
A Listeria monocytogenes-Based Vaccine Formulation Reduces Vertical Transmission and Leads to Enhanced Pup Survival in a Pregnant Neosporosis Mouse Model

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Host Cells, Parasite and Primers
2.2. Generation and Quality Control of the Mutant Vaccine Strain Lm3Dx_NcSAG1
2.3. Ethics Statement
2.4. Efficacy Evaluation of the Mutant Vaccine Strain Lm3Dx_NcSAG1 in Non-Pregnant and Pregnant Mice Infected with NcSpain-7 Tachyzoites
2.5. Analysis of Cerebral Parasite Burden by Real-Time (RT) qPCR and Histology
2.6. Analysis of Humoral Immune Responses by Enzyme-Linked Immunosorbent Assay (ELISA)
2.7. Determination of Cytokine Levels in Splenocyte Culture Supernatants by Multiplex Immunoassay
2.8. Statistical Analysis
3. Results
3.1. Safety and Efficacy of the Mutant Listeria Strain Lm3Dx_NcSAG1 in the Pregnant Neosporosis Mouse Model
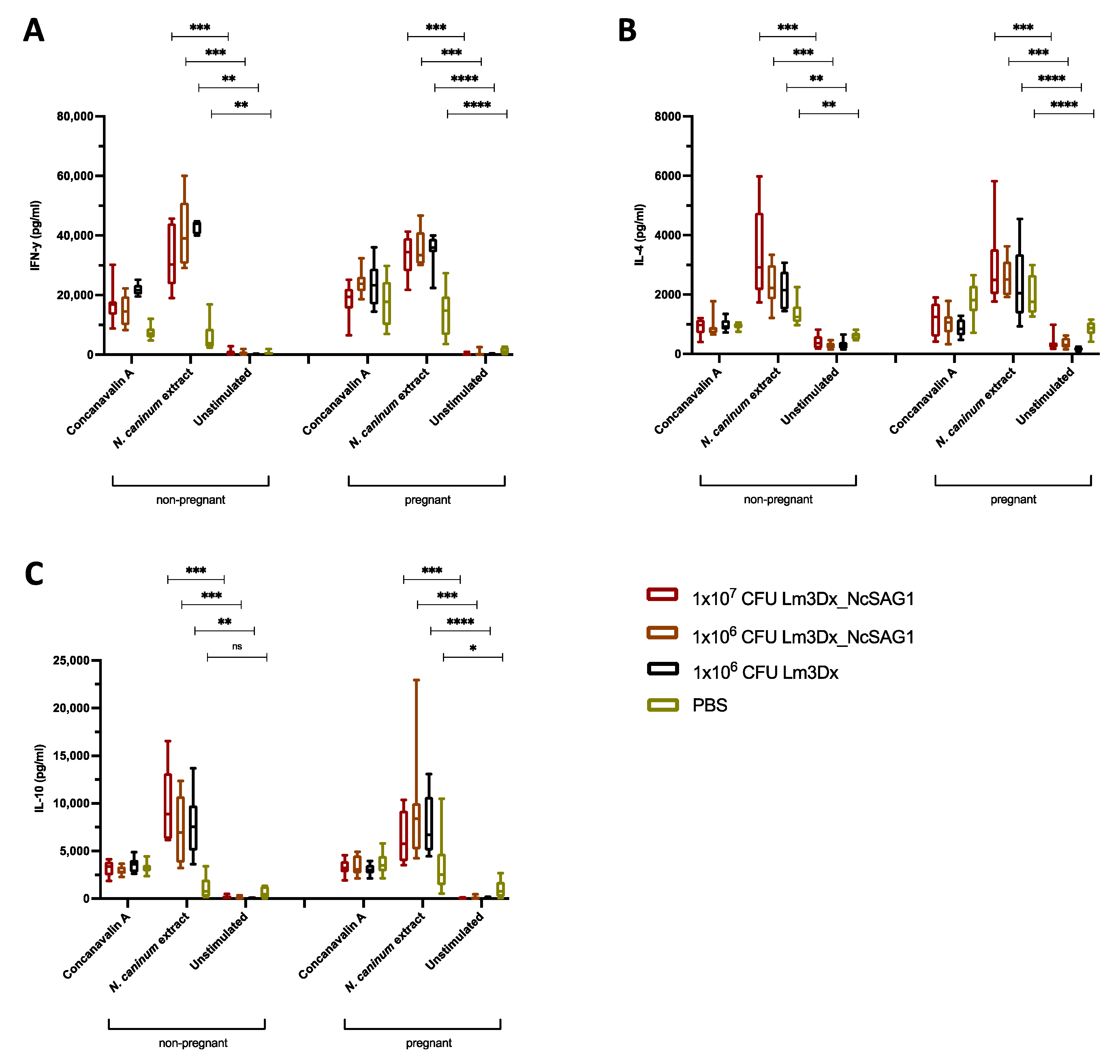
3.2. Humoral and Cellular Immune Responses
3.3. Histology
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Hemphill, A.; Calero-Bernal, R.; Schares, G. Neosporosis in Animals; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar] [CrossRef]
- Aguado-Martínez, A.; Basto, A.P.; Leitão, A.; Hemphill, A. Neospora caninum in non-pregnant and pregnant mouse models: Cross-talk between infection and immunity. Int. J. Parasitol. 2017, 47, 723–735. [Google Scholar] [CrossRef]
- Reichel, M.P.; Alejandra Ayanegui-Alcérreca, M.; Gondim, L.F.P.; Ellis, J.T. What is the global economic impact of Neospora caninum in cattle—The billion dollar question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [Green Version]
- Monney, T.; Hemphill, A. Vaccines against neosporosis: What can we learn from the past studies? Exp. Parasitol. 2014, 140, 52–70. [Google Scholar] [CrossRef]
- Eperon, S.; Brönnimann, K.; Hemphill, A.; Gottstein, B. Susceptibility of B-cell deficient C57BL/6 (μMT) mice to Neospora caninum infection. Parasite Immunol. 1999, 21, 225–236. [Google Scholar] [CrossRef]
- Hemphill, A.; Vonlaufen, N.; Naguleswaran, A. Cellular and immunological basis of the host-parasite relationship during infection with Neospora caninum. Parasitology 2006, 133, 261–278. [Google Scholar] [CrossRef]
- Williams, D.J.L.; Guy, C.S.; Smith, R.F.; Ellis, J.; Björkman, C.; Reichel, M.P.; Trees, A.J. Immunization of cattle with live tachyzoites of Neospora caninum confers protection against fetal death. Infect. Immun. 2007, 75, 1343–1348. [Google Scholar] [CrossRef] [Green Version]
- Monney, T.; Debache, K.; Hemphill, A. Vaccines against a major cause of abortion in cattle, Neospora caninum infection. Animals 2011, 1, 306–325. [Google Scholar] [CrossRef] [Green Version]
- Aguado-Martínez, A.; Basto, A.P.; Tanaka, S.; Ryser, L.T.; Nunes, T.P.; Ortega-Mora, L.M.; Arranz-Solís, D.; Leitão, A.; Hemphill, A. Immunization with a cocktail of antigens fused with OprI reduces Neospora caninum vertical transmission and postnatal mortality in mice. Vaccine 2018, 37, 473–483. [Google Scholar] [CrossRef] [Green Version]
- Monney, T.; Debache, K.; Grandgirard, D.; Leib, S.L.; Hemphill, A. Vaccination with the recombinant chimeric antigen recNcMIC3-1-R induces a non-protective Th2-type immune response in the pregnant mouse model for N. caninum infection. Vaccine 2012, 30, 6588–6594. [Google Scholar] [CrossRef]
- Chávez-Arroyo, A.; Portnoy, D.A. Why is Listeria monocytogenes such a potent inducer of CD8+ T-cells? Cell. Microbiol. 2020, 22, e13175. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.; Lian, K.; Zhao, D.; Tao, C.; Chen, X.; Tan, W.; Wang, X.; Xu, Z.; Hu, M.; Rao, Y.; et al. A promising listeria-vectored vaccine induces Th1-type immune responses and confers protection against tuberculosis. Front. Cell. Infect. Microbiol. 2017, 7, 407. [Google Scholar] [CrossRef] [Green Version]
- D’Orazio, S.E.F. Innate and Adaptive Immune Responses during Listeria monocytogenes Infection. Microbiol. Spectr. 2019, 7, 1–40. [Google Scholar] [CrossRef]
- Lecuit, M.; Ohayon, H.; Braun, L.; Mengaud, J.; Cossart, P. Internalin of Listeria monocytogenes with an intact leucine-rich repeat region is sufficient to promote internalization. Infect. Immunol. 1997, 65, 5309–5319. [Google Scholar] [CrossRef] [Green Version]
- Radoshevich, L.; Cossart, P. Listeria monocytogenes: Towards a complete picture of its physiology and pathogenesis. Nat. Rev. Microbiol. 2018, 16, 32–46. [Google Scholar] [CrossRef]
- Flickinger, J.C.; Rodeck, U.; Snook, A.E. Listeria monocytogenes as a vector for cancer immunotherapy: Current understanding and progress. Vaccines 2018, 6, 48. [Google Scholar] [CrossRef] [Green Version]
- Pownall, W.R.; Imhof, D.; Trigo, N.F.; Ganal-Vonarburg, S.C.; Plattet, P.; Monney, C.; Forterre, F.; Hemphill, A.; Oevermann, A. Safety of a Novel Listeria monocytogenes-based Vaccine Vector Expressing NcSAG1 (Neospora caninum Surface Antigen 1). Front. Cell. Infect. Microbiol. 2021, 11, 726. [Google Scholar] [CrossRef]
- Aguado-Martínez, A.; Basto, A.P.; Müller, J.; Balmer, V.; Manser, V.; Leitão, A.; Hemphill, A. N-terminal fusion of a toll-like receptor 2-ligand to a Neospora caninum chimeric antigen efficiently modifies the properties of the specific immune response. Parasitology 2016, 143, 606–616. [Google Scholar] [CrossRef]
- Whitten, M.K. Effect of exteroceptive factors on the œstrous cycle of mice. Nature 1957, 180, 1436. [Google Scholar] [CrossRef]
- Arranz-Solís, D.; Aguado-Martínez, A.; Müller, J.; Regidor-Cerrillo, J.; Ortega-Mora, L.M.; Hemphill, A. Dose-dependent effects of experimental infection with the virulent Neospora caninum Nc-Spain7 isolate in a pregnant mouse model. Vet. Parasitol. 2015, 211, 133–140. [Google Scholar] [CrossRef]
- Müller, N.; Zimmermann, V.; Hentrich, B.; Gottstein, B. Diagnosis of Neospora caninum and Toxoplasma gondii infection by PCR and DNA hybridization immunoassay. J. Clin. Microbiol. 1996, 34, 2850–2852. [Google Scholar] [CrossRef] [Green Version]
- Müller, N.; Vonlaufen, N.; Gianinazzi, C.; Leib, S.L.; Hemphill, A. Application of real-time fluorescent PCR for quantitative assessment of Neospora caninum infections in organotypic slice cultures of rat central nervous system tissue. J. Clin. Microbiol. 2002, 40, 252–255. [Google Scholar] [CrossRef] [Green Version]
- Longo, M.C.; Berninger, M.S.; Hartley, J.L. Use of uracil DNA glycosylase to control carry-over contamination in polymerase chain reactions. Gene 1990, 93, 125–128. [Google Scholar] [CrossRef]
- Debache, K.; Guionaud, C.; Alaeddine, F.; Mevissen, M.; Hemphill, A. Vaccination of mice with recombinant NcROP2 antigen reduces mortality and cerebral infection in mice infected with Neospora caninum tachyzoites. Int. J. Parasitol. 2008, 38, 1455–1463. [Google Scholar] [CrossRef]
- Debache, K.; Alaeddine, F.; Guionaud, C.; Monney, T.; Müller, J.; Strohbusch, M.; Leib, S.L.; Grandgirard, D.; Hemphill, A. Vaccination with recombinant NcROP2 combined with recombinant NcMIC1 and NcMIC3 reduces cerebral infection and vertical transmission in mice experimentally infected with Neospora caninum tachyzoites. Int. J. Parasitol. 2009, 39, 1373–1384. [Google Scholar] [CrossRef]
- Dunbar, S.A. Applications of Luminex® xMAPTM technology for rapid, high-throughput multiplexed nucleic acid detection. Clin. Chim. Acta 2006, 363, 71–82. [Google Scholar] [CrossRef]
- Rojo-Montejo, S.; Collantes-Fernández, E.; Pérez-Zaballos, F.; Rodríguez-Marcos, S.; Blanco-Murcia, J.; Rodríguez-Bertos, A.; Prenafeta, A.; Ortega-Mora, L.M. Effect of vaccination of cattle with the low virulence Nc-Spain 1H isolate of Neospora caninum against a heterologous challenge in early and mid-gestation. Vet. Res. 2013, 44, 106. [Google Scholar] [CrossRef] [Green Version]
- Rojo-Montejo, S.; Collantes-Fernández, E.; López-Pérez, I.; Risco-Castillo, V.; Prenafeta, A.; Ortega-Mora, L.M. Evaluation of the protection conferred by a naturally attenuated Neospora caninum isolate against congenital and cerebral neosporosis in mice. Vet. Res. 2012, 43, 62. [Google Scholar] [CrossRef] [Green Version]
- Weber, F.H.; Jackson, J.A.; Sobecki, B.; Choromanski, L.; Olsen, M.; Meinert, T.; Frank, R.; Reichel, M.P.; Ellis, J.T. On the efficacy and safety of vaccination with live tachyzoites of Neospora caninum for prevention of neospora-associated fetal loss in cattle. Clin. Vaccine Immunol. 2013, 20, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Penarete-Vargas, D.M.; Mévélec, M.N.; Dion, S.; Sèche, E.; Dimier-Poisson, I.; Fandeur, T. Protection against Lethal Neospora caninum Infection in Mice Induced by Heterologous Vaccination with a mic1 mic3 Knockout Toxoplasma gondii Strain. Infect. Immun. 2010, 78, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Marugán-Hernández, V.; Ortega-Mora, L.M.; Aguado-Martínez, A.; Jiménez-Ruíz, E.; Alvarez-García, G. Transgenic Neospora caninum strains constitutively expressing the bradyzoite NcSAG4 protein proved to be safe and conferred significant levels of protection against vertical transmission when used as live vaccines in mice. Vaccine 2011, 29, 7867–7874. [Google Scholar] [CrossRef]
- Hemphill, A.; Aguado-Martínez, A.; Müller, J. Approaches for the vaccination and treatment of Neospora caninum infections in mice and ruminant models. Parasitology 2016, 143, 245–259. [Google Scholar] [CrossRef]
- Horcajo, P.; Regidor-Cerrillo, J.; Aguado-Martínez, A.; Hemphill, A.; Ortega-Mora, L.M. Vaccines for bovine neosporosis: Current status and key aspects for development. Parasite Immunol. 2016, 38, 709–723. [Google Scholar] [CrossRef] [Green Version]
- Reichel, M.P.; Moore, D.P.; Hemphill, A.; Ortega-Mora, L.M.; Dubey, J.P.; Ellis, J.T. A live vaccine against Neospora caninum abortions in cattle. Vaccine 2015, 33, 1299–1301. [Google Scholar] [CrossRef]
- Marugan-Hernandez, V. Neospora caninum and Bovine Neosporosis: Current Vaccine Research. J. Comp. Pathol. 2017, 157, 193–200. [Google Scholar] [CrossRef] [Green Version]
- Hemphill, A.; Debache, K.; Monney, T.; Schorer, M.; Guionaud, C.; Alaeddine, F.; Müller, N.; Müller, J. Proteins mediating the Neospora caninum-host cell interaction as targets for vaccination. Front. Biosci. 2013, 5, 23–36. [Google Scholar] [CrossRef]
- Howe, D.K.; Crawford, A.C.; Lindsay, D.; Sibley, L.D. The p29 and p35 Immunodominant Antigens of Neospora caninum Tachyzoites Are Homologous to the Family of Surface Antigens of Toxoplasma gondii. Infect. Immun. 1998, 66, 5322–5328. [Google Scholar] [CrossRef] [Green Version]
- Sonda, S.; Fuchs, N.; Connolly, B.; Fernandez, P.; Gottstein, B.; Hemphill, A. The major 36 kDa Neospora caninum tachyzoite surface protein is closely related to the major Toxoplasma gondii surface antigen. Mol. Biochem. Parasitol. 1998, 97, 97–108. [Google Scholar] [CrossRef]
- Cannas, A.; Naguleswaran, A.; Müller, N.; Eperon, S.; Gottstein, B.; Hemphill, A. Vaccination of mice against experimental Neospora caninum infection using NcSAG1- and NcSRS2-based recombinant antigens and DNA vaccines. Parasitology 2003, 126, 303–312. [Google Scholar] [CrossRef]
- Xu, J.; Hiramatsu, R.; Suhaimi, H.; Kato, T.; Fujimoto, A.; Tokiwa, T.; Ike, K.; Park, E.Y. Neospora caninum antigens displaying virus-like particles as a bivalent vaccine candidate against neosporosis. Vaccine 2019, 37, 6426–6434. [Google Scholar] [CrossRef]
- Bengoa-Luoni, S.A.; Corigliano, M.G.; Sánchez-López, E.; Albarracín, R.M.; Legarralde, A.; Ganuza, A.; Clemente, M.; Sander, V.A. The potential of a DIVA-like recombinant vaccine composed by rNcSAG1 and rAtHsp81.2 against vertical transmission in a mouse model of congenital neosporosis. Acta Trop. 2019, 198, 105094. [Google Scholar] [CrossRef]
- Cho, J.H.; Chung, W.S.; Song, K.J.; Na, B.K.; Kang, S.W.; Song, C.Y.; Kim, T.S. Protective efficacy of vaccination with Neospora caninum multiple recombinant antigens against experimental Neospora caninum infection. Korean J. Parasitol. 2005, 43, 19–25. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccination | Challenge | Seropositive for N. caninum | N. caninum Brain Positive Non-Pregnant Mice | N. caninum Brain Positive Dams | Number of Pups per Dam | Neonatal Mortality a | Postnatal Mortality b | N. caninum Brain Positive Pups |
|---|---|---|---|---|---|---|---|---|
| Lm3Dx_NcSAG1 1 × 107 CFU | 1 × 105 NcSp7 tachyzoites | 16/16 | 5/8 | 7/8 | 43/8 ( 5.4) | 0/43 (0%) | 14/43 (33%) 2 | 17/43 (39%) 5 |
| Lm3Dx_NcSAG1 1 × 106 CFU | 1 × 105 NcSp7 tachyzoites | 16/16 | 1/8 1 | 5/8 | 48/8 ( 6) | 0/48 (0%) | 26/48 (54%) 3 | 37/48 (77%) 6 |
| Lm3Dx_NcSAG1 1 × 105 CFU | 1 × 105 NcSp7 tachyzoites | 15/15 + | 6/11 + | 2/4 | 19/4 ( 4.8) | 3/19 (16%) | 11/16 (69%) 4 | 14/16 (88%) |
| Lm3Dx 1 × 106 CFU | 1 × 105 NcSp7 tachyzoites | 16/16 | 5/6 | 10/10 | 65/10 ( 6.5) | 3/65 (5%) | 59/62 (95%) | 60/62 (97%) |
| PBS | 1 × 105 dermal fibroblasts | 0/16 | 0/6 | 0/10 | 57/10 ( 5.7) | 1/57 (2%) | 0/56 (0%) | 0/56 (0%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Imhof, D.; Pownall, W.R.; Monney, C.; Oevermann, A.; Hemphill, A. A Listeria monocytogenes-Based Vaccine Formulation Reduces Vertical Transmission and Leads to Enhanced Pup Survival in a Pregnant Neosporosis Mouse Model. Vaccines 2021, 9, 1400. https://doi.org/10.3390/vaccines9121400
Imhof D, Pownall WR, Monney C, Oevermann A, Hemphill A. A Listeria monocytogenes-Based Vaccine Formulation Reduces Vertical Transmission and Leads to Enhanced Pup Survival in a Pregnant Neosporosis Mouse Model. Vaccines. 2021; 9(12):1400. https://doi.org/10.3390/vaccines9121400
Chicago/Turabian StyleImhof, Dennis, William Robert Pownall, Camille Monney, Anna Oevermann, and Andrew Hemphill. 2021. "A Listeria monocytogenes-Based Vaccine Formulation Reduces Vertical Transmission and Leads to Enhanced Pup Survival in a Pregnant Neosporosis Mouse Model" Vaccines 9, no. 12: 1400. https://doi.org/10.3390/vaccines9121400