrishnan, A.; Dimri, U.; Saminathan, M.; Yatoo, M.I.; Priya, G.B.; Gopinath, D.; Sujatha, V.; Ajith, Y.; Suthar, A.; Lawrence, C.; et al. Virulence factors, intracellular survivability and mechanism of evasion from host immune response by Brucella: An overview. J. Anim. Plant Sci. 2016, 26, 1542–1555. [Google Scholar]
- Moriyón, I.; Grilló, M.J.; Monreal, D.; González, D.; Marín, C.; López-Goñi, I.; Mainar-Jaime, R.C.; Moreno, E.; Blasco, J.M. Rough vaccines in animal brucellosis: Structural and genetic basis and present status. Vet. Res. 2004, 35, 1–38. [Google Scholar] [CrossRef] [Green Version]
- Kurar, E.; Splitter, G.A. Nucleic acid vaccination of Brucella abortus ribosomal L7L12 gene elicits immune response. Vaccine 1997, 15, 1851–1857. [Google Scholar] [CrossRef]
- Ugalde, J.E.; Comerci, D.J.; Leguizamón, M.S.; Ugalde, R.A. Evaluation of Brucella abortus phosphoglucomutase (pgm) mutant as a new live rough-phenotype vaccine. Infect. Immun. 2003, 71, 6264–6269. [Google Scholar] [CrossRef] [Green Version]
- Schurig, G.G.; Sriranganathan, N.; Corbel, M.J. Brucellosis vaccines: Past, present and future. Vet. Microbiol. 2002, 90, 479–496. [Google Scholar] [CrossRef]
- Zhao, Z.; Li, M.; Luo, D.; Xing, L.; Wu, S.; Duan, Y.; Yang, P.; Wang, X. Protection of mice from Brucella infection by immunization with attenuated Salmonella enterica serovar typhimurium expressing AL7/L12 and BLS fusion antigen of Brucella. Vaccine 2009, 27, 5214–5219. [Google Scholar] [CrossRef]
- Pasquevich, K.A.; Estein, S.M.; García Samartino, C.; Samartino, C.G.; Zwerdling, A.; Coria, L.M.; Barrionuevo, P.; Fossati, C.A.; Giambartolomei, G.H.; Cassataro, J. Immunization with recombinant Brucella species outer membrane protein Omp16 or Omp19 in adjuvant induces specific CD4+ and CD8+ T cells as well as systemic and oral protection against Brucella abortus infection. Infect. Immun. 2009, 77, 436–445. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pakzad, I.; Rezaee, A.; Rasaee, M.J.; Tabbaraee, B.; Delpisheh, A. Immunogencity of HSA-L7/L12 (Brucella abortus ribosomal protein) in an animal model. Iran. J. Immunol. 2009, 6, 12–21. [Google Scholar]
- Harms, J.S.; Durward, M.A.; Magnani, D.M.; Splitter, G.A. Evaluation of recombinant invasive, nonpathogenic Eschericia coli as a vaccine vector against the intracellular pathogen, Brucella. J. Immune Based Ther. Vaccines 2009, 7, 1. [Google Scholar] [CrossRef] [Green Version]
- González, M.; Andrews, E.; Folch, H.; Sáez, D.; Cabrera, A.; Salgado, P.; Oñate, A. Cloning, expression and immunogenicity of the translation initiation factor 3 homologue of Brucella abortus. Immunobiology 2009, 214, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Stevens, M.G.; Hennager, S.G.; Olsen, S.C.; Cheville, N.F. Serologic responses in diagnostic tests for brucellosis in cattle vaccinated with Brucella abortus 19 or RB51. J. Clin. Microbiol. 1994, 32, 1065–1066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stevens, M.G.; Olsen, S.C.; Cheville, N.F. Comparative analysis of immune responses in cattle vaccinated with Brucella abortus strain 19 or strain RB51. Vet. Immunol. Immunopathol. 1995, 44, 223–235. [Google Scholar] [CrossRef]
- Diptee, M.D.; Asgarali, Z.; Campbell, M.; Fosgate, G.; Adesiyun, A.A. Post-exposure serological and bacteriological responses of water buffalo (Bubalus bubalis) to Brucella abortus biovar 1 following vaccination with Brucella abortus strain RB51. Rev. Sci. Tech. 2007, 26, 669–678. [Google Scholar] [CrossRef] [PubMed]
- Guiard, J.; Paszkiewicz, E.; Sadowska, J.M.; Bundle, D.R. Design and synthesis of a universal antigen to detect brucellosis. Angew Chem. Int. Ed. 2013, 52, 7181–7185. [Google Scholar] [CrossRef]
- Ganesh, V.N.; Sadowska, J.M.; Sarkar, S.; Howells, L.; McGiven, J.; Bundle, D.R. Molecular recognition of Brucella A and M antigens dissected by synthetic oligosaccharide glycoconjugates leads to a disaccharide diagnostic for brucellosis. J. Am. Chem. Soc. 2014, 136, 16260–16269. [Google Scholar] [CrossRef]
- McGiven, J.; Howells, L.; Duncombe, L.; Stack, J.; Ganesh, N.; Guiard, J.; Bundle, D.R. Improved serodiagnosis of bovine brucellosis by novel synthetic oligosaccharide antigens based on the capping M epitope elements of Brucella O-polysaccharide. J. Clin. Microbiol. 2015, 53, 1204–1210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandal, S.S.; Duncombe, L.; Ganesh, N.V.; Sarkar, S.; Howells, L.; Hogarth, P.; Bundle, D.R.; McGiven, J. Novel solutions for vaccines and diagnostics to combat brucellosis. ACS Cent. Sci. 2017, 3, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Bundle, D.R.; McGiven, J. Brucellosis: Improved diagnostics and vaccine insights from synthetic glycans. Acc. Chem. Res. 2017, 50, 2958–2967. [Google Scholar] [CrossRef]
- Ahmed, W.; Zheng, K.; Liu, Z.F. Establishment of chronic infection: Brucella’s stealth strategy. Front. Cell. Infect. Microbiol. 2016, 6, 30. [Google Scholar] [CrossRef] [Green Version]
- Allen, C.A.; Garry, A.L.; Ficht, T.A. Transposon-derived Brucella abortus rough mutants are attenuated and exhibit reduced intracellular survival. Infect. Immun. 1998, 66, 1008–1016. [Google Scholar] [CrossRef] [Green Version]
- Godfroid, F.; Taminiau, B.; Danese, I.; Denoel, P.; Tibor, A.; Weynants, V.; Cloeckaert, A.; Godfroid, J.; Letesson, J.J. Identification of perosamine synthetase gene of Brucella melitensis 16M and involvement of lipopolysaccharide O side chain in Brucella survival in mice and in macrophages. Infect. Immun. 1998, 66, 5485–5493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McQuiston, J.R.; Vemulapalli, R.; Inzana, T.J.; Schurig, G.G.; Sriranganathan, N.; Fritzinger, D.; Hadfield, T.L.; Warren, R.A.; Lindler, L.E.; Snellings, N.; et al. Genetic characterization of a Tn-5 disrupted glycosyl transferase gene homologin Brucella abortus and its effect on lipopolysaccharide composition and virulence. Infect. Immun. 1999, 67, 3830–3835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schurig, G.G.; Roop, R.M. II.; Bagchi, T.; Boyle, S.; Buhrman, D.; Sriranganathan, N. Biological properties of RB51; a stable rough strain of Brucella abortus. Vet. Microbiol. 1991, 28, 171–188. [Google Scholar] [CrossRef]
- Winter, A.J.; Schurig, G.G.; Boyle, S.M.; Sriranganathan, N.; Bevins, J.S.; Enright, F.M.; Elzer, P.H.; Kopec, J.D. Protection of BALB/c mice against homologous and heterologous species of Brucella by rough strain vaccines derived from Brucella melitensis and Brucella suis biovar 4. Am. J. Vet. Res. 1996, 57, 677–683. [Google Scholar]
- Eisenschenk, F.C.; Houle, J.J.; Hoffmann, E.M. Serum sensitivity of field isolates and laboratory strains of Brucella abortus. Am. J. Vet. Res. 1995, 56, 1592–1598. [Google Scholar]
- Gheibi, A.; Khanahmad, H.; Kashfi, K.; Sarmadi, M.; Khorramizadeh, M.R. Development of new generation of vaccines for Brucella abortus. Heliyon 2018, 4, e01079. [Google Scholar] [CrossRef] [Green Version]
- Monreal, D.; Grillo, M.J.; González, D.; Marin, C.M.; DeMiguel, M.J.; Lopez-Goñi, I.; Blasco, J.M.; Cloeckaert, A.; Moriyón, I. Characterization of Brucella abortus O-polysaccharide and core lipopolysaccharide mutants and demonstration that a complete core is required for rough vaccines to be efficient against Brucella abortus and Brucella ovis in the mouse model. Infect. Immun. 2003, 71, 3261–3271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheville, N.F.; Jensen, A.E.; Halling, S.M.; Tatum, F.M.; Morfitt, D.C.; Hennager, S.G.; Frerichs, W.M.; Schurig, G. Bacterial survival, lymph node changes and immunologic responses of cattle vaccinated with standard and mutant strains of Brucella abortus. Am. J. Vet. Res. 1992, 53, 1881–1888. [Google Scholar]
- Adesiyun, A.A.; Fosgate, G.T.; Persad, A.; Campbell, M.; Seebaransingh, R.; Stewart-Johnson, A. Comparative study on responses of cattle and water buffalo (Bubalus bubalis) to experimental inoculation of Brucella abortus biovar 1 by the intraconjunctival route—A preliminary report. Trop. Anim. Health Prod. 2010, 42, 1685–1694. [Google Scholar] [CrossRef] [PubMed]
- Olsen, S.C.; Cheville, N.F.; Kunkle, R.A.; Palmer, M.V.; Jensen, A.E. Bacterial survival, lymph node pathology, and serological responses of bison (Bison bison) vaccinated with Brucella abortus strain RB51 or strain 19. J. Wildl. Dis. 1997, 33, 146–151. [Google Scholar] [CrossRef] [Green Version]
- Caporale, V.; Bonfini, B.; DiGiannatale, E.; DiProvvido, A.; Forcella, S.; Giovannini, A.; Tittarelli, M.; Scacchia, M. Efficacy of Brucella abortus vaccine strain RB51 compared to the reference vaccine Brucella abortus strain 19 in water buffalo. Vet. Ital. 2010, 46, 13–19. [Google Scholar] [PubMed]
- Meyer, M.E.; Nelson, C.J. Persistence of Brucella abortus, strain 19 infection in immunized cattle. Proc. Annu. Meet US Anim. Health Assoc. 1969, 73, 159–165. [Google Scholar]
- Thomas, E.L.; Bracewell, C.D.; Corbel, M.J. Characterisation of Brucella abortus strain 19 cultures isolated from vaccinated cattle. Vet. Rec. 1981, 108, 90–93. [Google Scholar] [CrossRef]
- Davies, G.; Cocksi, E.; Hebert, N. Brucella abortus (strain 19) vaccine: (a) Determination of the minimum protective dose in cattle; (b) The effect of vaccinating calves previously inoculated with anti-Brucella abortus serum. J. Biol. Stand. 1980, 8, 171–178. [Google Scholar] [CrossRef]
- Plommet, M.; Fensterbank, R.; Souriau, A. Vaccination against bovine brucellosis with a low dose of strain 19 administered by the conjunctival route. III.—Serological response and immunity in the pregnant cow. Ann. Rech. Vet. 1976, 7, 9–23. [Google Scholar]
- Velasquez, L.N.; Delpino, M.V.; Ibanez, A.E.; Coria, L.M.; Miraglia, M.C.; Scian, R.; Cassataro, J.; Giambartolomei, G.H.; Barrionuevo, P. Brucella abortus induces apoptosis of human T lymphocytes. Microbes Infect. 2012, 14, 639–650. [Google Scholar] [CrossRef]
- Fosgate, G.T.; Adesiyun, A.A.; Hird, D.W.; Johnson, W.O.; Hietala, S.K.; Schurig, G.G.; Ryan, J.; Diptee, M.D. Evaluation of brucellosis RB51 vaccine for domestic water buffalo (Bubalus bubalis) in Trinidad. Prev. Vet. Med. 2003, 15, 211–225. [Google Scholar] [CrossRef]
- Ramnanan, A.; Campbell, M.; Asgarail, Z.; Diptee, M.; Adesiyun, A.A. Experimental study on Brucella abortus strain RB51 vaccinated water buffalo (Bubalus bubalis) challenged with virulent B. abortus strain during pregnancy. J. Buffalo Sci. 2018, 7, 17–29. [Google Scholar] [CrossRef]
- Zriba, S.; Garcia-Gonzalez, D.G.; Khalaf, O.H.; Wheeler, L.; Chaki, S.P.; Rice-Ficht, A.; Ficht, T.A.; Arenas-Gamboa, A.M. Vaccine safety studies of Brucella abortus S19 and S19∆vjbR in pregnant swine. Vaccine 2019, 3, 100041. [Google Scholar]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AST (U/L) | ALT (U/L) | AP (U/L) | Total Protein (g/dL) | Albumin (g/dL) | Direct Bilirubin (mg/dL) | Total Bilirubin (mg/dL) | CPK (U/L) | Creatinine (mg/dL) | |
|---|---|---|---|---|---|---|---|---|---|
| S19 | 80.25 ± 4.10 | 27.00 ± 1.83 | 416.39 ± 48.38 | 5.46 ± 0.09 | 1.75 ± 0.15 | 0.31 ± 0.05 | 0.38 ± 0.03 | 143.46 ± 14.69 | 1.60 ± 0.17 |
| S19ΔP | 79.38 ± 4.10 | 29.00 ± 1.83 | 426.35 ± 48.38 | 5.22 ± 0.09 | 1.88 ± 0.15 | 0.45 ± 0.05 | 0.24 ± 0.03 | 134.62 ± 14.69 | 1.43 ± 0.17 |
| S19ΔPr | 81.25 ± 4.73 | 24.33 ± 0.88 | 405.33 ± 36.60 | 5.73 ± 0.53 | 1.87 ± 0.15 | 0.33 ± 0.12 | 0.43 ± 0.05 | 153.22 ± 7.58 | 1.34 ± 0.23 |
| Vaccine Group | Animal No. | Days Post Immunization (dpi) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| 0 | 7 | 14 | 28 | 42 | 70 | 112 | 300 | ||
| S19 | 1 | − | +++ | +++ | +++ | +++ * | X | X | X |
| 2 | − | ++ | +++ | +++ | ++ * | X | X | X | |
| 3 | − | +++ | +++ | +++ | +++ * | X | X | X | |
| 4 | − | ++ | +++ | +++ | +++ * | X | X | X | |
| 5 | − | +++ | +++ | +++ | +++ | ++ | + | − | |
| 6 | − | +++ | +++ | +++ | +++ | ++ | + | − | |
| 7 | − | +++ | +++ | ++ | ++ | + | − | − | |
| S19Δper | 1 | − | − | + | − | − * | X | X | X |
| 2 | − | ++ | ++ | ++ | ++ * | X | X | X | |
| 3 | − | ++ | ++ | + | + * | X | X | X | |
| 4 | − | + | + | + | − * | X | X | X | |
| 5 | − | + | ++ | + | + | − | − | − | |
| 6 | − | + | + | + | + | − | − | − | |
| 7 | − | ++ | ++ | +++ | +++ | ++ | − | − | |
| S19Δper reduced dose | 1 | − | − | − | − | − * | X | X | X |
| 2 | − | − | − | − | − * | X | X | X | |
| 3 | − | − | + | + | − * | X | X | X | |
| 4 | − | + | + | + | + * | X | X | X | |
| 5 | − | − | − | − | − | − | − | − | |
| 6 | − | + | +++ | +++ | ++ | − | − | − | |
| 7 | − | ++ | ++ | ++ | ++ | − | − | − | |
| PBS control | 1 | − | ND | ND | − | ND | ND | ND | − |
| 2 | − | ND | ND | − | ND | ND | ND | − | |
| 3 | − | ND | ND | − | ND | ND | ND | − | |
 |  |  |  | ||||||
| +++ | ++ | + | − | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chaudhuri, P.; Saminathan, M.; Ali, S.A.; Kaur, G.; Singh, S.V.; Lalsiamthara, J.; Goswami, T.K.; Singh, A.K.; Singh, S.K.; Malik, P.; et al. Immunization with Brucella abortus S19Δper Conferred Protection in Water Buffaloes against Virulent Challenge with B. abortus Strain S544. Vaccines 2021, 9, 1423. https://doi.org/10.3390/vaccines9121423
Chaudhuri P, Saminathan M, Ali SA, Kaur G, Singh SV, Lalsiamthara J, Goswami TK, Singh AK, Singh SK, Malik P, et al. Immunization with Brucella abortus S19Δper Conferred Protection in Water Buffaloes against Virulent Challenge with B. abortus Strain S544. Vaccines. 2021; 9(12):1423. https://doi.org/10.3390/vaccines9121423
Chicago/Turabian StyleChaudhuri, Pallab, Mani Saminathan, Syed Atif Ali, Gurpreet Kaur, Shiv Varan Singh, Jonathan Lalsiamthara, Tapas K. Goswami, Ashwini K. Singh, Sandeep K. Singh, Praveen Malik, and et al. 2021. "Immunization with Brucella abortus S19Δper Conferred Protection in Water Buffaloes against Virulent Challenge with B. abortus Strain S544" Vaccines 9, no. 12: 1423. https://doi.org/10.3390/vaccines9121423