Environmental Risk Assessment for rVSVΔG-ZEBOV-GP, a Genetically Modified Live Vaccine for Ebola Virus Disease

 ,
,
Abstract
:1. Introduction
2. Environmental Assessment Requirements for GMO Vaccines
- Potential effects on the quality of the environment are likely to be highly controversial;
- Potential effects on human health are highly uncertain or involve unique or unknown risks;
- May have potential effects on an endangered/threatened species or its habitat;
- Potential effects may violate federal, state, or local laws or requirements imposed for the protection of the environment.
3. Developing an ERA for rVSVΔG-ZEBOV-GP
3.1. ERA Overview
- Identify any adverse environmental effects that the GMO may cause. In the case of a recombinant virus vaccine, this step should consider the vaccines’ host range, cell and tissue tropism (especially if genes involved in cellular entry are altered, such as with rVSVΔG-ZEBOV-GP), transmission route, infectivity, pathogenicity, replication mechanism, genetic stability, ability to transfer genetic materials to other organisms, and survivability. For this step, it makes most sense to first assess the relevant characteristics of the wild-type virus vector, and then consider and experimentally evaluate how those might be altered by the foreign genetic material [74].
- Evaluate the potential consequences of each of the identified adverse environmental effects, should it occur. For each of the adverse effects identified in the previous step, the extent of its negative impacts should be classified as high (i.e., significant changes that might affect ecosystem function), moderate, low, or negligible (i.e., no significant changes) on each potentially affected species, ecosystem, or the overall environment [74].
- Evaluate the likelihood of each identified adverse environmental effect to occur. The likelihood of most negative environmental impacts is difficult, even impossible, to quantify. The classification of likelihoods as high, moderate, low, or negligible is thus useful again. Alternatively, in the previous step, a worst-case scenario could be considered—if the consequences of that scenario can very conservatively be considered as acceptable, then actual quantification in step 3 may not be necessary. For recombinant virus vaccines, preclinical and clinical shedding studies of sufficiently long duration (especially in the case of replication-competent vaccines, such as rVSVΔG-ZEBOV-GP) should be conducted. Vaccine tropism should also be evaluated, which is typically done through in vitro methods assessing the vaccine’s ability to infect different cell types and/or a biodistribution study that quantifies vaccine virus in tissue samples by PCR methods; infectivity assays, such as plaque or TCID50 assays, are then employed to determine if observed virus is replicative [74].
- Estimate the risk posed by each adverse environmental effect. This step collates the information on magnitude and likelihood of each potential environmental hazard, perhaps in a risk matrix [74]. It is recommended that the risk be regarded as high if there is uncertainty around its likelihood or its consequences [74].
- Define mitigation strategies to minimize all of the risks associated with the GMO. Mitigation strategies must be defined, at the very least, for all environmental risks deemed as unacceptable in the previous step. Most mitigation measures will seek to minimize the likelihood of negative environmental effects. In the case of live recombinant vaccines, strategies may include adequate disinfection and disposal of materials used in the vaccine-to-recipient supply chain or during actual vaccine administration, protecting the injection site from contact with the environment, and even collection and disposal of the vaccine recipients’ bodily waste. A plan to monitor the effectiveness of mitigation strategies and help identify further, unanticipated hazards should also be developed [74].
- Determine the GMO’s overall environmental risk. The final step seeks to assess the overall risk by considering the totality of risks posed by each individual environmental hazard in light of the proposed risk management strategies and whether this overall risk profile is acceptable or not [74].
3.2. Environmental Risk Potential
3.2.1. Overview of Potential Environmental Issues with rVSVΔG-ZEBOV-GP
- Could rVSVΔG-ZEBOV-GP cause disease in humans, animals, and/or plants, including allergic or toxic effects?
- Could rVSVΔG-ZEBOV-GP have effects on the dynamics of populations of species in the receiving environment and the genetic diversity of these populations?
- Could rVSVΔG-ZEBOV-GP alter susceptibility of any organism to pathogens, thus facilitating the dissemination of infectious diseases and/or creating new reservoirs or vectors?
- Could rVSVΔG-ZEBOV-GP compromise any prophylactic or therapeutic medical, veterinary, or plant protection treatments?
- Could rVSVΔG-ZEBOV-GP have effects on biogeochemical cycles, particularly carbon and nitrogen recycling through changes in soil decomposition of organic material?
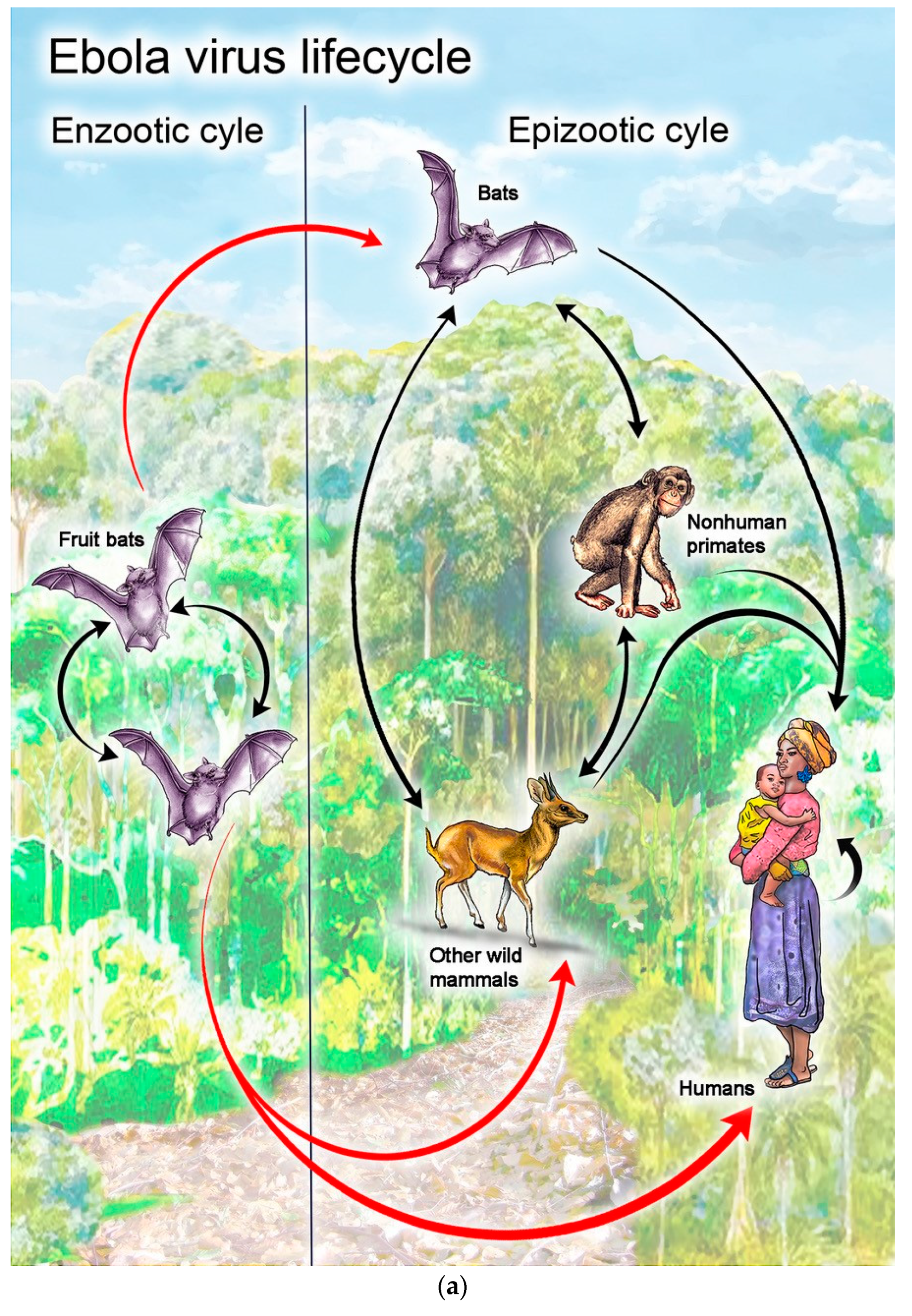
3.2.2. Potential to Cause Disease
3.2.3. Potential to Affect Populations and Genetic Diversity
3.2.4. Other Potential Effects
3.3. Exposure Risk Estimation and Risk Mitigation Strategies
3.4. Considering Environmental Risks vs. Health Benefits
4. Applying the Lessons Learned
4.1. Accelerating the Development of Live Recombinant Vaccines Overall
- Robust viral shedding and viremia data, generated from clinical and non-clinical studies using qualified methods that are applied consistently across clinical trials for the candidate vaccine. These data are essential from vaccine recipients and are also important from other species potentially susceptible to infection with the recombinant vaccine.
- Experiments that evaluate the likelihood for replication in potentially susceptible species and for transmission by biological and/or mechanical vector species.
- Studies on genetic stability of the vaccine virus and the potential for recombination with other viruses.
- Robust protocols for vaccine storage, transport/distribution, administration, safe handling, and disinfection/decontamination, all with the goal of preventing (a) contact with the vaccine (other than actual inoculation) and (b) release of the vaccine into the environment.
- Appropriate risk mitigation strategies.
- Knowledge of each country’s expectations and guidelines for relevant environmental assessments and having discussions with regulatory agencies prior to filing, in order to make sure that expectations will be met.
- Proper presentation of the environmental assessment with relevant appendices following applicable country guidelines.
- The likely requirement of detailed information on vaccine packaging and transport.
- The likely requirement of detailed handling instructions for the vaccine.
- The environmental assessment should be fully aligned with the proposed product label for the vaccine.
4.2. Accelerating the Development of rVSV-Based Vaccines Specifically
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jacob, S.T.; Crozier, I.; Fischer, W.A., 2nd; Hewlett, A.; Kraft, C.S.; Vega, M.A.; Soka, M.J.; Wahl, V.; Griffiths, A.; Bollinger, L.; et al. Ebola Virus Disease. Nat. Rev. Dis. Primers 2020, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Canady, D.; Weil, N.C.; Miller, C.; Shantha, J.G.; Bastien, G.; Yeh, S. Ophthalmic and Psychosocial Sequelae in Ebola Virus Disease Survivors: Ongoing Need for Health Systems Strengthening across Disciplines. Expert Rev. Anti. Infect. Ther. 2020, 1–3. [Google Scholar] [CrossRef]
- Etard, J.F.; Sow, M.S.; Leroy, S.; Toure, A.; Taverne, B.; Keita, A.K.; Msellati, P.; Magassouba, N.; Baize, S.; Raoul, H.; et al. Multidisciplinary Assessment of Post-Ebola Sequelae in Guinea (Postebogui): An Observational Cohort Study. Lancet Infect. Dis. 2017, 17, 545–552. [Google Scholar] [CrossRef] [Green Version]
- Mattia, J.G.; Vandy, M.J.; Chang, J.C.; Platt, D.E.; Dierberg, K.; Bausch, D.G.; Brooks, T.; Conteh, S.; Crozier, I.; Fowler, R.A.; et al. Early Clinical Sequelae of Ebola Virus Disease in Sierra Leone: A Cross-Sectional Study. Lancet Infect. Dis. 2016, 16, 331–338. [Google Scholar] [CrossRef] [Green Version]
- Wiedemann, A.; Foucat, E.; Hocini, H.; Lefebvre, C.; Hejblum, B.P.; Durand, M.; Kruger, M.; Keita, A.K.; Ayouba, A.; Mely, S.; et al. Long-Lasting Severe Immune Dysfunction in Ebola Virus Disease Survivors. Nat. Commun. 2020, 11, 3730. [Google Scholar] [CrossRef]
- Furuyama, W.; Marzi, A. Ebola Virus: Pathogenesis and Countermeasure Development. Annu. Rev. Virol. 2019, 6, 435–458. [Google Scholar] [PubMed]
- Patel, P.R.; Shah, S. Ebola Virus. In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- World Health Organization. Ebola Virus Disease. Available online: https://www.who.int/news-room/fact-sheets/detail/ebola-virus-disease (accessed on 20 August 2020).
- Gumusova, S.; Sunbul, M.; Leblebicioglu, H. Ebola Virus Disease and the Veterinary Perspective. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koch, L.K.; Cunze, S.; Kochmann, J.; Klimpel, S. Bats as Putative Zaire Ebolavirus Reservoir Hosts and Their Habitat Suitability in Africa. Sci. Rep. 2020, 10, 14268. [Google Scholar] [CrossRef] [PubMed]
- Carette, J.E.; Raaben, M.; Wong, A.C.; Herbert, A.S.; Obernosterer, G.; Mulherkar, N.; Kuehne, A.I.; Kranzusch, P.J.; Griffin, A.M.; Ruthel, G.; et al. Ebola Virus Entry Requires the Cholesterol Transporter Niemann-Pick C1. Nature 2011, 477, 340–343. [Google Scholar] [PubMed] [Green Version]
- Cote, M.; Misasi, J.; Ren, T.; Bruchez, A.; Lee, K.; Filone, C.M.; Hensley, L.; Li, Q.; Ory, D.; Chandran, K.; et al. Small Molecule Inhibitors Reveal Niemann-Pick C1 Is Essential for Ebola Virus Infection. Nature 2011, 477, 344–348. [Google Scholar] [CrossRef]
- Das, D.K.; Bulow, U.; Diehl, W.E.; Durham, N.D.; Senjobe, F.; Chandran, K.; Luban, J.; Munro, J.B. Conformational Changes in the Ebola Virus Membrane Fusion Machine Induced by Ph, Ca2+, and Receptor Binding. PLoS Biol. 2020, 18, e3000626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Banadyga, L.; Emeterio, K.; Wong, G.; Qiu, X. The Roles of Ebola Virus Soluble Glycoprotein in Replication, Pathogenesis, and Countermeasure Development. Viruses 2019, 11, 999. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozo-Lopez, P.; Drolet, B.S.; Londono-Renteria, B. Vesicular Stomatitis Virus Transmission: A Comparison of Incriminated Vectors. Insects 2018, 9, 190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jimenez, A.E.; Jimenez, C.; Castro, L.; Rodriguez, L. Serological Survey of Small Mammals in a Vesicular Stomatitis Virus Enzootic Area. J. Wildl. Dis. 1996, 32, 274–279. [Google Scholar] [CrossRef] [Green Version]
- Jimenez, A.E.; Herrera, F.V.; Salman, M.; Herrero, M.V. Survey of Small Rodents and Hematophagous Flies in Three Sentinel Farms in a Costa Rican Vesicular Stomatitis Endemic Region. Ann. N. Y. Acad. Sci. 2000, 916, 453–463. [Google Scholar] [CrossRef]
- Aguirre, A.A.; McLean, R.G.; Cook, R.S.; Quan, T.J. Serologic Survey for Selected Arboviruses and Other Potential Pathogens in Wildlife from Mexico. J. Wildl. Dis. 1992, 28, 435–442. [Google Scholar] [CrossRef]
- Tesh, R.B.; Boshell, J.; Modi, G.B.; Morales, A.; Young, D.G.; Corredor, A.; de Carrasquilla, C.F.; de Rodriguez, C.; Walters, L.L.; Gaitan, M.O. Natural Infection of Humans, Animals, and Phlebotomine Sand Flies with the Alagoas Serotype of Vesicular Stomatitis Virus in Colombia. Am. J. Trop. Med. Hyg. 1987, 36, 653–661. [Google Scholar] [CrossRef]
- Tesh, R.B.; Peralta, P.H.; Johnson, K.M. Ecologic Studies of Vesicular Stomatitis Virus. I. Prevalence of Infection among Animals and Humans Living in an Area of Endemic VSV Activity. Am. J. Epidemiol. 1969, 90, 255–261. [Google Scholar]
- Tesh, R.B.; Peralta, P.H.; Johnson, K.M. Ecologic Studies of Vesicular Stomatitis Virus. Ii. Results of Experimental Infection in Panamanian Wild Animals. Am. J. Epidemiol. 1970, 91, 216–224. [Google Scholar] [CrossRef]
- Comer, J.A.; Tesh, R.B.; Modi, G.B.; Corn, J.L.; Nettles, V.F. Vesicular Stomatitis Virus, New Jersey Serotype: Replication in and Transmission by Lutzomyia Shannoni (Diptera: Psychodidae). Am. J. Trop. Med. Hyg. 1990, 42, 483–490. [Google Scholar] [CrossRef]
- Sudia, W.D.; Fields, B.N.; Calisher, C.H. The Isolation of Vesiculay Stomatitis Virus (Indiana Strain) and Other Viruses from Mosquitoes in New Mexico, 1965. Am. J. Epidemiol. 1967, 86, 598–602. [Google Scholar] [CrossRef] [PubMed]
- Drolet, B.S.; Campbell, C.L.; Stuart, M.A.; Wilson, W.C. Vector Competence of Culicoides Sonorensis (Diptera: Ceratopogonidae) for Vesicular Stomatitis Virus. J. Med. Entomol. 2005, 42, 409–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drolet, B.S.; Stuart, M.A.; Derner, J.D. Infection of Melanoplus Sanguinipes Grasshoppers Following Ingestion of Rangeland Plant Species Harboring Vesicular Stomatitis Virus. Appl. Environ. Microbiol. 2009, 75, 3029–3033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cupp, E.W.; Mare, C.J.; Cupp, M.S.; Ramberg, F.B. Biological Transmission of Vesicular Stomatitis Virus (New Jersey) by Simulium Vittatum (Diptera: Simuliidae). J. Med. Entomol. 1992, 29, 137–140. [Google Scholar] [CrossRef]
- Francy, D.B.; Moore, C.G.; Smith, G.C.; Jakob, W.L.; Taylor, S.A.; Calisher, C.H. Epizootic Vesicular Stomatitis in Colorado, 1982: Isolation of Virus from Insects Collected Along the Northern Colorado Rocky Mountain Front Range. J. Med. Entomol. 1988, 25, 343–347. [Google Scholar] [CrossRef] [Green Version]
- Tesh, R.B.; Chaniotis, B.N.; Peralta, P.H.; Johnson, K.M. Ecology of Viruses Isolated from Panamanian Phlebotomine Sandflies. Am. J. Trop. Med. Hyg. 1974, 23, 258–269. [Google Scholar] [CrossRef]
- Cornish, T.E.; Stallknecht, D.E.; Brown, C.C.; Seal, B.S.; Howerth, E.W. Pathogenesis of Experimental Vesicular Stomatitis Virus (New Jersey Serotype) Infection in the Deer Mouse (Peromyscus Maniculatus). Vet. Pathol. 2001, 38, 396–406. [Google Scholar] [CrossRef]
- Mesquita, L.P.; Diaz, M.H.; Howerth, E.W.; Stallknecht, D.E.; Noblet, R.; Gray, E.W.; Mead, D.G. Pathogenesis of Vesicular Stomatitis New Jersey Virus Infection in Deer Mice ( Peromyscus Maniculatus) Transmitted by Black Flies ( Simulium Vittatum). Vet. Pathol. 2017, 54, 74–81. [Google Scholar] [CrossRef]
- Mead, D.G.; Ramberg, F.B.; Besselsen, D.G.; Mare, C.J. Transmission of Vesicular Stomatitis Virus from Infected to Noninfected Black Flies Co-Feeding on Nonviremic Deer Mice. Science 2000, 287, 485–487. [Google Scholar]
- Rozo-Lopez, P.; Londono-Renteria, B.; Drolet, B.S. Venereal Transmission of Vesicular Stomatitis Virus by Culicoides Sonorensis Midges. Pathogens 2020, 9, 316. [Google Scholar] [CrossRef]
- Ervebo® (Ebola Zaire Vaccine, Live) Suspension for Intramuscular Injection Prescribing Information; Merck Sharp &Dohme Corp. (Ed.) Subsidiary of Merck & Co., Inc.: Whitehouse Station, NJ, USA, 2019. [Google Scholar]
- Coller, B.G.; Blue, J.; Das, R.; Dubey, S.; Finelli, L.; Gupta, S.; Helmond, F.; Grant-Klein, R.J.; Liu, K.; Simon, J.; et al. Clinical Development of a Recombinant Ebola Vaccine in the Midst of an Unprecedented Epidemic. Vaccine 2017, 35 Pt A, 4465–4469. [Google Scholar] [CrossRef]
- International Federation of Pharmaceutical Manufacturers & Associations. The Complex Journey of a Vaccine. The Steps behind Developing a New Vaccine. IFMPA. Available online: https://www.ifpma.org/wp-content/uploads/2019/07/IFPMA-ComplexJourney-2019_FINAL.pdf (accessed on 22 August 2020).
- Halperin, S.A.; Arribas, J.R.; Rupp, R.; Andrews, C.P.; Chu, L.; Das, R.; Simon, J.K.; Onorato, M.T.; Liu, K.; Martin, J.; et al. Six-Month Safety Data of Recombinant Vesicular Stomatitis Virus-Zaire Ebola Virus Envelope Glycoprotein Vaccine in a Phase 3 Double-Blind, Placebo-Controlled Randomized Study in Healthy Adults. J. Infect. Dis. 2017, 215, 1789–1798. [Google Scholar] [CrossRef] [PubMed]
- Halperin, S.A.; Das, R.; Onorato, M.T.; Liu, K.; Martin, J.; Grant-Klein, R.J.; Nichols, R.; Coller, B.-A.; Helmond, F.A.; Simon, J.K.; et al. Immunogenicity, Lot Consistency, and Extended Safety of Recombinant Vesicular Stomatitis Virus–Zaire Ebola Virus Envelope Glycoprotein Vaccine: A Phase 3 Randomized, Double-Blind, Placebo-Controlled Study in Healthy Adults. J. Infect. Dis. 2019, 220, 1127–1136. [Google Scholar] [CrossRef] [PubMed]
- Samai, M.; Seward, J.F.; Goldstein, S.T.; Mahon, B.E.; Lisk, D.R.; Widdowson, M.A.; Jalloh, M.I.; Schrag, S.J.; Idriss, A.; Carter, R.J.; et al. The Sierra Leone Trial to Introduce a Vaccine against Ebola: An Evaluation of rVSVΔG-ZEBOV-GP Vaccine Tolerability and Safety during the West Africa Ebola Outbreak. J. Infect. Dis. 2018, 217 (Suppl. 1), S6–S15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, S.B.; Bolay, F.; Kieh, M.; Grandits, G.; Badio, M.; Ballou, R.; Eckes, R.; Feinberg, M.; Follmann, D.; Grund, B.; et al. Phase 2 Placebo-Controlled Trial of Two Vaccines to Prevent Ebola in Liberia. N. Engl. J. Med. 2017, 377, 1438–1447. [Google Scholar] [CrossRef] [Green Version]
- Henao-Restrepo, A.M.; Longini, I.M.; Egger, M.; Dean, N.E.; Edmunds, W.J.; Camacho, A.; Carroll, M.W.; Doumbia, M.; Draguez, B.; Duraffour, S.; et al. Efficacy and Effectiveness of an rVSV-Vectored Vaccine Expressing Ebola Surface Glycoprotein: Interim Results from the Guinea Ring Vaccination Cluster-Randomised Trial. Lancet 2015, 386, 857–866. [Google Scholar] [CrossRef]
- Monath, T.P.; Fast, P.E.; Modjarrad, K.; Clarke, D.K.; Martin, B.K.; Fusco, J.; Nichols, R.; Heppner, D.G.; Simon, J.K.; Dubey, S.; et al. rVSVΔG-ZEBOV-GP (Also Designated V920) Recombinant Vesicular Stomatitis Virus Pseudotyped with Ebola Zaire Glycoprotein: Standardized Template with Key Considerations for a Risk/Benefit Assessment. Vaccine X 2019, 1, 100009. [Google Scholar] [CrossRef]
- Huttner, A.; Agnandji, S.T.; Combescure, C.; Fernandes, J.F.; Bache, E.B.; Kabwende, L.; Ndungu, F.M.; Brosnahan, J.; Monath, T.P.; Lemaitre, B.; et al. Determinants of Antibody Persistence across Doses and Continents after Single-Dose -Zebov Vaccination for Ebola Virus Disease: An Observational Cohort Study. Lancet Infect. Dis. 2018, 18, 738–748. [Google Scholar]
- Huttner, A.; Combescure, C.; Grillet, S.; Haks, M.C.; Quinten, E.; Modoux, C.; Agnandji, S.T.; Brosnahan, J.; Dayer, J.A.; Harandi, A.M.; et al. A Dose-Dependent Plasma Signature of the Safety and Immunogenicity of the rVSV-Ebola Vaccine in Europe and Africa. Sci. Transl. Med. 2017, 9, eaaj1701. [Google Scholar] [CrossRef] [Green Version]
- Huttner, A.; Dayer, J.A.; Yerly, S.; Combescure, C.; Auderset, F.; Desmeules, J.; Eickmann, M.; Finckh, A.; Goncalves, A.R.; Hooper, J.W.; et al. The Effect of Dose on the Safety and Immunogenicity of the VSV Ebola Candidate Vaccine: A Randomised Double-Blind, Placebo-Controlled Phase 1/2 Trial. Lancet Infect. Dis. 2015, 15, 1156–1166. [Google Scholar] [CrossRef]
- Huttner, A.; Siegrist, C.A. Durability of Single-Dose rVSV-Zebov Vaccine Responses: What Do We Know? Expert. Rev. Vaccines 2018, 17, 1105–1110. [Google Scholar] [CrossRef] [PubMed]
- ElSherif, M.S.; Brown, C.; MacKinnon-Cameron, D.; Li, L.; Racine, T.; Alimonti, J.; Rudge, T.L.; Sabourin, C.; Silvera, P.; Hooper, J.W.; et al. Assessing the Safety and Immunogenicity of Recombinant Vesicular Stomatitis Virus Ebola Vaccine in Healthy Adults: A Randomized Clinical Trial. CMAJ 2017, 189, E819–E827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heppner, D.G., Jr.; Kemp, T.L.; Martin, B.K.; Ramsey, W.J.; Nichols, R.; Dasen, E.J.; Link, C.J.; Das, R.; Xu, Z.J.; Sheldon, E.A.; et al. Safety and Immunogenicity of the rVSV-Zebov-GP Ebola Virus Vaccine Candidate in Healthy Adults: A Phase 1b Randomised, Multicentre, Double-Blind, Placebo-Controlled, Dose-Response Study. Lancet Infect. Dis. 2017, 17, 854–866. [Google Scholar] [CrossRef] [Green Version]
- Simon, J.K.; Halperin, S.A.; Das, R.; Onorato, M.T.; Liu, K.; Martin, J.; Grant-Klein, R.J.; Nichols, R.; Helmond, F.A.; Coller, B.-A.; et al. Clinical Trial Experience with the Merck Rvsvδg-Zebov-Gp Ebola Vaccine: Updated Safety, Immunogenicity, and Efficacy. Paper Presented at the American Society of Tropical Medicine and Hygiene, New Orleans, LA, USA, 28 October–1 November 2018. [Google Scholar]
- Cobleigh, M.A.; Bradfield, C.; Liu, Y.; Mehta, A.; Robek, M.D. The Immune Response to a Vesicular Stomatitis Virus Vaccine Vector Is Independent of Particulate Antigen Secretion and Protein Turnover Rate. J. Virol. 2012, 86, 4253–4261. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clarke, D.K.; Hendry, R.M.; Singh, V.; Rose, J.K.; Seligman, S.J.; Klug, B.; Kochhar, S.; Mac, L.M.; Carbery, B.; Chen, R.T.; et al. Live Virus Vaccines Based on a Vesicular Stomatitis Virus (VSV) Backbone: Standardized Template with Key Considerations for a Risk/Benefit Assessment. Vaccine 2016, 34, 6597–6609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Letchworth, G.J.; Rodriguez, L.L.; Del Cbarrera, J. Vesicular Stomatitis. Vet. J. 1999, 157, 239–260. [Google Scholar] [CrossRef]
- Bram, R.A.; George, J.E.; Reichar, R.E.; Tabaciinic, W.J. Threat of Foreign Arthropod-Borne Pathogens to Livestock in the United States. J. Med. Entomol. 2002, 39, 405–416. [Google Scholar] [CrossRef]
- Rodriguez, L.L. Emergence and Re-Emergence of Vesicular Stomatitis in the United States. Virus Res. 2002, 85, 211–219. [Google Scholar] [CrossRef]
- Goodger, W.J.; Thurmond, M.; Nehay, J.; Mitchell, J.; Smith, P. Economic Impact of an Epizootic of Bovine Vesicular Stomatitis in California. J. Am. Vet. Med. Assoc. 1985, 186, 370–373. [Google Scholar]
- Hayek, A.M.; McCluskey, B.J.; Chavez, G.T.; Salman, M.D. Financial Impact of the 1995 Outbreak of Vesicular Stomatitis on 16 Beef Ranches in Colorado. J. Am. Vet. Med. Assoc. 1998, 212, 820–823. [Google Scholar]
- Awad, S.S.; Rodriguez, A.H.; Chuang, Y.C.; Marjanek, Z.; Pareigis, A.J.; Reis, G.; Scheeren, T.W.; Sanchez, A.S.; Zhou, X.; Saulay, M.; et al. A Phase 3 Randomized Double-Blind Comparison of Ceftobiprole Medocaril versus Ceftazidime Plus Linezolid for the Treatment of Hospital-Acquired Pneumonia. Clin. Infect. Dis. 2014, 59, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Lyles, D.S.; Rupprecht, C.E. Rhabdoviridae. In Fields Virology; Peter, K.M., David, H.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; pp. 1364–1408. [Google Scholar]
- Stallknecht, D.E.; Perzak, D.E.; Bauer, L.D.; Murphy, M.D.; Howerth, E.W. Contact Transmission of Vesicular Stomatitis Virus New Jersey in Pigs. Am. J. Vet. Res. 2001, 62, 516–520. [Google Scholar] [CrossRef] [PubMed]
- Martinez, I.; Wertz, G.W. Biological Differences between Vesicular Stomatitis Virus Indiana and New Jersey Serotype Glycoproteins: Identification of Amino Acid Residues Modulating Ph-Dependent Infectivity. J. Virol. 2005, 79, 3578–3585. [Google Scholar] [CrossRef] [Green Version]
- Ci, Y.; Yang, Y.; Xu, C.; Shi, L. Vesicular Stomatitis Virus G Protein Transmembrane Region Is Crucial for the Hemi-Fusion to Full Fusion Transition. Sci. Rep. 2018, 8, 10669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roche, S.; Rey, F.A.; Gaudin, Y.; Bressanelli, S. Structure of the Prefusion Form of the Vesicular Stomatitis Virus Glycoprotein G. Science 2007, 315, 843–848. [Google Scholar] [CrossRef] [Green Version]
- Finkelshtein, D.; Werman, A.; Novick, D.; Barak, S.; Rubinstein, M. Ldl Receptor and Its Family Members Serve as the Cellular Receptors for Vesicular Stomatitis Virus. Proc. Natl. Acad. Sci. USA 2013, 110, 7306–7311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Geisbert, T.W.; Daddario-Dicaprio, K.M.; Lewis, M.G.; Geisbert, J.B.; Grolla, A.; Leung, A.; Paragas, J.; Matthias, L.; Smith, M.A.; Jones, S.M.; et al. Vesicular Stomatitis Virus-Based Ebola Vaccine Is Well-Tolerated and Protects Immunocompromised Nonhuman Primates. PLoS Pathog. 2008, 4, e1000225. [Google Scholar]
- Garbutt, M.; Liebscher, R.; Wahl-Jensen, V.; Jones, S.; Moller, P.; Wagner, R.; Volchkov, V.; Klenk, H.D.; Feldmann, H.; Stroher, U. Properties of Replication-Competent Vesicular Stomatitis Virus Vectors Expressing Glycoproteins of Filoviruses and Arenaviruses. J. Virol. 2004, 78, 5458–5465. [Google Scholar] [CrossRef] [Green Version]
- Jones, S.M.; Feldmann, H.; Stroher, U.; Geisbert, J.B.; Fernando, L.; Grolla, A.; Klenk, H.D.; Sullivan, N.J.; Volchkov, V.E.; Fritz, E.A.; et al. Live Attenuated Recombinant Vaccine Protects Nonhuman Primates against Ebola and Marburg Viruses. Nat. Med. 2005, 11, 786–790. [Google Scholar] [CrossRef]
- Zimmer, B.; Summermatter, K.; Zimmer, G. Stability and Inactivation of Vesicular Stomatitis Virus, a Prototype Rhabdovirus. Vet. Microbiol. 2013, 162, 78–84. [Google Scholar] [CrossRef]
- Geisbert, T.W.; Feldmann, H. Recombinant Vesicular Stomatitis Virus-Based Vaccines against Ebola and Marburg Virus Infections. J. Infect. Dis. 2011, 204 (Suppl. 3), S1075–S1081. [Google Scholar] [CrossRef] [PubMed]
- Bergren, N.A.; Miller, M.R.; Monath, T.P.; Kading, R.C. Assessment of the Ability of V920 Recombinant Vesicular Stomatitis-Zaire Ebolavirus Vaccine to Replicate in Relevant Arthropod Cell Cultures and Vector Species. Hum. Vaccines Immunother. 2018, 14, 994–1002. [Google Scholar] [CrossRef] [Green Version]
- Nunamaker, R.A.; Lockwood, J.A.; Stith, C.E.; Campbell, C.L.; Schell, S.P.; Drolet, B.S.; Wilson, W.C.; White, D.M.; Letchworth, G.J. Grasshoppers (Orthoptera: Acrididae) Could Serve as Reservoirs and Vectors of Vesicular Stomatitis Virus. J. Med. Entomol. 2003, 40, 957–963. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferris, D.H.; Hanson, R.P.; Dicke, R.J.; Roberts, R.H. Experimental Transmission of Vesicular Stomatitis Virus by Diptera. J. Infect. Dis. 1955, 96, 184–192. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food and Drug Administration. Determining the Need for and Content of Environmental Assessments for Gene Therapies, Vectored Vaccines, and Related Recombinant Viral or Microbial Products; US Food and Drug Administration: Silver Spring, MD, USA, 2015. [Google Scholar]
- Nakayama, Y.; Aruga, A. Comparison of Current Regulatory Status for Gene-Based Vaccines in the U.S., Europe and Japan. Vaccines 2015, 3, 186–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- European Medicines Agency. Guideline on Environmental Risk Assessments for Medicinal Products Consisting of, or Containing, Genetically Modified Organisms; EMA: London, UK, 2006. [Google Scholar]
- Anliker, B.; Longhurst, S.; Buchholz, C.J. Environmental Risk Assessment for Medicinal Products Containing Genetically Modified Organisms. Bundesgesundheitsblatt Gesundheitsforschung Gesundheitsschutz 2010, 53, 52–57. [Google Scholar] [CrossRef]
- Regules, J.A.; Beigel, J.H.; Paolino, K.M.; Voell, J.; Castellano, A.R.; Hu, Z.; Munoz, P.; Moon, J.E.; Ruck, R.C.; Bennett, J.W.; et al. A Recombinant Vesicular Stomatitis Virus Ebola Vaccine. N. Engl. J. Med. 2017, 376, 330–341. [Google Scholar] [CrossRef]
- Agnandji, S.T.; Huttner, A.; Zinser, M.E.; Njuguna, P.; Dahlke, C.; Fernandes, J.F.; Yerly, S.; Dayer, J.A.; Kraehling, V.; Kasonta, R.; et al. Phase 1 Trials of rVSV Ebola Vaccine in Africa and Europe. N. Engl. J. Med. 2016, 374, 1647–1660. [Google Scholar]
- Agnandji, S.T.; Fernandes, J.F.; Bache, E.B.; Mba, R.M.O.; Brosnahan, J.S.; Kabwende, L.; Pitzinger, P.; Staarink, P.; Massinga-Loembe, M.; Krahling, V.; et al. Safety and Immunogenicity of rVSVΔG-ZEBOV-GP Ebola Vaccine in Adults and Children in Lambarene, Gabon: A Phase I Randomised Trial. PLoS Med. 2017, 14, e1002402. [Google Scholar] [CrossRef] [Green Version]
- De Wit, E.; Marzi, A.; Bushmaker, T.; Brining, D.; Scott, D.; Richt, J.A.; Geisbert, T.W.; Feldmann, H. Safety of Recombinant Vsv-Ebola Virus Vaccine Vector in Pigs. Emerg. Infect. Dis. 2015, 21, 702–704. [Google Scholar] [CrossRef]
- Jones, S.M.; Stroher, U.; Fernando, L.; Qiu, X.; Alimonti, J.; Melito, P.; Bray, M.; Klenk, H.D.; Feldmann, H. Assessment of a Vesicular Stomatitis Virus-Based Vaccine by Use of the Mouse Model of Ebola Virus Hemorrhagic Fever. J. Infect. Dis. 2007, 196 (Suppl. 2), S404–S412. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, Y.; Safronetz, D.; Brown, K.; LaCasse, R.; Marzi, A.; Ebihara, H.; Feldmann, H. Protective Efficacy of a Bivalent Recombinant Vesicular Stomatitis Virus Vaccine in the Syrian Hamster Model of Lethal Ebola Virus Infection. J. Infect. Dis. 2011, 2011 (Suppl. 3), S1090–S1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mire, C.E.; Miller, A.D.; Carville, A.; Westmoreland, S.V.; Geisbert, J.B.; Mansfield, K.G.; Feldmann, H.; Hensley, L.E.; Geisbert, T.W. Recombinant Vesicular Stomatitis Virus Vaccine Vectors Expressing Filovirus Glycoproteins Lack Neurovirulence in Nonhuman Primates. PLoS Negl. Trop. Dis. 2012, 6, e1567. [Google Scholar] [CrossRef] [PubMed]
- Barber, G.N. Vesicular Stomatitis Virus as an Oncolytic Vector. Viral. Immunol. 2004, 17, 516–527. [Google Scholar] [CrossRef]
- Obuchi, M.; Fernandez, M.; Barber, G.N. Development of Recombinant Vesicular Stomatitis Viruses That Exploit Defects in Host Defense to Augment Specific Oncolytic Activity. J. Virol. 2003, 77, 8843–8856. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. Assessment Report. Ervebo. Common Name: Ebola Zaire Vaccine (rVSV∆G-ZEBOV-GP, Live); European Medicines Agency: Amsterdam, The Netherlands, 2019; Available online: https://www.ema.europa.eu/en/documents/assessment-report/ervebo-epar-public-assessment-report_en.pdf (accessed on 14 December 2020).
- Quiñones-Kochs, M.I.; Schnell, M.J.; Buonocore, L.; Rose, J.K. Mechanisms of loss of foreign gene expression in recombinant vesicular stomatitis viruses. Virology 2001, 287, 427–435. [Google Scholar] [CrossRef] [Green Version]
- Sanjuan, R.; Domingo-Calap, P. Mechanisms of Viral Mutation. Cell Mol. Life Sci. 2016, 73, 4433–4448. [Google Scholar] [CrossRef] [Green Version]
- Chare, E.R.; Gould, E.A.; Holmes, E.C. Phylogenetic Analysis Reveals a Low Rate of Homologous Recombination in Negative-Sense Rna Viruses. J. Gen. Virol. 2003, 84 Pt 10, 2691–2703. [Google Scholar] [CrossRef]
- Simon-Loriere, E.; Holmes, E.C. Why Do Rna Viruses Recombine? Nat. Rev. Microbiol. 2011, 9, 617–626. [Google Scholar] [CrossRef]
- Marzi, A.; Ebihara, H.; Callison, J.; Groseth, A.; Williams, K.J.; Geisbert, T.W.; Feldmann, H. Vesicular Stomatitis Virus-Based Ebola Vaccines with Improved Cross-Protective Efficacy. J. Infect. Dis. 2011, 2011 (Suppl. 3), S1066–S1074. [Google Scholar]
- Condit, R.C.; Williamson, A.L.; Sheets, R.; Seligman, S.J.; Monath, T.P.; Excler, J.L.; Gurwith, M.; Bok, K.; Robertson, J.S.; Kim, D.; et al. Unique Safety Issues Associated with Virus-Vectored Vaccines: Potential for and Theoretical Consequences of Recombination with Wild Type Virus Strains. Vaccine 2016, 34, 6610–6616. [Google Scholar] [CrossRef] [PubMed]
- U.S. Food & Drug Administration. Ervebo. Available online: https://www.fda.gov/vaccines-blood-biologics/ervebo (accessed on 2 October 2020).
- Wolf, J.; Bruno, S.; Eichberg, M.; Jannat, R.; Rudo, S.; VanRheenen, S.; Coller, B.A. Applying Lessons from the Ebola Vaccine Experience for Sars-Cov-2 and Other Epidemic Pathogens. NPJ Vaccines 2020, 5, 51. [Google Scholar] [CrossRef] [PubMed]
- Kaur, S.P.; Gupta, V. Covid-19 Vaccine: A Comprehensive Status Report. Virus Res. 2020, 288, 198114. [Google Scholar] [CrossRef] [PubMed]
- The European Parliament and the Council of the European Union. Regulation (Eu) 2020/1043 of the European Parliament and of the Council of 15 July 2020 on the Conduct of Clinical Trials with and Supply of Medicinal Products for Human Use Containing or Consisting of Genetically Modified Organisms Intended to Treat or Prevent Coronavirus Disease (Covid-19); The European Parliament and the Council of the European Union, Ed.; Official Journal of the European Union: Brussels, Belgium, 2020. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Small-Molecule Medicinal Product | GMO Medicinal Product |
|---|---|
Hazardous characteristics
| Hazardous characteristics
|
Type and extent of release
| Type and extent of release
|
Hazard effect levels/concentration
| Hazard effect levels/concentration
|
Potential for persistence and bioaccumulation
| Potential for persistence and bioaccumulation
|
| Quantitative assessment based on PEC/PNEC ratios | Qualitative assessment based on likelihood of above factors in natural environment |
Labeling
| Labeling
|
| Study Name, Location, Literature Citation | Number of Participants | Dose Levels Evaluated, pfu | Viremia Observed | Shedding Observed |
|---|---|---|---|---|
| Phase 1 | ||||
| V920-001—USA [75] | 30 | 3 × 106, 2 × 107, 1 × 108 | Y | Y |
| Caco-2V920-002—Caco-2USA a [75] | 30 | 3 × 106, 2 × 107, 1 × 108 | Y | Y |
| Caco-2V920-003—Caco-2Canada [46] | 30 | 1 × 105, 5 × 105, 3 × 106 | Y | N |
| Caco-2V920-004—Caco-2USA [47] | 418 | 3 × 103, 3 × 104, 3 × 105, 3 × 106, 9 × 106, 2 × 107, 1 × 108 | Y | Y c |
| Caco-2V920-005—Caco-2Switzerland [76] | 102 | 3 × 105, 1 × 107, 5 × 107 | Y | N |
| Caco-2V920-006—Caco-2Germany [76] | 30 | 3 × 105, 3 × 106, 2 × 107 | Y | N |
| Caco-2V920-007—Caco-2Gabon b [77] | 155 | 3 × 103, 3 × 104, 3 × 105, 3 × 106, 2 × 107 | Y | Y d |
| Caco-2V920-008—Caco-2Kenya [76] | 40 | 3 × 106, 1 × 107 | Y | N |
| Potential Exposure | Measure |
|---|---|
| Accidental breakage/spillage during transport or administration | Medical personnel involved in the administration of rVSVΔG-ZEBOV-GP should be wearing personal protective equipment in order to minimize exposure. Disinfectants such as aldehydes, alcohols, and detergents should always be available in case breakage/spillage were to occur, in order to inactivate the vaccine through chemical disinfection and prevent release into the environment. Detailed instructions on how to handle accidental breakage/spillage have been developed and accompany each shipment of rVSVΔG-ZEBOV-GP. Any unused vaccine or waste material should be disposed of in compliance with applicable institutional guidelines for GMOs or biohazardous waste, as appropriate. |
| Direct human contact with rVSVΔG-ZEBOV-GP shed by vaccinated individuals | Vaccinated individuals should be informed about the potential for shedding and the need to avoid close association with high-risk individuals (i.e., immunocompromised and children < 1 year old) for up to 6 weeks following vaccination, particularly exposing them to blood and bodily fluids. People who develop vesicular rash after receiving the vaccine should cover the vesicles until they heal. The vaccination site or any vesicles should be covered with an adequate bandage (e.g., adhesive bandage, gauze, and tape) that provides a physical barrier protecting against direct contact. The covering may be removed when there is no visible fluid leakage. Vaccinated individuals should not donate blood for 6 weeks following vaccination. |
| Accidental needle stick injury | The injection site should be disinfected immediately and covered, in the same fashion as advised for vaccinated individuals (see above). If this were to occur in the context of a clinical trial, the injured individual should be followed up for safety in the same fashion as a purposely vaccinated trial participant. |
| Direct contact of animals with rVSVΔG-ZEBOV-GP shed by vaccinated individuals | Vaccinated individuals should avoid exposing livestock to their blood and bodily fluids for at least 6 weeks following vaccination. The vaccination site or any vesicles should be covered (see above). |
| Unintended use or misuse | rVSVΔG-ZEBOV-GP shipments delivered to hospital centers for vaccination should be well controlled and be handled as GMOs per local regulations. Only appropriately trained medical personnel should have access to the vaccine. If eye contact were to occur, eyes should be flushed with tepid tap water for 5 min. If skin contact were to occur, exposed areas should be washed with ordinary soap and tap water. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tell, J.G.; Coller, B.-A.G.; Dubey, S.A.; Jenal, U.; Lapps, W.; Wang, L.; Wolf, J. Environmental Risk Assessment for rVSVΔG-ZEBOV-GP, a Genetically Modified Live Vaccine for Ebola Virus Disease. Vaccines 2020, 8, 779. https://doi.org/10.3390/vaccines8040779
Tell JG, Coller B-AG, Dubey SA, Jenal U, Lapps W, Wang L, Wolf J. Environmental Risk Assessment for rVSVΔG-ZEBOV-GP, a Genetically Modified Live Vaccine for Ebola Virus Disease. Vaccines. 2020; 8(4):779. https://doi.org/10.3390/vaccines8040779
Chicago/Turabian StyleTell, Joan G., Beth-Ann G. Coller, Sheri A. Dubey, Ursula Jenal, William Lapps, Liman Wang, and Jayanthi Wolf. 2020. "Environmental Risk Assessment for rVSVΔG-ZEBOV-GP, a Genetically Modified Live Vaccine for Ebola Virus Disease" Vaccines 8, no. 4: 779. https://doi.org/10.3390/vaccines8040779