The Potential of Soluble Human Leukocyte Antigen Molecules for Early Cancer Detection and Therapeutic Vaccine Design



Abstract
:1. Introduction
2. sHLA in Malignancies
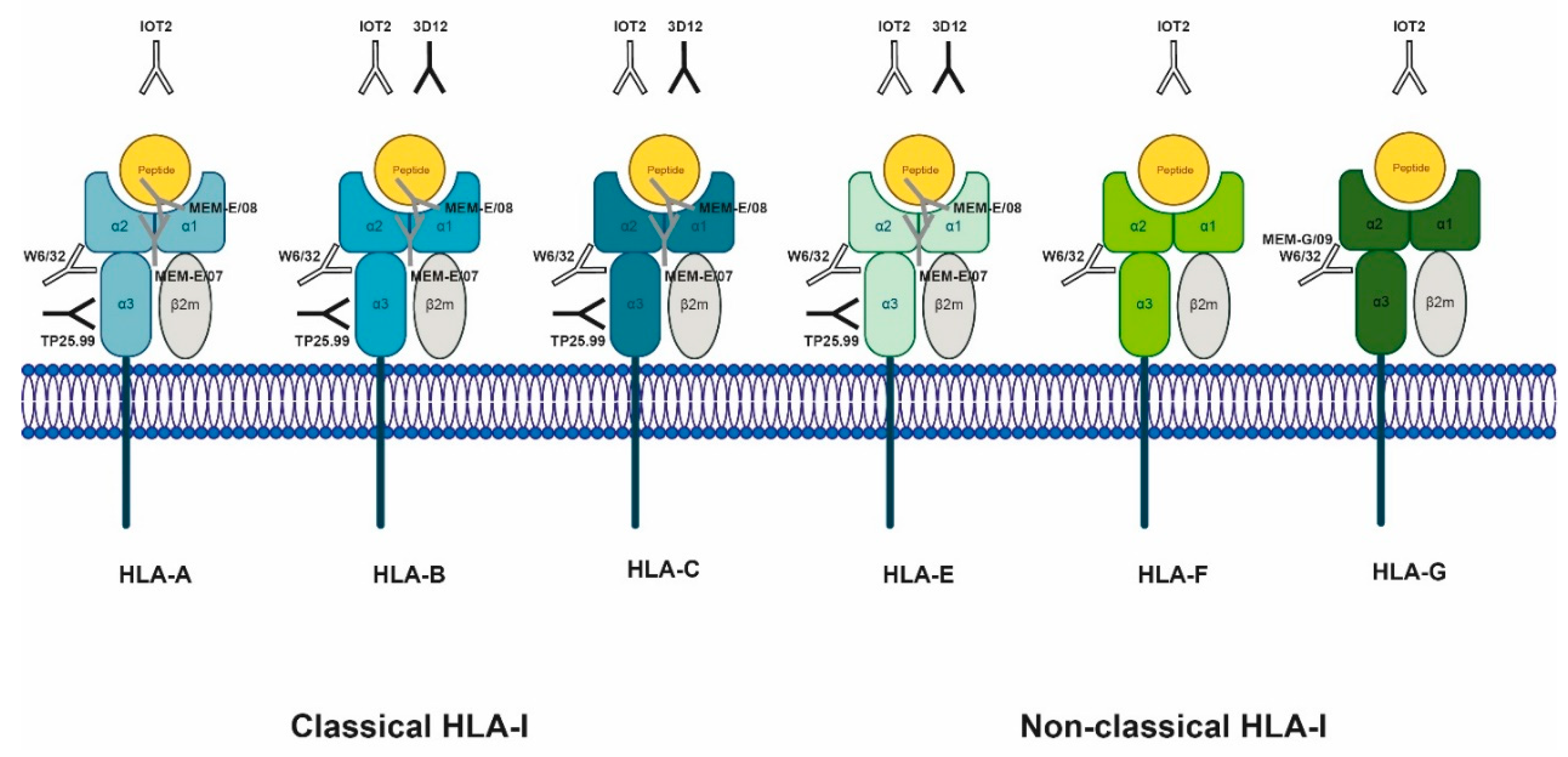
2.1. Classical sHLA Class I
2.2. Non-Classical sHLA Class I
2.3. sHLA Class II
2.4. Confounding Factors Affecting sHLA Levels and/or Detection
2.5. Future Diagnostic Utilization of sHLA
3. sHLA Complexes: Mechanism of Secretion and Immunological Effects
3.1. sHLA-I Complexes
3.2. sHLA-II Complexes
4. sHLA Peptides for Cancer Detection and Therapeutic Vaccine Design
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kamisawa, T.; Wood, L.D.; Itoi, T.; Takaori, K. Pancreatic cancer. Lancet 2016, 388, 73–85. [Google Scholar] [CrossRef]
- Molina, J.R.; Yang, P.; Cassivi, S.D.; Schild, S.E.; Adjei, A.A. Non-small cell lung cancer: Epidemiology, risk factors, treatment, and survivorship. Mayo Clin. Proc. 2008, 83, 584–594. [Google Scholar] [CrossRef]
- Balogh, J.; Victor, D., 3rd; Asham, E.H.; Burroughs, S.G.; Boktour, M.; Saharia, A.; Li, X.; Ghobrial, R.M.; Monsour, H.P., Jr. Hepatocellular carcinoma: A review. J. Hepatocell. Carcinoma 2016, 3, 41–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Rood, J.J.; van Leeuwen, A.; van Santen, M.C. Anti Hl-A2 inhibitor in normal human serum. Nature 1970, 226, 366–367. [Google Scholar] [CrossRef]
- Pistoia, V.; Morandi, F.; Wang, X.; Ferrone, S. Soluble HLA-G: Are they clinically relevant? Semin. Cancer Biol. 2007, 17, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, A.; Rebmann, V.; LeMaoult, J.; Horn, P.A.; Carosella, E.D.; Alegre, E. The immunosuppressive molecule HLA-G and its clinical implications. Crit. Rev. Clin. Lab. Sci. 2012, 49, 63–84. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, S.; Han, Y.; Wang, Y.; Sun, Y. Human leukocyte antigen-g expression and polymorphisms promote cancer development and guide cancer diagnosis/treatment. Oncol. Lett. 2018, 15, 699–709. [Google Scholar] [CrossRef]
- Boegel, S.; Lower, M.; Bukur, T.; Sorn, P.; Castle, J.C.; Sahin, U. Hla and proteasome expression body map. BMC Med. Genomics 2018, 11, 36. [Google Scholar] [CrossRef]
- Lo Monaco, E.; Sibilio, L.; Melucci, E.; Tremante, E.; Suchanek, M.; Horejsi, V.; Martayan, A.; Giacomini, P. HLA-E: Strong association with beta2-microglobulin and surface expression in the absence of HLA class I signal sequence-derived peptides. J. Immunol. 2008, 181, 5442–5450. [Google Scholar] [CrossRef] [Green Version]
- Celik, A.A.; Kraemer, T.; Huyton, T.; Blasczyk, R.; Bade-Doding, C. The diversity of the HLA-E-restricted peptide repertoire explains the immunological impact of the arg107gly mismatch. Immunogenetics 2016, 68, 29–41. [Google Scholar] [CrossRef] [Green Version]
- Tremante, E.; Lo Monaco, E.; Ingegnere, T.; Sampaoli, C.; Fraioli, R.; Giacomini, P. Monoclonal antibodies to HLA-E bind epitopes carried by unfolded beta2 m-free heavy chains. Eur. J. Immunol. 2015, 45, 2356–2364. [Google Scholar] [CrossRef] [PubMed]
- Lepin, E.J.; Bastin, J.M.; Allan, D.S.; Roncador, G.; Braud, V.M.; Mason, D.Y.; van der Merwe, P.A.; McMichael, A.J.; Bell, J.I.; Powis, S.H.; et al. Functional characterization of HLA-F and binding of HLA-F tetramers to ilt2 and ilt4 receptors. Eur. J. Immunol. 2000, 30, 3552–3561. [Google Scholar] [CrossRef]
- Amirghofran, Z.; Sheikhi, A.K.; Kumar, P.V.; Saberi Firouzi, M. Soluble HLA class I molecules in malignant pleural and peritoneal effusions and its possible role on nk and lak cytotoxicity. J. Cancer Res. Clin. Oncol. 2002, 128, 443–448. [Google Scholar] [PubMed]
- Shimura, T.; Hagihara, M.; Yamamoto, K.; Takebe, K.; Munkhbat, B.; Ogoshi, K.; Mitomi, T.; Nagamachi, Y.; Tsuji, K. Quantification of serum-soluble HLA class I antigens in patients with gastric cancer. Hum. Immunol. 1994, 40, 183–186. [Google Scholar] [CrossRef]
- Albitar, M.; Vose, J.M.; Johnson, M.M.; Do, K.A.; Day, A.; Jilani, I.; Kantarjian, H.; Keating, M.; O’Brien, S.M.; Verstovsek, S.; et al. Clinical relevance of soluble hla-i and beta2-microglobulin levels in non-hodgkin’s lymphoma and hodgkin’s disease. Leuk. Res. 2007, 31, 139–145. [Google Scholar] [CrossRef]
- Albitar, M.; Johnson, M.; Do, K.A.; Day, A.; Jilani, I.; Pierce, S.; Estey, E.; Kantarjian, H.; Keating, M.; Verstovsek, S.; et al. Levels of soluble hla-i and beta2m in patients with acute myeloid leukemia and advanced myelodysplastic syndrome: Association with clinical behavior and outcome of induction therapy. Leukemia 2007, 21, 480–488. [Google Scholar] [CrossRef]
- Shimura, T.; Tsutsumi, S.; Hosouchi, Y.; Kojima, T.; Kon, Y.; Yonezu, M.; Kuwano, H. Clinical significance of soluble form of HLA class I molecule in japanese patients with pancreatic cancer. Hum. Immunol. 2001, 62, 615–619. [Google Scholar] [CrossRef]
- Wierenga, A.P.A.; Gezgin, G.; van Beelen, E.; Eikmans, M.; Spruyt-Gerritse, M.; Brouwer, N.J.; Versluis, M.; Verdijk, R.M.; van Duinen, S.G.; Marinkovic, M.; et al. Soluble hla in the aqueous humour of uveal melanoma is associated with unfavourable tumour characteristics. Cancers 2019, 11, 1202. [Google Scholar] [CrossRef] [Green Version]
- Schutt, P.; Schutt, B.; Switala, M.; Bauer, S.; Stamatis, G.; Opalka, B.; Eberhardt, W.; Schuler, M.; Horn, P.A.; Rebmann, V. Prognostic relevance of soluble human leukocyte antigen-g and total human leukocyte antigen class i molecules in lung cancer patients. Hum. Immunol. 2010, 71, 489–495. [Google Scholar] [CrossRef]
- Nocito, M.; Montalban, C.; Gonzalez-Porque, P.; Villar, L.M. Increased soluble serum HLA class I antigens in patients with lymphoma. Hum. Immunol. 1997, 58, 106–111. [Google Scholar] [CrossRef]
- Schutt, P.; Rebmann, V.; Brandhorst, D.; Wiefelsputz, J.; Ebeling, P.; Opalka, B.; Seeber, S.; Nowrousian, M.R.; Moritz, T.; Grosse-Wilde, H. The clinical significance of soluble human leukocyte antigen class-i, ictp, and rankl molecules in multiple myeloma patients. Hum. Immunol. 2008, 69, 79–87. [Google Scholar] [CrossRef] [PubMed]
- Leleu, X.; Le Friec, G.; Facon, T.; Amiot, L.; Fauchet, R.; Hennache, B.; Coiteux, V.; Yakoub-Agha, I.; Dubucquoi, S.; Avet-Loiseau, H.; et al. Total soluble HLA class I and soluble HLA-G in multiple myeloma and monoclonal gammopathy of undetermined significance. Clin. Cancer Res. 2005, 11, 7297–7303. [Google Scholar] [CrossRef] [Green Version]
- Moreau, A.S.; Sebti, Y.; Duhamel, A.; Roccaro, A.M.; Coiteux, V.; Gastinne, T.; Le Friec, G.; Burwick, N.; Amiot, L.; Ho, A.W.; et al. Clinical relevance of soluble HLA class I molecules in waldenstrom macroglobulinemia. Eur. J. Haematol. 2008, 80, 503–509. [Google Scholar] [CrossRef] [PubMed]
- Contini, P.; Zocchi, M.R.; Pierri, I.; Albarello, A.; Poggi, A. In vivo apoptosis of cd8(+) lymphocytes in acute myeloid leukemia patients: Involvement of soluble hla-i and fas ligand. Leukemia 2007, 21, 253–260. [Google Scholar] [CrossRef] [PubMed]
- Le Discorde, M.; Moreau, P.; Sabatier, P.; Legeais, J.M.; Carosella, E.D. Expression of HLA-G in human cornea, an immune-privileged tissue. Hum. Immunol. 2003, 64, 1039–1044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mallet, V.; Blaschitz, A.; Crisa, L.; Schmitt, C.; Fournel, S.; King, A.; Loke, Y.W.; Dohr, G.; Le Bouteiller, P. HLA-G in the human thymus: A subpopulation of medullary epithelial but not cd83(+) dendritic cells expresses HLA-G as a membrane-bound and soluble protein. Int. Immunol. 1999, 11, 889–898. [Google Scholar] [CrossRef] [Green Version]
- Cirulli, V.; Zalatan, J.; McMaster, M.; Prinsen, R.; Salomon, D.R.; Ricordi, C.; Torbett, B.E.; Meda, P.; Crisa, L. The class i hla repertoire of pancreatic islets comprises the nonclassical class ib antigen HLA-G. Diabetes 2006, 55, 1214–1222. [Google Scholar] [CrossRef] [Green Version]
- Lee, N.; Ishitani, A.; Geraghty, D.E. HLA-F is a surface marker on activated lymphocytes. Eur. J. Immunol. 2010, 40, 2308–2318. [Google Scholar] [CrossRef] [Green Version]
- Ishitani, A.; Sageshima, N.; Lee, N.; Dorofeeva, N.; Hatake, K.; Marquardt, H.; Geraghty, D.E. Protein expression and peptide binding suggest unique and interacting functional roles for HLA-E, F, and G in maternal-placental immune recognition. J. Immunol. 2003, 171, 1376–1384. [Google Scholar] [CrossRef]
- Lee, N.; Goodlett, D.R.; Ishitani, A.; Marquardt, H.; Geraghty, D.E. HLA-E surface expression depends on binding of tap-dependent peptides derived from certain HLA class I signal sequences. J. Immunol. 1998, 160, 4951–4960. [Google Scholar]
- Bukur, J.; Jasinski, S.; Seliger, B. The role of classical and non-classical HLA class I antigens in human tumors. Semin. Cancer Biol. 2012, 22, 350–358. [Google Scholar] [CrossRef]
- Li, J.B.; Ruan, Y.Y.; Hu, B.; Dong, S.S.; Bi, T.N.; Lin, A.; Yan, W.H. Importance of the plasma soluble HLA-G levels for prognostic stratification with traditional prognosticators in colorectal cancer. Oncotarget 2017, 8, 48854–48862. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agnihotri, V.; Gupta, A.; Kumar, L.; Dey, S. Serum sHLA-G: Significant diagnostic biomarker with respect to therapy and immunosuppressive mediators in head and neck squamous cell carcinoma. Sci. Rep. 2020, 10, 3806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lazaro-Sanchez, A.D.; Salces-Ortiz, P.; Velasquez, L.I.; Orozco-Beltran, D.; Diaz-Fernandez, N.; Juarez-Marroqui, A. HLA-G as a new tumor biomarker: Detection of soluble isoforms of HLA-G in the serum and saliva of patients with colorectal cancer. Clin. Transl. Oncol. 2020, 22, 1166–1171. [Google Scholar] [CrossRef]
- Zhu, C.B.; Wang, C.X.; Zhang, X.; Zhang, J.; Li, W. Serum sHLA-G levels: A useful indicator in distinguishing colorectal cancer from benign colorectal diseases. Int. J. Cancer 2011, 128, 617–622. [Google Scholar] [CrossRef]
- Pan, Y.Q.; Ruan, Y.Y.; Peng, J.B.; Han, Q.Y.; Zhang, X.; Lin, A.; Yan, W.H. Diagnostic significance of soluble human leukocyte antigen-g for gastric cancer. Hum. Immunol. 2016, 77, 317–324. [Google Scholar] [CrossRef]
- Wang, Y.; Ye, Z.; Meng, X.Q.; Zheng, S.S. Expression of HLA-G in patients with hepatocellular carcinoma. Hepatobiliary Pancreat Dis. Int. 2011, 10, 158–163. [Google Scholar] [CrossRef]
- Heidari, M.H.; Movafagh, A.; Abdollahifar, M.A.; Abdi, S.; Barez, M.M.; Azimi, H.; Moradi, A.; Bagheri, A.; Heidari, M.; Hessam Mohseni, J.; et al. Evaluation of sHLA-G levels in serum of patients with prostate cancer identify as a potential of tumor marker. Anat. Cell Biol. 2017, 50, 69–72. [Google Scholar] [CrossRef] [Green Version]
- Sipak-Szmigiel, O.; Wlodarski, P.; Ronin-Walknowska, E.; Niedzielski, A.; Karakiewicz, B.; Sluczanowska-Glabowska, S.; Laszczynska, M.; Malinowski, W. Serum and peritoneal fluid concentrations of soluble human leukocyte antigen, tumor necrosis factor alpha and interleukin 10 in patients with selected ovarian pathologies. J. Ovarian Res. 2017, 10, 25. [Google Scholar] [CrossRef] [Green Version]
- Akin, M.; Aral, L.A.; Yavuz, A.; Karabacak, H.; Dikmen, K.; Bostanci, H. Plasma human leukocyte antigen-g (HLA-G) in patients with thyroid cancer. Turk. J. Med. Sci. 2017, 47, 1263–1266. [Google Scholar] [CrossRef]
- Bertol, B.C.; de Araujo, J.N.G.; Sadissou, I.A.; Sonon, P.; Dias, F.C.; Bortolin, R.H.; de Figueiredo-Feitosa, N.L.; de Freitas, L.C.C.; de Miranda Henrique Tarrapp, S.R.; de Oliveira Ramos, C.C.; et al. Plasma levels of soluble HLA-G and cytokines in papillary thyroid carcinoma before and after thyroidectomy. Int. J. Clin. Pract. 2020, 74, e13585. [Google Scholar] [CrossRef] [PubMed]
- de Figueiredo Feitosa, N.L.; Crispim, J.C.; Zanetti, B.R.; Magalhaes, P.K.; Soares, C.P.; Soares, E.G.; Neder, L.; Donadi, E.A.; Maciel, L.M. HLA-G is differentially expressed in thyroid tissues. Thyroid 2014, 24, 585–592. [Google Scholar] [CrossRef] [PubMed]
- Kirana, C.; Ruszkiewicz, A.; Stubbs, R.S.; Hardingham, J.E.; Hewett, P.J.; Maddern, G.J.; Hauben, E. Soluble HLA-G is a differential prognostic marker in sequential colorectal cancer disease stages. Int. J. Cancer 2017, 140, 2577–2586. [Google Scholar] [CrossRef] [PubMed]
- Adolf, I.C.; Akan, G.; Mselle, T.F.; Dharsee, N.; Namkinga, L.A.; Atalar, F. Implication of soluble HLA-G and HLA-G + 3142g/c polymorphism in breast cancer patients receiving adjuvant therapy in tanzania. Asian Pac. J. Cancer Prev. 2019, 20, 3465–3472. [Google Scholar] [CrossRef]
- Ben Yahia, H.; Babay, W.; Bortolotti, D.; Boujelbene, N.; Laaribi, A.B.; Zidi, N.; Kehila, M.; Chelbi, H.; Boudabous, A.; Mrad, K.; et al. Increased plasmatic soluble HLA-G levels in endometrial cancer. Mol. Immunol. 2018, 99, 82–86. [Google Scholar] [CrossRef]
- Morandi, F.; Pozzi, S.; Carlini, B.; Amoroso, L.; Pistoia, V.; Corrias, M.V. Soluble HLA-G and HLA-E levels in bone marrow plasma samples are related to disease stage in neuroblastoma patients. J. Immunol. Res. 2016, 2016, 7465741. [Google Scholar] [CrossRef] [Green Version]
- Motawi, T.M.; Zakhary, N.I.; Salman, T.M.; Tadros, S.A. Serum human leukocyte antigen-g and soluble interleukin 2 receptor levels in acute lymphoblastic leukemic pediatric patients. Asian Pac. J. Cancer Prev. 2012, 13, 5399–5403. [Google Scholar] [CrossRef] [Green Version]
- Morandi, F.; Cangemi, G.; Barco, S.; Amoroso, L.; Giuliano, M.; Gigliotti, A.R.; Pistoia, V.; Corrias, M.V. Plasma levels of soluble HLA-E and HLA-F at diagnosis may predict overall survival of neuroblastoma patients. Biomed. Res. Int. 2013, 2013, 956878. [Google Scholar] [CrossRef]
- Scarabel, L.; Garziera, M.; Fortuna, S.; Asaro, F.; Toffoli, G.; Geremia, S. Soluble HLA-G expression levels and HLA-G/irinotecan association in metastatic colorectal cancer treated with irinotecan-based strategy. Sci. Rep. 2020, 10, 8773. [Google Scholar] [CrossRef]
- Farjadian, S.; Tabebordbar, M.; Mokhtari, M.; Safaei, A.; Malekzadeh, M.; Ghaderi, A. HLA-G expression in tumor tissues and soluble HLA-G plasma levels in patients with gastrointestinal cancer. Asian Pac. J. Cancer Prev. 2018, 19, 2731–2735. [Google Scholar]
- Konig, L.; Kasimir-Bauer, S.; Hoffmann, O.; Bittner, A.K.; Wagner, B.; Manvailer, L.F.; Schramm, S.; Bankfalvi, A.; Giebel, B.; Kimmig, R.; et al. The prognostic impact of soluble and vesicular HLA-G and its relationship to circulating tumor cells in neoadjuvant treated breast cancer patients. Hum. Immunol. 2016, 77, 791–799. [Google Scholar] [CrossRef]
- Ugurel, S.; Rebmann, V.; Ferrone, S.; Tilgen, W.; Grosse-Wilde, H.; Reinhold, U. Soluble human leukocyte antigen—G serum level is elevated in melanoma patients and is further increased by interferon-alpha immunotherapy. Cancer 2001, 92, 369–376. [Google Scholar] [CrossRef]
- Almeida, R.D.S.; Ramos, A.M.L.; Luna, C.F.; Pedrosa, F.; Donadi, E.A.; Lucena-Silva, N. Cytokines and soluble HLA-G levels in bone marrow stroma and their association with the survival rate of patients exhibiting childhood t-cell acute lymphoblastic leukemia. Cytokine 2018, 102, 94–101. [Google Scholar] [CrossRef] [PubMed]
- Hamed, N.A.; El Halawani, N.; Nafea, D.; El Rahman, M.A.; Kasber, A. Soluble HLA-G: A novel marker in acute myeloid leukemia patients. Acta Med. Int. 2016, 4, 51–55. [Google Scholar] [CrossRef]
- Caocci, G.; Greco, M.; Arras, M.; Cusano, R.; Orru, S.; Martino, B.; Abruzzese, E.; Galimberti, S.; Mulas, O.; Trucas, M.; et al. HLA-G molecules and clinical outcome in chronic myeloid leukemia. Leuk. Res. 2017, 61, 1–5. [Google Scholar] [CrossRef]
- Allard, M.; Oger, R.; Vignard, V.; Percier, J.M.; Fregni, G.; Perier, A.; Caignard, A.; Charreau, B.; Bernardeau, K.; Khammari, A.; et al. Serum soluble HLA-E in melanoma: A new potential immune-related marker in cancer. PLoS ONE 2011, 6, e21118. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Yang, H.; Ying, S.; Lu, H.; Wang, W.; Lv, J.; Xiong, H.; Hu, W. High HLA-F expression is a poor prognosis factor in patients with nasopharyngeal carcinoma. Anal. Cell. Pathol. 2018, 2018, 7691704. [Google Scholar] [CrossRef] [Green Version]
- Wagner, B.; da Silva Nardi, F.; Schramm, S.; Kraemer, T.; Celik, A.A.; Durig, J.; Horn, P.A.; Duhrsen, U.; Nuckel, H.; Rebmann, V. HLA-E allelic genotype correlates with HLA-E plasma levels and predicts early progression in chronic lymphocytic leukemia. Cancer 2017, 123, 814–823. [Google Scholar] [CrossRef]
- Xu, Y.P.; Wieten, L.; Wang, S.X.; Cai, Y.; Olieslagers, T.; Zhang, L.; He, L.M.; Tilanus, M.G.J.; Hong, W.X. Clinical significance of HLA-E genotype and surface/soluble expression levels between healthy individuals and patients with acute leukemia. Leuk. Lymphoma 2019, 60, 208–215. [Google Scholar] [CrossRef]
- Rebmann, V.; Ugurel, S.; Tilgen, W.; Reinhold, U.; Grosse-Wilde, H. Soluble hla-dr is a potent predictive indicator of disease progression in serum from early-stage melanoma patients. Int. J. Cancer 2002, 100, 580–585. [Google Scholar] [CrossRef]
- Hassan, N.; Dhaliwal, J.S.; Mohd Ibrahim, H.; Osman, R.; SZ, H.I.; Lee le, J.; Abdullah, M. Increased soluble hla-drb1in b-cell acute lymphoblastic leukaemia. Malays. J. Pathol. 2015, 37, 83–90. [Google Scholar] [PubMed]
- Contini, P.; Ghio, M.; Poggi, A.; Filaci, G.; Indiveri, F.; Ferrone, S.; Puppo, F. Soluble hla-a,-b,-c and -g molecules induce apoptosis in t and nk cd8 + cells and inhibit cytotoxic t cell activity through cd8 ligation. Eur. J. Immunol. 2003, 33, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Rebmann, V.; Pfeiffer, K.; Passler, M.; Ferrone, S.; Maier, S.; Weiss, E.; Grosse-Wilde, H. Detection of soluble HLA-G molecules in plasma and amniotic fluid. Tissue Antigens 1999, 53, 14–22. [Google Scholar] [CrossRef] [PubMed]
- Desai, S.A.; Wang, X.; Noronha, E.J.; Zhou, Q.; Rebmann, V.; Grosse-Wilde, H.; Moy, F.J.; Powers, R.; Ferrone, S. Structural relatedness of distinct determinants recognized by monoclonal antibody TP25.99 on β2-microglobulin-associated and β2-microglobulin-free HLA class I heavy chains. J. Immunol. 2000, 165, 3275–3283. [Google Scholar] [CrossRef] [Green Version]
- Pouletty, P.; Ferrone, S.; Amesland, F.; Cohen, N.; Westhoff, U.; Charron, D.; Shimizu, R.M.; Grosse-Wilde, H. Summary report from the first international workshop on soluble hla antigens. Paris, august 1992. Tissue Antigens 1993, 42, 45–54. [Google Scholar] [CrossRef]
- Ravindranath, M.H.; Pham, T.; El-Awar, N.; Kaneku, H.; Terasaki, P.I. Anti-HLA-E mab 3d12 mimics mem-e/02 in binding to HLA-B and HLA-C alleles: Web-tools validate the immunogenic epitopes of HLA-E recognized by the antibodies. Mol. Immunol. 2011, 48, 423–430. [Google Scholar] [CrossRef]
- Grumet, F.C.; Buelow, R.; Grosse-Wilde, H.; Kubens, B.; Garovoy, M.; Pouletty, P. Report of the second international soluble HLA (sHLA) workshop. Phoenix, arizona, october 2, 1993. Hum. Immunol. 1994, 40, 153–165. [Google Scholar] [CrossRef]
- Comuzio, S.; Puppo, F.; Ruzzenenti, R.; Orlandini, A.; Grillo, F.; Brenci, S.; Lanza, L.; Scudeletti, M.; Indiveri, F. Simple elisa method for the evaluation of soluble HLA class I antigens in human serum. J. Clin. Lab. Anal. 1991, 5, 278–283. [Google Scholar] [CrossRef]
- Rebmann, V.; Lemaoult, J.; Rouas-Freiss, N.; Carosella, E.D.; Grosse-Wilde, H. Report of the wet workshop for quantification of soluble HLA-G in essen, 2004. Hum. Immunol. 2005, 66, 853–863. [Google Scholar] [CrossRef]
- Polakova, K.; Bandzuchova, E.; Russ, G. Impact of blood processing on estimation of soluble HLA-G. Neoplasma 2011, 58, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Stevenson, F.K.; George, A.J.; Walters, M.T.; Hamblin, T.J. Analysis of soluble HLA class Ii antigenic material in patients with immunological diseases using monoclonal antibodies. J. Immunol. Methods 1986, 86, 187–190. [Google Scholar] [CrossRef]
- O’Connell, G.C.; Alder, M.L.; Webel, A.R.; Moore, S.M. Neuro biomarker levels measured with high-sensitivity digital elisa differ between serum and plasma. Bioanalysis 2019, 11, 2087–2094. [Google Scholar] [CrossRef]
- Rudstein-Svetlicky, N.; Loewenthal, R.; Horejsi, V.; Gazit, E. HLA-G levels in serum and plasma. Tissue Antigens 2006, 67, 111–116. [Google Scholar] [CrossRef]
- Lee, J.E.; Kim, J.W.; Han, B.G.; Shin, S.Y. Impact of whole-blood processing conditions on plasma and serum concentrations of cytokines. Biopreserv. Biobank. 2016, 14, 51–55. [Google Scholar] [CrossRef]
- Hillebrand, K.; Moritz, T.; Westhoff, U.; Niederle, N.; Grosse-Wilde, H. Soluble HLA class I and beta-2-microglobulin plasma concentrations during interferon treatment of chronic myelogenous leukemia. Vox Sang. 1994, 67, 310–314. [Google Scholar] [CrossRef]
- Keskinen, P.; Ronni, T.; Matikainen, S.; Lehtonen, A.; Julkunen, I. Regulation of HLA class I and ii expression by interferons and influenza a virus in human peripheral blood mononuclear cells. Immunology 1997, 91, 421–429. [Google Scholar] [CrossRef]
- Coomans de Brachene, A.; Dos Santos, R.S.; Marroqui, L.; Colli, M.L.; Marselli, L.; Mirmira, R.G.; Marchetti, P.; Eizirik, D.L. Ifn-alpha induces a preferential long-lasting expression of MHC class I in human pancreatic beta cells. Diabetologia 2018, 61, 636–640. [Google Scholar] [CrossRef] [Green Version]
- Lefebvre, S.; Moreau, P.; Guiard, V.; Ibrahim, E.C.; Adrian-Cabestre, F.; Menier, C.; Dausset, J.; Carosella, E.D.; Paul, P. Molecular mechanisms controlling constitutive and ifn-gamma-inducible HLA-G expression in various cell types. J. Reprod. Immunol. 1999, 43, 213–224. [Google Scholar] [CrossRef]
- Alegre, E.; Rizzo, R.; Bortolotti, D.; Fernandez-Landazuri, S.; Fainardi, E.; Gonzalez, A. Some basic aspects of HLA-G biology. J. Immunol. Res. 2014, 2014, 657625. [Google Scholar] [CrossRef] [Green Version]
- Budhwani, M.; Mazzieri, R.; Dolcetti, R. Plasticity of type i interferon-mediated responses in cancer therapy: From anti-tumor immunity to resistance. Front. Oncol. 2018, 8, 322. [Google Scholar] [CrossRef] [Green Version]
- Kao, K.J.; Scornik, J.C.; Riley, W.J.; McQueen, C.F. Association between HLA phenotype and HLA concentration in plasma or platelets. Hum. Immunol. 1988, 21, 115–124. [Google Scholar] [CrossRef]
- Kubysheva, N.; Soodaeva, S.; Novikov, V.; Eliseeva, T.; Li, T.; Klimanov, I.; Kuzmina, E.; Baez-Medina, H.; Solovyev, V.; Ovsyannikov, D.Y.; et al. Soluble hla-i and hla-ii molecules are potential prognostic markers of progression of systemic and local inflammation in patients with copd. Dis. Markers 2018, 2018, 3614341. [Google Scholar] [CrossRef]
- Adamashvili, I.; Pressly, T.; Gebel, H.; Milford, E.; Wolf, R.; Mancini, M.; Sittg, K.; Ghali, G.E.; Hall, V.; McDonald, J.C. Soluble hla in saliva of patients with autoimmune rheumatic diseases. Rheumatol. Int. 2002, 22, 71–76. [Google Scholar] [CrossRef]
- Heiligenhaus, A.; Rebmann, V.; Neubert, A.; Plewa, S.; Ferencik, S.; Vogeler, U.; Steuhl, K.P.; Grosse-Wilde, H. Soluble HLA class I and HLA-DR plasma levels in patients with anterior uveitis. Tissue Antigens 2004, 63, 369–375. [Google Scholar] [CrossRef]
- Huang, J.; Al-Mozaini, M.; Rogich, J.; Carrington, M.F.; Seiss, K.; Pereyra, F.; Lichterfeld, M.; Yu, X.G. Systemic inhibition of myeloid dendritic cells by circulating HLA class I molecules in HIV-1 infection. Retrovirology 2012, 9, 11. [Google Scholar] [CrossRef] [Green Version]
- Murdaca, G.; Contini, P.; Cagnati, P.; Marenco, S.; Pieri, G.; Lantieri, F.; Picciotto, A.; Puppo, F. Behavior of soluble HLA-A, -B, -C and HLA-G molecules in patients with chronic hepatitis c virus infection undergoing pegylated interferon-alpha and ribavirin treatment: Potential role as markers of response to antiviral therapy. Clin. Exp. Med. 2017, 17, 93–100. [Google Scholar] [CrossRef]
- Campoli, M.; Ferrone, S. Tumor escape mechanisms: Potential role of soluble hla antigens and nk cells activating ligands. Tissue Antigens 2008, 72, 321–334. [Google Scholar] [CrossRef] [Green Version]
- Ritz, D.; Gloger, A.; Weide, B.; Garbe, C.; Neri, D.; Fugmann, T. High-sensitivity HLA class I peptidome analysis enables a precise definition of peptide motifs and the identification of peptides from cell lines and patients’ sera. Proteomics 2016, 16, 1570–1580. [Google Scholar] [CrossRef] [Green Version]
- Ritz, D.; Gloger, A.; Neri, D.; Fugmann, T. Purification of soluble HLA class I complexes from human serum or plasma deliver high quality immuno peptidomes required for biomarker discovery. Proteomics 2017, 17. [Google Scholar] [CrossRef] [Green Version]
- Demaria, S.; Bushkin, Y. Soluble hla proteins with bound peptides are released from the cell surface by the membrane metalloproteinase. Hum. Immunol. 2000, 61, 1332–1338. [Google Scholar] [CrossRef]
- Bassani-Sternberg, M.; Barnea, E.; Beer, I.; Avivi, I.; Katz, T.; Admon, A. Soluble plasma hla peptidome as a potential source for cancer biomarkers. Proc. Natl. Acad. Sci. USA 2010, 107, 18769–18776. [Google Scholar] [CrossRef] [Green Version]
- Bakela, K.; Athanassakis, I. Soluble major histocompatibility complex molecules in immune regulation: Highlighting class ii antigens. Immunology 2018, 153, 315–324. [Google Scholar] [CrossRef] [Green Version]
- van Niel, G.; D’Angelo, G.; Raposo, G. Shedding light on the cell biology of extracellular vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Krangel, M.S. Secretion of HLA-A and -B antigens via an alternative rna splicing pathway. J. Exp. Med. 1986, 163, 1173–1190. [Google Scholar] [CrossRef] [PubMed]
- Concha-Benavente, F.; Srivastava, R.; Ferrone, S.; Ferris, R.L. Immunological and clinical significance of HLA class I antigen processing machinery component defects in malignant cells. Oral. Oncol. 2016, 58, 52–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haynes, L.D.; Bushkin, Y.; Love, R.B.; Burlingham, W.J. Interferon-gamma drives the metalloproteinase-dependent cleavage of HLA class I soluble forms from primary human bronchial epithelial cells. Hum. Immunol. 2002, 63, 893–901. [Google Scholar] [CrossRef]
- He, X.; Le, J. Alternative splicing of HLA class I transcripts induced by ifn-gamma and tnf in fibroblasts: Release of soluble HLA class I heavy chain and an associate protein. Cell. Immunol. 1995, 162, 159–168. [Google Scholar] [CrossRef]
- Brieva, J.A.; Villar, L.M.; Leoro, G.; Alvarez-Cermeno, J.C.; Roldan, E.; Gonzalez-Porque, P. Soluble HLA class I antigen secretion by normal lymphocytes: Relationship with cell activation and effect of interferon-gamma. Clin. Exp. Immunol. 1990, 82, 390–395. [Google Scholar] [CrossRef]
- Le, J.; Hua, J.C. Production of soluble HLA-class-I molecules by ifn-γ-induced colon-adenocarcinoma cells. Int. J. Cancer 1995, 60, 576–581. [Google Scholar] [CrossRef]
- Derre, L.; Corvaisier, M.; Charreau, B.; Moreau, A.; Godefroy, E.; Moreau-Aubry, A.; Jotereau, F.; Gervois, N. Expression and release of HLA-E by melanoma cells and melanocytes: Potential impact on the response of cytotoxic effector cells. J. Immunol. 2006, 177, 3100–3107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gros, F.; Sebti, Y.; de Guibert, S.; Branger, B.; Bernard, M.; Fauchet, R.; Amiot, L. Soluble HLA-G molecules increase during acute leukemia, especially in subtypes affecting monocytic and lymphoid lineages. Neoplasia 2006, 8, 223–230. [Google Scholar] [CrossRef]
- Datema, G.; Mulder, A.; Claas, F.H.; van den Elsen, P.J. Detection and characterization of HLA class I molecules in the supernatant of an hepatocarcinoma cell line and of ebv-transformed b cell lines. Hum. Immunol. 1999, 60, 435–441. [Google Scholar] [CrossRef]
- Allard, M.; Oger, R.; Benlalam, H.; Florenceau, L.; Echasserieau, K.; Bernardeau, K.; Labarriere, N.; Lang, F.; Gervois, N. Soluble HLA-I/peptide monomers mediate antigen-specific cd8 t cell activation through passive peptide exchange with cell-bound hla-i molecules. J. Immunol. 2014, 192, 5090–5097. [Google Scholar] [CrossRef] [Green Version]
- Ge, Q.; Stone, J.D.; Thompson, M.T.; Cochran, J.R.; Rushe, M.; Eisen, H.N.; Chen, J.; Stern, L.J. Soluble peptide-mhc monomers cause activation of cd8+ t cells through transfer of the peptide to t cell mhc molecules. Proc. Natl. Acad. Sci. USA 2002, 99, 13729–13734. [Google Scholar] [CrossRef] [Green Version]
- Poggi, A.; Contini, P.; Catellani, S.; Setti, M.; Murdaca, G.; Zocchi, M.R. Regulation of gammadelta t cell survival by soluble hla-i: Involvement of cd8 and activating killer ig-like receptors. Eur. J. Immunol. 2005, 35, 2670–2678. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Contini, P.; Negrini, S.; Dondero, A.; Carosio, R.; Ghio, M.; Puppo, F.; Indiveri, F.; Zocchi, M.R.; Poggi, A. Ifn-gamma production in human nk cells through the engagement of cd8 by soluble or surface HLA class I molecules. Eur. J. Immunol. 2003, 33, 3049–3059. [Google Scholar] [CrossRef]
- Spaggiari, G.M.; Contini, P.; Carosio, R.; Arvigo, M.; Ghio, M.; Oddone, D.; Dondero, A.; Zocchi, M.R.; Puppo, F.; Indiveri, F.; et al. Soluble HLA class I molecules induce natural killer cell apoptosis through the engagement of cd8: Evidence for a negative regulation exerted by members of the inhibitory receptor superfamily. Blood 2002, 99, 1706–1714. [Google Scholar] [CrossRef] [Green Version]
- Jendro, M.; Goronzy, J.J.; Weyand, C.M. Structural and functional characterization of hla-dr molecules circulating in the serum. Autoimmunity 1991, 8, 289–296. [Google Scholar] [CrossRef]
- Bakela, K.; Kountourakis, N.; Aivaliotis, M.; Athanassakis, I. Soluble mhc-ii proteins promote suppressive activity in cd4 + T cells. Immunology 2015, 144, 158–169. [Google Scholar] [CrossRef] [Green Version]
- Ritz, D.; Sani, E.; Debiec, H.; Ronco, P.; Neri, D.; Fugmann, T. Membranal and blood-soluble HLA class Ii peptidome analyses using data-dependent and independent acquisition. Proteomics 2018, 18, e1700246. [Google Scholar] [CrossRef]
- Aultman, D.; Adamashvili, I.; Yaturu, K.; Langford, M.; Gelder, F.; Gautreaux, M.; Ghali, G.E.; McDonald, J. Soluble hla in human body fluids. Hum. Immunol. 1999, 60, 239–244. [Google Scholar] [CrossRef]
- Adamashvili, I.; Minagar, A.; Gonzalez-Toledo, E.; Featherston, L.; Kelley, R.E. Soluble hla measurement in saliva and cerebrospinal fluid in caucasian patients with multiple sclerosis: A preliminary study. J. Neuroinflamm. 2005, 2, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buschow, S.I.; Nolte-’t Hoen, E.N.; van Niel, G.; Pols, M.S.; ten Broeke, T.; Lauwen, M.; Ossendorp, F.; Melief, C.J.; Raposo, G.; Wubbolts, R.; et al. Mhc ii in dendritic cells is targeted to lysosomes or t cell-induced exosomes via distinct multivesicular body pathways. Traffic 2009, 10, 1528–1542. [Google Scholar] [CrossRef] [PubMed]
- Buschow, S.I.; van Balkom, B.W.; Aalberts, M.; Heck, A.J.; Wauben, M.; Stoorvogel, W. Mhc class ii-associated proteins in b-cell exosomes and potential functional implications for exosome biogenesis. Immunol. Cell. Biol. 2010, 88, 851–856. [Google Scholar] [CrossRef] [PubMed]
- MacKay, P.A.; Leibundgut-Landmann, S.; Koch, N.; Dunn, A.C.; Reith, W.; Jack, R.W.; McLellan, A.D. Circulating, soluble forms of major histocompatibility complex antigens are not exosome-associated. Eur. J. Immunol. 2006, 36, 2875–2884. [Google Scholar] [CrossRef] [PubMed]
- Athanassakis, I.; Ranella, A.; Vassiliadis, S. IFN-γ facilitates release of class II-loaded intracellular pools in trophoblast cells: A novel property independent of protein synthesis. J. Interferon Cytokine Res. 2000, 20, 823–830. [Google Scholar] [CrossRef]
- Marios-Frankiskos, S.; Panagiota, M.; Katerina, B.; Athanassakis, I. Serum-derived mhc class II molecules: Potent regulators of the cellular and humoral immune response. Immunobiology 2010, 215, 194–205. [Google Scholar] [CrossRef]
- Bakela, K.; Dimakopoulou, M.; Batsou, P.; Manidakis, N.; Athanassakis, I. Soluble mhc class ii-driven therapy for a systemic lupus erythematosus murine experimental in vitro and in vivo model. Scand. J. Immunol. 2018, 87. [Google Scholar] [CrossRef] [Green Version]
- Nag, B.; Kendrick, T.; Arimilli, S.; Yu, S.C.; Sriram, S. Soluble mhc ii-peptide complexes induce antigen-specific apoptosis in t cells. Cell. Immunol. 1996, 170, 25–33. [Google Scholar] [CrossRef]
- Shraibman, B.; Barnea, E.; Kadosh, D.M.; Haimovich, Y.; Slobodin, G.; Rosner, I.; Lopez-Larrea, C.; Hilf, N.; Kuttruff, S.; Song, C.; et al. Identification of tumor antigens among the hla peptidomes of glioblastoma tumors and plasma. Mol. Cell. Proteomics 2019, 18, 1255–1268. [Google Scholar] [CrossRef] [Green Version]
- Purcell, A.W.; Ramarathinam, S.H.; Ternette, N. Mass spectrometry-based identification of mhc-bound peptides for immunopeptidomics. Nat. Protoc. 2019, 14, 1687–1707. [Google Scholar] [CrossRef]
- Scull, K.E.; Dudek, N.L.; Corbett, A.J.; Ramarathinam, S.H.; Gorasia, D.G.; Williamson, N.A.; Purcell, A.W. Secreted hla recapitulates the immunopeptidome and allows in-depth coverage of HLA A * 02:01 ligands. Mol. Immunol. 2012, 51, 136–142. [Google Scholar] [CrossRef]
- Barnea, E.; Beer, I.; Patoka, R.; Ziv, T.; Kessler, O.; Tzehoval, E.; Eisenbach, L.; Zavazava, N.; Admon, A. Analysis of endogenous peptides bound by soluble mhc class i molecules: A novel approach for identifying tumor-specific antigens. Eur. J. Immunol. 2002, 32, 213–222. [Google Scholar] [CrossRef]
- Axelrod, M.L.; Cook, R.S.; Johnson, D.B.; Balko, J.M. Biological consequences of MHC-II expression by tumor cells in cancer. Clin. Cancer Res. 2019, 25, 2392–2402. [Google Scholar] [CrossRef]
- Borst, J.; Ahrends, T.; Babala, N.; Melief, C.J.M.; Kastenmuller, W. Cd4(+) t cell help in cancer immunology and immunotherapy. Nat. Rev. Immunol. 2018, 18, 635–647. [Google Scholar] [CrossRef]
- Bantscheff, M.; Schirle, M.; Sweetman, G.; Rick, J.; Kuster, B. Quantitative mass spectrometry in proteomics: A critical review. Anal. Bioanal. Chem. 2007, 389, 1017–1031. [Google Scholar] [CrossRef] [Green Version]
- Marino, F.; Chong, C.; Michaux, J.; Bassani-Sternberg, M. High-throughput, fast, and sensitive immunopeptidomics sample processing for mass spectrometry. Methods Mol. Biol. 2019, 1913, 67–79. [Google Scholar]
- Altman, D.G.; McShane, L.M.; Sauerbrei, W.; Taube, S.E. Reporting recommendations for tumor marker prognostic studies (remark): Explanation and elaboration. PLoS Med. 2012, 9, e1001216. [Google Scholar] [CrossRef] [Green Version]
- Hickman, H.D.; Yewdell, J.W. Mining the plasma immunopeptidome for cancer peptides as biomarkers and beyond. Proc. Natl. Acad. Sci. USA 2010, 107, 18747–18748. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Disease | N for sHLA Measurements | Treatment | Sample Type | Antibodies | sHLA Type | sHLA Levels Normal Controls | Correlation with Progression/Prognosis | sHLA Compared to Normal Controls | Notes |
|---|---|---|---|---|---|---|---|---|---|
| Solid tumors | |||||||||
| Gastric cancer (GC) | Healthy: 15, GC: 74 | Unknown | Serum | W6/32 + IOT2 | sHLA-I | 868.9 ± 715.0 ng/mL * | Lower sHLA-I in higher stadia | ↓ | Lowest sHLA levels in stage IV. HLA-A24+ patients contained higher sHLA-I levels [14] |
| Pancreatic cancer (PC), hepatocellular carcinoma (HCC), gallbladder cancer (GBC), cholangiocellular carcinoma (CCC) | Healthy: 22, PC: 19, HCC: 51, GBC: 6, CCC: 6 | Unknown | Serum | W6/32 + IOT2 | sHLA-I | 1470 ± 95.3 ng/mL ** | N.A. | ↑ | [17] |
| Uveal melanoma | 108 | Yes (4), no (104). All undergoing enucleation | Aqueous humor | W6/32 + anti-β2M | sHLA-I | N.A. | Higher sHLA-I in higher stadia, bigger tumor prominence, ciliary body involvement, metastasis and poorer survival | ↑ | Magnetic Luminex; highest levels in stage III and IV [18] |
| Small-cell lung cancer (SCLC) and non-small-cell lung cancer (NSCLC) | Healthy: 84, SCLC: 23, NSCLC: 114 | n = 6 for chemotherapy >3 months before blood collection | Plasma | W6/32 + anti-β2M | sHLA-I | sHLA-I: 1370 ng/mL (274–2670) *** | sHLA-I under cut off had significant prolonged OS | ↑ | [19] |
| Various malignancies | Randomly selected malignancies: 75; Infection that was cause of effusion: 21; Other pathologies: 21 | Unknown | Body cavity fluids (70 pleural and 53 peritoneal) | W6/32 | sHLA-I | Other diseases: 0.49 ± 0.45 ug/mL ** | N.A. | N.A. | Competitive ELISA. Trend towards higher sHLA-I in malignant and infected patients compared to others, but not significantly different [13] |
| Hematological tumors | |||||||||
| Non-Hodgkin’s lymphoma (NHL) and Hodgkin’s disease (HD) | Healthy: ?, NHL: 65, HD: 37 | Treatment naive | Serum | W6/32 + anti-β2M | sHLA-I | Unknown | NHL: higher sHLA-I levels are associated with shorter survival. HD: none found | ↑ | sHLA-I levels are higher pre-therapy [15] |
| NHL and HD | Healthy: 50, NHL: 40, HD: 9 | Blood collection before initiation treatment | Serum | W6/32 + FG2/2 (anti-β2m) | sHLA-I | 0.89 ± 0.08 ug/mL ** | None found | ↑ | No statistical differences between patients in remission and normal controls [20] |
| Multiple myeloma (MM) | Healthy: 96; MM: 103 | Chemotherapy naive: 44. ≥1 round of systemic anti-myeloma treatment ≥19 days ago: 59 | Serum | W6/32 + anti-β2M | sHLA-I | 261 (44–1010) µg/L *** | Higher sHLA-I in stage III compared to stage II. sHLA-I is an independent prognostic factor for OS | ↑ | [21] |
| MM and monoclonal gammopathy of undetermined significance (MGUS) | Healthy: 30, MGUS: 30, MM: 103 | After collection. Conventional chemotherapy (53), high-dose therapy with autologous PBSC support (38), untreated (12) | Serum | W6/32 (sHLA-I) + anti-β2M | sHLA-I | 1.04 ± 0.92 ug/mL * | Higher in MM stages II–III compared to stage I, above cut off predicted shorter OS | ↑ | sHLA-I higher in more advanced stages [22] |
| Waldenstrom macroglobulinemia (WM) and IgM-MGUS | Healthy: 41, IgM-MGUS: 63, WM: 42 | After collection. WM patients (19/42) received alkylating agents (15) or fludarabine (4) ± rituximab | Serum | W6/32 (sHLA-I) + anti-β2M | sHLA-I | 0.69 (0.4–0.8) ug/mL **** | Higher median sHLA-I in WM patients with adverse prognosis markers | ↑ | sHLA-I higher in WM [23] |
| Acute myeloid leukemia (AML) | Healthy: 12, AML: 20 | Blood collection before initiation chemotherapy | Serum | W6/32 + TP25.99 | sHLA-I (minus sHLA-G) | Range: 0.1–0.6 ug/mL | N.A. | ↑ | [24] |
| AML and advanced myelodysplastic syndrome (MDS) | Healthy: ?, AML: 209, MDS: 98 | Treatment naive | Plasma | W6/32 + anti-β2M | sHLA-I | Unknown | AML: correlation with survival, complete response and cumulative response duration. MDS: none found | ↑ | Earlier transfusion or growth factor administration at earlier points could not be ruled out [16] |
| Disease | N for sHLA Measurements | Treatment | Sample Type | Antibodies | sHLA Type | sHLA Levels Normal Controls | Correlation with Progression/Prognosis | sHLA Compared to Normal Controls | Notes |
|---|---|---|---|---|---|---|---|---|---|
| Solid tumors | |||||||||
| Papillary thyroid carcinoma (PTC) | Healthy: 80, PTC: 85 | Unknown | Plasma | MEM-G/9 + anti-β2M | sHLA-G | 3.51 (0–10.60) ng/mL **** | Decreased in patients with invasion. No difference before/after thyroidectomy. | ↓ | Tendency towards lower sHLA-G, not statistically different [41]. |
| Thyroid cancer (TC) | Healthy: 45, TC: 40 | Unknown | Serum | ELISA kit (Bio-Vendor) | sHLA-G | Unknown | N.A. | ↓ | Significantly lower serum HLA-G positivity [40]. |
| Colorectal cancer (CRC) | Healthy: 113, CRC: 178 | Treatment naive | Plasma | ELISA kit (Exbio) | sHLA-G1 and -G5 | 25.4 (3.6–97.1) U/mL *** | Higher in dead than alive patients. Above cut off associated with worse prognosis. | ↑ | [32]. |
| Colorectal cancer (CRC) | Normal: 60, hyperplastic polyp: 72, IBD: 57, adenoma: 65, CRC: 144 | Treatment naive | Serum | ELISA kit (Exbio) | sHLA-G1 and -G5 | 25.0 (13.8–39.5) U/mL **** | None found. | ↑ | Higher sHLA-G in all diseases, highest in CRC [35]. |
| Colorectal cancer (CRC) | Mucinous carcinoma: 16, adenocarcinoma: 117 | Unknown | Plasma | ELISA kit (Exbio) | sHLA-G1 and -G5 | N.A. | Stage II: Negative correlation sHLA-G and liver metastasis free survival (LMFS). Stage III: Positive correlation sHLA-G and LMFS. | N.A. | [43]. |
| Colorectal cancer (CRC) | Healthy: 10, CRC: 20 | No prior treatment for CRC | Saliva and serum | ELISA kit (Bio-Vendor) | sHLA-G1 and –G5 | Saliva: 6.3 U/mL, (3.4–10.4 U/mL) ^. Serum: undetermined | Higher saliva and serum sHLA-G levels in stage III–IV than stage I–II. | ↑ | Higher serum sHLA-G in age group >70 years [34]. |
| Metastatic colorectal cancer (mCRC) | mCRC: 40 | Untreated | Plasma | ELISA kit (Bio-Vendor) | sHLA-G1 and -G5 | N.A. | Higher sHLA-G levels in patients with more than 1 metastatic site. | N.A. | [49] |
| Gastric cancer and colorectal cancer (CRC)) | Healthy: 45, gastrointestinal cancer: 82 | ≈ 50% pre-operative chemo- or radiotherapy | Plasma | ELISA kit (Exbio) | sHLA-G1 and -G5 | 56.99 ± 48.45 U/mL ** | Correlation between increased sHLA-G and stage I tumors. | ↑ | [50]. |
| Gastric cancer (GC) | Healthy: 77, Benign disease: 53, GC: 81 | No preoperative chemotherapy or radiotherapy | EDTA plasma | ELISA kit (Exbio) | sHLA-G1 and -G5 | 29.4 (24.0–39.3) U/mL **** | None found. | ↑ | Also higher sHLA-G levels in cancer compared to benign disease [36]. |
| Hepatocellular carcinoma (HCC) | Healthy: 25, liver cirrhosis: 25, HCC: 36 | Liver cirrhosis treatment naive, HCC unknown | Serum | ELISA kit (Bio-Vendor) | sHLA-G | 47.0 ± 15.5 U/mL * | N.A. | ↑ | Also higher sHLA-G levels in cancer compared to liver cirrhosis [37]. |
| Prostate cancer (PC) | Healthy: 26, Benign PC: 26, Malignant PC: 26 | Unknown | Serum | ELISA kit (Elab-science Bio-technology) | sHLA-G | 0.83 ng/mL # | A trend towards higher levels in more advanced stage. | ↑ | A statistical difference observed between the three groups, but no apparent difference between benign/malignant PC [38]. |
| Endometrial cancer (EC) | Healthy: 45, EC: 40 | Treatment naive | Plasma | ELISA kit (Exbio) | sHLA-G1 and -G5 | 0.145 ± 0.102 ng/mL ** | Higher sHLA-G in high grade disease than low grade (trend). | ↑ | Indicated as trend, as borderline significant (p = 0.057) [45]. |
| Ovarian pathologies (including ovarian cancer) | Benign serous ovarian cysts (G1): 54, Endometriosis (G2): 43, Ovarian cancer (G3): 38 | Before chemotherapy: 31, after: 7 (ovarian cancer patients) | Perito-neal fluid (PF) and serum | ELISA kit (Bio-Vendor) | sHLA-G | N.A. | PF sHLA-G concentrations and the difference between sHLA-G in serum and PF can differentiate G2 and G3. | N.A. | No significant differences in both PF and serum sHLA-G levels between groups. The median difference between PF and serum sHLA-G levels significantly higher in ovarian cancer patients [39]. |
| (Neoadjuvant-treated) breast cancer (BC) | Healthy: 16, BC: 190 | All undergoing neoadjuvant chemotherapy | EDTA plasma | MEM-G/9 and anti-β2M | sHLA-G | 16.3 (4.0–37.8) ng/mL *** | Before therapy: sHLA-Gev above cut off associated with worse PFS, sHLA-Gfree above cut off with better PFS and OS. | ↑ | [51]. |
| Breast cancer (BC) | Healthy: 84, BC: 75 | Most patients received chemo-, radio- or hormone therapy | Plasma | ELISA kit (Qayee Biological Technology Co., Ltd.) | sHLA-G | Median: 22.71 ng/mL. Mean: 31.58 ± 28.60 ng/mL | sHLA-G levels could identify mastectomized patients. | ↓ | 61 patients were mastectomized [44]. |
| Head and neck squamous cell carcinoma (HNSCC) | Healthy: 99, HNSCC: 120 | Treatment naive | Serum | SPR: Mem-G/9, ELISA: kit (Cloud Clone Corp.) | sHLA-G | ELISA: 2.47 ± 0.10 ng/mL * | Seemingly higher sHLA-G in advanced stages. | ↑ | Surface plasmon resonance (SPR) and ELISA [33]. |
| Neuroblastoma (NB) | Healthy: 13, NB: 31 | Unknown | Bone marrow plasma | ELISA kit (Exbio) | sHLA-G | 25.16 ± 7.38 ng/mL $ | sHLA-G higher in stage 3–4 than stage 1–2. | __ | [46]. |
| Melanoma | Healthy: 126, Melanoma: 190 | Cytostatics (24), IFN-α (31), or treatment naive (135) | Serum | ELISA: W6/32 + anti-β2M | sHLA-G | 22.92 ± 1.51 ng/mL ** | None found. | ↑ | Prior to ELISA serum depletion with TP25.99. IFN-α treatment was the only factor of impact on increased sHLA-G levels [52]. |
| Hematological tumors | |||||||||
| Acute lymphoblastic leukemia (ALL) | Healthy: 14, ALL: 33 | Before/after intensification phase of chemo-therapy (IPC) | Serum | ELISA kit (Biovendor) | sHLA-G | 31.0 ± 21.1 U/mL * | None found. | ↑ | Before IPC non-significantly increased, after IPC significantly increased [47]. |
| T cell acute lymphoblastic leukemia (T-ALL) | T-ALL: 32 | Chemotherapy | Bone marrow (BM) aspirates | ELISA kit (Biovendor) | sHLA-G | N.A. | Association with leukocytosis at diagnosis. Not related to survival. | N.A. | Patients’ age: 2–16 years. sHLA-G levels did not change during treatment [53]. |
| Acute myeloid leukemia (AML) | Healthy: 15, AML: 30 | Unknown for newly diagnosed patients, conventional chemotherapy for relapsed patients | Serum | ELISA kit (Glory Science) | sHLA-G1 and -G5 | 329.8 ± 57.54 ng/mL * | Higher in relapsed patients than newly diagnosed. patients and related to bone marrow blasts percentage. | ↑ | Newly diagnosed: 15, Relapsed: 15 [54]. |
| Chronic myeloid leukemia (CML) | CML: 68 | Imatinib, nilotinib or dasatinib | EDTA plasma | ELISA kit (Bio-Vendor) | sHLA-G1 and -G5 | N.A. | High secretor haplotype is associated with lower event-free survival during TKI treatment. | N.A. | Higher sHLA-G in high secretor haplotype (HLA-G*01:01:01/02) than low secretor haplotyope (HLA-G*01:01:03) [55]. |
| Waldenstrom macroglobulinemia (WM) and IgM-MGUS | Healthy: 41, IgM-MGUS: 63, WM: 42 | After serum collection. WM patients (19/42) received alkylating agents (15) or fludarabine (4) ± rituximab | Serum | MEM-G/9 (sHLA-G) + anti-β2M | sHLA-G | 17.3 (12–21) ng/mL **** | None found. | ↑ | sHLA-G higher in WM and IgM-MGUS [23]. |
| Disease | N for sHLA Measurements | Treatment | Sample Type | Antibodies | sHLA Type | sHLA Levels Normal Controls | Correlation with Progression/Prognosis | sHLA Levels Compared to Normal Controls | Notes |
|---|---|---|---|---|---|---|---|---|---|
| Solid tumors | |||||||||
| Melanoma | Healthy: 94, Melanoma: 127 | No current therapy | Serum | MEM-E/08 + MEM-E/07 | sHLA-E | 0 (0–1224) pg/mL *** | None found. | ↑ | Mostly higher in stage III [56]. |
| Neuroblastoma (NB) | Healthy: 75, NB: 84 | Unknown | Plasma | sHLA-E: 3D12 + anti-β2M, sHLA-F: polyclonal anti-HLA-F + anti-β2M | sHLA-E and sHLA-F | sHLA-E: 0.07 ± 0.02 units/mL * and sHLA-F: 0.1 ± 0.03 units/mL * | Higher sHLA-E and -F levels (above cut off) associated with better EFS and OS. | ↑ | [48]. |
| Neuroblastoma (NB) | Healthy: 13, NB: 31 | Unknown | Bone marrow plasma | 3D12 + anti-β2M (Exbio) | sHLA-E | 48.01 ± 10.93 U/mL $ | sHLA-E higher in stage 3–4 than stage 1–2. | ↓ | Tendency towards lower sHLA-E, not statistically different [46]. |
| Nasopharyngeal carcinoma (NPC) | Healthy: 65, NPC: 81 | Unknown | Plasma | Commercial kit (TSZ) | sHLA-F | 10.06 (3.22–74.05) pg/mL ^ | N.A. | ↑ | Tendency towards higher sHLA-F, not statistically different [57]. |
| Hematological tumors | |||||||||
| Chronic lymphocytic leukemia (CLL) | Healthy: 35, CLL: 50 | Chemotherapy naive | Plasma | 3D12 + MEM/07 | sHLA-E | 1222 ± 101 pg/mL * | HLA-E*01:03 is an independent predictor for disease progression. | ___ | No difference between healthy and CLL, but significantly higher in stage C than stage A. sHLA decreases after therapy [58]. |
| Acute leukemia (AL) | Healthy: 64, AL: 54 | Unknown | Plasma | Commercial kit (Shang-hai Jianglai Biotech. Co. Ltd.) | sHLA-E | 18.34 pg/mL # | N.A. | ↑ | [59]. |
| Disease | N for sHLA Measurements | Treatment | Sample Type | Antibodies | sHLA Type | sHLA Levels Normal Controls | Correlation with Progression/Prognosis | sHLA Levels Compared to Normal Controls | Notes |
|---|---|---|---|---|---|---|---|---|---|
| Solid tumors | |||||||||
| Malignant melanoma | Healthy (random): 86, Healthy (old): 35, Melanoma: 183 | Cytostatic drugs, immunomodulatory agents or nothing. | Serum | 1.243 + CR3/43 | sHLA-DR | Healthy (random): 1.38 ± 0.16 ug/mL; Healthy (old): 1.519 ± 0.1 ug/mL ** | Independent predictor of progression-free survival. | ↓ | [60]. |
| Hematological tumors | |||||||||
| Pre-B acute lymphoblastic leukemia (ALL) | Healthy: 31, ALL; 30 | Treatment naive | Plasma | Polyclonal rabbit HLA-DRB1 + mAb anti-HLA-DR clone L243 | sHLA-DRB1 | 0.051 ± 0.007 ug/mL ** | N.A. | ↑ | [61]. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kessler, A.L.; Bruno, M.J.; Buschow, S.I. The Potential of Soluble Human Leukocyte Antigen Molecules for Early Cancer Detection and Therapeutic Vaccine Design. Vaccines 2020, 8, 775. https://doi.org/10.3390/vaccines8040775
Kessler AL, Bruno MJ, Buschow SI. The Potential of Soluble Human Leukocyte Antigen Molecules for Early Cancer Detection and Therapeutic Vaccine Design. Vaccines. 2020; 8(4):775. https://doi.org/10.3390/vaccines8040775
Chicago/Turabian StyleKessler, Amy L., Marco J. Bruno, and Sonja I. Buschow. 2020. "The Potential of Soluble Human Leukocyte Antigen Molecules for Early Cancer Detection and Therapeutic Vaccine Design" Vaccines 8, no. 4: 775. https://doi.org/10.3390/vaccines8040775