
Enhancement of the Local CD8+ T-Cellular Immune Response to Mycobacterium tuberculosis in BCG-Primed Mice after Intranasal Administration of Influenza Vector Vaccine Carrying TB10.4 and HspX Antigens

 , , , ,
, , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Vaccine Strain
2.3. Experimental Design
2.4. Cell Preparation
2.5. Flow Cytometry Analysis
2.6. Protection Study in Mice
2.7. Mycobacterial Load
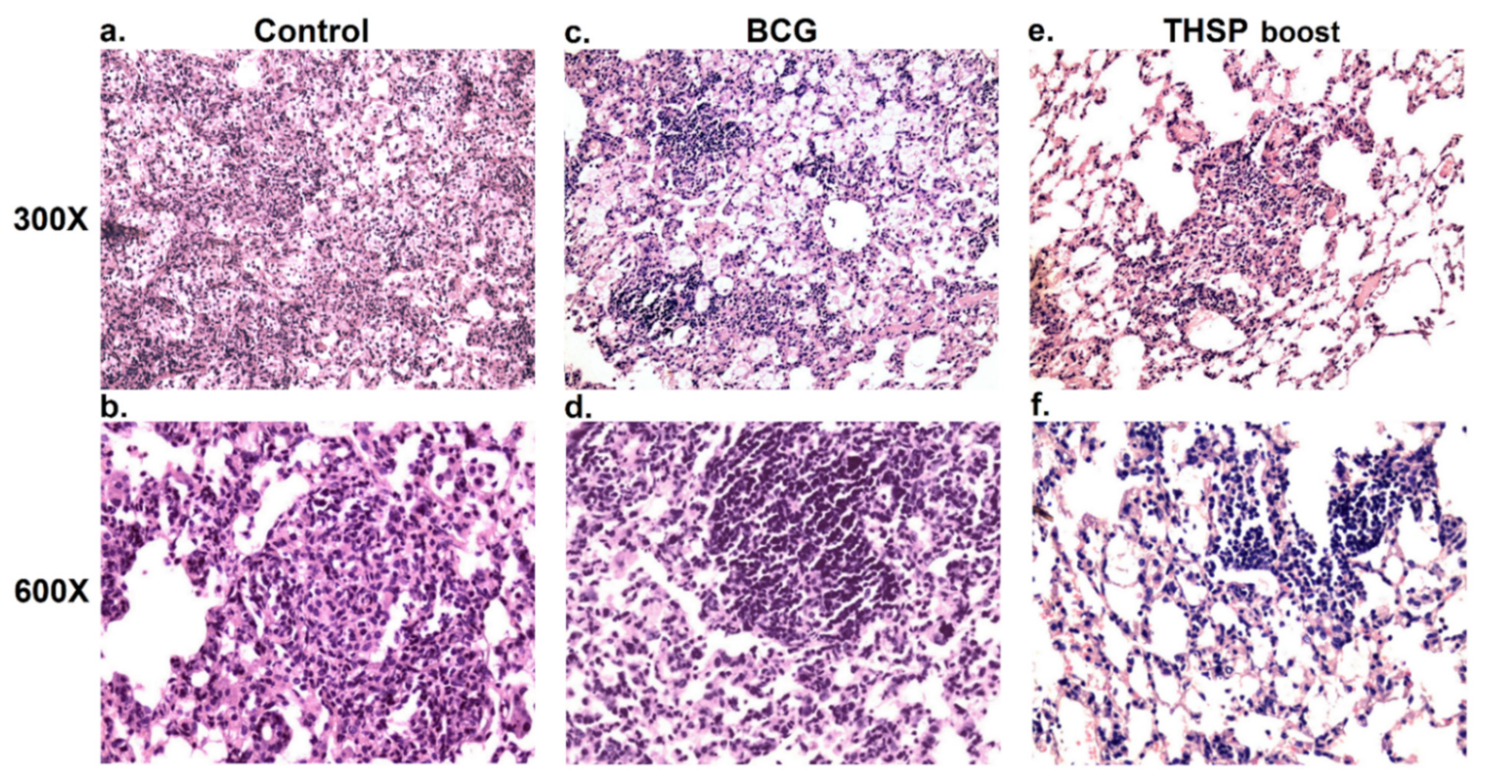
2.8. Histopathology
2.9. Statistical Analysis
3. Results
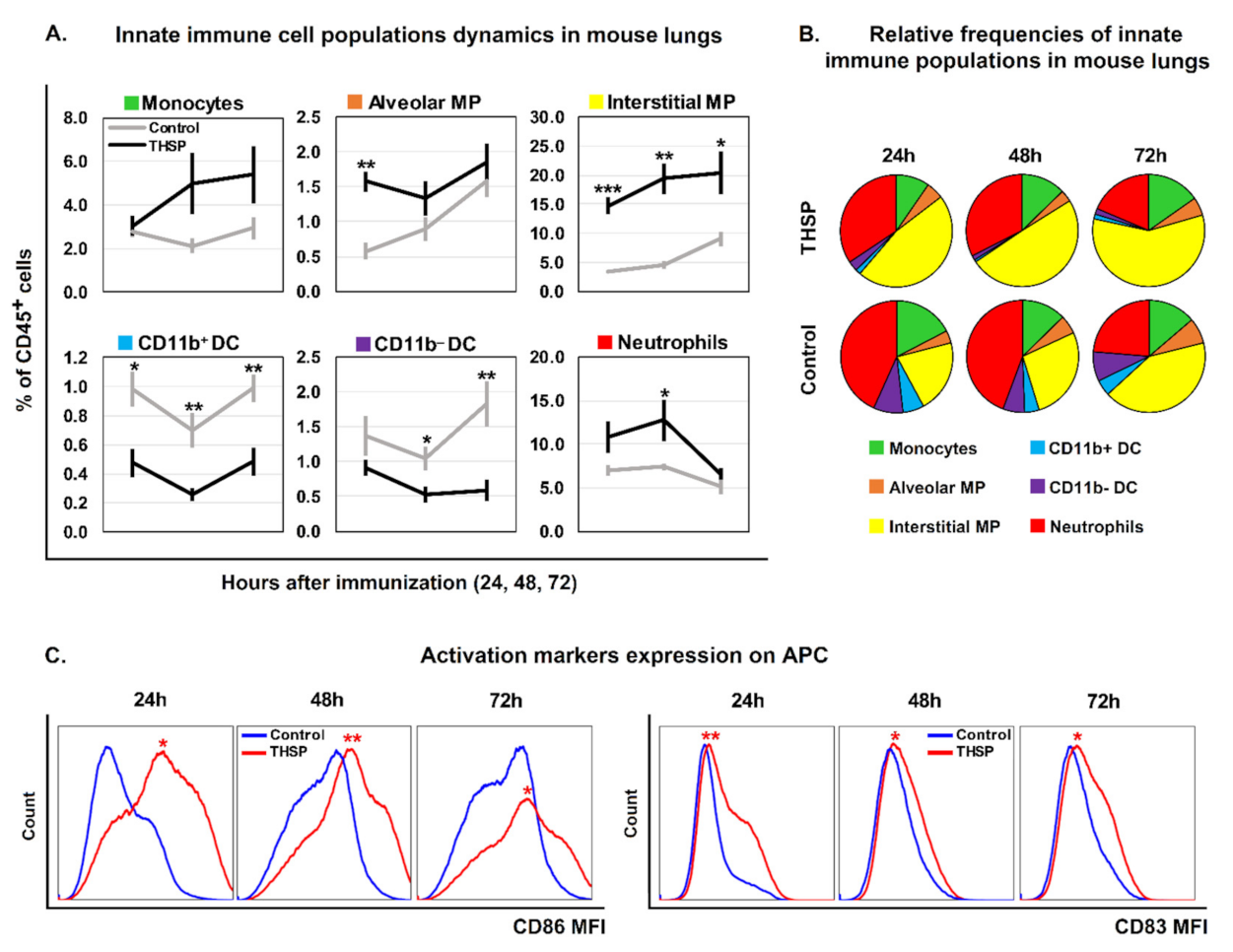
3.1. Flu/THSP Recombinant Virus Efficiently Activates Innate Immune Response at the Site of Administration
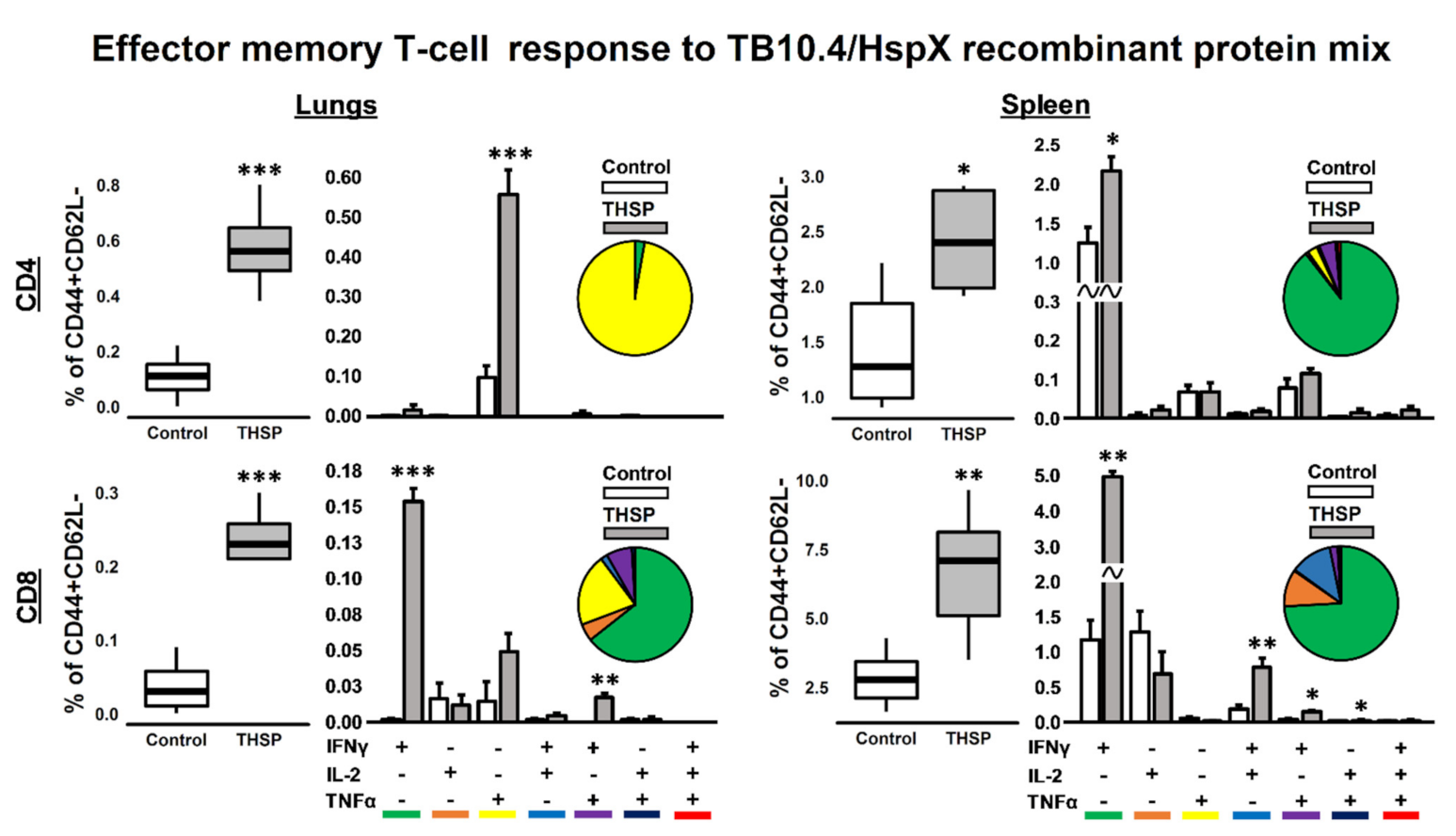
3.2. Single Flu/THSP Immunization Induces Both Local and Systemic Antigen-Specific T-Cell Response
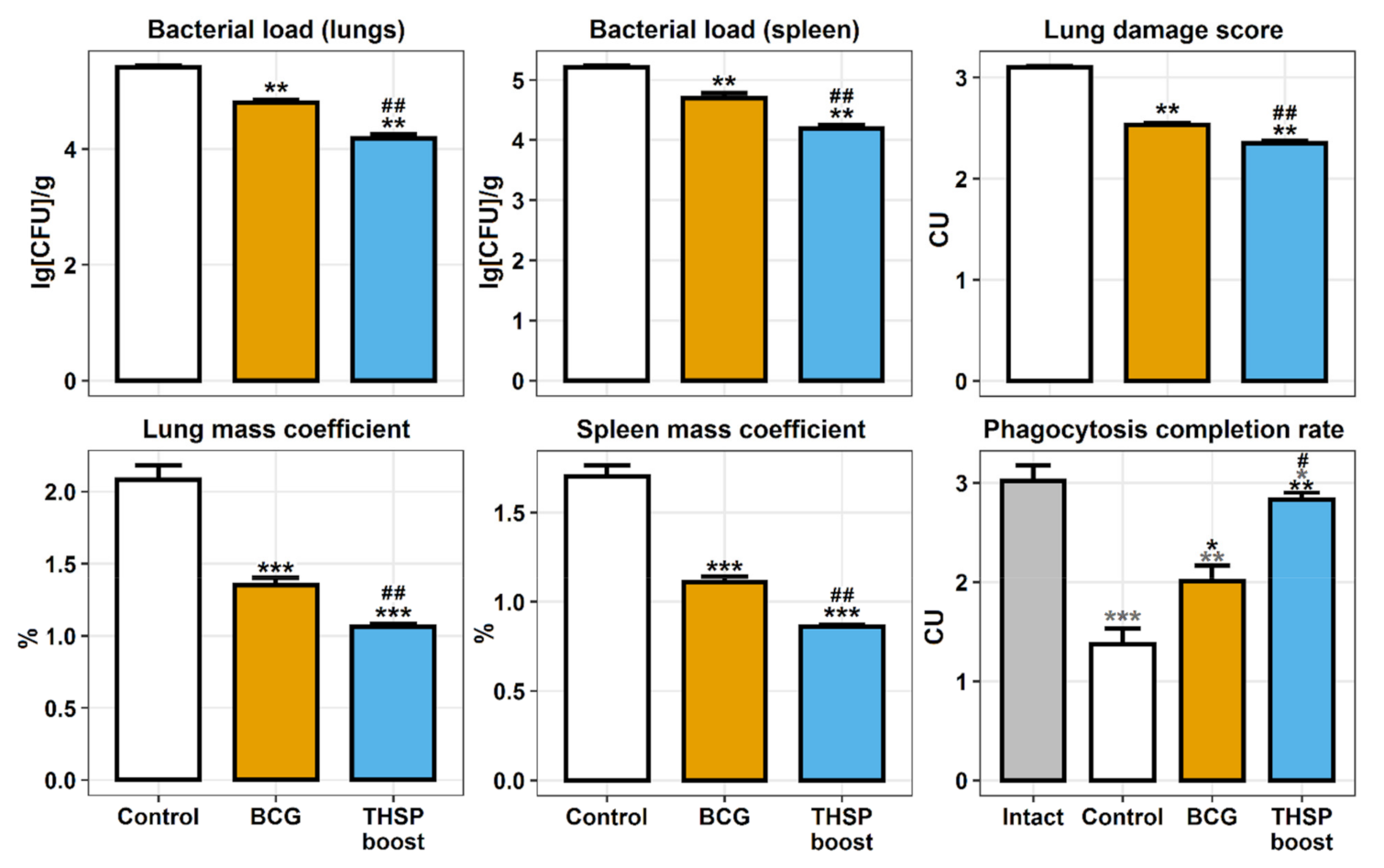
3.3. Prime-Boost Immunization with Flu/THSP Induces Better Protection against M. tb Strain H37Rv Compared to Single BCG Immunization
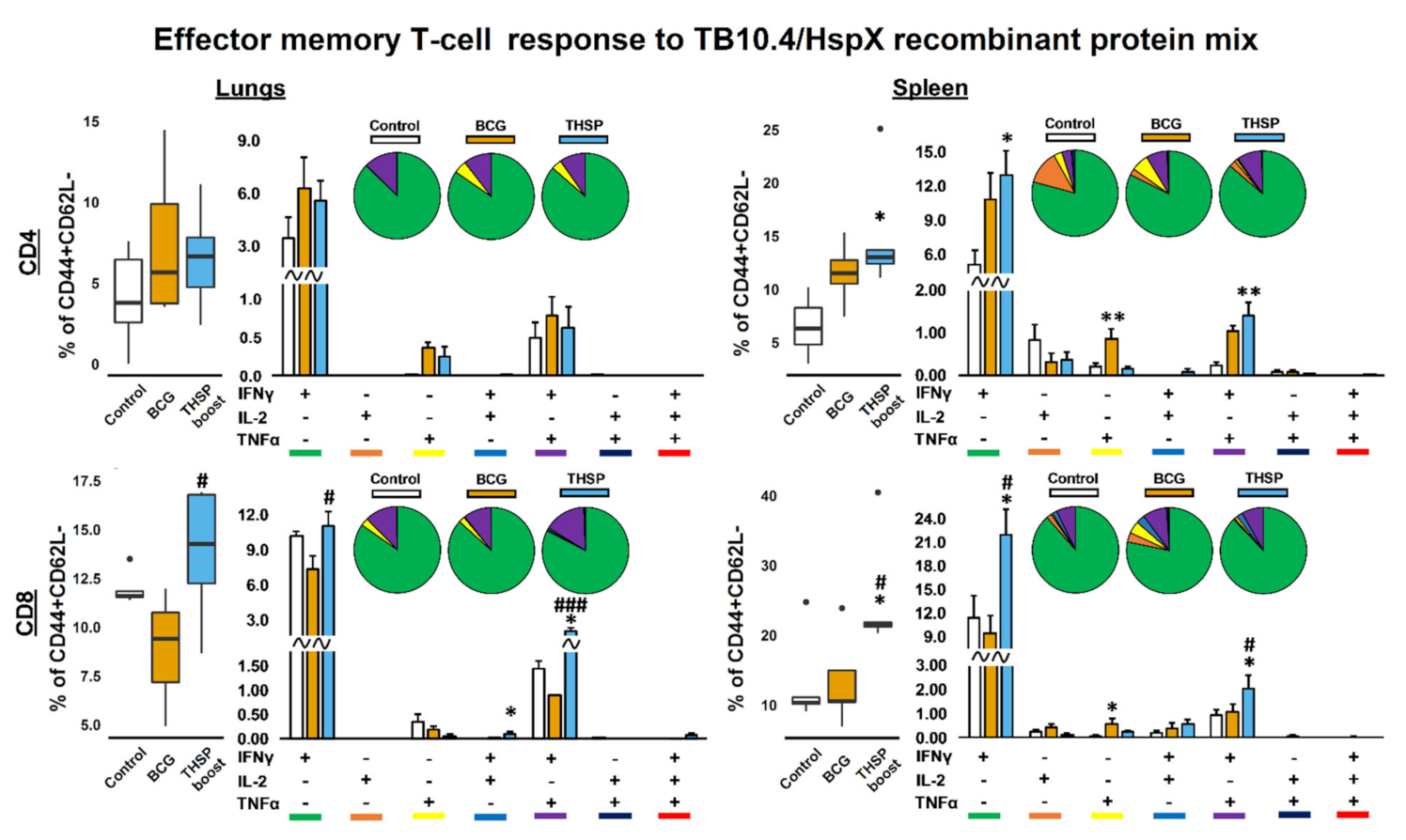
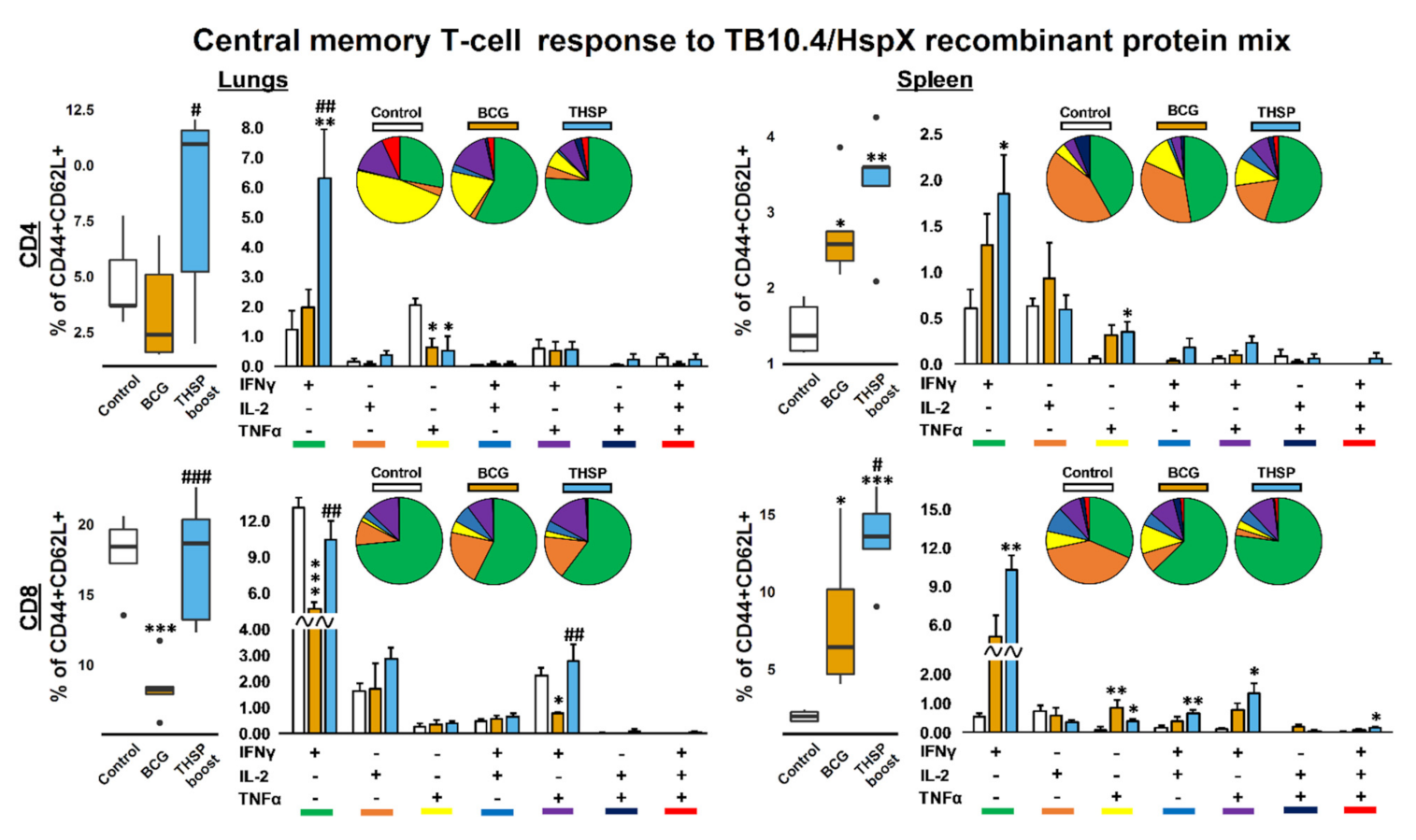
3.4. Prime-Boost Immunization with Flu/THSP Induces Higher Local and Systemic Antigen-Specific T-Cell Response Than Single BCG Vaccination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, J.; Zhao, A.; Tang, J.; Wang, G.; Shi, Y.; Zhan, L.; Qin, C. Tuberculosis vaccine development: From classic to clinical candidates. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1405–1425. [Google Scholar] [CrossRef]
- Sable, S.B.; Posey, J.E.; Scriba, T.J. Tuberculosis Vaccine Development: Progress in Clinical Evaluation. Clin. Microbiol. Rev. 2019, 33, 100–119. [Google Scholar] [CrossRef] [PubMed]
- Schrager, L.K.; Vekemens, J.; Drager, N.; Lewinsohn, D.M.; Olesen, O.F. The status of tuberculosis vaccine development. Lancet Infect. Dis. 2020, 20, e28–e37. [Google Scholar] [CrossRef]
- Orme, I.M. The Achilles heel of BCG. Tuberculosis 2010, 90, 329–332. [Google Scholar] [CrossRef]
- Flynn, J.L. Immunology of tuberculosis and implications in vaccine development. Tuberculosis 2004, 84, 93–101. [Google Scholar] [CrossRef] [PubMed]
- Ryan, A.A.; Nambiar, J.K.; Wozniak, T.; Roediger, B.; Shklovskaya, E.; Britton, W.J.; Groth, B.F.D.S.; Triccas, J. Antigen Load Governs the Differential Priming of CD8 T Cells in Response to the Bacille Calmette Guérin Vaccine orMycobacterium tuberculosisInfection. J. Immunol. 2009, 182, 7172–7177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rafi, W.; Ribeiro-Rodrigues, R.; Ellner, J.; Salgame, P. Coinfection-helminthes and tuberculosis. Curr. Opin. HIV AIDS 2012, 7, 239–244. [Google Scholar] [CrossRef] [PubMed]
- Henao-Tamayo, M.I.; Ordway, D.J.; Irwin, S.M.; Shang, S.; Shanley, C.; Orme, I.M. Phenotypic definition of effector and memory T-lymphocyte subsets in mice chronically infected with Mycobacterium tuberculosis. Clin. Vaccine Immunol. 2010, 17, 618–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beverley, P.C.L.; Sridhar, S.; Lalvani, A.; Tchilian, E. Harnessing local and systemic immunity for vaccines against tuberculosis. Mucosal Immunol. 2013, 7, 20–26. [Google Scholar] [CrossRef]
- Stylianou, E.; Paul, M.J.; Reljic, R.; McShane, H. Mucosal delivery of tuberculosis vaccines: A review of current approaches and challenges. Expert Rev. Vaccines 2019, 18, 1271–1284. [Google Scholar] [CrossRef] [PubMed]
- Hervas-Stubbs, S.; Majlessi, L.; Simsova, M.; Morova, J.; Rojas, M.-J.; Nouzé, C.; Brodin, P.; Sebo, P.; Leclerc, C. High Frequency of CD4 + T Cells Specific for the TB10.4 Protein Correlates with Protection against Mycobacterium tuberculosis Infection. Infect. Immun. 2006, 74, 3396–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.-H.; Shin, S.J.; Park, Y.-M.; Jung, I.D.; Ryu, S.-W.; Kim, D.-J.; Park, J.-H.; Park, J.-H. Critical role of TRIF and MyD88 in Mycobacterium tuberculosis Hsp70-mediated activation of dendritic cells. Cytokine 2015, 71, 139–144. [Google Scholar] [CrossRef]
- Lin, M.Y.; Geluk, A.; Smith, S.G.; Stewart, A.L.; Friggen, A.H.; Franken, K.L.; Verduyn, M.J.; van Meijgaarden, K.E.; Voskuil, M.I.; Dockrell, H.M.; et al. Lack of immune responses to Mycobacterium tuberculosis DosR regulon proteins following Mycobacterium bovis BCG vaccination. Infect. Immun. 2007, 75, 3523–3530. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Q.; Yu, H.; Zhang, Y.; Wang, B.; Jiang, W.; Da, Z.; Xian, Q.; Wang, Y.; Liu, X.; Zhu, B. Immunogenicity and Protective Efficacy of a Fusion Protein Vaccine Consisting of Antigen Ag85B and HspX against Mycobacterium tuberculosis Infection in Mice. Scand. J. Immunol. 2011, 73, 568–576. [Google Scholar] [CrossRef] [PubMed]
- de Sousa, E.M.; da Costa, A.C.; Trentini, M.M.; de AraújoFilho, J.A.; Kipnis, A.; Junqueira-Kipnis, A.P. Immunogenicity of a fusion protein containing immunodominant epitopes of Ag85C, MPT51, and HspX from Mycobacterium tuberculosis in mice and active TB infection. PLoS ONE 2012, 7, e47781. [Google Scholar] [CrossRef]
- Soleimanpour, S.; Farsiani, H.; Mosavat, A.; Ghazvini, K.; Eydgahi, M.R.; Sankian, M.; Sadeghian, H.; Meshkat, Z.; Rezaee, S.A. APC targeting enhances immunogenicity of a novel multistage Fc-fusion tuberculosis vaccine in mice. Appl. Microbiol. Biotechnol. 2015, 99, 10467–10480. [Google Scholar] [CrossRef] [PubMed]
- Yousefi-Avarvand, A.; Tafaghodi, M.; Soleimanpour, S.; Khademi, F. HspX protein as a candidate vaccine against Mycobacterium tuberculosis: An overview. Front. Biol. 2018, 13, 293–296. [Google Scholar] [CrossRef]
- Singh, S.; Saraav, I.; Sharma, S. Immunogenic potential of latency associated antigens against Mycobacterium tuberculosis. Vaccine 2014, 32, 712–716. [Google Scholar] [CrossRef] [PubMed]
- Egorov, A.; Brandt, S.; Sereinig, S.; Romanova, J.; Ferko, B.; Katinger, D.; Grassauer, A.; Alexandrova, G.; Katinger, H.; Muster, T. Transfectant Influenza A Viruses with Long Deletions in the NS1 Protein Grow Efficiently in Vero Cells. J. Virol. 1998, 72, 6437–6441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Sastre, A.; Egorov, A.; Matassova, D.; Brandt, S.; Levy, D.; Durbin, J.E.; Palese, P.; Musterbc, T. Influenza A Virus Lacking the NS1 Gene Replicates in Interferon-Deficient Systems. Virology 1998, 252, 324–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferko, B.; Stasakova, J.; Romanova, J.; Kittel, C.; Sereinig, S.; Katinger, H.; Egorov, A. Immunogenicity and Protection Efficacy of Replication-Deficient Influenza A Viruses with Altered NS1 Genes. J. Virol. 2004, 78, 13037–13045. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasilyev, K.A.; Shurygina, A.-P.S.; Stukova, M.A.; Egorov, A.Y. Enhanced CD8+ T-cell response in mice immunized with NS1-truncated influenza virus. Microbiol. Indep. Res. J. 2020, 7, 24–33. [Google Scholar] [CrossRef]
- Romanova, J.; Krenn, B.M.; Wolschek, M.; Ferko, B.; Romanovskaja-Romanko, E.; Morokutti, A.; Shurygina, A.-P.; Nakowitsch, S.; Ruthsatz, T.; Kiefmann, B.; et al. Preclinical Evaluation of a Replication-Deficient Intranasal ΔNS1 H5N1 Influenza Vaccine. PLoS ONE 2009, 4, e5984. [Google Scholar] [CrossRef] [PubMed]
- Stasakova, J.; Ferko, B.; Kittel, C.; Sereinig, S.; Romanova, J.; Katinger, H.; Egorov, A. Influenza A mutant viruses with altered NS1 protein function provoke caspase-1 activation in primary human macrophages, resulting in fast apoptosis and release of high levels of interleukins 1β and 18. J. Gen. Virol. 2005, 86, 185–195. [Google Scholar] [CrossRef] [PubMed]
- Stukova, M.A.; Sereinig, S.; Zabolotnyh, N.V.; Ferko, B.; Kittel, C.; Romanova, J.; Vinogradova, T.I.; Katinger, H.; Kiselev, O.I.; Egorov, A. Vaccine potential of influenza vectors expressing Mycobacterium tuberculosis ESAT-6 protein. Tuberculosis 2006, 86, 236–246. [Google Scholar] [CrossRef] [PubMed]
- Vasilyev, K.A.; Yukhneva, M.A.; Shurygina, A.-P.S.; Stukova, M.A.; Egorov, A.Y. Enhancement of the immunogenicity of influenza A virus by the inhibition of immunosuppressive function of NS1 protein. Microbiol. Indep. Res. J. 2018, 5, 48–58. [Google Scholar] [CrossRef]
- Vasilyev, K.; Shurygina, A.-P.; Sergeeva, M.; Stukova, M.; Egorov, A. Intranasal Immunization with the Influenza A Virus Encoding Truncated NS1 Protein Protects Mice from Heterologous Challenge by Restraining the Inflammatory Response in the Lungs. Microorganisms 2021, 9, 690. [Google Scholar] [CrossRef]
- Sereinig, S.; Stukova, M.; Zabolotnyh, N.; Ferko, B.; Kittel, C.; Romanova, J.; Vinogradova, T.; Katinger, H.; Kiselev, O.; Egorov, A. Influenza Virus NS Vectors Expressing the Mycobacterium tuberculosis ESAT-6 Protein Induce CD4 + Th1 Immune Response and Protect Animals against Tuberculosis Challenge. Clin. Vaccine Immunol. 2006, 13, 898–904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, G.; Watanabe, T.; Ito, H.; Watanabe, S.; Goto, H.; Gao, P.; Hughes, M.; Perez, D.; Donis, R.; Hoffmann, E.; et al. Generation of influenza A viruses entirely from cloned cDNAs. Proc. Natl. Acad. Sci. USA 1999, 96, 9345–9350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Misharin, A.; Morales-Nebreda, L.; Mutlu, G.M.; Budinger, G.R.S.; Perlman, H. Flow Cytometric Analysis of Macrophages and Dendritic Cell Subsets in the Mouse Lung. Am. J. Respir. Cell Mol. Biol. 2013, 49, 503–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.-R.A.; O’Koren, E.G.; Hotten, D.F.; Kan, M.J.; Kopin, D.; Nelson, E.R.; Que, L.; Gunn, M.D. A Protocol for the Comprehensive Flow Cytometric Analysis of Immune Cells in Normal and Inflamed Murine Non-Lymphoid Tissues. PLoS ONE 2016, 11, e0150606. [Google Scholar] [CrossRef] [Green Version]
- Salgame, P. Host innate and Th1 responses and the bacterial factors that control Mycobacterium tuberculosis infection. Curr. Opin. Immunol. 2005, 17, 374–380. [Google Scholar] [CrossRef]
- Lyadova, I.V.; Panteleev, A.V. Th1 and Th17 cells in tuberculosis: Protection, pathology, and biomarkers. Mediat. Inflamm. 2015, 2015, 854507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeng, G.; Zhang, G.; Chen, X. Th1 cytokines, true functional signatures for protective immunity against TB? Cell. Mol. Immunol. 2018, 15, 206–215. [Google Scholar] [CrossRef] [Green Version]
- Lalvani, A.; Brookes, R.; Wilkinson, R.J.; Malin, A.S.; Pathan, A.A.; Andersen, P.; Dockrell, H.; Pasvol, G.; Hill, A.V.S. Human cytolytic and interferon γ-secreting CD8+ T lymphocytes specific for Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 1998, 95, 270–275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caccamo, N.; Pietra, G.; Sullivan, L.C.; Brooks, A.G.; Prezzemolo, T.; La Manna, M.P.; Di Liberto, D.; Joosten, S.A.; van Meijgaarden, K.E.; Di Carlo, P.; et al. Human CD8 T lymphocytes recognize Mycobacterium tuberculosis antigens presented by HLA-E during active tuberculosis and express type 2 cytokines. Eur. J. Immunol. 2015, 45, 1069–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Darrah, P.A.; Patel, D.T.; De Luca, P.M.; Lindsay, R.W.B.; Davey, D.F.; Flynn, B.J.; Hoff, S.T.; Andersen, P.; Reed, S.G.; Morris, S.L.; et al. Multifunctional TH1 cells define a correlate of vaccine-mediated protection against Leishmania major. Nat. Med. 2007, 13, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Sergeeva, M.; Romanovskaya-Romanko, E.; Zabolotnyh, N.; Pulkina, A.; Vasilyev, K.; Shurigina, A.; Buzitskaya, J.; Zabrodskaya, Y.; Fadeev, A.; Vasin, A.; et al. Mucosal Influenza Vector Vaccine Carrying TB10.4 and HspX Antigens Provides Protection against Mycobacterium tuberculosis in Mice and Guinea Pigs. Vaccines 2021, 9, 394. [Google Scholar] [CrossRef] [PubMed]
- Schweitzer, A.N.; Borriello, F.; Wong, R.C.; Abbas, A.K.; Sharpe, A.H. Role of costimulators in T cell differentiation: Studies using antigen-presenting cells lacking expression of CD80 or CD86. J. Immunol. 1997, 158, 2713–2722. [Google Scholar]
- Lechmann, M.; Berchtold, S.; Steinkasserer, A.; Hauber, J. CD83 on dendritic cells: More than just a marker for maturation. Trends Immunol. 2002, 23, 273–275. [Google Scholar] [CrossRef]
- O’Garra, A.; Redford, P.S.; McNab, F.W.; Bloom, C.I.; Wilkinson, R.J.; Berry, M.P.R. The immune response in tuberculosis. Annu. Rev. Immunol. 2013, 31, 475–527. [Google Scholar] [CrossRef] [PubMed]
- Flynn, J.L.; Chan, J.; Triebold, K.J.; Dalton, D.K.; A Stewart, T.; Bloom, B.R. An essential role for interferon gamma in resistance to Mycobacterium tuberculosis infection. J. Exp. Med. 1993, 178, 2249–2254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, A.M.; Dalton, D.K.; A Stewart, T.; Griffin, J.P.; Russell, D.; Orme, I.M. Disseminated tuberculosis in interferon gamma gene-disrupted mice. J. Exp. Med. 1993, 178, 2243–2247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forbes, E.K.; Sander, C.; Ronan, E.O.; McShane, H.; Hill, A.V.S.; Beverley, P.C.L.; Tchilian, E.Z. Multifunctional, High-Level Cytokine-Producing Th1 Cells in the Lung, but Not Spleen, Correlate with Protection against Mycobacterium tuberculosis Aerosol Challenge in Mice. J. Immunol. 2008, 181, 4955–4964. [Google Scholar] [CrossRef] [Green Version]
- Caccamo, N.R.; Guggino, G.; Joosten, S.A.; Gelsomino, G.; Di Carlo, P.; Titone, L.; Galati, D.; Bocchino, M.; Matarese, A.; Salerno, A.; et al. Multifunctional CD4+ T cells correlate with active Mycobacterium tuberculosis infection. Eur. J. Immunol. 2010, 40, 2211–2220. [Google Scholar] [CrossRef] [PubMed]
- Derrick, S.C.; Yabe, I.M.; Yang, A.; Morris, S.L. Vaccine-induced anti-tuberculosis protective immunity in mice correlates with the magnitude and quality of multifunctional CD4 T cells. Vaccine 2011, 29, 2902–2909. [Google Scholar] [CrossRef] [PubMed]
- Lewinsohn, D.A.; Lewinsohn, D.M.; Scriba, T. Polyfunctional CD4+ T Cells As Targets for Tuberculosis Vaccination. Front. Immunol. 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vasilyev, K.; Shurygina, A.-P.; Zabolotnykh, N.; Sergeeva, M.; Romanovskaya-Romanko, E.; Pulkina, A.; Buzitskaya, J.; Dogonadze, M.Z.; Vinogradova, T.I.; Stukova, M.A. Enhancement of the Local CD8+ T-Cellular Immune Response to Mycobacterium tuberculosis in BCG-Primed Mice after Intranasal Administration of Influenza Vector Vaccine Carrying TB10.4 and HspX Antigens. Vaccines 2021, 9, 1273. https://doi.org/10.3390/vaccines9111273
Vasilyev K, Shurygina A-P, Zabolotnykh N, Sergeeva M, Romanovskaya-Romanko E, Pulkina A, Buzitskaya J, Dogonadze MZ, Vinogradova TI, Stukova MA. Enhancement of the Local CD8+ T-Cellular Immune Response to Mycobacterium tuberculosis in BCG-Primed Mice after Intranasal Administration of Influenza Vector Vaccine Carrying TB10.4 and HspX Antigens. Vaccines. 2021; 9(11):1273. https://doi.org/10.3390/vaccines9111273
Chicago/Turabian StyleVasilyev, Kirill, Anna-Polina Shurygina, Natalia Zabolotnykh, Mariia Sergeeva, Ekaterina Romanovskaya-Romanko, Anastasia Pulkina, Janna Buzitskaya, Marine Z. Dogonadze, Tatiana I. Vinogradova, and Marina A. Stukova. 2021. "Enhancement of the Local CD8+ T-Cellular Immune Response to Mycobacterium tuberculosis in BCG-Primed Mice after Intranasal Administration of Influenza Vector Vaccine Carrying TB10.4 and HspX Antigens" Vaccines 9, no. 11: 1273. https://doi.org/10.3390/vaccines9111273