Lactobacilli Expressing Broadly Neutralizing Nanobodies against HIV-1 as Potential Vectors for HIV-1 Prophylaxis?

 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression of Recombinant Soluble optC gp140 (optC.664) SOSIP Proteins
2.2. Expression of VHH Proteins in E. coli
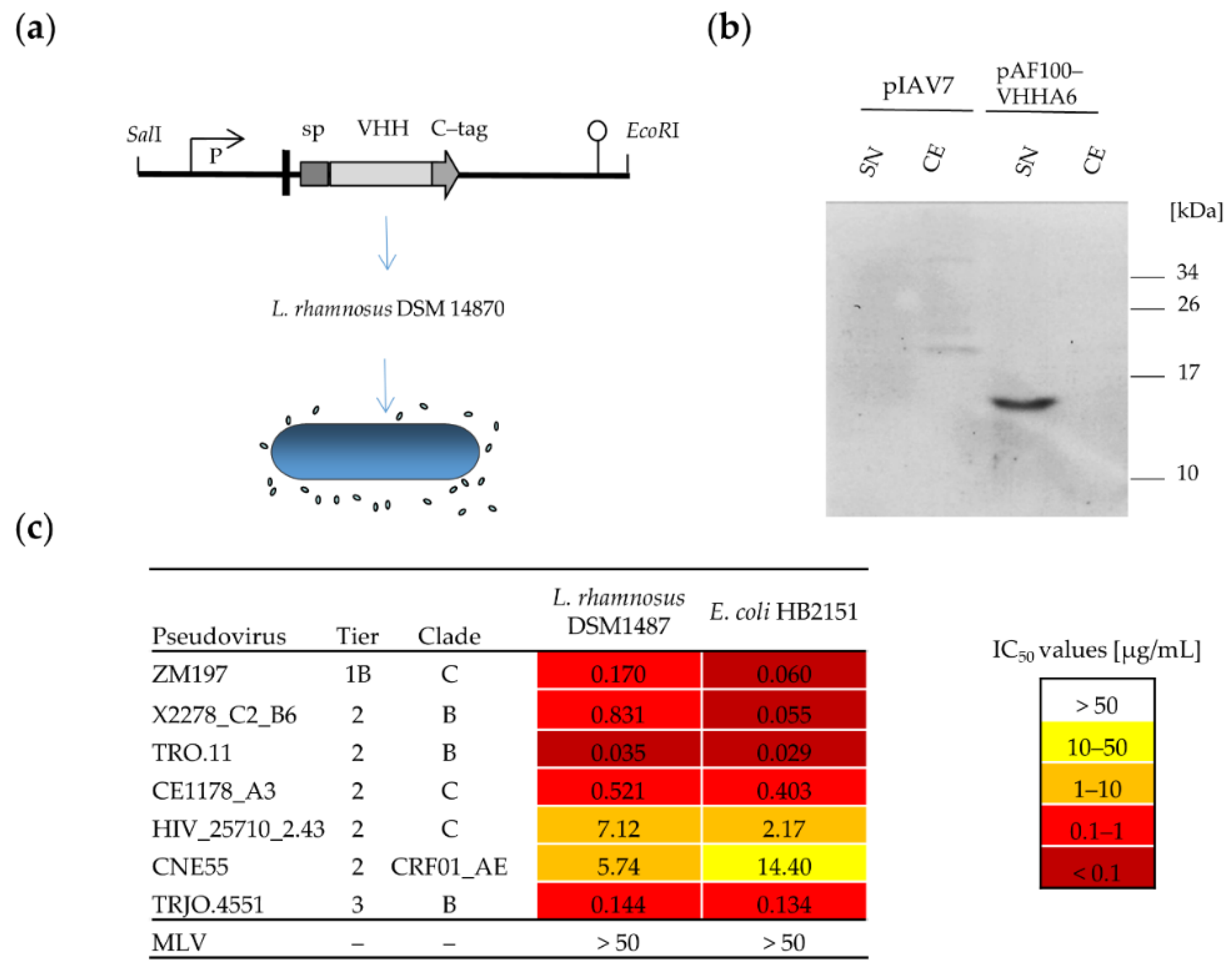
2.3. Cloning and Expression of VHH Proteins in L. rhamnosus
2.4. Expression of Recombinant VHH-Fc Proteins
2.5. Acidic pH ELISA
2.6. Neutralization Assays for Soluble Nanobodies
2.7. Infectivity Depletion Assay for Cell Surface-Anchored Nanobodies
2.8. Production of pCHIV Pseudovirus
2.9. Analysis of Lactobacilli by Flow Cytometry
2.10. p24 ELISA
2.11. Mouse Experiments
3. Results
3.1. Functionality of Nanobody VHHA6 Expressed from E. coli
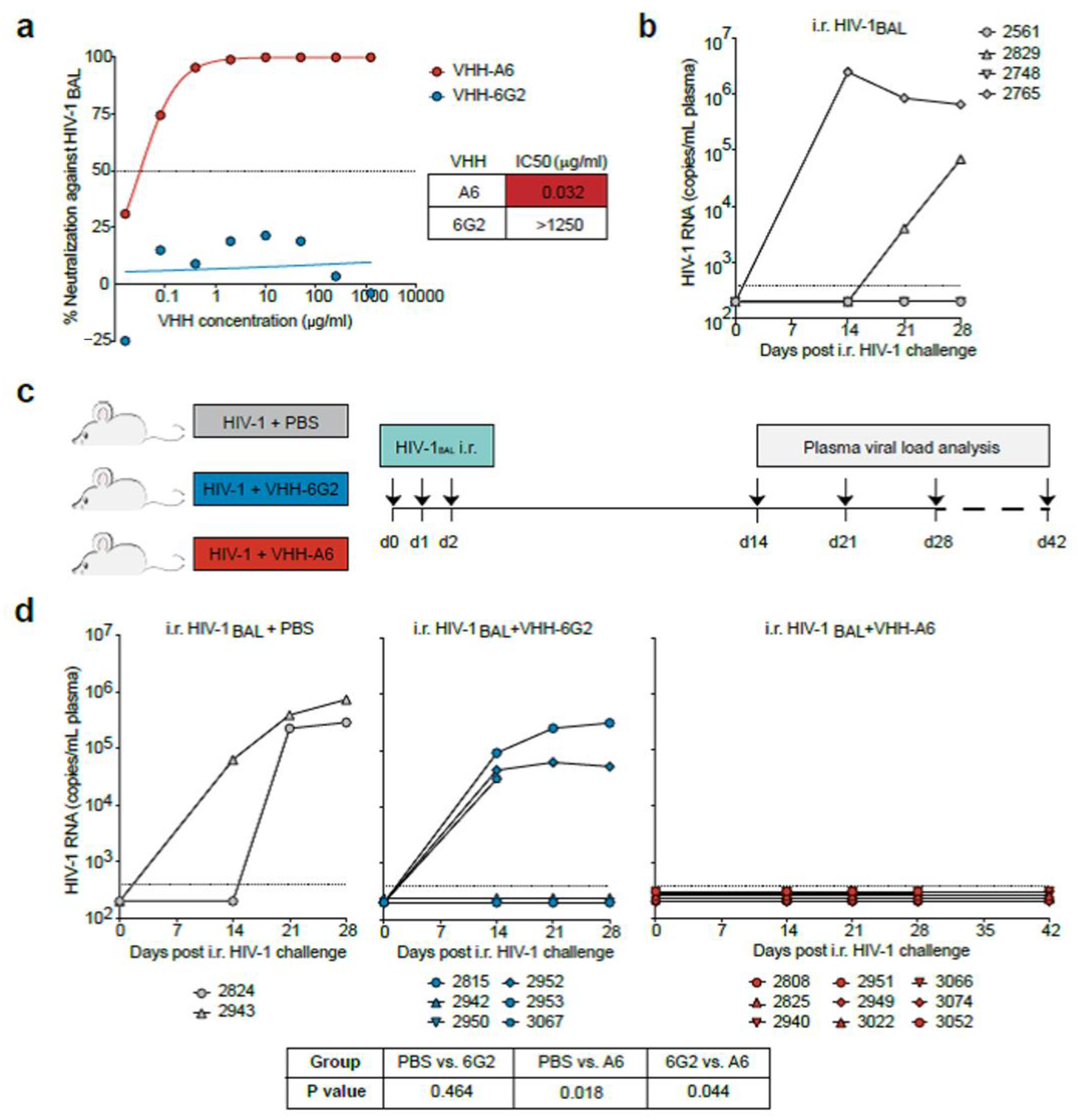
3.1.1. Determination of In Vivo Half-Life of VHHA6 in Sera of Mice
3.1.2. Soluble VHHA6 Blocks Infection of Humanized Mice by HIV-1
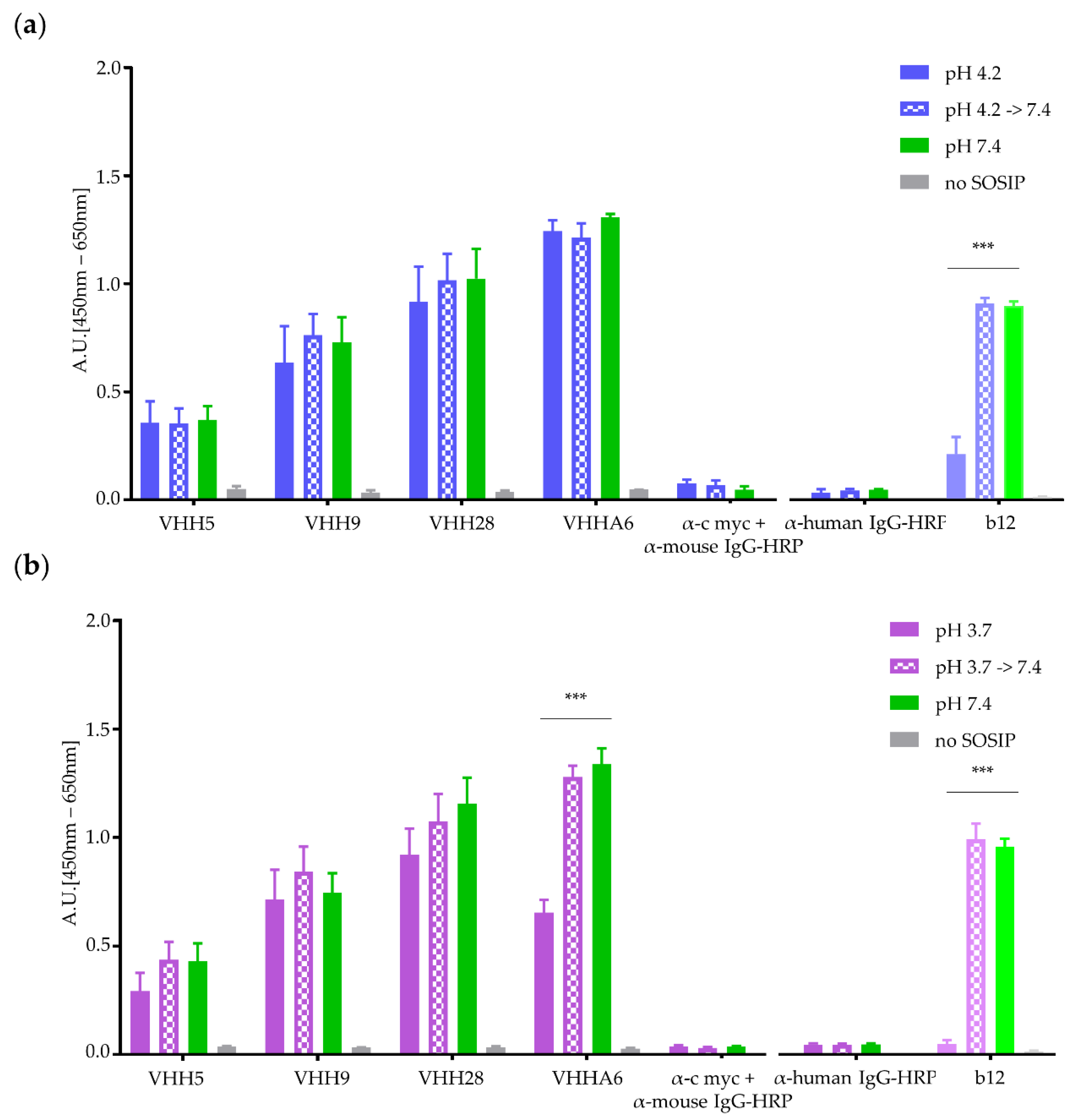
3.1.3. Nanobodies Retain Functionality at Acidic pH
3.2. Functionality of Nanobody VHHA6 Expressed from L. rhamnosus DSM 14870
3.2.1. Soluble VHHA6 Expressed from L. rhamnosus Retains Neutralizing Activity against HIV-1
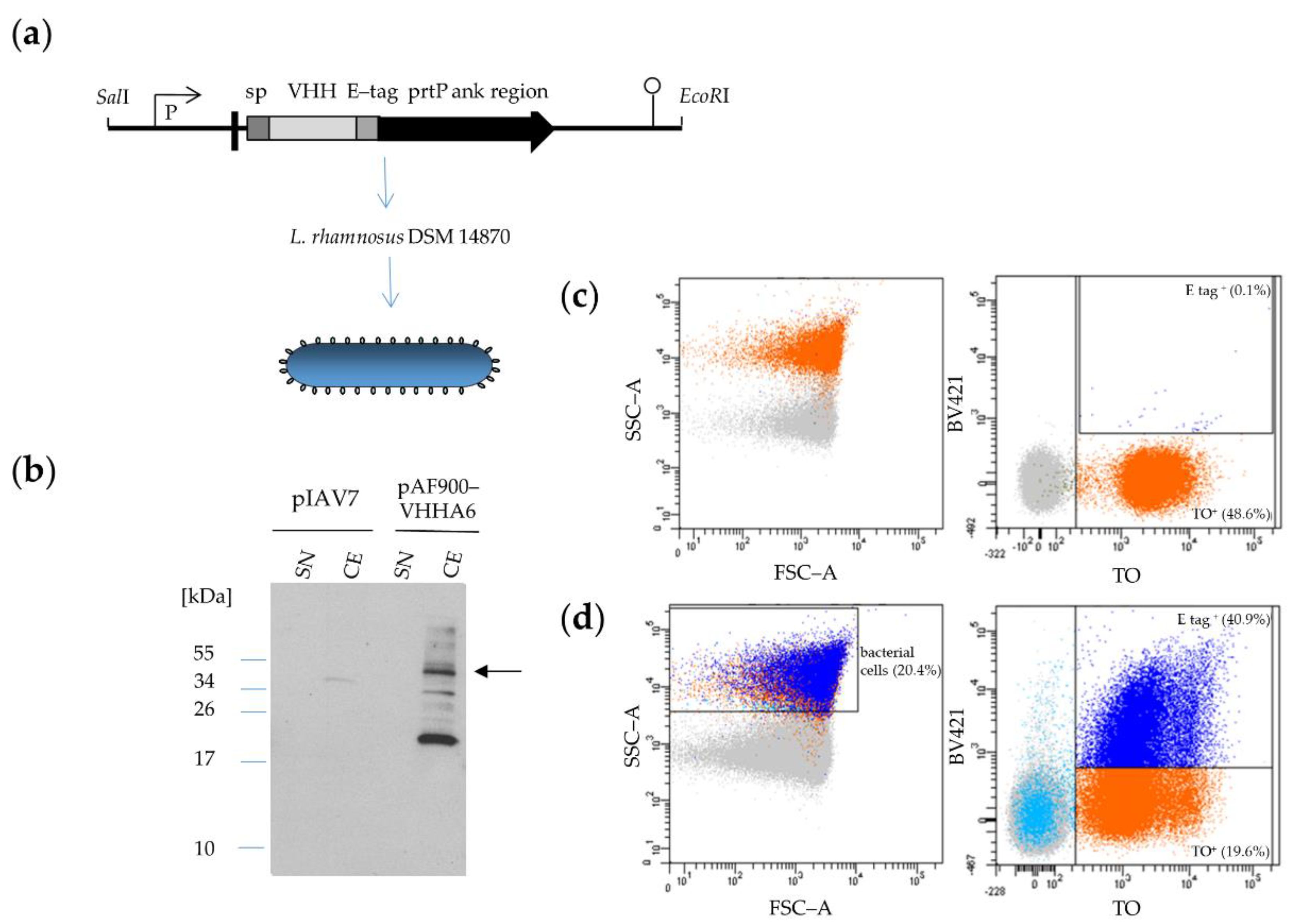
3.2.2. Expression of VHHA6 Anchored on the Surface of L. rhamnosus DSM 14870 pAF900-VHHA6
Lactobacilli with Surface-Anchored VHHA6 Bind GFP-Labeled HIV-1 Virions
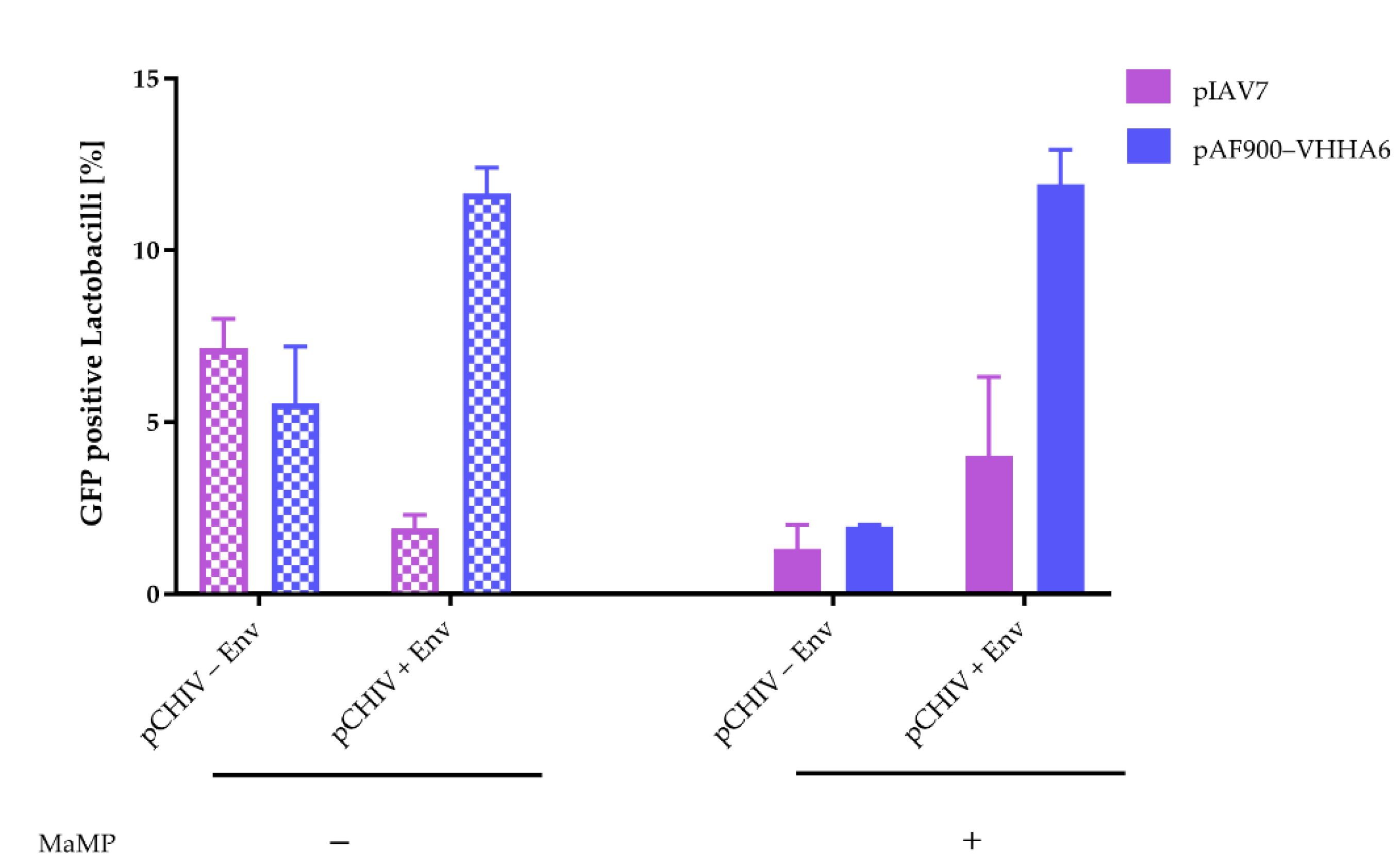
Lactobacilli with Surface-Anchored VHHA6 Neutralize HIV-1 in an Infectivity Depletion Assay
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- UNAIDS Fact Sheet. 2020. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 10 November 2020).
- Cohen, M.S.; Baden, L.R. Preexposure prophylaxis for HIV--where do we go from here? N. Engl. J. Med. 2012, 367, 459–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stephenson, K.E.; Wagh, K.; Korber, B.; Barouch, D.H. Vaccines and Broadly Neutralizing Antibodies for HIV-1 Prevention. Annu. Rev. Immunol. 2020, 38, 673–703. [Google Scholar] [CrossRef] [PubMed]
- Bar, K.J.; Sneller, M.C.; Harrison, L.J.; Justement, J.S.; Overton, E.T.; Petrone, M.E.; Salantes, D.B.; Seamon, C.A.; Scheinfeld, B.; Kwan, R.W.; et al. Effect of HIV Antibody VRC01 on Viral Rebound after Treatment Interruption. N. Engl. J. Med. 2016, 375, 2037–2050. [Google Scholar] [CrossRef] [PubMed]
- Caskey, M.; Klein, F.; Lorenzi, J.C.; Seaman, M.S.; West, A.P., Jr.; Buckley, N.; Kremer, G.; Nogueira, L.; Braunschweig, M.; Scheid, J.F.; et al. Viraemia suppressed in HIV-1-infected humans by broadly neutralizing antibody 3BNC117. Nature 2015, 522, 487–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caskey, M.; Schoofs, T.; Gruell, H.; Settler, A.; Karagounis, T.; Kreider, E.F.; Murrell, B.; Pfeifer, N.; Nogueira, L.; Oliveira, T.Y.; et al. Antibody 10-1074 suppresses viremia in HIV-1-infected individuals. Nat. Med. 2017, 23, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Gautam, R.; Nishimura, Y.; Pegu, A.; Nason, M.C.; Klein, F.; Gazumyan, A.; Golijanin, J.; Buckler-White, A.; Sadjadpour, R.; Wang, K.; et al. A single injection of anti-HIV-1 antibodies protects against repeated SHIV challenges. Nature 2016, 533, 105–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynch, R.M.; Wong, P.; Tran, L.; O’Dell, S.; Nason, M.C.; Li, Y.; Wu, X.; Mascola, J.R. HIV-1 fitness cost associated with escape from the VRC01 class of CD4 binding site neutralizing antibodies. J. Virol. 2015, 89, 4201–4213. [Google Scholar] [CrossRef] [Green Version]
- Moldt, B.; Rakasz, E.G.; Schultz, N.; Chan-Hui, P.Y.; Swiderek, K.; Weisgrau, K.L.; Piaskowski, S.M.; Bergman, Z.; Watkins, D.I.; Poignard, P.; et al. Highly potent HIV-specific antibody neutralization in vitro translates into effective protection against mucosal SHIV challenge in vivo. Proc. Natl. Acad. Sci. USA 2012, 109, 18921–18925. [Google Scholar] [CrossRef] [Green Version]
- Scheid, J.F.; Horwitz, J.A.; Bar-On, Y.; Kreider, E.F.; Lu, C.L.; Lorenzi, J.C.; Feldmann, A.; Braunschweig, M.; Nogueira, L.; Oliveira, T.; et al. HIV-1 antibody 3BNC117 suppresses viral rebound in humans during treatment interruption. Nature 2016, 535, 556–560. [Google Scholar] [CrossRef] [Green Version]
- Schommers, P.; Gruell, H.; Abernathy, M.E.; Tran, M.K.; Dingens, A.S.; Gristick, H.B.; Barnes, C.O.; Schoofs, T.; Schlotz, M.; Vanshylla, K.; et al. Restriction of HIV-1 Escape by a Highly Broad and Potent Neutralizing Antibody. Cell 2020, 180, 471–489.e22. [Google Scholar] [CrossRef] [Green Version]
- Sok, D.; Burton, D.R. Recent progress in broadly neutralizing antibodies to HIV. Nat. Immunol. 2018, 19, 1179–1188. [Google Scholar] [CrossRef] [PubMed]
- Balazs, A.B.; Chen, J.; Hong, C.M.; Rao, D.S.; Yang, L.; Baltimore, D. Antibody-based protection against HIV infection by vectored immunoprophylaxis. Nature 2011, 481, 81–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garber, D.A.; Adams, D.R.; Guenthner, P.; Mitchell, J.; Kelley, K.; Schoofs, T.; Gazumyan, A.; Nason, M.; Seaman, M.S.; McNicholl, J.; et al. Durable protection against repeated penile exposures to simian-human immunodeficiency virus by broadly neutralizing antibodies. Nat. Commun 2020, 11, 3195. [Google Scholar] [CrossRef] [PubMed]
- Julg, B.; Barouch, D.H. Neutralizing antibodies for HIV-1 prevention. Curr. Opin. HIV AIDS 2019, 14, 318–324. [Google Scholar] [CrossRef]
- Pegu, A.; Borate, B.; Huang, Y.; Pauthner, M.G.; Hessell, A.J.; Julg, B.; Doria-Rose, N.A.; Schmidt, S.D.; Carpp, L.N.; Cully, M.D.; et al. A Meta-analysis of Passive Immunization Studies Shows that Serum-Neutralizing Antibody Titer Associates with Protection against SHIV Challenge. Cell Host Microbe 2019, 26, 336–346.e3. [Google Scholar] [CrossRef]
- Shingai, M.; Donau, O.K.; Plishka, R.J.; Buckler-White, A.; Mascola, J.R.; Nabel, G.J.; Nason, M.C.; Montefiori, D.; Moldt, B.; Poignard, P.; et al. Passive transfer of modest titers of potent and broadly neutralizing anti-HIV monoclonal antibodies block SHIV infection in macaques. J. Exp. Med. 2014, 211, 2061–2074. [Google Scholar] [CrossRef]
- Bar-On, Y.; Gruell, H.; Schoofs, T.; Pai, J.A.; Nogueira, L.; Butler, A.L.; Millard, K.; Lehmann, C.; Suarez, I.; Oliveira, T.Y.; et al. Safety and antiviral activity of combination HIV-1 broadly neutralizing antibodies in viremic individuals. Nat. Med. 2018, 24, 1701–1707. [Google Scholar] [CrossRef]
- Caskey, M.; Klein, F.; Nussenzweig, M.C. Broadly neutralizing anti-HIV-1 monoclonal antibodies in the clinic. Nat. Med. 2019, 25, 547–553. [Google Scholar] [CrossRef]
- Mendoza, P.; Gruell, H.; Nogueira, L.; Pai, J.A.; Butler, A.L.; Millard, K.; Lehmann, C.; Suarez, I.; Oliveira, T.Y.; Lorenzi, J.C.C.; et al. Combination therapy with anti-HIV-1 antibodies maintains viral suppression. Nature 2018, 561, 479–484. [Google Scholar] [CrossRef]
- Klein, F.; Halper-Stromberg, A.; Horwitz, J.A.; Gruell, H.; Scheid, J.F.; Bournazos, S.; Mouquet, H.; Spatz, L.A.; Diskin, R.; Abadir, A.; et al. HIV therapy by a combination of broadly neutralizing antibodies in humanized mice. Nature 2012, 492, 118–122. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, P.B.; Juraska, M.; deCamp, A.C.; Karuna, S.; Edupuganti, S.; Mgodi, N.; Donnell, D.J.; Bentley, C.; Sista, N.; Andrew, P.; et al. Basis and Statistical Design of the Passive HIV-1 Antibody Mediated Prevention (AMP) Test-of-Concept Efficacy Trials. Stat. Commun. Infect. Dis. 2017, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- National Institutes of Health (NIH), U.S; National Library of Medicine. Clinical Trials. Available online: https://www.clinicaltrials.gov (accessed on 10 November 2020).
- Morris, G.C.; Wiggins, R.C.; Woodhall, S.C.; Bland, J.M.; Taylor, C.R.; Jespers, V.; Vcelar, B.A.; Lacey, C.J. MABGEL 1: First phase 1 trial of the anti-HIV-1 monoclonal antibodies 2F5, 4E10 and 2G12 as a vaginal microbicide. PLoS ONE 2014, 9, e116153. [Google Scholar] [CrossRef] [PubMed]
- Muyldermans, S. Generation and design of nanobodies. FEBS J. 2020. [Google Scholar] [CrossRef] [PubMed]
- Wesolowski, J.; Alzogaray, V.; Reyelt, J.; Unger, M.; Juarez, K.; Urrutia, M.; Cauerhff, A.; Danquah, W.; Rissiek, B.; Scheuplein, F.; et al. Single domain antibodies: Promising experimental and therapeutic tools in infection and immunity. Med. Microbiol. Immunol. 2009, 198, 157–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Guan, Y. Immunologic Basis for Long HCDR3s in Broadly Neutralizing Antibodies Against HIV-1. Front. Immunol. 2014, 5, 250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCoy, L.E.; Quigley, A.F.; Strokappe, N.M.; Bulmer-Thomas, B.; Seaman, M.S.; Mortier, D.; Rutten, L.; Chander, N.; Edwards, C.J.; Ketteler, R.; et al. Potent and broad neutralization of HIV-1 by a llama antibody elicited by immunization. J. Exp. Med. 2012, 209, 1091–1103. [Google Scholar] [CrossRef] [Green Version]
- Koch, K.; Kalusche, S.; Torres, J.L.; Stanfield, R.L.; Danquah, W.; Khazanehdari, K.; von Briesen, H.; Geertsma, E.R.; Wilson, I.A.; Wernery, U.; et al. Selection of nanobodies with broad neutralizing potential against primary HIV-1 strains using soluble subtype C gp140 envelope trimers. Sci. Rep. 2017, 7, 8390. [Google Scholar] [CrossRef]
- Petrova, M.I.; van den Broek, M.; Balzarini, J.; Vanderleyden, J.; Lebeer, S. Vaginal microbiota and its role in HIV transmission and infection. FEMS Microbiol. Rev. 2013, 37, 762–792. [Google Scholar] [CrossRef] [Green Version]
- Welch, J.L.; Xiang, J.; Okeoma, C.M.; Schlievert, P.M.; Stapleton, J.T. Glycerol Monolaurate, an Analogue to a Factor Secreted by Lactobacillus, Is Virucidal against Enveloped Viruses, Including HIV-1. mBio 2020, 11. [Google Scholar] [CrossRef]
- Gosmann, C.; Anahtar, M.N.; Handley, S.A.; Farcasanu, M.; Abu-Ali, G.; Bowman, B.A.; Padavattan, N.; Desai, C.; Droit, L.; Moodley, A.; et al. Lactobacillus-Deficient Cervicovaginal Bacterial Communities Are Associated with Increased HIV Acquisition in Young South African Women. Immunity 2017, 46, 29–37. [Google Scholar] [CrossRef] [Green Version]
- Kaul, R.; Pettengell, C.; Sheth, P.M.; Sunderji, S.; Biringer, A.; MacDonald, K.; Walmsley, S.; Rebbapragada, A. The genital tract immune milieu: An important determinant of HIV susceptibility and secondary transmission. J. Reprod. Immunol. 2008, 77, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, B.; Krogh-Andersen, K.; Tellgren-Roth, C.; Martinez, N.; Gunaydin, G.; Lin, Y.; Martin, M.C.; Alvarez, M.A.; Hammarstrom, L.; Marcotte, H. An Exopolysaccharide-Deficient Mutant of Lactobacillus rhamnosus GG Efficiently Displays a Protective Llama Antibody Fragment against Rotavirus on Its Surface. Appl. Environ. Microbiol. 2015, 81, 5784–5793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, K.K.; Strokappe, N.M.; Hultberg, A.; Truusalu, K.; Smidt, I.; Mikelsaar, R.H.; Mikelsaar, M.; Verrips, T.; Hammarstrom, L.; Marcotte, H. Neutralization of Clostridium Difficile Toxin B Mediated by Engineered Lactobacilli That Produce Single-Domain Antibodies. Infect. Immun. 2016, 84, 395–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, T.L.; Chang, C.H.; Simpson, D.A.; Xu, Q.; Martin, P.K.; Lagenaur, L.A.; Schoolnik, G.K.; Ho, D.D.; Hillier, S.L.; Holodniy, M.; et al. Inhibition of HIV infectivity by a natural human isolate of Lactobacillus jensenii engineered to express functional two-domain CD4. Proc. Natl. Acad. Sci. USA 2003, 100, 11672–11677. [Google Scholar] [CrossRef] [Green Version]
- Gunaydin, G.; Alvarez, B.; Lin, Y.; Hammarstrom, L.; Marcotte, H. Co-expression of anti-rotavirus proteins (llama VHH antibody fragments) in Lactobacillus: Development and functionality of vectors containing two expression cassettes in tandem. PLoS ONE 2014, 9, e96409. [Google Scholar] [CrossRef] [Green Version]
- Kruger, C.; Hu, Y.; Pan, Q.; Marcotte, H.; Hultberg, A.; Delwar, D.; van Dalen, P.J.; Pouwels, P.H.; Leer, R.J.; Kelly, C.G.; et al. In situ delivery of passive immunity by lactobacilli producing single-chain antibodies. Nat. Biotechnol. 2002, 20, 702–706. [Google Scholar] [CrossRef]
- Lagenaur, L.A.; Sanders-Beer, B.E.; Brichacek, B.; Pal, R.; Liu, X.; Liu, Y.; Yu, R.; Venzon, D.; Lee, P.P.; Hamer, D.H. Prevention of vaginal SHIV transmission in macaques by a live recombinant Lactobacillus. Mucosal Immunol. 2011, 4, 648–657. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Lagenaur, L.A.; Simpson, D.A.; Essenmacher, K.P.; Frazier-Parker, C.L.; Liu, Y.; Tsai, D.; Rao, S.S.; Hamer, D.H.; Parks, T.P.; et al. Engineered vaginal lactobacillus strain for mucosal delivery of the human immunodeficiency virus inhibitor cyanovirin-N. Antimicrob. Agents Chemother. 2006, 50, 3250–3259. [Google Scholar] [CrossRef] [Green Version]
- Marcobal, A.; Liu, X.; Zhang, W.; Dimitrov, A.S.; Jia, L.; Lee, P.P.; Fouts, T.R.; Parks, T.P.; Lagenaur, L.A. Expression of Human Immunodeficiency Virus Type 1 Neutralizing Antibody Fragments Using Human Vaginal Lactobacillus. AIDS Res. Hum. Retrovir. 2016, 32, 964–971. [Google Scholar] [CrossRef] [Green Version]
- Martin, M.C.; Pant, N.; Ladero, V.; Gunaydin, G.; Andersen, K.K.; Alvarez, B.; Martinez, N.; Alvarez, M.A.; Hammarstrom, L.; Marcotte, H. Integrative expression system for delivery of antibody fragments by lactobacilli. Appl. Environ. Microbiol. 2011, 77, 2174–2179. [Google Scholar] [CrossRef] [Green Version]
- Pant, N.; Hultberg, A.; Zhao, Y.; Svensson, L.; Pan-Hammarstrom, Q.; Johansen, K.; Pouwels, P.H.; Ruggeri, F.M.; Hermans, P.; Frenken, L.; et al. Lactobacilli expressing variable domain of llama heavy-chain antibody fragments (lactobodies) confer protection against rotavirus-induced diarrhea. J. Infect. Dis. 2006, 194, 1580–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandenbroucke, K.; de Haard, H.; Beirnaert, E.; Dreier, T.; Lauwereys, M.; Huyck, L.; Van Huysse, J.; Demetter, P.; Steidler, L.; Remaut, E.; et al. Orally administered L. lactis secreting an anti-TNF Nanobody demonstrate efficacy in chronic colitis. Mucosal Immunol. 2010, 3, 49–56. [Google Scholar] [CrossRef] [PubMed]
- Larsson, P.G.; Brandsborg, E.; Forsum, U.; Pendharkar, S.; Andersen, K.K.; Nasic, S.; Hammarstrom, L.; Marcotte, H. Extended antimicrobial treatment of bacterial vaginosis combined with human lactobacilli to find the best treatment and minimize the risk of relapses. BMC Infect. Dis. 2011, 11, 223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marcotte, H.; Andersen, K.K.; Lin, Y.; Zuo, F.; Zeng, Z.; Larsson, P.G.; Brandsborg, E.; Bronstad, G.; Hammarstrom, L. Characterization and complete genome sequences of L. rhamnosus DSM 14870 and L. gasseri DSM 14869 contained in the EcoVag((R)) probiotic vaginal capsules. Microbiol. Res. 2017, 205, 88–98. [Google Scholar] [CrossRef]
- Marcotte, H.; Larsson, P.G.; Andersen, K.K.; Zuo, F.; Mikkelsen, L.S.; Brandsborg, E.; Gray, G.; Laher, F.; Otwombe, K. An exploratory pilot study evaluating the supplementation of standard antibiotic therapy with probiotic lactobacilli in south African women with bacterial vaginosis. BMC Infect. Dis. 2019, 19, 824. [Google Scholar] [CrossRef]
- Pendharkar, S.; Brandsborg, E.; Hammarstrom, L.; Marcotte, H.; Larsson, P.G. Vaginal colonisation by probiotic lactobacilli and clinical outcome in women conventionally treated for bacterial vaginosis and yeast infection. BMC Infect. Dis. 2015, 15, 255. [Google Scholar] [CrossRef] [Green Version]
- Sanders, R.W.; Derking, R.; Cupo, A.; Julien, J.P.; Yasmeen, A.; de Val, N.; Kim, H.J.; Blattner, C.; de la Pena, A.T.; Korzun, J.; et al. A next-generation cleaved, soluble HIV-1 Env trimer, BG505 SOSIP.664 gp140, expresses multiple epitopes for broadly neutralizing but not non-neutralizing antibodies. PLoS Pathog. 2013, 9, e1003618. [Google Scholar] [CrossRef] [Green Version]
- Perez-Arellano, I.; Zuniga, M.; Perez-Martinez, G. Construction of compatible wide-host-range shuttle vectors for lactic acid bacteria and Escherichia coli. Plasmid 2001, 46, 106–116. [Google Scholar] [CrossRef]
- Montefiori, D.C. Measuring HIV neutralization in a luciferase reporter gene assay. Methods Mol. Biol. (Clifton N.J.) 2009, 485, 395–405. [Google Scholar]
- Lampe, M.; Briggs, J.A.; Endress, T.; Glass, B.; Riegelsberger, S.; Krausslich, H.G.; Lamb, D.C.; Brauchle, C.; Muller, B. Double-labelled HIV-1 particles for study of virus-cell interaction. Virology 2007, 360, 92–104. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.J.; Hatziioannou, T.; Zang, T.; Braaten, D.; Luban, J.; Goff, S.P.; Bieniasz, P.D. Envelope-dependent, cyclophilin-independent effects of glycosaminoglycans on human immunodeficiency virus type 1 attachment and infection. J. Virol. 2002, 76, 6332–6343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanshylla, K.; Held, K.; Eser, T.M.; Gruell, H.; Jain, K.; Weiland, D.; Muench, J.; Gruettner, B.; Geldmacher, C.; Klein, F. A novel humanized mouse model to study mucosal HIV-1 transmission and prevention. BioRxiv 2020, 2020. [Google Scholar] [CrossRef]
- Palmer, S.; Wiegand, A.P.; Maldarelli, F.; Bazmi, H.; Mican, J.M.; Polis, M.; Dewar, R.L.; Planta, A.; Liu, S.; Metcalf, J.A.; et al. New real-time reverse transcriptase-initiated PCR assay with single-copy sensitivity for human immunodeficiency virus type 1 RNA in plasma. J. Clin. Microbiol. 2003, 41, 4531–4536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navarre, W.W.; Schneewind, O. Surface proteins of gram-positive bacteria and mechanisms of their targeting to the cell wall envelope. Microbiol. Mol. Biol. Rev. 1999, 63, 174–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nygren, J.; Svanvik, N.; Kubista, M. The interactions between the fluorescent dye thiazole orange and DNA. Biopolymers 1998, 46, 39–51. [Google Scholar] [CrossRef]
- Lakhtin, V.M.; Aleshkin, V.A.; Lakhtin, M.V.; Afanas’ev, S.S.; Pospelova, V.V.; Shenderov, B.A. Lectins, adhesins, and lectin-like substances of lactobacilli and bifidobacteria. Vestn. Ross. Akad. Med. Nauk 2006, 1, 28–34. [Google Scholar]
- Gruell, H.; Klein, F. Antibody-mediated prevention and treatment of HIV-1 infection. Retrovirology 2018, 15, 73. [Google Scholar] [CrossRef] [Green Version]
- Walker, L.M.; Burton, D.R. Passive immunotherapy of viral infections: ‘super-antibodies’ enter the fray. Nat. Rev. Immunol. 2018, 18, 297–308. [Google Scholar] [CrossRef]
- Ledgerwood, J.E.; Coates, E.E.; Yamshchikov, G.; Saunders, J.G.; Holman, L.; Enama, M.E.; DeZure, A.; Lynch, R.M.; Gordon, I.; Plummer, S.; et al. Safety, pharmacokinetics and neutralization of the broadly neutralizing HIV-1 human monoclonal antibody VRC01 in healthy adults. Clin. Exp. Immunol. 2015, 182, 289–301. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.; Balazs, A.B. Adeno-associated virus gene delivery of broadly neutralizing antibodies as prevention and therapy against HIV-1. Retrovirology 2018, 15, 66. [Google Scholar] [CrossRef]
- Martinez-Navio, J.M.; Fuchs, S.P.; Pantry, S.N.; Lauer, W.A.; Duggan, N.N.; Keele, B.F.; Rakasz, E.G.; Gao, G.; Lifson, J.D.; Desrosiers, R.C. Adeno-Associated Virus Delivery of Anti-HIV Monoclonal Antibodies Can Drive Long-Term Virologic Suppression. Immunity 2019, 50, 567–575.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priddy, F.H.; Lewis, D.J.M.; Gelderblom, H.C.; Hassanin, H.; Streatfield, C.; LaBranche, C.; Hare, J.; Cox, J.H.; Dally, L.; Bendel, D.; et al. Adeno-associated virus vectored immunoprophylaxis to prevent HIV in healthy adults: A phase 1 randomised controlled trial. Lancet HIV 2019, 6, e230–e239. [Google Scholar] [CrossRef] [Green Version]
- van den Berg, F.T.; Makoah, N.A.; Ali, S.A.; Scott, T.A.; Mapengo, R.E.; Mutsvunguma, L.Z.; Mkhize, N.N.; Lambson, B.E.; Kgagudi, P.D.; Crowther, C.; et al. AAV-Mediated Expression of Broadly Neutralizing and Vaccine-like Antibodies Targeting the HIV-1 Envelope V2 Region. Mol. Ther. Methods Clin. Dev. 2019, 14, 100–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuchs, S.P.; Desrosiers, R.C. Promise and problems associated with the use of recombinant AAV for the delivery of anti-HIV antibodies. Mol. Ther. Methods Clin. Dev. 2016, 3, 16068. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardner, M.R.; Fetzer, I.; Kattenhorn, L.M.; Davis-Gardner, M.E.; Zhou, A.S.; Alfant, B.; Weber, J.A.; Kondur, H.R.; Martinez-Navio, J.M.; Fuchs, S.P.; et al. Anti-drug Antibody Responses Impair Prophylaxis Mediated by AAV-Delivered HIV-1 Broadly Neutralizing Antibodies. Mol. Ther. 2019, 27, 650–660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ablynx Clinical Programs. Available online: www.ablynx.com (accessed on 10 November 2020).
- Bartunek, J.; Barbato, E.; Heyndrickx, G.; Vanderheyden, M.; Wijns, W.; Holz, J.B. Novel antiplatelet agents: ALX-0081, a Nanobody directed towards von Willebrand factor. J. Cardiovasc. Transl. Res. 2013, 6, 355–363. [Google Scholar] [CrossRef] [PubMed]
- Cortez-Retamozo, V.; Lauwereys, M.; Hassanzadeh Gh, G.; Gobert, M.; Conrath, K.; Muyldermans, S.; De Baetselier, P.; Revets, H. Efficient tumor targeting by single-domain antibody fragments of camels. Int. J. Cancer 2002, 98, 456–462. [Google Scholar] [CrossRef]
- Jovcevska, I.; Muyldermans, S. The Therapeutic Potential of Nanobodies. BioDrugs 2020, 34, 11–26. [Google Scholar] [CrossRef] [Green Version]
- Steeland, S.; Vandenbroucke, R.E.; Libert, C. Nanobodies as therapeutics: Big opportunities for small antibodies. Drug Discov. Today 2016, 21, 1076–1113. [Google Scholar] [CrossRef]
- Ulrichts, H.; Silence, K.; Schoolmeester, A.; de Jaegere, P.; Rossenu, S.; Roodt, J.; Priem, S.; Lauwereys, M.; Casteels, P.; Van Bockstaele, F.; et al. Antithrombotic drug candidate ALX-0081 shows superior preclinical efficacy and safety compared with currently marketed antiplatelet drugs. Blood 2011, 118, 757–765. [Google Scholar] [CrossRef] [Green Version]
- Papadopoulos, K.P.; Isaacs, R.; Bilic, S.; Kentsch, K.; Huet, H.A.; Hofmann, M.; Rasco, D.; Kundamal, N.; Tang, Z.; Cooksey, J.; et al. Unexpected hepatotoxicity in a phase I study of TAS266, a novel tetravalent agonistic Nanobody(R) targeting the DR5 receptor. Cancer Chemother. Pharmacol. 2015, 75, 887–895. [Google Scholar] [CrossRef] [PubMed]
- Chancey, C.J.; Khanna, K.V.; Seegers, J.F.; Zhang, G.W.; Hildreth, J.; Langan, A.; Markham, R.B. Lactobacilli-expressed single-chain variable fragment (scFv) specific for intercellular adhesion molecule 1 (ICAM-1) blocks cell-associated HIV-1 transmission across a cervical epithelial monolayer. J. Immunol. 2006, 176, 5627–5636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangelista, L.; Secchi, M.; Liu, X.; Bachi, A.; Jia, L.; Xu, Q.; Lusso, P. Engineering of Lactobacillus jensenii to secrete RANTES and a CCR5 antagonist analogue as live HIV-1 blockers. Antimicrob. Agents Chemother. 2010, 54, 2994–3001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brichacek, B.; Lagenaur, L.A.; Lee, P.P.; Venzon, D.; Hamer, D.H. In vivo evaluation of safety and toxicity of a Lactobacillus jensenii producing modified cyanovirin-N in a rhesus macaque vaginal challenge model. PLoS ONE 2013, 8, e78817. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrova, M.I.; Imholz, N.C.; Verhoeven, T.L.; Balzarini, J.; Van Damme, E.J.; Schols, D.; Vanderleyden, J.; Lebeer, S. Lectin-Like Molecules of Lactobacillus rhamnosus GG Inhibit Pathogenic Escherichia coli and Salmonella Biofilm Formation. PLoS ONE 2016, 11, e0161337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, D.H.; McBride, B.W.; Roff, M.A.; Farrar, G.H. Efficient purification and rigorous characterisation of a recombinant gp120 for HIV vaccine studies. Vaccine 1995, 13, 991–999. [Google Scholar] [CrossRef]
- Dean, S.N.; Leary, D.H.; Sullivan, C.J.; Oh, E.; Walper, S.A. Isolation and characterization of Lactobacillus-derived membrane vesicles. Sci. Rep. 2019, 9, 877. [Google Scholar] [CrossRef]
- Nahui Palomino, R.A.; Vanpouille, C.; Laghi, L.; Parolin, C.; Melikov, K.; Backlund, P.; Vitali, B.; Margolis, L. Extracellular vesicles from symbiotic vaginal lactobacilli inhibit HIV-1 infection of human tissues. Nat. Commun. 2019, 10, 5656. [Google Scholar] [CrossRef] [Green Version]
- Zuo, F.; Zeng, Z.; Hammarstrom, L.; Marcotte, H. Inducible Plasmid Self-Destruction (IPSD) Assisted Genome Engineering in Lactobacilli and Bifidobacteria. ACS Synth. Biol. 2019, 8, 1723–1729. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IOD50 | |||||||
|---|---|---|---|---|---|---|---|
| −MaMP | +MaMP | ||||||
| Pseudovirus | Tier | Clade | pIAV7 | pAF900-VHHA6 | pIAV7 | pAF900-VHHA6 | |
| ZM197 | 1B | C | 1.43 | 0.643 | 5.00 | 0.274 | IOD50 [OD600/well] |
| X2278-C2_B6 | 2 | B | 2.83 | 0.683 | 3.95 | 0.846 | |
| TRO.11 | 2 | B | 4.41 | 0.637 | 4.95 | 0.551 | |
| CE1178_A3 | 2 | C | 2.90 | 0.512 | 3.20 | 0.744 | <1 |
| HIV_25710_2.43 | 2 | C | 2.74 | 0.520 | 2.06 | 0.468 | 1–2.5 |
| optC | n.d. | C | 3.13 | 0.760 | 5.00 | 1.73 | 2.5–5 |
| CNE55 | 2 | CRF01_AE | - | - | 3.52 | 2.54 | |
| TRJO.4551 | 3 | B | 1.65 | 1.25 | 3.04 | 0.935 | |
| MLV | - | - | 4.87 | 4.99 | 4.38 | 4.48 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kalusche, S.; Vanshylla, K.; Kleipass, F.; Gruell, H.; Müller, B.; Zeng, Z.; Koch, K.; Stein, S.; Marcotte, H.; Klein, F.; et al. Lactobacilli Expressing Broadly Neutralizing Nanobodies against HIV-1 as Potential Vectors for HIV-1 Prophylaxis? Vaccines 2020, 8, 758. https://doi.org/10.3390/vaccines8040758
Kalusche S, Vanshylla K, Kleipass F, Gruell H, Müller B, Zeng Z, Koch K, Stein S, Marcotte H, Klein F, et al. Lactobacilli Expressing Broadly Neutralizing Nanobodies against HIV-1 as Potential Vectors for HIV-1 Prophylaxis? Vaccines. 2020; 8(4):758. https://doi.org/10.3390/vaccines8040758
Chicago/Turabian StyleKalusche, Sarah, Kanika Vanshylla, Franziska Kleipass, Henning Gruell, Barbara Müller, Zhu Zeng, Kathrin Koch, Stefan Stein, Harold Marcotte, Florian Klein, and et al. 2020. "Lactobacilli Expressing Broadly Neutralizing Nanobodies against HIV-1 as Potential Vectors for HIV-1 Prophylaxis?" Vaccines 8, no. 4: 758. https://doi.org/10.3390/vaccines8040758