
Novel Recombinant Newcastle Disease Virus-Based In Ovo Vaccines Bypass Maternal Immunity to Provide Full Protection from Early Virulent Challenge

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses
2.2. Eggs
2.3. Vaccination
2.4. Hatching, Sampling, Mortality
2.5. Challenge
2.6. Real-Time RT-PCR
2.7. HI Assay
2.8. Statistical Analyses
3. Results
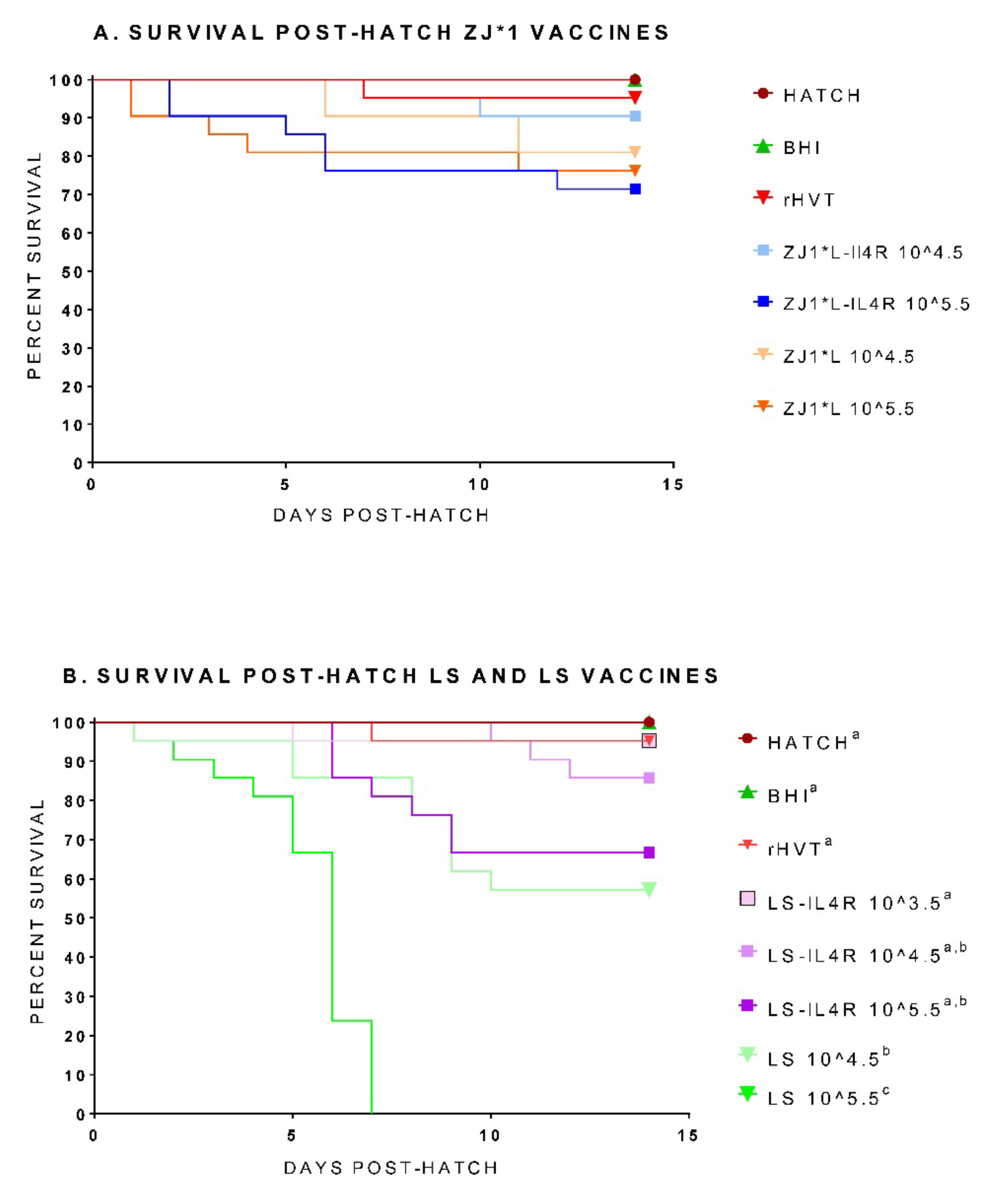
3.1. Hatchability, Post-Hatch Survival, and Body Weights
3.2. Shedding of Vaccine Viruses Post-Vaccination, as Demonstrated by rRT-PCR
3.3. Humoral Immune Response Post-Hatch
3.4. Clinical Signs and Survival after Early Challenge with vNDV
3.5. Shedding Post-Challenge as Demonstrated by rRT-PCR
3.6. Post-Challenge Serology
3.7. Inoculated SPF Control Groups
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Suarez, D.L.; Miller, P.J.; Koch, G.; Mundt, E.; Rautenschlein, S. Newcastle disease, Other Avian Paramyxoviruses, and Avian Metapneumovirus infections. In Diseases of Poultry, 13th ed.; Swayne, D.E., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2013; pp. 89–138. [Google Scholar] [CrossRef]
- International Committee on Taxonomy of Viruses. Virus Taxonomy: 2018 Release. Available online: https://talk.ictvonline.org/taxonomy/ (accessed on 8 September 2020).
- Amarasinghe, G.K.; Ayllon, M.A.; Bao, Y.; Basler, C.F.; Bavari, S.; Blasdell, K.R.; Briese, T.; Brown, P.A.; Bukreyev, A.; Balkema-Buschmann, A.; et al. Taxonomy of the order Mononegavirales: Update 2019. Arch. Virol. 2019, 164, 1967–1980. [Google Scholar] [CrossRef] [Green Version]
- Lamb, R.A.; Parks, G.D. Paramyxoviridae. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott, Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 957–995. [Google Scholar]
- Czeglédi, A.; Ujvári, D.; Somogyi, E.; Wehmann, E.; Werner, O.; Lomniczi, B. Third genome size category of avian paramyxovirus serotype 1 (Newcastle disease virus) and evolutionary implications. Virus Res. 2006, 120, 36–48. [Google Scholar] [CrossRef] [PubMed]
- The World Bank; The TAFS forum. World Livestock Disease Atlas. A Quantitative Analysis of Global Animal Health Data (2006–2009); The International Bank for Reconstruction and Development: Washington, DC, USA, 2011. [Google Scholar]
- Dimitrov, K.M.; Ramey, A.M.; Qiu, X.; Bahl, J.; Afonso, C.L. Temporal, geographic, and host distribution of avian paramyxovirus 1 (Newcastle disease virus). Infect. Genet. Evol. 2016, 39, 22–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mayers, J.; Mansfield, K.L.; Brown, I.H. The role of vaccination in risk mitigation and control of Newcastle disease in poultry. Vaccine 2017, 35, 5974–5980. [Google Scholar] [CrossRef] [PubMed]
- Dimitrov, K.M.; Afonso, C.L.; Yu, Q.; Miller, P.J. Newcastle disease vaccines—A solved problem or a continuous challenge? Vet. Microbiol. 2017, 206, 126–136. [Google Scholar] [CrossRef] [PubMed]
- Senne, D.A.; King, D.J.; Kapczynski, D.R. Control of Newcastle disease by vaccination. Dev. Biol. 2004, 119, 165–170. [Google Scholar] [PubMed]
- OIE. World Animal Health Information Database (WAHIS Interface)—Version 1. Available online: http://www.oie.int/wahis_2/public/wahid.php/Wahidhome/Home (accessed on 27 November 2019).
- Armour, N.K.; García, M. Current and Future Applications of Viral-Vectored Recombinant Vaccines. In The Poultry Informed Professional; Department of Population Health, University of Georgia: Athens, GA, USA, 2014; pp. 1–9. [Google Scholar]
- Alexander, D.J. Newcastle disease virus—An avian Paramyxovirus. In Newcastle Disease; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 11–22. [Google Scholar]
- Alexander, D.J.; Manvell, R.J.; Lowings, J.P.; Frost, K.M.; Collins, M.S.; Russell, P.H.; Smith, J.E. Antigenic diversity and similarities detected in avian paramyxovirus type 1 (Newcastle disease virus) isolates using monoclonal antibodies. Avian Pathol. 1997, 26, 399–418. [Google Scholar] [CrossRef]
- Miller, P.J.; King, D.J.; Afonso, C.L.; Suarez, D.L. Antigenic differences among Newcastle disease virus strains of different genotypes used in vaccine formulation affect viral shedding after a virulent challenge. Vaccine 2007, 25, 7238–7246. [Google Scholar] [CrossRef]
- Miller, P.J.; Estevez, C.; Yu, Q.; Suarez, D.L.; King, D.J. Comparison of viral shedding following vaccination with inactivated and live Newcastle disease vaccines formulated with wild-type and recombinant viruses. Avian Dis. 2009, 53, 39–49. [Google Scholar] [CrossRef]
- Xiao, S.; Nayak, B.; Samuel, A.; Paldurai, A.; Kanabagatte Basavarajappa, M.; Prajitno, T.Y.; Bharoto, E.E.; Collins, P.L.; Samal, S.K. Generation by reverse genetics of an effective, stable, live-attenuated newcastle disease virus vaccine based on a currently circulating, highly virulent Indonesian strain. PLoS ONE 2012, 7, e52751. [Google Scholar] [CrossRef] [Green Version]
- Kapczynski, D.R.; Martin, A.; Haddad, E.E.; King, D.J. Protection from clinical disease against three highly virulent strains of Newcastle disease virus after in ovo application of an antibody-antigen complex vaccine in maternal antibody-positive chickens. Avian Dis. 2012, 56, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Schat, K. Back to the Past: Do Vector Vaccines Represent the Future? Cornell University: Ithaca, NY, USA, 2015; pp. 1–12. [Google Scholar]
- Johnston, P.A.; Liu, H.; O’Connell, T.; Phelps, P.; Bland, M.; Tyczkowski, J.; Kemper, A.; Harding, T.; Avakian, A.; Haddad, E.; et al. Applications in in ovo technology. Poult. Sci. 1997, 76, 165–178. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.J.; Zedek, A.S. Comparative field evaluations of in ovo applied technology. Poult. Sci. 2010, 89, 189–193. [Google Scholar] [CrossRef]
- Wen, G.; Li, L.; Yu, Q.; Wang, H.; Luo, Q.; Zhang, T.; Zhang, R.; Zhang, W.; Shao, H. Evaluation of a thermostable Newcastle disease virus strain TS09-C as an in-ovo vaccine for chickens. PLoS ONE 2017, 12, e0172812. [Google Scholar] [CrossRef]
- Avakian, A.P.; Poston, R.M.; Kong, F.K.; Van Kampen, K.R.; Tang, D.C. Automated mass immunization of poultry: The prospect for nonreplicating human adenovirus-vectored in ovo vaccines. Expert Rev. Vaccines 2007, 6, 457–465. [Google Scholar] [CrossRef]
- Wakenell, P.S.; Bryan, T.; Schaeffer, J.; Avakian, A.; Williams, C.; Whitfill, C. Effect of in ovo vaccine delivery route on herpesvirus of turkeys/SB-1 efficacy and viremia. Avian. Dis. 2002, 46, 274–280. [Google Scholar] [CrossRef]
- Karaca, K.; Sharma, J.M.; Winslow, B.J.; Junker, D.E.; Reddy, S.; Cochran, M.; McMillen, J. Recombinant Fowlpox Viruses Coexpressing Chicken Type I IFN and Newcastle Disease Virus HN and F Genes: Influence of IFN on Protective Efficacy and Humoral Responses of Chickens Following in ovo or Post-Hatch Administration of Recombinant Viruses. Vaccine 1998, 16, 1496–1503. [Google Scholar] [CrossRef]
- Rautenschlein, S.; Sharma, J.M.; Winslow, B.J.; McMillen, J.; Junker, D.; Cochran, M. Embryo vaccination of turkeys against Newcastle disease infection with recombinant fowlpox virus constructs containing interferons as adjuvants. Vaccine 1999, 18, 426–433. [Google Scholar] [CrossRef]
- United States Department of Agriculture, Animal and Plant Health Inspection Service. Veterinary Biological Products in Licensed Establishments; Center for Veterinary Biologics: Ames, IA, USA, 2019; pp. 1–39.
- Esaki, M.; Godoy, A.; Rosenberger, J.K.; Rosenberger, S.C.; Gardin, Y.; Yasuda, A.; Dorsey, K.M. Protection and antibody response caused by turkey herpesvirus vector Newcastle disease vaccine. Avian Dis. 2013, 57, 750–755. [Google Scholar] [CrossRef]
- Palya, V.; Kiss, I.; Tatar-Kis, T.; Mato, T.; Felfoldi, B.; Gardin, Y. Advancement in vaccination against Newcastle disease: Recombinant HVT NDV provides high clinical protection and reduces challenge virus shedding with the absence of vaccine reactions. Avian Dis. 2012, 56, 282–287. [Google Scholar] [CrossRef]
- Toro, H.; Tang, D.C.; Suarez, D.L.; Zhang, J.; Shi, Z. Protection of chickens against avian influenza with non-replicating adenovirus-vectored vaccine. Vaccine 2008, 26, 2640–2646. [Google Scholar] [CrossRef]
- Schultz-Cherry, S.; Dybing, J.K.; Davis, N.L.; Williamson, C.; Suarez, D.L.; Johnston, R.; Perdue, M.L. Influenza virus (A/HK/156/97) hemagglutinin expressed by an alphavirus replicon system protects chickens against lethal infection with Hong Kong-origin H5N1 viruses. Virology 2000, 278, 55–59. [Google Scholar] [CrossRef] [Green Version]
- Mebatsion, T.; Verstegen, S.; De Vaan, L.T.; Romer-Oberdorfer, A.; Schrier, C.C. A recombinant newcastle disease virus with low-level V protein expression is immunogenic and lacks pathogenicity for chicken embryos. J. Virol. 2001, 75, 420–428. [Google Scholar] [CrossRef] [Green Version]
- Steel, J.; Burmakina, S.V.; Thomas, C.; Spackman, E.; Garcia-Sastre, A.; Swayne, D.E.; Palese, P. A combination in-ovo vaccine for avian influenza virus and Newcastle disease virus. Vaccine 2008, 26, 522–531. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, J.; Sharma, J.M. Evaluation of a modified-live virus vaccine administered in ovo to protect chickens against Newcastle disease. Am. J. Vet. Res. 1992, 53, 1999–2004. [Google Scholar] [PubMed]
- Ge, J.; Wang, X.; Tian, M.; Wen, Z.; Feng, Q.; Qi, X.; Gao, H.; Wang, X.; Bu, Z. Novel in-ovo chimeric recombinant Newcastle disease vaccine protects against both Newcastle disease and infectious bursal disease. Vaccine 2014, 32, 1514–1521. [Google Scholar] [CrossRef] [PubMed]
- Ramp, K.; Topfstedt, E.; Wackerlin, R.; Hoper, D.; Ziller, M.; Mettenleiter, T.C.; Grund, C.; Romer-Oberdorfer, A. Pathogenicity and immunogenicity of different recombinant Newcastle disease virus clone 30 variants after in ovo vaccination. Avian Dis. 2012, 56, 208–217. [Google Scholar] [CrossRef] [PubMed]
- Dilaveris, D.; Chen, C.; Kaiser, P.; Russell, P.H. The safety and immunogenicity of an in ovo vaccine against Newcastle disease virus differ between two lines of chicken. Vaccine 2007, 25, 3792–3799. [Google Scholar] [CrossRef] [PubMed]
- Marcano, V.C.; Cardenas-Garcia, S.; Diel, D.G.; Antoniassi da Silva, L.H.; Gogal, R.M.; Miller, P.J.; Brown, C.C.; Butt, S.L.; Goraichuk, I.V.; Dimitrov, K.M.; et al. A Novel Recombinant Newcastle Disease Vaccine Improves Post-In Ovo Vaccination Survival with Sustained Protection against Virulent Challenge. Vaccines 2021, 9, 953. [Google Scholar] [CrossRef] [PubMed]
- Cardenas Garcia, S.; Dunwoody, R.P.; Marcano, V.; Diel, D.G.; Williams, R.J.; Gogal, R.M.; Brown, C.C., Jr.; Miller, P.J.; Afonso, C.L. Effects of Chicken Interferon Gamma on Newcastle Disease Virus Vaccine Immunogenicity. PLoS ONE 2016, 11, e0159153. [Google Scholar] [CrossRef] [PubMed]
- Susta, L.; Cornax, I.; Diel, D.G.; Garcia, S.C.; Miller, P.J.; Liu, X.; Hu, S.; Brown, C.C.; Afonso, C.L. Expression of interferon gamma by a highly virulent strain of Newcastle disease virus decreases its pathogenicity in chickens. Microb. Pathog. 2013, 61–62, 73–83. [Google Scholar] [CrossRef]
- Estevez, C.; King, D.; Seal, B.; Yu, Q. Evaluation of Newcastle disease virus chimeras expressing the hemagglutinin-neuraminidase protein of velogenic strains in the context of a mesogenic recombinant virus backbone. Virus Res. 2007, 129, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Senne, D.A. Virus propagation in embryonating eggs. In A Laboratory Manual for the Isolation, Identification and Characterization of Avian Pathogens, 5th ed.; Dufour-Zavala, L., Swayne, D.E., Glisson, J.R., Pearson, J.E., Reed, W.M., Jackwood, M.W., Woolcock, P.R., Eds.; The American Association of Avian Pathologists: Athens, GA, USA, 2008; pp. 204–208. [Google Scholar]
- United States Department of Agriculture, Animal and Plant Health Inspection Service. Supplemental Assay Method for Titration of Marek’s Serotype 3; USDA/CVB, SAM 407.05; Center for Veterinary Biologics: Ames, IA, USA, 2014; pp. 1–10.
- Das, A.; Spackman, E.; Pantin-Jackwood, M.J.; Suarez, D.L. Removal of real-time reverse transcription polymerase chain reaction (RT-PCR) inhibitors associated with cloacal swab samples and tissues for improved diagnosis of Avian influenza virus by RT-PCR. J. Vet. Diagn Invest. 2009, 21, 771–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, M.G.; Suarez, D.L.; Seal, B.S.; Pedersen, J.C.; Senne, D.A.; King, D.J.; Kapczynski, D.R.; Spackman, E. Development of a real-time reverse-transcription PCR for detection of newcastle disease virus RNA in clinical samples. J. Clin. Microbiol. 2004, 42, 329–338. [Google Scholar] [CrossRef] [Green Version]
- OIE. Newcastle Disease, Biological Standards Commission. Manual of Diagnostic Tests and Vaccines for Terrestrial Animals: Mammals, Birds and Bees, 7th ed.; World Organisation for Animal Health: Paris, France, 2012; pp. 555–574. [Google Scholar]
- Vrdoljak, A.; Halas, M.; Suli, T. Vaccination of broilers against Newcastle disease in the presence of maternally derived antibodies. Tierarztl. Prax. Ausg. G Grosstiere/Nutztiere 2017, 45, 151–158. [Google Scholar] [CrossRef]
- Westbury, H.A.; Parsons, G.; Allan, W.H. Comparison of the immunogenicity of Newcastle disease virus strains V4, Hitchner B1 and La Sota in chickens. 2. Tests in chickens with maternal antibody to the virus. Aust. Vet. J. 1984, 61, 10–13. [Google Scholar] [CrossRef]
- Kapczynski, D.R.; King, D.J. Protection of chickens against overt clinical disease and determination of viral shedding following vaccination with commercially available Newcastle disease virus vaccines upon challenge with highly virulent virus from the California 2002 exotic Newcastle disease outbreak. Vaccine 2005, 23, 3424–3433. [Google Scholar] [CrossRef]
- Sonoda, K.; Sakaguchi, M.; Okamura, H.; Yokogawa, K.; Tokunaga, E.; Tokiyoshi, S.; Kawaguchi, Y.; Hirai, K. Development of an Effective Polyvalent Vaccine Against Both Marek’s and Newcastle Diseases Based on Recombinant Marek’s Disease Virus Type 1 in Commercial Chickens with Maternal Antibodies. J. Virol. 2000, 74, 3217–3226. [Google Scholar] [CrossRef] [Green Version]
- Elazab, M.F.; Fukushima, Y.; Fujita, Y.; Horiuchi, H.; Matsuda, H.; Furusawa, S. Induction of immune suppression in the chick by an optimal dose of an immunizing antigen in the presence of its specific maternal antibody. J. Vet. Med. Sci. 2010, 72, 257–262. [Google Scholar] [CrossRef] [Green Version]
- Lancaster, J.E. Chapter 3: Control Measures. Live and inactivated vaccines compared. In Newcastle Disease: A Review 1926–1964 Monograph No. 3; Canadian Department of Agriculture: Ottawa, ON, Canada, 1966; pp. 80–115. [Google Scholar]
- Faulkner, O.B.; Estevez, C.; Yu, Q.; Suarez, D.L. Passive antibody transfer in chickens to model maternal antibody after avian influenza vaccination. Vet. Immunol. Immunopathol. 2013, 152, 341–347. [Google Scholar] [CrossRef]
- Sarfati-Mizrahi, D.; Lozano-Dubernard, B.; Soto-Priante, E.; Castro-Peralta, F.; Flores-Castro, R.; Loza-Rubio, E.; Gay-Gutierrez, M. Protective dose of a recombinant Newcastle disease LaSota-avian influenza virus H5 vaccine against H5N2 highly pathogenic avian influenza virus and velogenic viscerotropic Newcastle disease virus in broilers with high maternal antibody levels. Avian Dis. 2010, 54, 239–241. [Google Scholar] [CrossRef]
- Schroer, D.; Veits, J.; Keil, G.; Romer-Oberdorfer, A.; Weber, S.; Mettenleiter, T.C. Efficacy of Newcastle disease virus recombinant expressing avian influenza virus H6 hemagglutinin against Newcastle disease and low pathogenic avian influenza in chickens and turkeys. Avian Dis. 2011, 55, 201–211. [Google Scholar] [CrossRef]
- Van Boven, M.; Bouma, A.; Fabri, T.H.; Katsma, E.; Hartog, L.; Koch, G. Herd immunity to Newcastle disease virus in poultry by vaccination. Avian. Pathol. 2008, 37, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Alexander, D.J.; Bell, J.G.; Alders, R.G. A Technology Review: Newcastle Disease, with Special Emphasis on its Effect on Village Chickens; FAO: Rome, Italy, 2004; pp. 1–26. [Google Scholar]
- Jochemsen, P.; Jeurissen, S.H. The localization and uptake of in ovo injected soluble and particulate substances in the chicken. Poult. Sci. 2002, 81, 1811–1817. [Google Scholar] [CrossRef] [PubMed]
- Palya, V.; Tatar-Kis, T.; Mato, T.; Felfoldi, B.; Kovacs, E.; Gardin, Y. Onset and long-term duration of immunity provided by a single vaccination with a turkey herpesvirus vector ND vaccine in commercial layers. Vet. Immunol. Immunopathol. 2014, 158, 105–115. [Google Scholar] [CrossRef]
- Alexander, D.J. Historical Aspects. In Newcastle Disease; Kluwer Academic Publishers: Boston, MA, USA, 1988; pp. 1–10. [Google Scholar]
- Miller, P.J.; Afonso, C.L.; El Attrache, J.; Dorsey, K.M.; Courtney, S.C.; Guo, Z.; Kapczynski, D.R. Effects of Newcastle disease virus vaccine antibodies on the shedding and transmission of challenge viruses. Dev. Comp. Immunol. 2013, 41, 505–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, M.S.; Steel, J.; Garcia-Sastre, A.; Swayne, D.; Palese, P. Engineered viral vaccine constructs with dual specificity: Avian influenza and Newcastle disease. Proc. Natl. Acad. Sci. USA 2006, 103, 8203–8208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Veits, J.; Wiesner, D.; Fuchs, W.; Hoffmann, B.; Granzow, H.; Starick, E.; Mundt, E.; Schirrmeier, H.; Mebatsion, T.; Mettenleiter, T.C.; et al. Newcastle disease virus expressing H5 hemagglutinin gene protects chickens against Newcastle disease and avian influenza. Proc. Natl. Acad. Sci. USA 2006, 103, 8197–8202. [Google Scholar] [CrossRef] [Green Version]
- Stanford, M.M.; McFadden, G. The ‘supervirus’? Lessons from IL-4-expressing poxviruses. Trends Immunol. 2005, 26, 339–345. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DPH or DPC | Mean Weights (Standard Deviations) of Chickens in Control and Vaccine Groups Presented in Grams | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HATCH Control | BHI Control | rHVT-ND Control | LS 104.5 Control | LS 105.5 Control | ZJ1*L 104.5 | ZJ1*L 105.5 | ZJ1*L-IL4R 104.5 | ZJ1*L-IL4R 105.5 | LS-IL4R 103.5 | LS-IL4R 104.5 | LS-IL4R 105.5 | |
| 1 DPH | 35.52 a,b (3.27) | 39.67 a,b (2.31) | 42.14 b,c (2.95) | 38.50 a (3.00) | 40.25 a,b(3.178) | 38.05 a (2.44) | 40.21 a,b (1.90) | 42.57 b,c (3.30) | 40.71 a,b (2.64) | 40.24 a,b (2.76) | 39.05 a,b (2.66) | 40.01 a,b (2.77) |
| 8 DPH | 76.81 a,b (9.95) | 81.76 a,b (6.38) | 81.95a,b (7.46) | 53.5 c (21.24) | NA | 54.26 c (15.34) | 58.06 c (6.74) | 67.50 a,c (14.73) | 69.85 a,c (13.24) | 71.55 a,c (13.74) | 63.33 c (14.19) | 50.00 c (14.23) |
| 13 DPH | 123.80 a,b(14.87) | 123.50 a,b (9.67) | 129.40a,b (11.78) | 95.75 c (27.18) | NA | 99.59 c (19.21) | 97.38c (13.84) | 113.40 a,c (13.88) | 119.40 a,c (11.89) | 116.70 a,c (23.30) | 110.70c (16.13) | 88.93 c (24.88) |
| 14 DPC | 265.90 (65.85) | 255.40 (67.3) | 256.30 (86.05) | 291.10 (56.29) | NA | 299.90 (53.22) | 289.50 (42.17) | 296.90 (38.99) | 328.30 (32.76) | 305.90 (54.93) | 313.80 (47.10) | 282.30 (61.71) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dimitrov, K.M.; Taylor, T.L.; Marcano, V.C.; Williams-Coplin, D.; Olivier, T.L.; Yu, Q.; Gogal, R.M., Jr.; Suarez, D.L.; Afonso, C.L. Novel Recombinant Newcastle Disease Virus-Based In Ovo Vaccines Bypass Maternal Immunity to Provide Full Protection from Early Virulent Challenge. Vaccines 2021, 9, 1189. https://doi.org/10.3390/vaccines9101189
Dimitrov KM, Taylor TL, Marcano VC, Williams-Coplin D, Olivier TL, Yu Q, Gogal RM Jr., Suarez DL, Afonso CL. Novel Recombinant Newcastle Disease Virus-Based In Ovo Vaccines Bypass Maternal Immunity to Provide Full Protection from Early Virulent Challenge. Vaccines. 2021; 9(10):1189. https://doi.org/10.3390/vaccines9101189
Chicago/Turabian StyleDimitrov, Kiril M., Tonya L. Taylor, Valerie C. Marcano, Dawn Williams-Coplin, Timothy L. Olivier, Qingzhong Yu, Robert M. Gogal, Jr., David L. Suarez, and Claudio L. Afonso. 2021. "Novel Recombinant Newcastle Disease Virus-Based In Ovo Vaccines Bypass Maternal Immunity to Provide Full Protection from Early Virulent Challenge" Vaccines 9, no. 10: 1189. https://doi.org/10.3390/vaccines9101189