Fusion Cytokines IL-7-Linker-IL-15 Promote Mycobacterium Tuberculosis Subunit Vaccine to Induce Central Memory like T Cell-Mediated Immunity


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Mice
2.3. Preparation of Subunit Vaccines and Single Mycobacterial Antigens
2.4. Construction of rAd-IL-7, rAd-IL-15, rAd-IL-7-IL-15, rAd-IL-7-Linker-IL-15 and rAd-Vector
2.5. Vaccine Immunization
2.6. Cultured IFN-γ ELISPOT assay In Vitro
2.7. EdU Incorporation and Proliferation Assay
2.8. IFN-γ Secretion Following Twice-Stimulation with Antigens
2.9. TB10.44-12 Pentamer Staining
2.10. BCG Challenge and Enumeration of Bacteria-Load
2.11. Statistical Analysis
3. Results
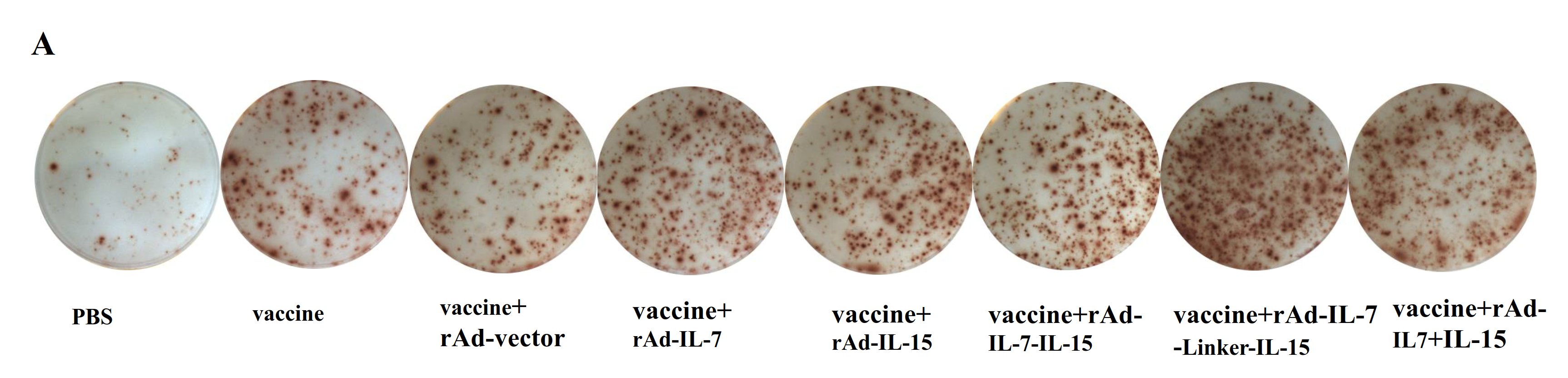
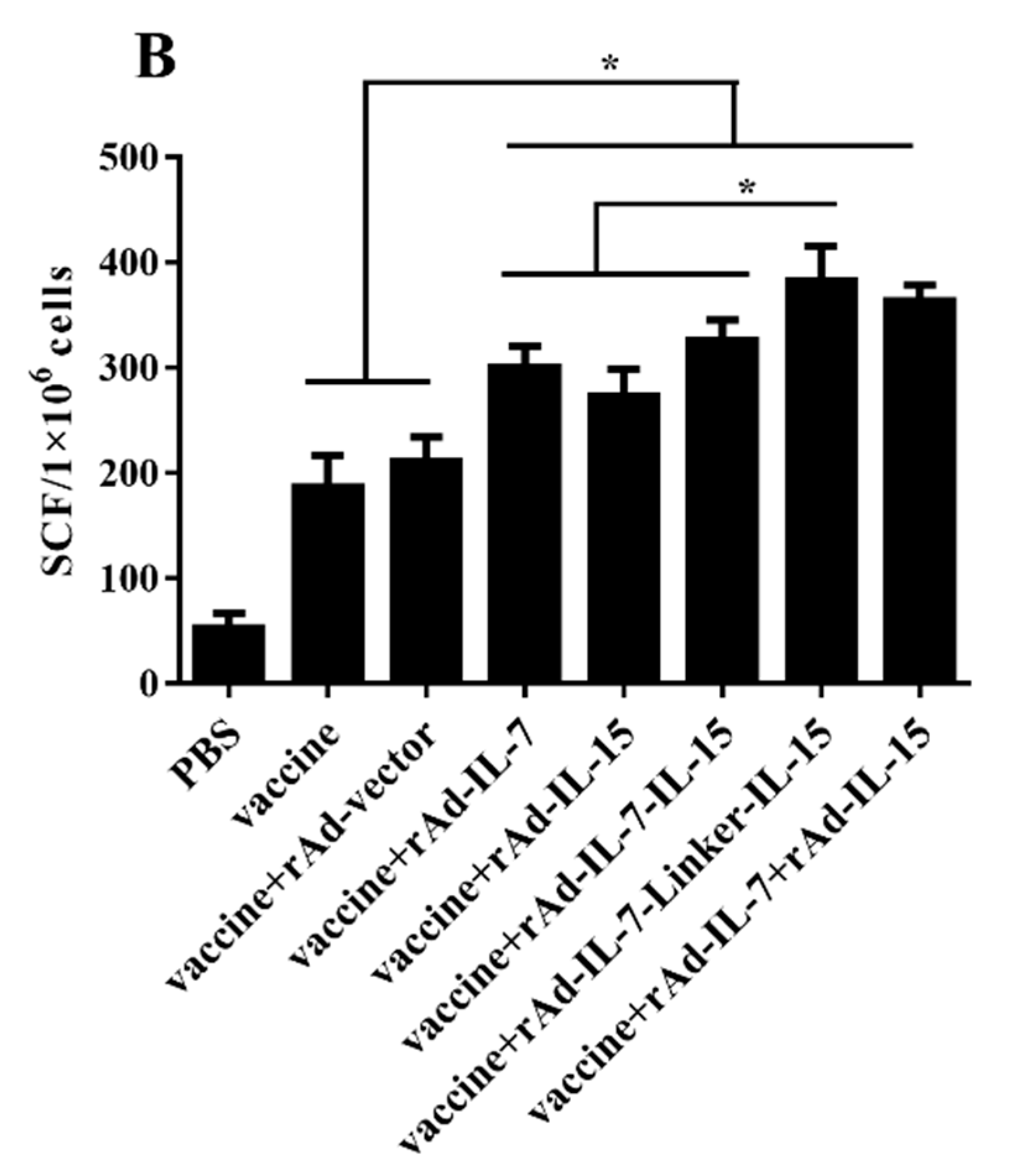
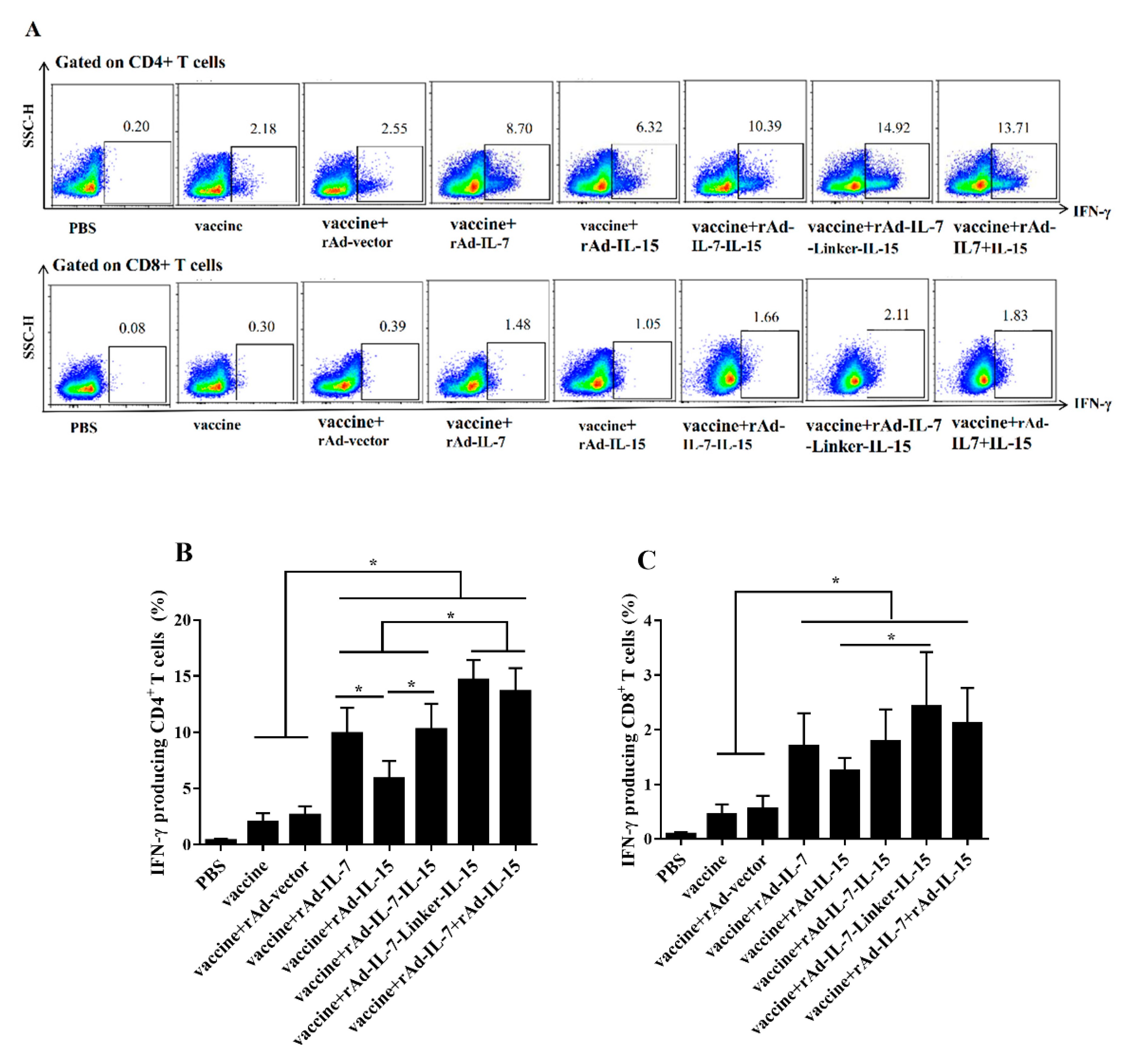
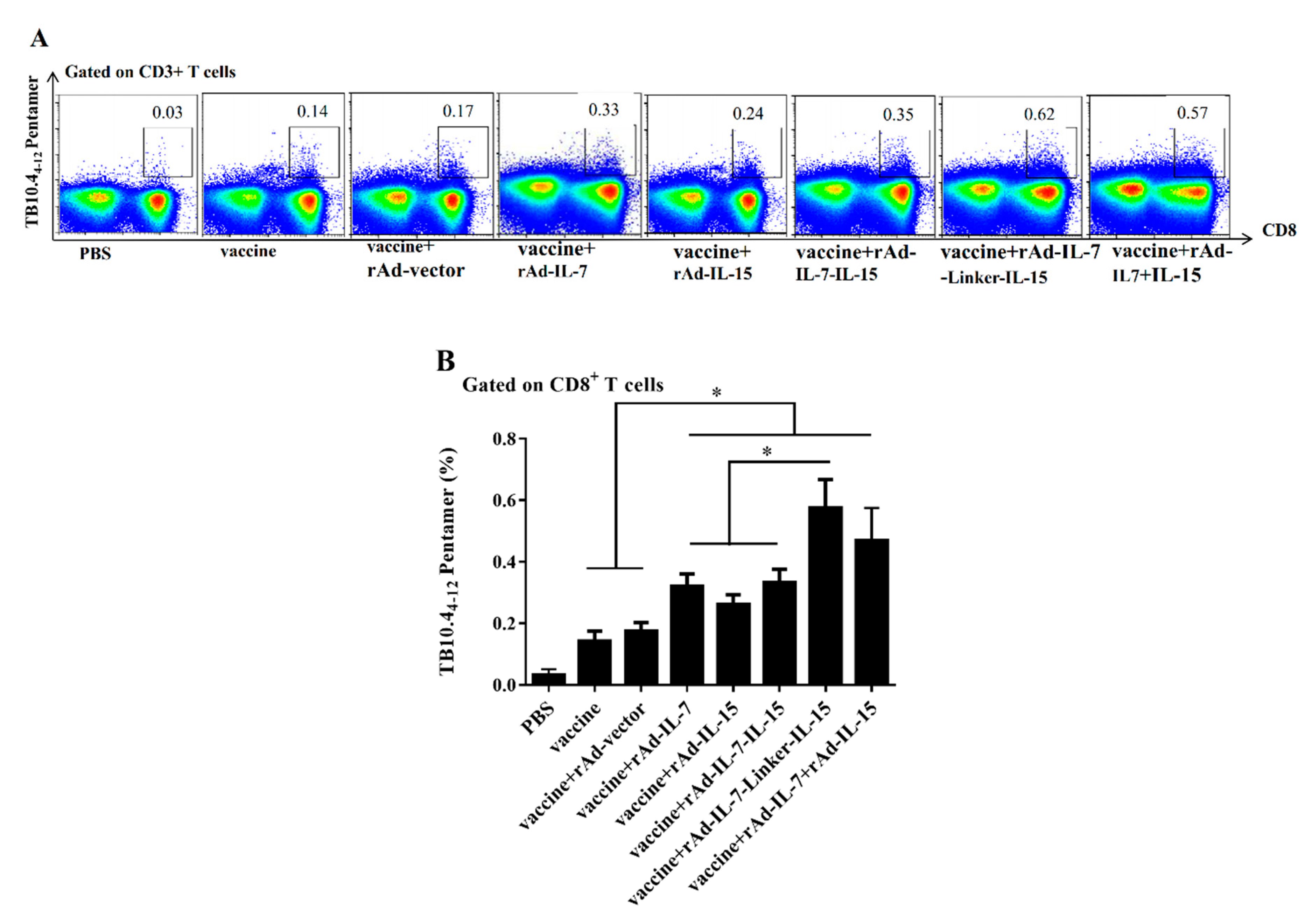
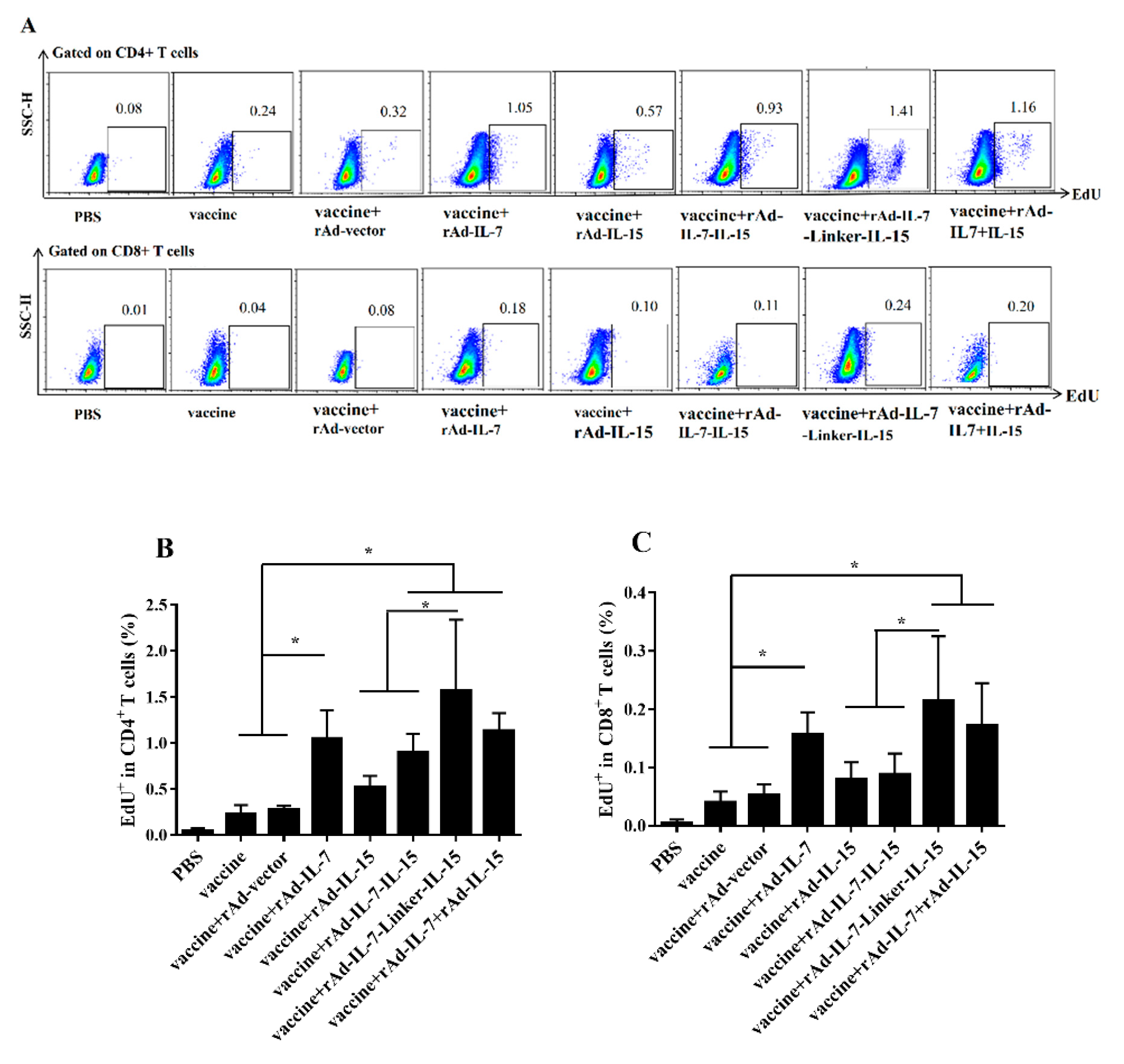
3.1. rAd-IL-7-Linker-IL-15 Enhanced Quality and Quantity of TCM Like Cells
3.2. rAd-IL-7-Linker-IL-15 Induced Higher Proliferation Capability of TCM Like Cells
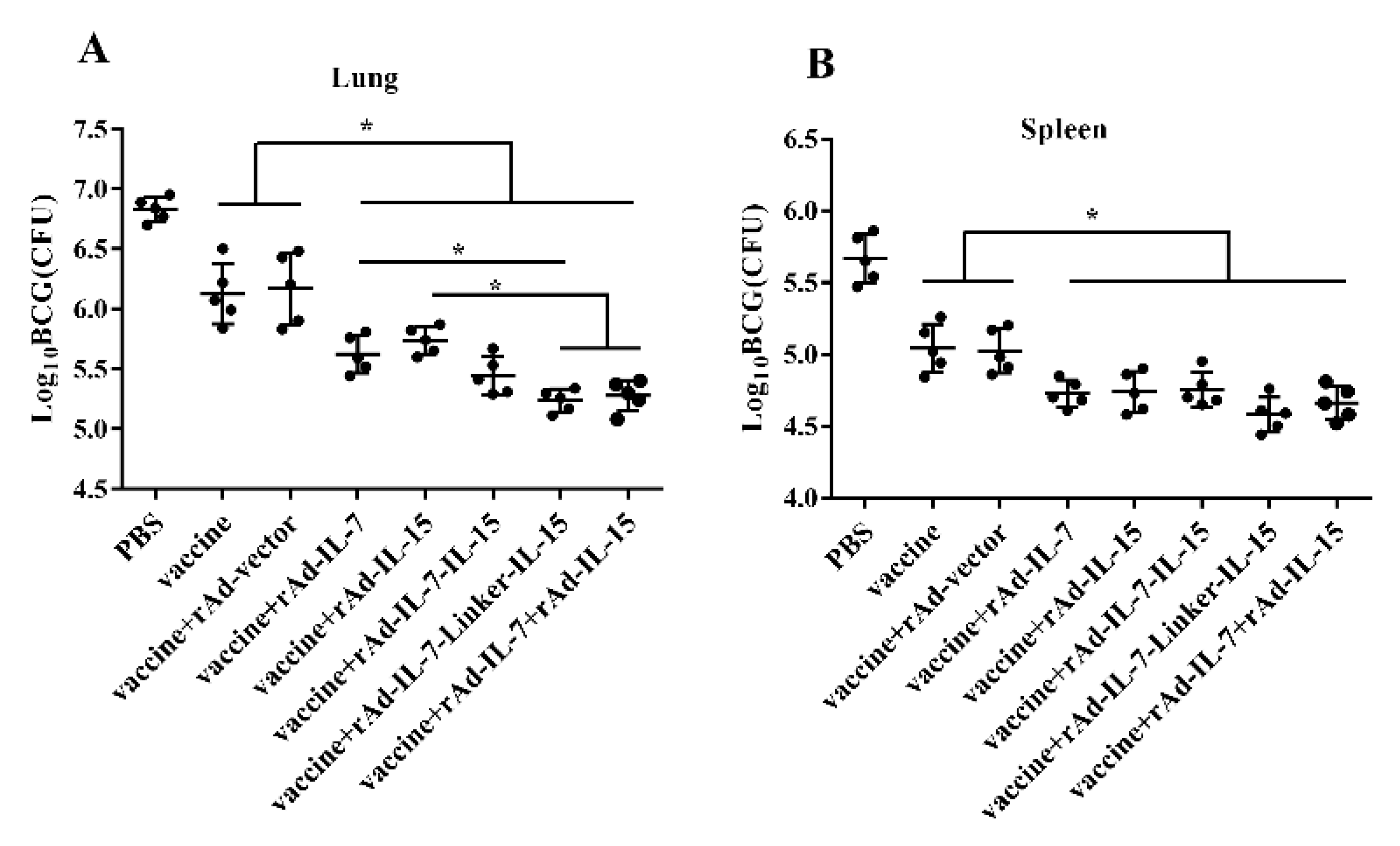
3.3. rAd-IL-7-Linker-IL-15 Promoted the Protective Efficacy of Vaccine
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TB—Tuberculosis |
| M. tuberculosis—Mycobacterium tuberculosis |
| BCG—Mycobacterium bovis Bacilli Calmette-Guerin |
| TCM like cells—central memory-like T cells |
| TCM—central memory T cells |
| TEM—effector memory T cells |
| DDA—dioctadecylammonium bromide |
| Poly (I:C)—polyinosinic-polycytidylic acid |
| IL—Interleukin |
| LT70-ESAT6-Ag85B-MPT64<190-198>-Mtb8.4-Rv2626c |
| MH—Mtb10.4-HspX |
| PBS—Phosphate-buffered saline |
| rAd—recombined adenoviral |
| CPE—cytopathic effect |
| i.p.—intraperitoneal injection |
| APCs—antigen presenting cells |
| Linker—(Gly-Gly-Gly-Ser)3 |
References
- WHO. Global Tuberculosis Report 2020. Available online: https://apps.who.int/iris/bitstream/handle/10665/336069/9789240013131-eng.pdf (accessed on 22 November 2020).
- Andersen, P.; Doherty, T.M. The success and failure of BCG—Implications for a novel tuberculosis vaccine. Nat. Rev. Microbiol. 2005, 3, 656–662. [Google Scholar] [CrossRef]
- Barker, L.F.; Leadman, A.E.; Clagett, B. The challenges of developing new tuberculosis vaccines. Health Aff. 2011, 30, 1073–1079. [Google Scholar] [CrossRef] [Green Version]
- Orme, I.M. The Achilles heel of BCG. Tuberculosis 2010, 90, 329–332. [Google Scholar] [CrossRef] [PubMed]
- Kaveh, D.A.; Garcia-Pelayo, M.C.; Hogarth, P.J. Persistent BCG bacilli perpetuate CD4 T effector memory and optimal protection against tuberculosis. Vaccine 2014, 32, 6911–6918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Youngblood, B.; Hale, J.S.; Ahmed, R. T-cell memory differentiation: Insights from transcriptional signatures and epigenetics. Immunology 2013, 139, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Voss, G.; Casimiro, D.; Neyrolles, O.; Williams, A.; Kaufmann, S.H.E.; McShane, H.; Hatherill, M.; Fletcher, H.A. Progress and challenges in TB vaccine development. F1000Research 2018, 7, 199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted influenza vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Da, Z.; Wang, Y.; Niu, H.; Li, R.; Yu, H.; He, S.; Guo, M.; Luo, Y.; Ma, X.; et al. A novel liposome adjuvant DPC mediates Mycobacterium tuberculosis subunit vaccine well to induce cell-mediated immunity and high protective efficacy in mice. Vaccine 2016, 34, 1370–1378. [Google Scholar] [CrossRef]
- Vogelzang, A.; Perdomo, C.; Zedler, U.; Kuhlmann, S.; Hurwitz, R.; Gengenbacher, M.; Kaufmann, S.H. Central memory CD4+ T cells are responsible for the recombinant Bacillus Calmette-Guerin DeltaureC::hly vaccine’s superior protection against tuberculosis. J. Infect. Dis. 2014, 210, 1928–1937. [Google Scholar] [CrossRef]
- Lindenstrom, T.; Knudsen, N.P.H.; Agger, E.M.; Andersen, P. Control of Chronic Mycobacterium tuberculosis Infection by CD4 KLRG1(-) IL-2-Secreting Central Memory Cells. J. Immunol. 2013, 190, 6311–6319. [Google Scholar] [CrossRef] [Green Version]
- Okoye, A.A.; Rohankhedkar, M.; Konfe, A.L.; Abana, C.O.; Reyes, M.D.; Clock, J.A.; Duell, D.M.; Sylwester, A.W.; Sammader, P.; Legasse, A.W.; et al. Effect of IL-7 Therapy on Naive and Memory T Cell Homeostasis in Aged Rhesus Macaques. J. Immunol. 2015, 195, 4292–4305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melchionda, F.; Fry, T.J.; Milliron, M.J.; McKirdy, M.A.; Tagaya, Y.; Mackall, C.L. Adjuvant IL-7 or IL-15 overcomes immunodominance and improves survival of the CD8+ memory cell pool. J. Clin. Investig. 2005, 115, 1177–1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Purton, J.F.; Martin, C.E.; Surh, C.D. Enhancing T cell memory: IL-7 as an adjuvant to boost memory T-cell generation. Immunol. Cell Biol. 2008, 86, 385–386. [Google Scholar] [CrossRef] [PubMed]
- Yeon, S.M.; Halim, L.; Chandele, A.; Perry, C.J.; Kim, S.H.; Kim, S.U.; Byun, Y.; Yuk, S.H.; Kaech, S.M.; Jung, Y.W. IL-7 plays a critical role for the homeostasis of allergen-specific memory CD4 T cells in the lung and airways. Sci. Rep. 2017, 7, 11155. [Google Scholar] [CrossRef] [Green Version]
- Knop, L.; Deiser, K.; Bank, U.; Witte, A.; Mohr, J.; Philipsen, L.; Fehling, H.J.; Muller, A.J.; Kalinke, U.; Schuler, T. IL-7 derived from lymph node fibroblastic reticular cells is dispensable for naive T cell homeostasis but crucial for central memory T cell survival. Eur. J. Immunol. 2020, 50, 846–857. [Google Scholar] [CrossRef] [Green Version]
- DeGottardi, M.Q.; Okoye, A.A.; Vaidya, M.; Talla, A.; Konfe, A.L.; Reyes, M.D.; Clock, J.A.; Duell, D.M.; Legasse, A.W.; Sabnis, A.; et al. Effect of Anti-IL-15 Administration on T Cell and NK Cell Homeostasis in Rhesus Macaques. J. Immunol. 2016, 197, 1183–1198. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Li, Z.Y.; Petersen, E.; Liu, W.G.; Zhu, X.Q. Co-administration of interleukins 7 and 15 with DNA vaccine improves protective immunity against Toxoplasma gondii. Exp. Parasitol. 2016, 162, 18–23. [Google Scholar] [CrossRef]
- Gao, Q.; Zhang, N.Z.; Zhang, F.K.; Wang, M.; Hu, L.Y.; Zhu, X.Q. Immune response and protective effect against chronic Toxoplasma gondii infection induced by vaccination with a DNA vaccine encoding profilin. BMC Infect. Dis. 2018, 18, 117. [Google Scholar] [CrossRef] [Green Version]
- Calarota, S.A.; Dai, A.; Trocio, J.N.; Weiner, D.B.; Lori, F.; Lisziewicz, J. IL-15 as memory T-cell adjuvant for topical HIV-1 DermaVir vaccine. Vaccine 2008, 26, 5188–5195. [Google Scholar] [CrossRef] [Green Version]
- Singh, V.; Gowthaman, U.; Jain, S.; Parihar, P.; Banskar, S.; Gupta, P.; Gupta, U.D.; Agrewala, J.N. Coadministration of interleukins 7 and 15 with bacille Calmette-Guerin mounts enduring T cell memory response against Mycobacterium tuberculosis. J. Infect. Dis. 2010, 202, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Tang, C.; Yamada, H.; Shibata, K.; Yoshida, S.; Wajjwalku, W.; Yoshikai, Y. IL-15 protects antigen-specific CD8+ T cell contraction after Mycobacterium bovis bacillus Calmette-Guerin infection. J. Leukoc. Biol. 2009, 86, 187–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Peng, J.; Hu, L.; Luo, Y.; Niu, H.; Bai, C.; Wang, Q.; Li, F.; Yu, H.; Wang, B.; et al. A multistage mycobacterium tuberculosis subunit vaccine LT70 including latency antigen Rv2626c induces long-term protection against tuberculosis. Hum. Vaccines Immunother. 2016, 12, 1670–1677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niu, H.; Hu, L.; Li, Q.; Da, Z.; Wang, B.; Tang, K.; Xin, Q.; Yu, H.; Zhang, Y.; Wang, Y.; et al. Construction and evaluation of a multistage Mycobacterium tuberculosis subunit vaccine candidate Mtb10.4-HspX. Vaccine 2011, 29, 9451–9458. [Google Scholar] [CrossRef] [PubMed]
- Xin, Q.; Niu, H.; Li, Z.; Zhang, G.; Hu, L.; Wang, B.; Li, J.; Yu, H.; Liu, W.; Wang, Y.; et al. Subunit vaccine consisting of multi-stage antigens has high protective efficacy against Mycobacterium tuberculosis infection in mice. PLoS ONE 2013, 8, e72745. [Google Scholar] [CrossRef]
- Huston, J.S.; Levinson, D.; Mudgett-Hunter, M.; Tai, M.S.; Novotny, J.; Margolies, M.N.; Ridge, R.J.; Bruccoleri, R.E.; Haber, E.; Crea, R.; et al. Protein engineering of antibody binding sites: Recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in Escherichia coli. Proc. Natl. Acad. Sci. USA 1988, 85, 5879–5883. [Google Scholar] [CrossRef] [Green Version]
- Darling, A.J.; Boose, J.A.; Spaltro, J. Virus assay methods: Accuracy and validation. Biol. J. Int. Assoc. Biol. Stand. 1998, 26, 105–110. [Google Scholar] [CrossRef]
- Bai, C.; He, J.; Niu, H.; Hu, L.; Luo, Y.; Liu, X.; Peng, L.; Zhu, B. Prolonged intervals during Mycobacterium tuberculosis subunit vaccine boosting contributes to eliciting immunity mediated by central memory-like T cells. Tuberculosis 2018, 110, 104–111. [Google Scholar] [CrossRef]
- Billeskov, R.; Vingsbo-Lundberg, C.; Andersen, P.; Dietrich, J. Induction of CD8 T cells against a novel epitope in TB10.4: Correlation with mycobacterial virulence and the presence of a functional region of difference-1. J. Immunol. 2007, 179, 3973–3981. [Google Scholar] [CrossRef]
- Calarota, S.A.; Baldanti, F. Enumeration and characterization of human memory T cells by enzyme-linked immunospot assays. Clin. Dev. Immunol. 2013, 2013, 637649. [Google Scholar] [CrossRef] [Green Version]
- Aagaard, C.; Hoang, T.; Dietrich, J.; Cardona, P.J.; Izzo, A.; Dolganov, G.; Schoolnik, G.K.; Cassidy, J.P.; Billeskov, R.; Andersen, P. A multistage tuberculosis vaccine that confers efficient protection before and after exposure. Nat. Med. 2011, 17, 189–194. [Google Scholar] [CrossRef]
- Niu, H.; Peng, J.; Bai, C.; Liu, X.; Hu, L.; Luo, Y.; Wang, B.; Zhang, Y.; Chen, J.; Yu, H.; et al. Multi-Stage Tuberculosis Subunit Vaccine Candidate LT69 Provides High Protection against Mycobacterium tuberculosis Infection in Mice. PLoS ONE 2015, 10, e0130641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rouanet, C.; Debrie, A.S.; Lecher, S.; Locht, C. Subcutaneous boosting with heparin binding haemagglutinin increases BCG-induced protection against tuberculosis. Microbes Infect. 2009, 11, 995–1001. [Google Scholar] [CrossRef] [PubMed]
- Bohme, J.; Martinez, N.; Li, S.; Lee, A.; Marzuki, M.; Tizazu, A.M.; Ackart, D.; Frenkel, J.H.; Todd, A.; Lachmandas, E.; et al. Metformin enhances anti-mycobacterial responses by educating CD8+ T-cell immunometabolic circuits. Nat. Commun. 2020, 11, 5225. [Google Scholar] [CrossRef] [PubMed]
- Lenz, D.C.; Kurz, S.K.; Lemmens, E.; Schoenberger, S.P.; Sprent, J.; Oldstone, M.B.; Homann, D. IL-7 regulates basal homeostatic proliferation of antiviral CD4+ T cell memory. Proc. Natl. Acad. Sci. USA 2004, 101, 9357–9362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lugli, E.; Goldman, C.K.; Perera, L.P.; Smedley, J.; Pung, R.; Yovandich, J.L.; Creekmore, S.P.; Waldmann, T.A.; Roederer, M. Transient and persistent effects of IL-15 on lymphocyte homeostasis in nonhuman primates. Blood 2010, 116, 3238–3248. [Google Scholar] [CrossRef]
- Castro, I.; Yu, A.; Dee, M.J.; Malek, T.R. The basis of distinctive IL-2- and IL-15-dependent signaling: Weak CD122-dependent signaling favors CD8+ T central-memory cell survival but not T effector-memory cell development. J. Immunol. 2011, 187, 5170–5182. [Google Scholar] [CrossRef] [Green Version]
- Ku, C.C.; Murakami, M.; Sakamoto, A.; Kappler, J.; Marrack, P. Control of homeostasis of CD8+ memory T cells by opposing cytokines. Science 2000, 288, 675–678. [Google Scholar] [CrossRef]
- Surh, C.D.; Sprent, J. Homeostasis of naive and memory T cells. Immunity 2008, 29, 848–862. [Google Scholar] [CrossRef] [Green Version]
- Boyman, O.; Letourneau, S.; Krieg, C.; Sprent, J. Homeostatic proliferation and survival of naive and memory T cells. Eur. J. Immunol. 2009, 39, 2088–2094. [Google Scholar] [CrossRef]
- Reddy, S.T.; Ge, X.; Miklos, A.E.; Hughes, R.A.; Kang, S.H.; Hoi, K.H.; Chrysostomou, C.; Hunicke-Smith, S.P.; Iverson, B.L.; Tucker, P.W.; et al. Monoclonal antibodies isolated without screening by analyzing the variable-gene repertoire of plasma cells. Nat. Biotechnol. 2010, 28, 965–969. [Google Scholar] [CrossRef]
- Alfthan, K.; Takkinen, K.; Sizmann, D.; Soderlund, H.; Teeri, T.T. Properties of a single-chain antibody containing different linker peptides. Protein Eng. 1995, 8, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Brown, V.I.; Hulitt, J.; Fish, J.; Sheen, C.; Bruno, M.; Xu, Q.; Carroll, M.; Fang, J.; Teachey, D.; Grupp, S.A. Thymic stromal-derived lymphopoietin induces proliferation of pre-B leukemia and antagonizes mTOR inhibitors, suggesting a role for interleukin-7Ralpha signaling. Cancer Res. 2007, 67, 9963–9970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, J.; Pfannenstiel, V.; Waldmann, A.; Bergs, J.W.J.; Brill, B.; Huenecke, S.; Klingebiel, T.; Rodel, F.; Buchholz, C.J.; Wels, W.S.; et al. A Two-Phase Expansion Protocol Combining Interleukin (IL)-15 and IL-21 Improves Natural Killer Cell Proliferation and Cytotoxicity against Rhabdomyosarcoma. Front. Immunol. 2017, 8, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, K.C.; Lugli, E.; Welles, H.C.; Rosenberg, S.A.; Fojo, A.T.; Morris, J.C.; Fleisher, T.A.; Dubois, S.P.; Perera, L.P.; Stewart, D.M.; et al. Redistribution, hyperproliferation, activation of natural killer cells and CD8 T cells, and cytokine production during first-in-human clinical trial of recombinant human interleukin-15 in patients with cancer. J. Clin. Oncol. 2015, 33, 74–82. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bai, C.; Zhou, L.; Tang, J.; He, J.; Han, J.; Niu, H.; Zhu, B. Fusion Cytokines IL-7-Linker-IL-15 Promote Mycobacterium Tuberculosis Subunit Vaccine to Induce Central Memory like T Cell-Mediated Immunity. Vaccines 2020, 8, 715. https://doi.org/10.3390/vaccines8040715
Bai C, Zhou L, Tang J, He J, Han J, Niu H, Zhu B. Fusion Cytokines IL-7-Linker-IL-15 Promote Mycobacterium Tuberculosis Subunit Vaccine to Induce Central Memory like T Cell-Mediated Immunity. Vaccines. 2020; 8(4):715. https://doi.org/10.3390/vaccines8040715
Chicago/Turabian StyleBai, Chunxiang, Lijun Zhou, Junxia Tang, Juanjuan He, Jiangyuan Han, Hongxia Niu, and Bingdong Zhu. 2020. "Fusion Cytokines IL-7-Linker-IL-15 Promote Mycobacterium Tuberculosis Subunit Vaccine to Induce Central Memory like T Cell-Mediated Immunity" Vaccines 8, no. 4: 715. https://doi.org/10.3390/vaccines8040715