Epitope Selection for Fighting Visceral Leishmaniosis: Not All Peptides Function the Same Way

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals, Parasites, and Production of Leishmania Soluble Antigen
2.2. Prediction of MHC Class I and II Binding Epitopes
2.3. Multiepitope Peptide Design and Synthesis
2.4. Immunization Protocol
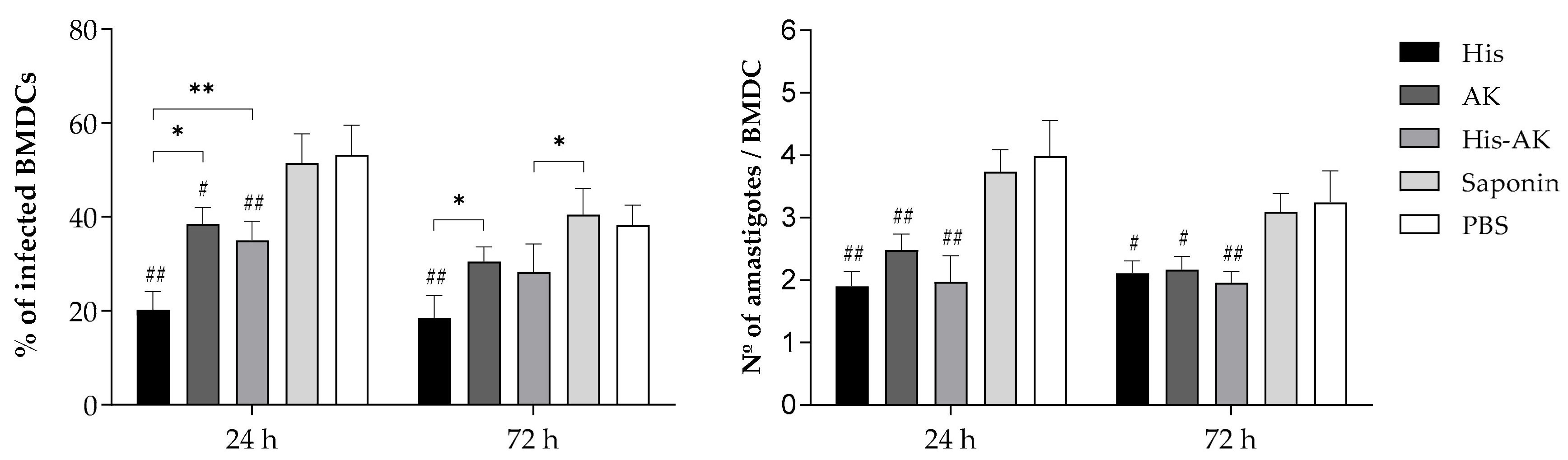
2.5. Maturation of Bone-Marrow-Derived Dendritic Cells (BMDCs) for Post- and Preinfection Assays
2.6. Immune Response Induced by Multiepitope Peptide Immunizations Prior to Infection
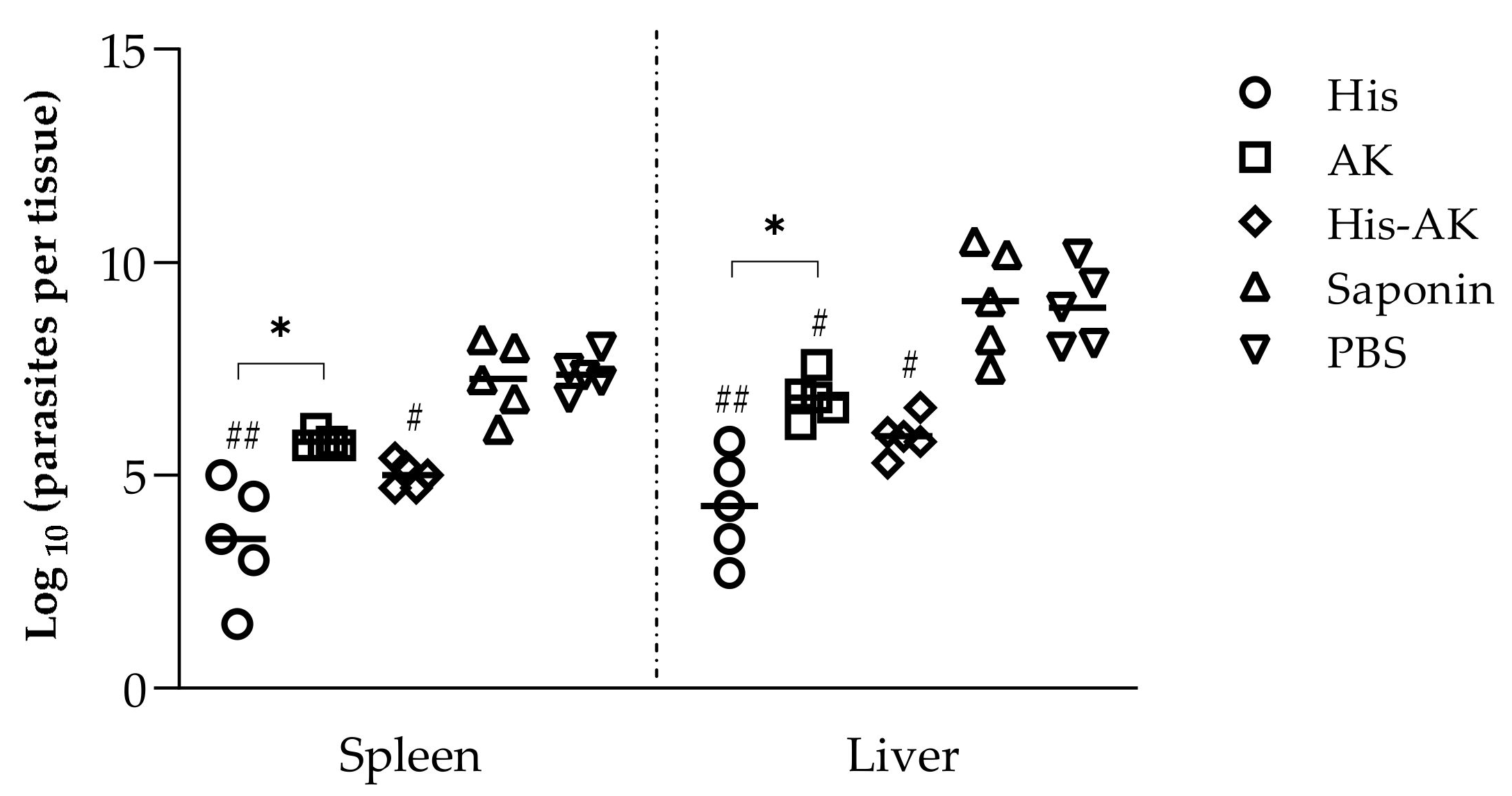
2.7. Infection and Parasite Burden
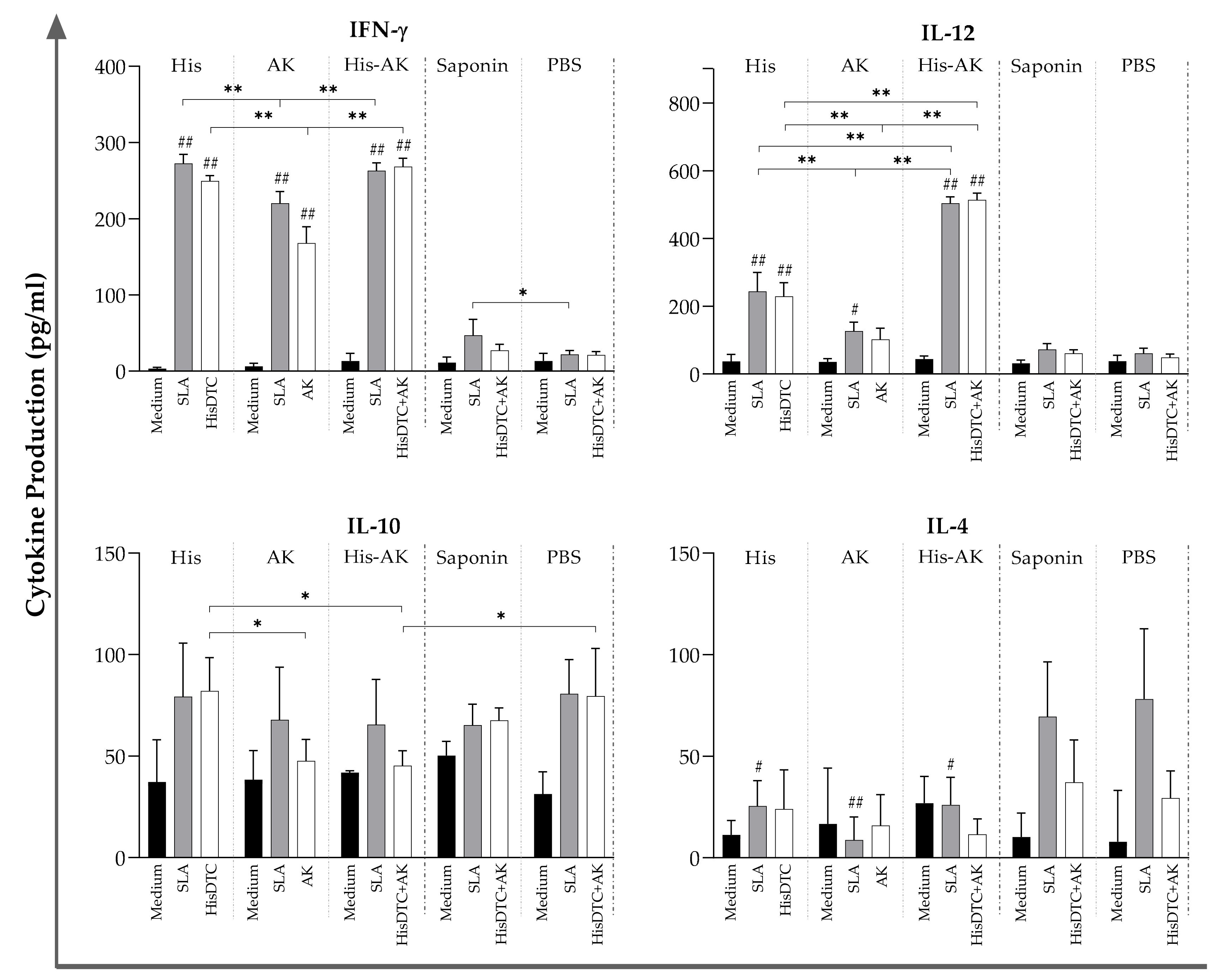
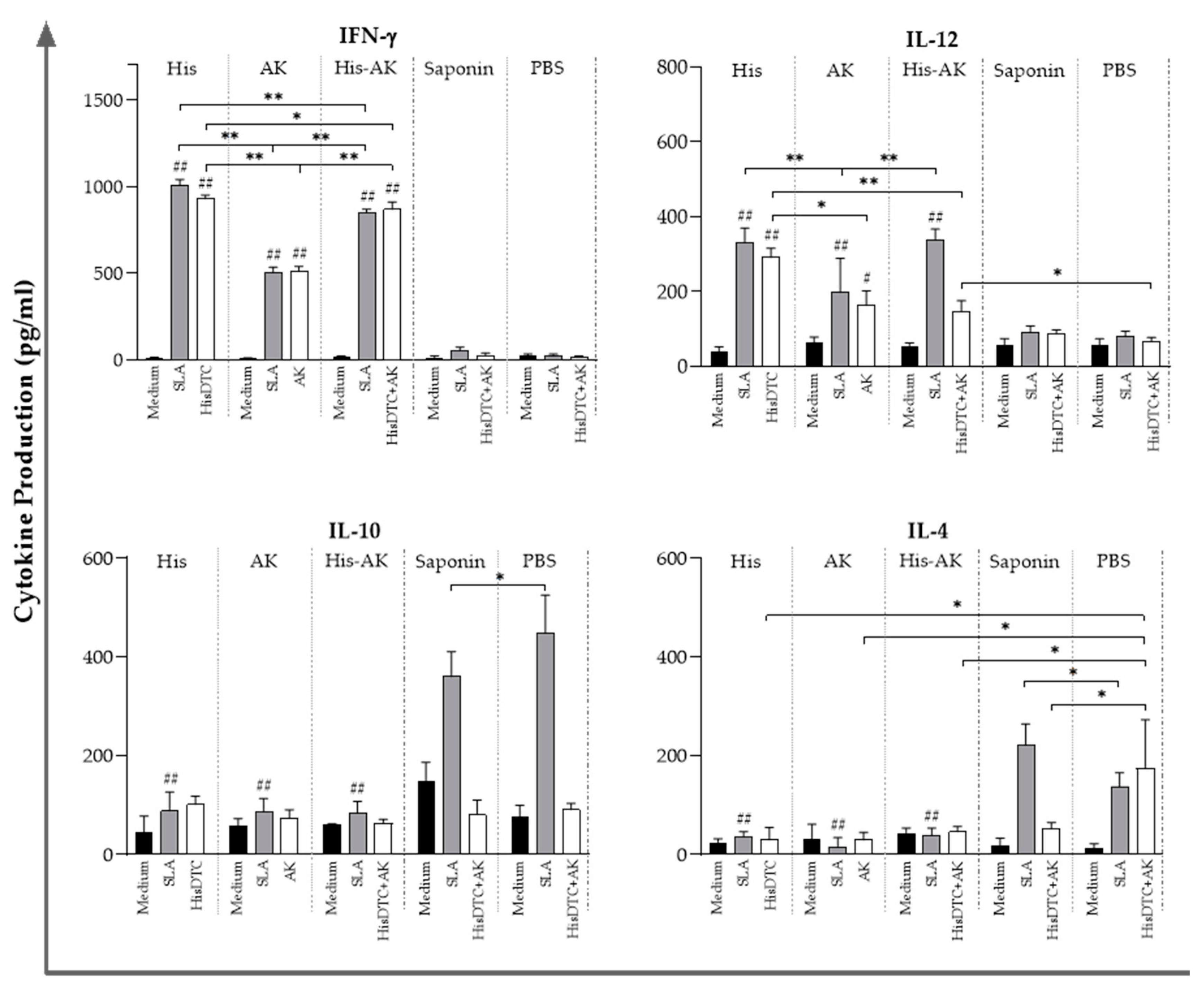
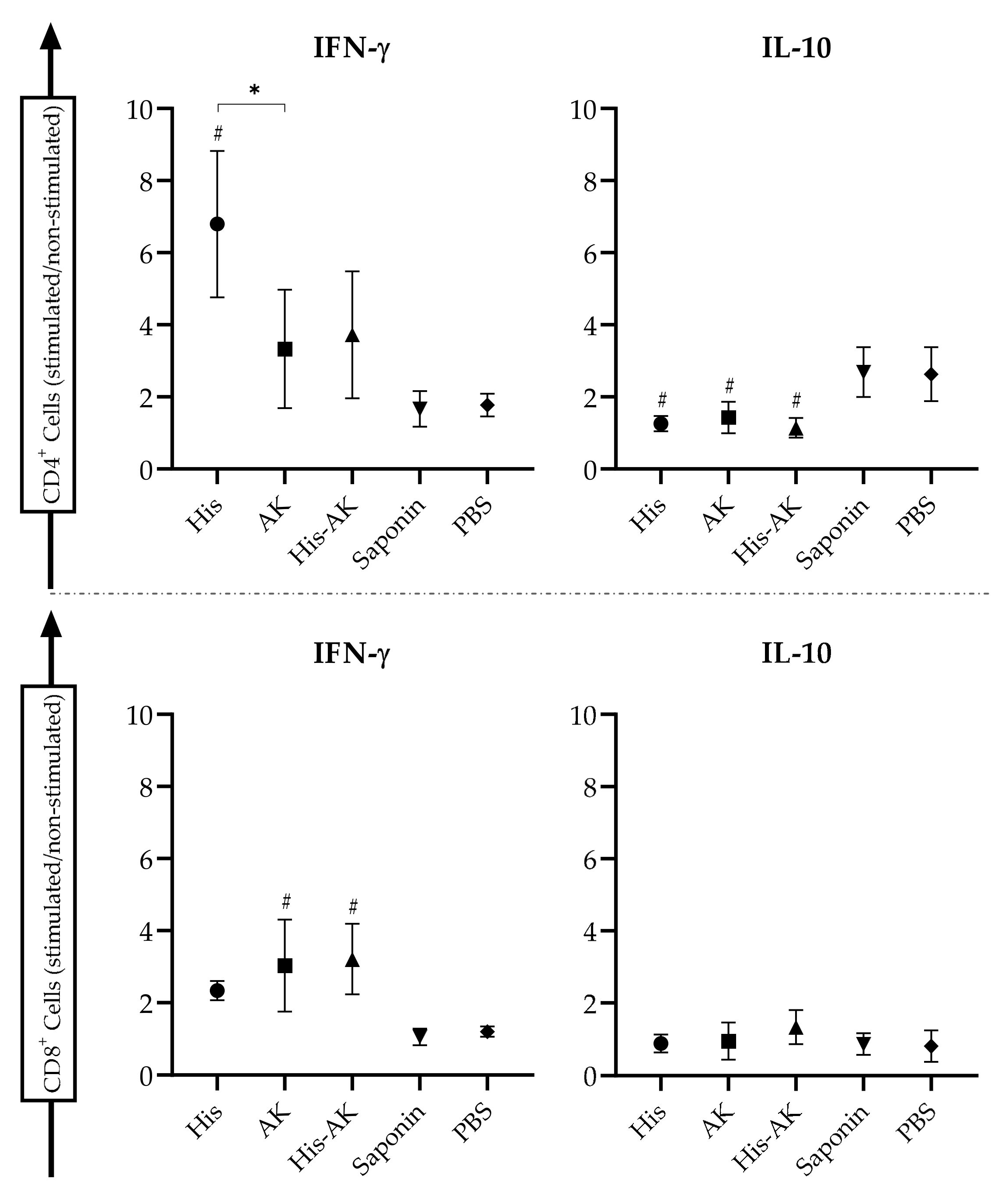
2.8. Cellular Immune Response in the Spleen: Cytokine Production
2.9. Enzyme Modulation after Infection
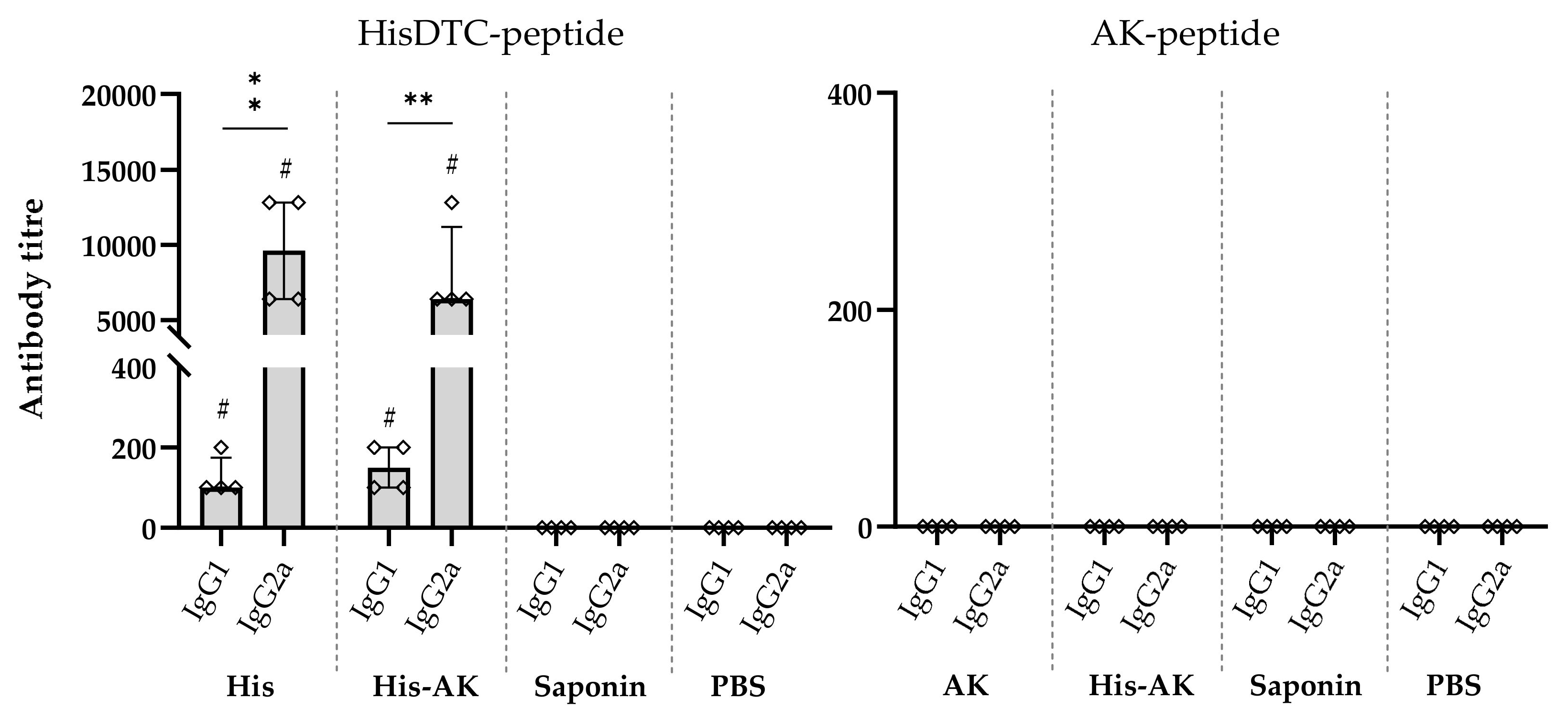
2.10. Multiepitope Peptide Immunization, and Postinfection Humoral Response Assessment
2.11. Statistical Analysis
3. Results
3.1. Used of Artificial Neural Network Analysis for Predicting Strongly Binding Epitopes for Human and Mouse MHC Class I and II Molecules
3.2. In Silico Analysis of Chimeric Multiepitope Peptides
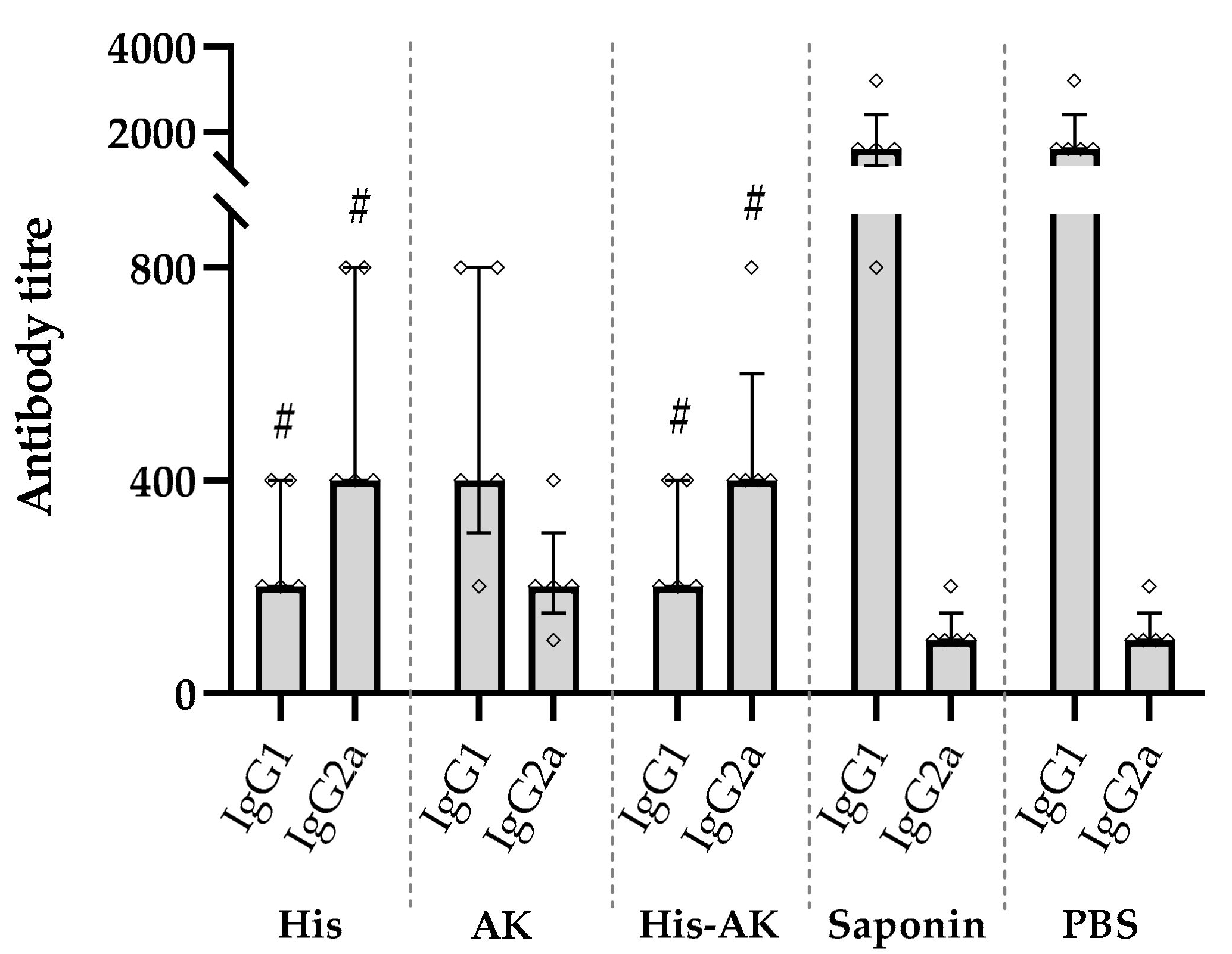
3.3. Evaluation of the Efficiency of Multiepitope Peptide Immunogenicity in Mice
3.4. Immunization with Multiepitope Peptides Induced a Diminished Parasite Load in Target Organs
3.5. Effects of Immunization on the Induction of the Immune Response After L. infantum Challenge
3.6. Multiepitope Immunization Mitigated the Impact of L. Infantum Parasites on Host L-arginine Metabolism
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Conflicts of Interest
References
- WHO Organization. Global Health Observatory (GHO) data. In Leishmaniasis: Situation and Trends; WHO: Geneva, Switzerland, 2017. [Google Scholar]
- WHO. Leishmaniasis. Available online: http://www.who.int/leishmaniasis/en/ (accessed on 15 December 2019).
- Martin-Martin, I.; Jimenez, M.; Gonzalez, E.; Eguiluz, C.; Molina, R. Natural transmission of Leishmania infantum through experimentally infected Phlebotomus perniciosus highlights the virulence of Leishmania parasites circulating in the human visceral leishmaniasis outbreak in Madrid, Spain. Vet. Res. 2015, 46, 138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aguado, M.; Espinosa, P.; Romero-Mate, A.; Tardio, J.C.; Cordoba, S.; Borbujo, J. Outbreak of Cutaneous Leishmaniasis in Fuenlabrada, Madrid. Actas Dermosifiliogr. 2013, 104, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Bernal, G.; Jimenez, M.; Molina, R.; Ordonez-Gutierrez, L.; Martinez-Rodrigo, A.; Mas, A.; Cutuli, M.T.; Carrion, J. Characterisation of the ex vivo virulence of Leishmania infantum isolates from Phlebotomus perniciosus from an outbreak of human leishmaniosis in Madrid, Spain. Parasites Vectors 2014, 7, 499. [Google Scholar] [CrossRef] [PubMed]
- Roth-Damas, P.; Sempere-Manuel, M.; Mialaret-Lahiguera, A.; Fernandez-Garcia, C.; Gil-Tomas, J.J.; Colomina-Rodriguez, J.; Palop-Larrea, V. Community outbreak of cutaneous leishmaniasis in La Ribera region of Valencia, Spain: Public Health measures. Enferm. Infecc. Microbiol. Clin. 2016. [Google Scholar] [CrossRef]
- Jimenez, M.; Gonzalez, E.; Martin-Martin, I.; Hernandez, S.; Molina, R. Could wild rabbits (Oryctolagus cuniculus) be reservoirs for Leishmania infantum in the focus of Madrid, Spain? Vet. Parasitol. 2014, 202, 296–300. [Google Scholar] [CrossRef]
- Molina, R.; Jimenez, M.I.; Cruz, I.; Iriso, A.; Martin-Martin, I.; Sevillano, O.; Melero, S.; Bernal, J. The hare (Lepus granatensis) as potential sylvatic reservoir of Leishmania infantum in Spain. Vet. Parasitol. 2012, 190, 268–271. [Google Scholar] [CrossRef]
- Miro, G.; Ruperez, C.; Checa, R.; Galvez, R.; Hernandez, L.; Garcia, M.; Canorea, I.; Marino, V.; Montoya, A. Current status of L. infantum infection in stray cats in the Madrid region (Spain): Implications for the recent outbreak of human leishmaniosis? Parasites Vectors 2014, 7, 112. [Google Scholar] [CrossRef] [Green Version]
- Akhoundi, M.; Kuhls, K.; Cannet, A.; Votypka, J.; Marty, P.; Delaunay, P.; Sereno, D. A Historical Overview of the Classification, Evolution, and Dispersion of Leishmania Parasites and Sandflies. PLoS Negl. Trop. Dis. 2016, 10, e0004349. [Google Scholar] [CrossRef]
- Fenwick, A. The global burden of neglected tropical diseases. Public Health 2012, 126, 233–236. [Google Scholar] [CrossRef]
- Bern, C.; Maguire, J.H.; Alvar, J. Complexities of assessing the disease burden attributable to leishmaniasis. PLoS Negl. Trop. Dis. 2008, 2, e313. [Google Scholar] [CrossRef]
- Maroli, M.; Feliciangeli, M.D.; Bichaud, L.; Charrel, R.N.; Gradoni, L. Phlebotomine sandflies and the spreading of leishmaniases and other diseases of public health concern. Med. Vet. Entomol. 2013, 27, 123–147. [Google Scholar] [CrossRef] [PubMed]
- Antoniou, M.; Gramiccia, M.; Molina, R.; Dvorak, V.; Volf, P. The role of indigenous phlebotomine sandflies and mammals in the spreading of leishmaniasis agents in the Mediterranean region. Euro Surveill. Bull. Eur. Mal. Transm. 2013, 18, 20540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iborra, S.; Solana, J.C.; Requena, J.M.; Soto, M. Vaccine candidates against leishmania under current research. Expert Rev. Vaccines 2018, 17, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Glennie, N.D.; Scott, P. Memory T cells in cutaneous leishmaniasis. Cell Immunol. 2016, 309, 50–54. [Google Scholar] [CrossRef] [Green Version]
- Scott, P.; Novais, F.O. Cutaneous leishmaniasis: Immune responses in protection and pathogenesis. Nat. Rev. Immunol. 2016, 16, 581–592. [Google Scholar] [CrossRef]
- Velez, R.; Gallego, M. Commercially approved vaccines for canine leishmaniosis: A review of available data on their safety and efficacy. Trop. Med. Int. Health 2020. [Google Scholar] [CrossRef] [Green Version]
- De Brito, R.C.F.; Cardoso, J.M.D.O.; Reis, L.E.S.; Mathias, F.A.S.; Aguiar-Soares, R.D.D.O.; Teixeira-Carvalho, A.; Roatt, B.M.; Corrêa-Oliveira, R.; Ruiz, J.C.; Resende, D.D.M.; et al. Synthetic Peptides Elicit Strong Cellular Immunity in Visceral Leishmaniasis Natural Reservoir and Contribute to Long-Lasting Polyfunctional T-Cells in BALB/c Mice. Vaccines 2019, 7, 162. [Google Scholar] [CrossRef] [Green Version]
- Carcelen, J.; Iniesta, V.; Fernandez-Cotrina, J.; Serrano, F.; Parejo, J.C.; Corraliza, I.; Gallardo-Soler, A.; Maranon, F.; Soto, M.; Alonso, C.; et al. The chimerical multi-component Q protein from Leishmania in the absence of adjuvant protects dogs against an experimental Leishmania infantum infection. Vaccine 2009, 27, 5964–5973. [Google Scholar] [CrossRef]
- Ratnapriya, S.; Keerti; Sahasrabuddhe, A.A.; Dube, A. Visceral leishmaniasis: An overview of vaccine adjuvants and their applications. Vaccine 2019, 37, 3505–3519. [Google Scholar] [CrossRef]
- Ribeiro, P.A.F.; Dias, D.S.; Novais, M.V.M.; Lage, D.P.; Tavares, G.S.V.; Mendonca, D.V.C.; Oliveira, J.S.; Chavez-Fumagalli, M.A.; Roatt, B.M.; Duarte, M.C.; et al. A Leishmania hypothetical protein-containing liposome-based formulation is highly immunogenic and induces protection against visceral leishmaniasis. Cytokine 2018, 111, 131–139. [Google Scholar] [CrossRef]
- Athanasiou, E.; Agallou, M.; Tastsoglou, S.; Kammona, O.; Hatzigeorgiou, A.; Kiparissides, C.; Karagouni, E. A Poly(Lactic-co-Glycolic) Acid Nanovaccine Based on Chimeric Peptides from Different Leishmania infantum Proteins Induces Dendritic Cells Maturation and Promotes Peptide-Specific IFNgamma-Producing CD8(+) T Cells Essential for the Protection against Experimental Visceral Leishmaniasis. Front. Immunol. 2017, 8, 684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Agallou, M.; Athanasiou, E.; Koutsoni, O.; Dotsika, E.; Karagouni, E. Experimental Validation of Multi-Epitope Peptides Including Promising MHC Class I- and II-Restricted Epitopes of Four Known Leishmania infantum Proteins. Front. Immunol. 2014, 5, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alves-Silva, M.V.; Nico, D.; Morrot, A.; Palatnik, M.; Palatnik-de-Sousa, C.B. A Chimera Containing CD4+ and CD8+ T-Cell Epitopes of the Leishmania donovani Nucleoside Hydrolase (NH36) Optimizes Cross-Protection against Leishmania amazonesis Infection. Front. Immunol. 2017, 8, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cunha-Neto, E. MHC-restricted antigen presentation and recognition: Constraints on gene, recombinant and peptide vaccines in humans. Braz. J. Med. Biol. Res. 1999, 32, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Soto, M.; Requena, J.M.; Gomez, L.C.; Navarrete, I.; Alonso, C. Molecular characterization of a Leishmania donovani infantum antigen identified as histone H2A. Eur. J. Biochem. 1992, 205, 211–216. [Google Scholar] [CrossRef]
- Soto, M.; Requena, J.M.; Morales, G.; Alonso, C. The Leishmania infantum histone H3 possesses an extremely divergent N-terminal domain. Biochim. Biophys. Acta 1994, 1219, 533–535. [Google Scholar] [CrossRef]
- Zhang, W.W.; Matlashewski, G. Characterization of the A2-A2rel gene cluster in Leishmania donovani: Involvement of A2 in visceralization during infection. Mol. Microbiol. 2001, 39, 935–948. [Google Scholar] [CrossRef]
- Agallou, M.; Margaroni, M.; Karagouni, E. Cellular vaccination with bone marrow-derived dendritic cells pulsed with a peptide of Leishmania infantum KMP-11 and CpG oligonucleotides induces protection in a murine model of visceral leishmaniasis. Vaccine 2011, 29, 5053–5064. [Google Scholar] [CrossRef]
- Sachdeva, R.; Banerjea, A.C.; Malla, N.; Dubey, M.L. Immunogenicity and efficacy of single antigen Gp63, polytope and polytopeHSP70 DNA vaccines against visceral Leishmaniasis in experimental mouse model. PLoS ONE 2009, 4, e7880. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bernal, G.; Horcajo, P.; Orden, J.A.; De La Fuente, R.; Herrero-Gil, A.; Ordonez-Gutierrez, L.; Carrion, J. Mitigating an undesirable immune response of inherent susceptibility to cutaneous leishmaniosis in a mouse model: The role of the pathoantigenic HISA70 DNA vaccine. Vet. Res. 2012, 43, 59. [Google Scholar] [CrossRef] [Green Version]
- Dominguez-Bernal, G.; Horcajo, P.; Orden, J.A.; Ruiz-Santa-Quiteria, J.A.; De La Fuente, R.; Ordonez-Gutierrez, L.; Martinez-Rodrigo, A.; Mas, A.; Carrion, J. HisAK70: Progress towards a vaccine against different forms of leishmaniosis. Parasites Vectors 2015, 8, 629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez-Rodrigo, A.; Mas, A.; Fernández-Cotrina, J.; Belinchón-Lorenzo, S.; Orden, J.A.; Arias, P.; de la Fuente, R.; Carrión, J.; Domínguez-Bernal, G. Strength and medium-term impact of HisAK70 immunization in dogs: Vaccine safety and biomarkers of effectiveness for ex vivo Leishmania infantum infection. Comp. Immunol. Microbiol. Infect. Dis. 2019. [Google Scholar] [CrossRef] [PubMed]
- Carrion, J.; Abengozar, M.A.; Fernandez-Reyes, M.; Sanchez-Martin, C.; Rial, E.; Dominguez-Bernal, G.; Gonzalez-Barroso, M.M. UCP2 deficiency helps to restrict the pathogenesis of experimental cutaneous and visceral leishmaniosis in mice. PLoS Negl. Trop. Dis. 2013, 7, e2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Scott, P.; Pearce, E.; Natovitz, P.; Sher, A. Vaccination against cutaneous leishmaniasis in a murine model. I. Induction of protective immunity with a soluble extract of promastigotes. J. Immunol. 1987, 139, 221–227. [Google Scholar]
- Andreatta, M.; Nielsen, M. Gapped sequence alignment using artificial neural networks: Application to the MHC class I system. Bioinformatics 2016, 32, 511–517. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, M.; Lundegaard, C.; Worning, P.; Lauemoller, S.L.; Lamberth, K.; Buus, S.; Brunak, S.; Lund, O. Reliable prediction of T-cell epitopes using neural networks with novel sequence representations. Protein Sci. 2003, 12, 1007–1017. [Google Scholar] [CrossRef]
- Jensen, K.K.; Andreatta, M.; Marcatili, P.; Buus, S.; Greenbaum, J.A.; Yan, Z.; Sette, A.; Peters, B.; Nielsen, M. Improved methods for predicting peptide binding affinity to MHC class II molecules. Immunology 2018, 154, 394–406. [Google Scholar] [CrossRef]
- Trolle, T.; Nielsen, M. NetTepi: An integrated method for the prediction of T cell epitopes. Immunogenetics 2014, 66, 449–456. [Google Scholar] [CrossRef]
- Lutz, M.B.; Kukutsch, N.; Ogilvie, A.L.; Rossner, S.; Koch, F.; Romani, N.; Schuler, G. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]
- Dominguez-Bernal, G.; Martinez-Rodrigo, A.; Mas, A.; Blanco, M.M.; Orden, J.A.; De La Fuente, R.; Carrion, J. Alternative strategy for visceral leishmaniosis control: HisAK70-Salmonella Choleraesuis-pulsed dendritic cells. Comp. Immunol. Microbiol. Infect. Dis. 2017, 54, 13–19. [Google Scholar] [CrossRef]
- Martínez-Rodrigo, A.; Dias, D.S.; Ribeiro, P.A.F.; Roatt, B.M.; Mas, A.; Carrion, J.; Coelho, E.A.F.; Domínguez-Bernal, G. Immunization with the HisAK70 DNA Vaccine Induces Resistance against Leishmania Amazonensis Infection in BALB/c Mice. Vaccines 2019, 7, 183. [Google Scholar] [CrossRef] [Green Version]
- Buffet, P.A.; Sulahian, A.; Garin, Y.J.F.; Nassar, N.; Derouin, F. Culture Microtitration-a Sensitive Method for Quantifying Leishmania-Infantum in Tissues of Infected Mice. Antimicrob. Agents Chemother. 1995, 39, 2167–2168. [Google Scholar] [CrossRef] [Green Version]
- Duarte, M.C.; Lage, D.P.; Martins, V.T.; Chávez-Fumagalli, M.A.; Roatt, B.M.; Menezes-Souza, D.; Goulart, L.R.; Soto, M.; Tavares, C.A.P.; Coelho, E.A.F. Recent updates and perspectives on approaches for the development of vaccines against visceral leishmaniasis. Rev. Soc. Bras. Med. Trop. 2016, 49, 398–407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ding, A.H.; Nathan, C.F.; Stuehr, D.J. Release of Reactive Nitrogen Intermediates and Reactive Oxygen Intermediates from Mouse Peritoneal-Macrophages-Comparison of Activating Cytokines and Evidence for Independent Production. J. Immunol. 1988, 141, 2407–2412. [Google Scholar] [PubMed]
- Garrido, V.V.; Dulgerian, L.R.; Stempin, C.C.; Cerban, F.M. The Increase in Mannose Receptor Recycling Favors Arginase Induction and Trypanosoma Cruzi Survival in Macrophages. Int. J. Biol. Sci. 2011, 7, 1257–1272. [Google Scholar] [CrossRef] [Green Version]
- Rath, M.; Muller, I.; Kropf, P.; Closs, E.I.; Munder, M. Metabolism via arginase or nitric oxide synthase: Two competing arginine pathways in macrophages. Front. Immunol. 2014, 5, 532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, A.C.; Mattila, J.T. “Of mice and men”: Arginine metabolism in macrophages. Front. Immunol. 2014, 5, 479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duthie, M.S.; Reed, S.G. Not All Antigens Are Created Equally: Progress, Challenges, and Lessons Associated with Developing a Vaccine for Leishmaniasis. Clin. Vaccine Immunol. 2017, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glennie, N.D.; Volk, S.W.; Scott, P. Skin-resident CD4+ T cells protect against Leishmania major by recruiting and activating inflammatory monocytes. PLoS Pathog. 2017, 13, e1006349. [Google Scholar] [CrossRef]
- Agallou, M.; Pantazi, E.; Tsiftsaki, E.; Toubanaki, D.K.; Gaitanaki, C.; Smirlis, D.; Karagouni, E. Induction of protective cellular immune responses against experimental visceral leishmaniasis mediated by dendritic cells pulsed with the N-terminal domain of Leishmania infantum elongation factor-2 and CpG oligodeoxynucleotides. Mol. Immunol. 2018, 103, 7–20. [Google Scholar] [CrossRef]
- Dias, D.S.; Ribeiro, P.A.F.; Martins, V.T.; Lage, D.P.; Costa, L.E.; Chavez-Fumagalli, M.A.; Ramos, F.F.; Santos, T.T.O.; Ludolf, F.; Oliveira, J.S.; et al. Vaccination with a CD4(+) and CD8(+) T-cell epitopes-based recombinant chimeric protein derived from Leishmania infantum proteins confers protective immunity against visceral leishmaniasis. Transl. Res. 2018. [Google Scholar] [CrossRef] [PubMed]
- Martins, V.T.; Duarte, M.C.; Lage, D.P.; Costa, L.E.; Carvalho, A.M.; Mendes, T.A.; Roatt, B.M.; Menezes-Souza, D.; Soto, M.; Coelho, E.A. A recombinant chimeric protein composed of human and mice-specific CD4(+) and CD8(+) T-cell epitopes protects against visceral leishmaniasis. Parasite Immunol. 2017, 39. [Google Scholar] [CrossRef] [PubMed]
- Barth, S.M.; Schreitmuller, C.M.; Proehl, F.; Oehl, K.; Lumpp, L.M.; Kowalewski, D.J.; Di Marco, M.; Sturm, T.; Backert, L.; Schuster, H.; et al. Characterization of the Canine MHC Class I DLA-88*50101 Peptide Binding Motif as a Prerequisite for Canine T Cell Immunotherapy. PLoS ONE 2016, 11, e0167017. [Google Scholar] [CrossRef] [PubMed]
- Debenham, S.L.; Hart, E.A.; Ashurst, J.L.; Howe, K.L.; Quail, M.A.; Ollier, W.E.R.; Binns, M.M. Genomic sequence of the class II region of the canine MHC: Comparison with the MHC of other mammalian species. Genomics 2005, 85, 48–59. [Google Scholar] [CrossRef]
- Agallou, M.; Margaroni, M.; Athanasiou, E.; Toubanaki, D.K.; Kontonikola, K.; Karidi, K.; Kammona, O.; Kiparissides, C.; Karagouni, E. Identification of BALB/c Immune Markers Correlated with a Partial Protection to Leishmania infantum after Vaccination with a Rationally Designed Multi-epitope Cysteine Protease A Peptide-Based Nanovaccine. PLoS Negl. Trop. Dis. 2017, 11, e0005311. [Google Scholar] [CrossRef]
- Kaye, P.; Scott, P. Leishmaniasis: Complexity at the host-pathogen interface. Nat. Rev. 2011, 9, 604–615. [Google Scholar] [CrossRef]
- Uzonna, J.E.; Spath, G.F.; Beverley, S.M.; Scott, P. Vaccination with phosphoglycan-deficient Leishmania major protects highly susceptible mice from virulent challenge without inducing a strong Th1 response. J Immunol. 2004, 172, 3793–3797. [Google Scholar] [CrossRef] [Green Version]
- Kaye, P.M.; Cruz, I.; Picado, A.; Van Bocxlaer, K.; Croft, S.L. Leishmaniasis immunopathology-impact on design and use of vaccines, diagnostics and drugs. Semin. Immunopathol. 2020. [Google Scholar] [CrossRef]
- Martins, V.T.; Chavez-Fumagalli, M.A.; Lage, D.P.; Duarte, M.C.; Garde, E.; Costa, L.E.; da Silva, V.G.; Oliveira, J.S.; de Magalhaes-Soares, D.F.; Teixeira, S.M.R.; et al. Antigenicity, Immunogenicity and Protective Efficacy of Three Proteins Expressed in the Promastigote and Amastigote Stages of Leishmania infantum against Visceral Leishmaniasis. PLoS ONE 2015, 10, e0141496. [Google Scholar] [CrossRef] [Green Version]
- Lage, D.P.; Martins, V.T.; Duarte, M.C.; Garde, E.; Chavez-Fumagalli, M.A.; Menezes-Souza, D.; Roatt, B.M.; Tavares, C.A.P.; Soto, M.; Coelho, E.A.F. Prophylactic properties of a Leishmania-specific hypothetical protein in a murine model of visceral leishmaniasis. Parasite Immunol. 2015, 37, 646–656. [Google Scholar] [CrossRef] [Green Version]
- Gerth, A.J.; Lin, L.; Peng, S.L. T-bet regulates T-independent IgG2a class switching. Int. Immunol. 2003, 15, 937–944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bordbar, A.; Bagheri, K.P.; Ebrahimi, S.; Parvizi, P. Bioinformatics analyses of immunogenic T-cell epitopes of LeIF and PpSP15 proteins from Leishmania major and sand fly saliva used as model antigens for the design of a multi-epitope vaccine to control leishmaniasis. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2020, 80, 104189. [Google Scholar] [CrossRef] [PubMed]
- Maroof, A.; Brown, N.; Smith, B.; Hodgkinson, M.R.; Maxwell, A.; Losch, F.O.; Fritz, U.; Walden, P.; Lacey, C.N.J.; Smith, D.F.; et al. Therapeutic Vaccination With Recombinant Adenovirus Reduces Splenic Parasite Burden in Experimental Visceral Leishmaniasis. J. Infect. Dis. 2012, 205, 853–863. [Google Scholar] [CrossRef] [PubMed]
- Margaroni, M.; Agallou, M.; Athanasiou, E.; Kammona, O.; Kiparissides, C.; Gaitanaki, C.; Karagouni, E. Vaccination with poly(D,L-lactide-co-glycolide) nanoparticles loaded with soluble Leishmania antigens and modified with a TNFalpha-mimicking peptide or monophosphoryl lipid A confers protection against experimental visceral leishmaniasis. Int. J. Nanomed. 2017, 12, 6169–6184. [Google Scholar] [CrossRef] [Green Version]
- Peleteiro, M.; Presas, E.; Gonzalez-Aramundiz, J.V.; Sanchez-Correa, B.; Simon-Vazquez, R.; Csaba, N.; Alonso, M.J.; Gonzalez-Fernandez, A. Polymeric Nanocapsules for Vaccine Delivery: Influence of the Polymeric Shell on the Interaction With the Immune System. Front. Immunol. 2018, 9. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MHC-I | MHC-II | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Protein | Selected Peptide | HLA-A | HLA-B | HLA-C and E | H-2 | HLA-DR | HLA-DP | HLA-DQ | H-2 |
| H2A | TAVLEYLTAELLELS | 13 | 6 | 8 | 1 | 5 | 9 | 15 | X |
| H2B | RSLKAINAQMSMSHRTMKIVNSYV | 16 | 12 | 9 | X | 16 | 2 | 11 | 4 |
| H3 | EGLRFQSSAIMALQE | 10 | 3 | 8 | X | 17 | 5 | 12 | 3 |
| H4 | ITRGCVRRMARRGGVK | X | 6 | X | X | 6 | 1 | 3 | 2 |
| A2 | SAEPHKAAVDVGPLSVGPQSVGPLSVGPQA | X | X | X | 4 | 7 | 7 | 8 | 10 |
| KMP-11 | KMHE12HSEHFKQKFAELLEQQKA | 3 | 16 | 5 | X | 2 | 7 | 2 | 2 |
| Groups: | His | AK | His-AK | Saponin | PBS |
|---|---|---|---|---|---|
| 24 h | 3.80 ± 0.43 ##,a | 9.60 ± 1.40 ## | 6.94 ± 2.00 ## | 19.23 ± 2.18 | 21.17 ± 3.67 |
| 72 h | 3.97 ± 1.28 ## | 6.59 ± 1.30 # | 5.60 ± 1.38 ## | 12.63 ± 2.62 | 12.43 ± 2.16 |
| Groups: | mU Arginase Activity | µM Nitrites |
|---|---|---|
| His | 7.70 ± 1.10 ##,a | 14.53 ± 6.79 ##,a |
| AK | 20.70 ± 4.40 b | 7.57 ± 4.35 # |
| His-AK | 12.18 ± 1.81 ## | 10.77 ± 5.47 # |
| Saponin | 29.64 ± 6.32 c | 1.67 ± 0.35 |
| PBS | 38.97 ± 3.10 | 2.20 ± 1.14 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-Rodrigo, A.; Mas, A.; Álvarez-Campos, D.; Orden, J.A.; Domínguez-Bernal, G.; Carrión, J. Epitope Selection for Fighting Visceral Leishmaniosis: Not All Peptides Function the Same Way. Vaccines 2020, 8, 352. https://doi.org/10.3390/vaccines8030352
Martínez-Rodrigo A, Mas A, Álvarez-Campos D, Orden JA, Domínguez-Bernal G, Carrión J. Epitope Selection for Fighting Visceral Leishmaniosis: Not All Peptides Function the Same Way. Vaccines. 2020; 8(3):352. https://doi.org/10.3390/vaccines8030352
Chicago/Turabian StyleMartínez-Rodrigo, Abel, Alicia Mas, Daniel Álvarez-Campos, José A. Orden, Gustavo Domínguez-Bernal, and Javier Carrión. 2020. "Epitope Selection for Fighting Visceral Leishmaniosis: Not All Peptides Function the Same Way" Vaccines 8, no. 3: 352. https://doi.org/10.3390/vaccines8030352