Genetically Modified Mouse Mesenchymal Stem Cells Expressing Non-Structural Proteins of Hepatitis C Virus Induce Effective Immune Response

 , , , , , ,
, , , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Isolation of Primary MSCs
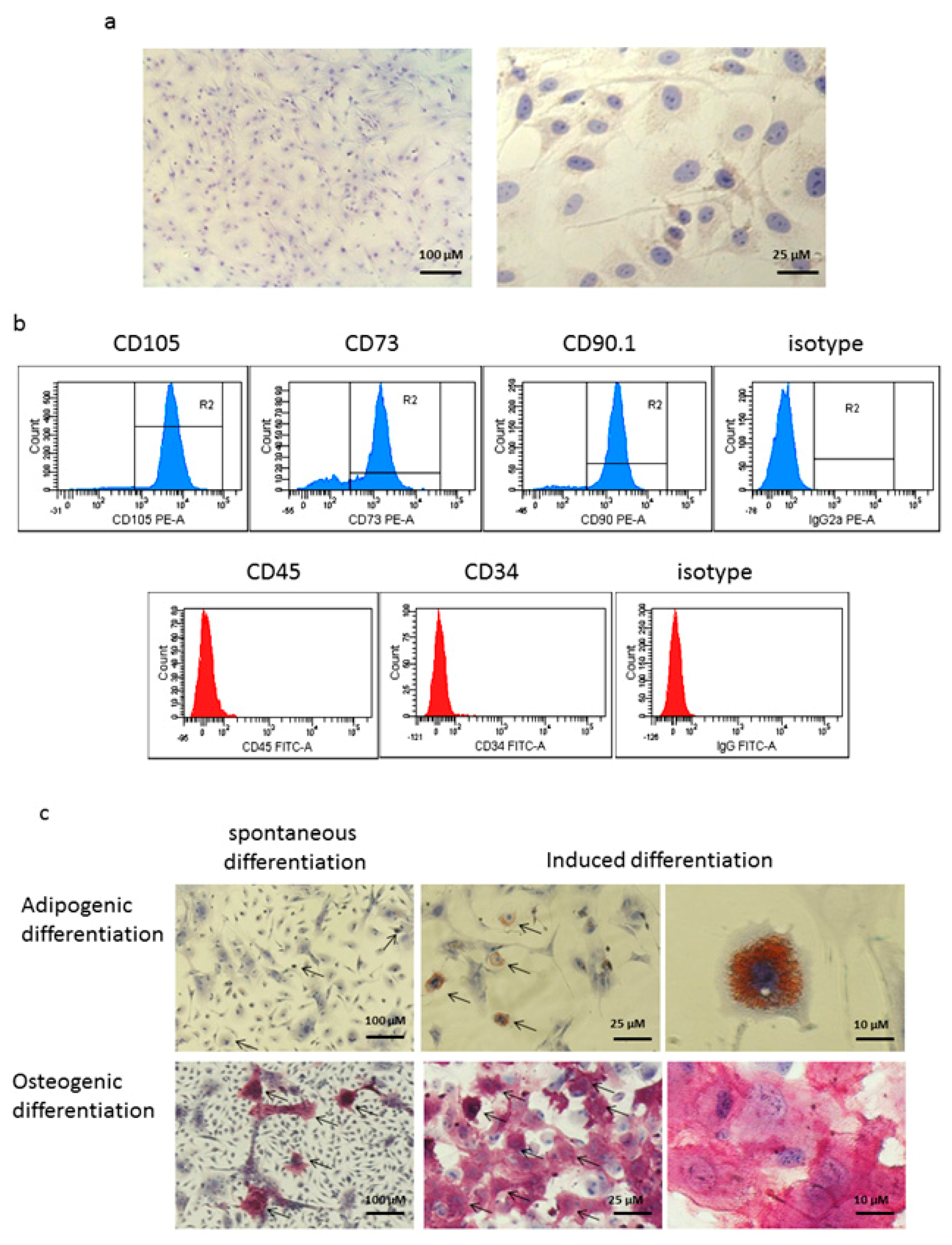
2.3. Characterization of MSCs
2.4. Assessment of Adipogenic and Osteogenic Potencies of MSCs
2.5. Plasmid and Transfection of MSC Culture
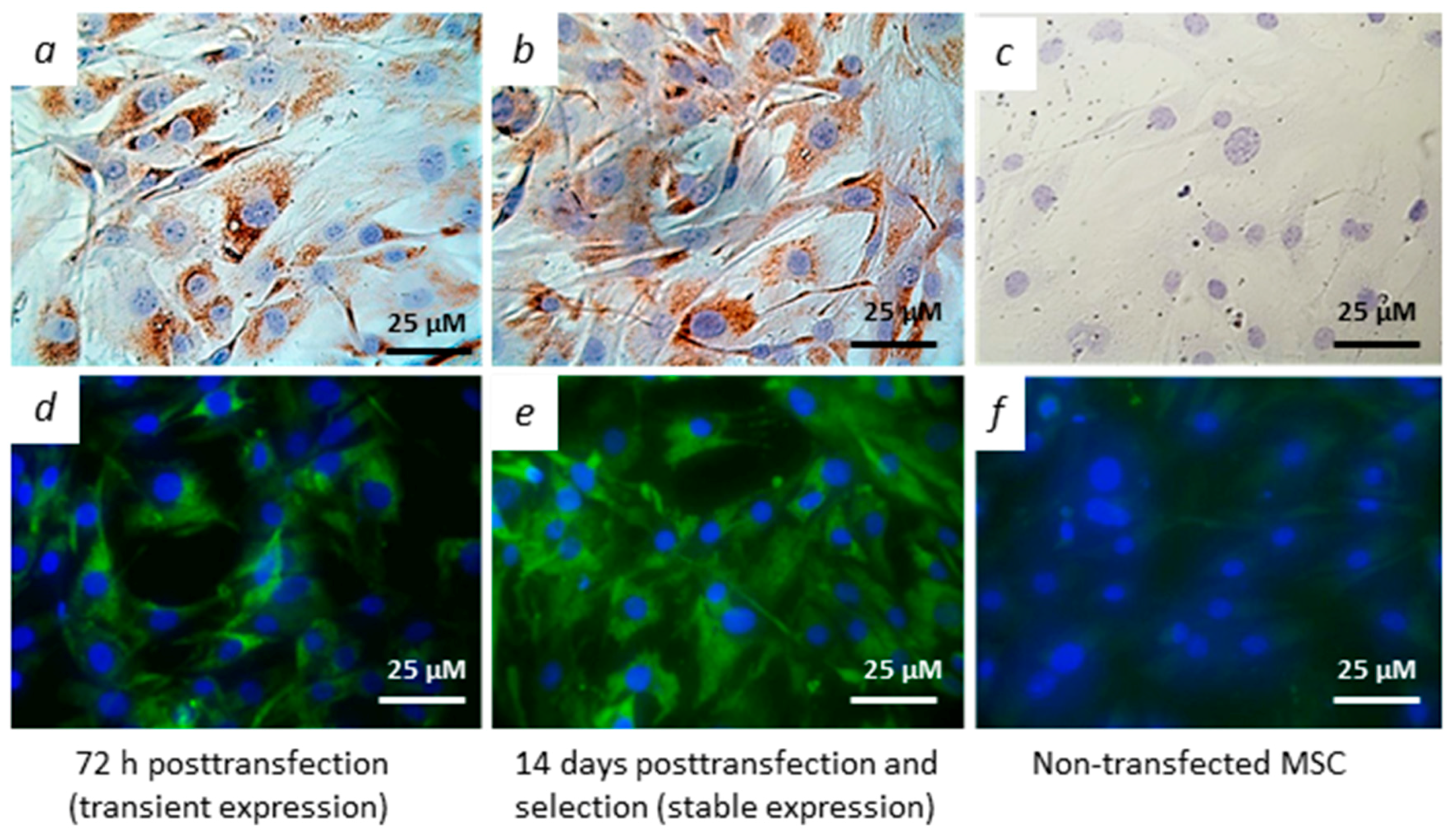
2.6. Immunocytochemical and Immunoblot Detection of Viral Proteins
2.7. Immunization of Animals
2.8. The Recombinant HCV Proteins
2.9. Humoral Immune Response
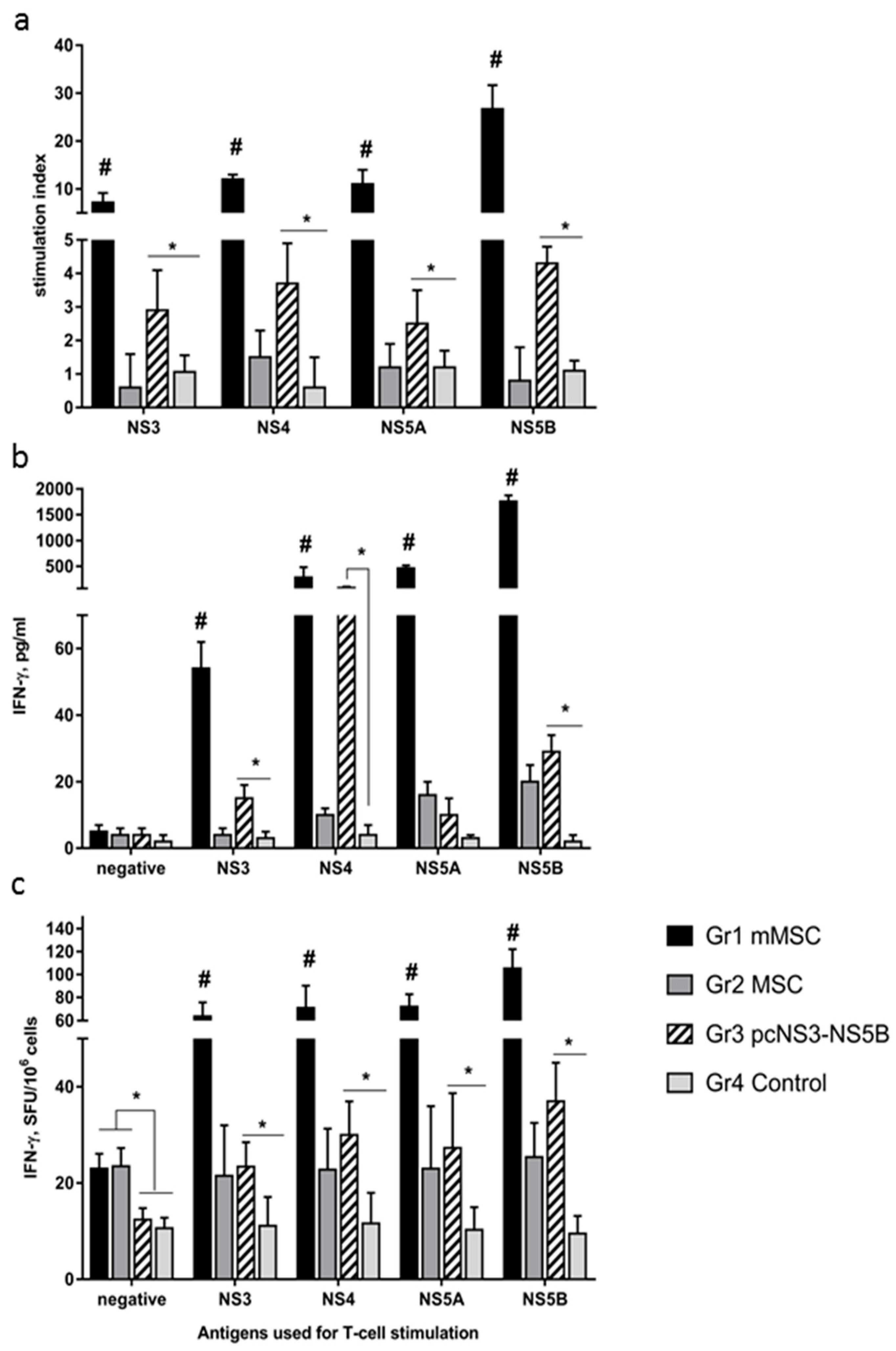
2.10. T-Cell Proliferation and ELISpot Assays
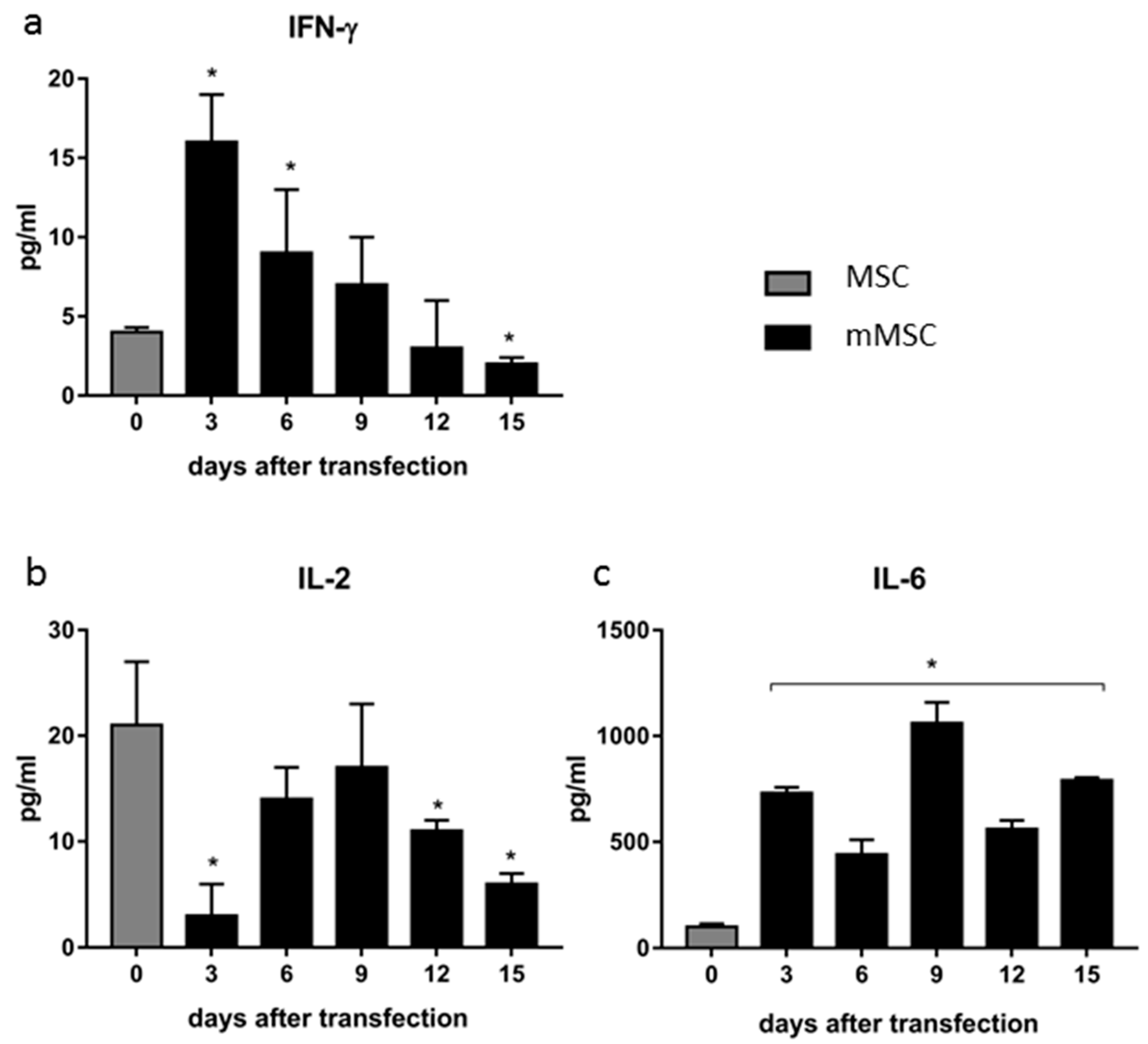
2.11. Detection of Cytokines in Cell Culture Fluids by Sandwich ELISA
2.12. Flow Cytometry
2.13. Determination of Type I IFNs (IFN-α/β) Production by Immune Cells
2.14. Phagocytic Activity
2.15. Statistical Analysis
3. Results
3.1. Cells Isolated From the Bone Marrow of Mice Display Features of Mesenchymal Stem Cells
3.2. Modified MSCs Express Genes of Hepatitis C Virus
3.3. Dynamics of Cytokine Production in Transfected MSCs
3.4. Immune Response to Administration of Modified MSCs to Mice Exceeds Immune Response to Plasmid
3.5. Decrease in Proportion of Myeloid Derived Suppressor Cells in Spleens of Mice Immunized with MSCs
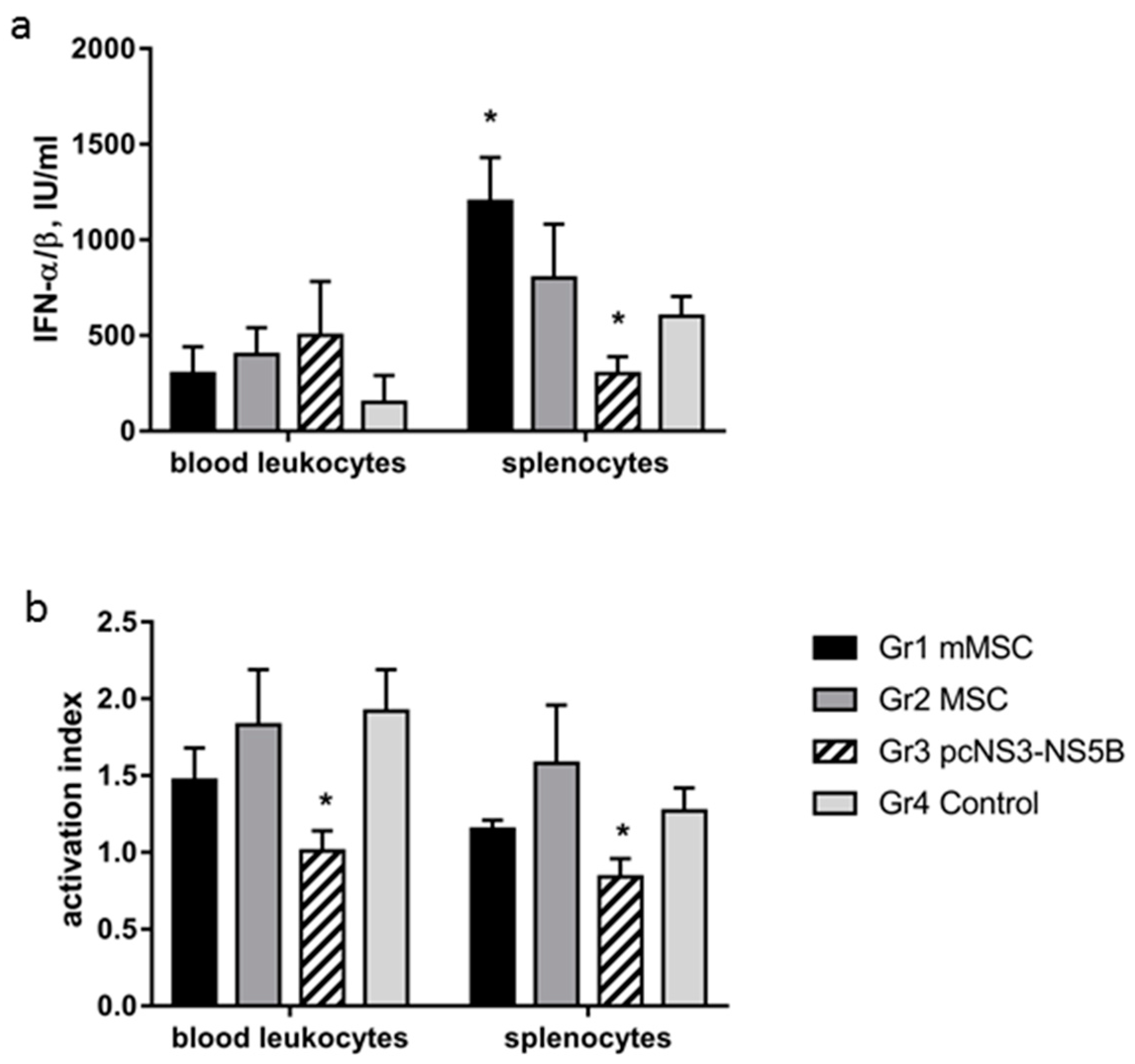
3.6. Changes in Activity of IFN-α/β and in Phagocytic Activity of Immune Cells of Immunized Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Yuan, X.; Logan, T.M.; Ma, T. Metabolism in human mesenchymal stromal cells: A missing link between hmsc biomanufacturing and therapy? Front. Immunol. 2019, 10, 977. [Google Scholar] [CrossRef] [Green Version]
- Dameshghi, S.; Zavaran-Hosseini, A.; Soudi, S.; Shirazi, F.J.; Nojehdehi, S.; Hashemi, S.M. Mesenchymal stem cells alter macrophage immune responses to leishmania major infection in both susceptible and resistance mice. Immunol. Lett. 2016, 170, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Goodarzi, P.; Larijani, B.; Alavi-Moghadam, S.; Tayanloo-Beik, A.; Mohamadi-Jahani, F.; Ranjbaran, N.; Payab, M.; Falahzadeh, K.; Mousavi, M.; Arjmand, B. Mesenchymal stem cells-derived exosomes for wound regeneration. Adv. Exp. Med. Biol. 2018, 1119, 119–131. [Google Scholar]
- Lukashyk, S.P.; Tsyrkunov, V.M.; Isaykina, Y.I.; Romanova, O.N.; Shymanskiy, A.T.; Aleynikova, O.V.; Kravchuk, R.I. Mesenchymal bone marrow-derived stem cells transplantation in patients with HCV related liver cirrhosis. J. Clin. Transl. Hepatol. 2014, 2, 217–221. [Google Scholar] [PubMed] [Green Version]
- Munir, H.; McGettrick, H.M. Mesenchymal stem cell therapy for autoimmune disease: Risks and rewards. Stem Cells Dev. 2015, 24, 2091–2100. [Google Scholar] [CrossRef] [PubMed]
- Cobo, M.; Anderson, P.; Benabdellah, K.; Toscano, M.G.; Munoz, P.; Garcia-Perez, A.; Gutierrez, I.; Delgado, M.; Martin, F. Mesenchymal stem cells expressing vasoactive intestinal peptide ameliorate symptoms in a model of chronic multiple sclerosis. Cell Transp. 2013, 22, 839–854. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wyse, R.D.; Dunbar, G.L.; Rossignol, J. Use of genetically modified mesenchymal stem cells to treat neurodegenerative diseases. Int. J. Mol. Sci. 2014, 15, 1719–1745. [Google Scholar] [CrossRef] [Green Version]
- Marin-Banasco, C.; Benabdellah, K.; Melero-Jerez, C.; Oliver, B.; Pinto-Medel, M.J.; Hurtado-Guerrero, I.; de Castro, F.; Clemente, D.; Fernandez, O.; Martin, F.; et al. Gene therapy with mesenchymal stem cells expressing ifn-ss ameliorates neuroinflammation in experimental models of multiple sclerosis. Br. J. Pharmacol. 2017, 174, 238–253. [Google Scholar] [CrossRef]
- Xue, X.; Liu, Y.; Zhang, J.; Liu, T.; Yang, Z.; Wang, H. BCL-XL genetic modification enhanced the therapeutic efficacy of mesenchymal stem cell transplantation in the treatment of heart infarction. Stem Cells Int. 2015, 2015, 176409. [Google Scholar] [CrossRef] [Green Version]
- Haber, T.; Baruch, L.; Machluf, M. Ultrasound-mediated mesenchymal stem cells transfection as a targeted cancer therapy platform. Sci. Rep. 2017, 7, 42046. [Google Scholar] [CrossRef] [Green Version]
- Tomchuck, S.L.; Norton, E.B.; Garry, R.F.; Bunnell, B.A.; Morris, C.A.; Freytag, L.C.; Clements, J.D. Mesenchymal stem cells as a novel vaccine platform. Front. Cell. Infect. Microbiol. 2012, 2, 140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.H.; Rehermann, B. Immune responses to hcv and other hepatitis viruses. Immunity 2014, 40, 13–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dustin, L.B. Innate and adaptive immune responses in chronic HCV infection. Curr. Drug Targets 2017, 18, 826–843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pawlotsky, J.M. Hepatitis C virus: Standard-of-care treatment. Adv. Pharmacol. 2013, 67, 169–215. [Google Scholar]
- Spearman, C.W.; Dusheiko, G.M.; Hellard, M.; Sonderup, M. Hepatitis C. Lancet 2019, 394, 1451–1466. [Google Scholar] [CrossRef]
- Wang, Y.; Rao, H.; Chi, X.; Li, B.; Liu, H.; Wu, L.; Zhang, H.; Liu, S.; Zhou, G.; Li, N.; et al. Detection of residual HCV-RNA in patients who have achieved sustained virological response is associated with persistent histological abnormality. EBioMedicine 2019, 46, 227–235. [Google Scholar] [CrossRef] [Green Version]
- Elmasry, S.; Wadhwa, S.; Bang, B.R.; Cook, L.; Chopra, S.; Kanel, G.; Kim, B.; Harper, T.; Feng, Z.; Jerome, K.R.; et al. Detection of occult hepatitis C virus infection in patients who achieved a sustained virologic response to direct-acting antiviral agents for recurrent infection after liver transplantation. Gastroenterology 2017, 152, 550–553. [Google Scholar] [CrossRef] [Green Version]
- Di Lorenzo, C.; Angus, A.G.; Patel, A.H. Hepatitis C virus evasion mechanisms from neutralizing antibodies. Viruses 2011, 3, 2280–2300. [Google Scholar] [CrossRef]
- Ahlen, G.; Frelin, L. Methods to evaluate novel hepatitis C virus vaccines. Methods Mol. Biol. 2016, 1403, 221–244. [Google Scholar]
- Verma, R.; Khanna, P.; Chawla, S. Hepatitis C vaccine. Need of the hour. Hum. Vaccin. Immunother. 2014, 10, 1927–1929. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.; Fernandez, S.; Toro, F.; De Sanctis, J.B. An overview of hepatitis C vaccines. Recent Pat. Inflamm. Allergy Drug Discov. 2014, 8, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Wintermeyer, P.; Gehring, S.; Eken, A.; Wands, J.R. Generation of cellular immune responses to HCV ns5 protein through in vivo activation of dendritic cells. J. Viral Hepat. 2010, 17, 705–713. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.; Babiuk, L.A.; van Drunen Littel-van den Hurk, S. Strategies for loading dendritic cells with hepatitis c ns5a antigen and inducing protective immunity. J. Viral Hepat. 2008, 15, 459–470. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Y.; Yao, Z.; Moorman, J.P.; Jia, Z. Dendritic cell-based immunity and vaccination against hepatitis C virus infection. Immunology 2012, 136, 385–396. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chernykh, E.; Leplina, O.; Oleynik, E.; Tikhonova, M.; Tyrinova, T.; Starostina, N.; Ostanin, A. Immunotherapy with interferon-alpha-induced dendritic cells for chronic hcv infection (the results of pilot clinical trial). Immunol. Res. 2018, 66, 31–43. [Google Scholar] [CrossRef]
- Zabaleta, A.; D’Avola, D.; Echeverria, I.; Llopiz, D.; Silva, L.; Villanueva, L.; Riezu-Boj, J.I.; Larrea, E.; Pereboev, A.; Lasarte, J.J.; et al. Clinical testing of a dendritic cell targeted therapeutic vaccine in patients with chronic hepatitis c virus infection. Mol. Ther. Methods Clin. Dev. 2015, 2, 15006. [Google Scholar] [CrossRef]
- Bailey, J.R.; Barnes, E.; Cox, A.L. Approaches, progress, and challenges to hepatitis C vaccine development. Gastroenterology 2019, 156, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Thimme, R.; Oldach, D.; Chang, K.M.; Steiger, C.; Ray, S.C.; Chisari, F.V. Determinants of viral clearance and persistence during acute hepatitis C virus infection. J. Exp. Med. 2001, 194, 1395–1406. [Google Scholar] [CrossRef]
- Wijesundara, D.K.; Gummow, J.; Li, Y.; Yu, W.; Quah, B.J.; Ranasinghe, C.; Torresi, J.; Gowans, E.J.; Grubor-Bauk, B. Induction of genotype cross-reactive, hepatitis C virus-specific, cell-mediated immunity in DNA-vaccinated mice. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [Green Version]
- Masalova, O.V.; Lesnova, E.I.; Ivanov, A.V.; Pichugin, A.V.; Permiakova, K.; Smirnova, O.A.; Tynitskaia, V.L.; Ulanova, T.I.; Burkov, A.N.; Kochetkov, S.N.; et al. Comparative analysis of the immune response to DNA constructions encoding hepatitis c virus nonstructural proteins. Vopr. Virusol. 2013, 58, 21–28. [Google Scholar]
- Tolosa, L.; Donato, M.T.; Gomez-Lechon, M.J. General cytotoxicity assessment by means of the MTT assay. Methods Mol. Biol. 2015, 1250, 333–348. [Google Scholar] [PubMed]
- Schlueter, N.; Lussi, A.; Ganss, C.; Gruber, R. L929 fibroblast bioassay on the in vitro toxicity of SNCL2, h3po4, clearfil se primer and combinations thereof. Swiss Dent. J. 2016, 126, 566–572. [Google Scholar] [PubMed]
- Rechkina, E.A.; Denisova, G.F.; Masalova, O.V.; Lideman, L.F.; Denisov, D.A.; Lesnova, E.I.; Ataullakhanov, R.I.; Gur’ianova, S.V.; Kushch, A.A. Epitope mapping of antigenic determinants of hepatitis C virus proteins by phage display. Mol. Biol. (Mosk) 2006, 40, 312–323. [Google Scholar] [CrossRef]
- Masalova, O.V.; Lesnova, E.I.; Solyev, P.N.; Zakirova, N.F.; Prassolov, V.S.; Kochetkov, S.N.; Ivanov, A.V.; Kushch, A.A. Modulation of cell death pathways by hepatitis C virus proteins in huh7.5 hepatoma cells. Int. J. Mol. Sci. 2017, 18, 2346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masalova, O.V.; Lesnova, E.I.; Pichugin, A.V.; Melnikova, T.M.; Grabovetsky, V.V.; Petrakova, N.V.; Smirnova, O.A.; Ivanov, A.V.; Zaberezhny, A.D.; Ataullakhanov, R.I.; et al. The successful immune response against hepatitis C nonstructural protein 5a (ns5a) requires heterologous DNA/protein immunization. Vaccine 2010, 28, 1987–1996. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Smirnova, O.A.; Ivanova, O.N.; Masalova, O.V.; Kochetkov, S.N.; Isaguliants, M.G. Hepatitis C virus proteins activate nrf2/are pathway by distinct ROS-dependent and independent mechanisms in huh7 cells. PLoS ONE 2011, 6, e24957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukovnya, A.V.; Tunitskaya, V.L.; Khandazhinskaya, A.L.; Golubeva, N.A.; Zakirova, N.F.; Ivanov, A.V.; Kukhanova, M.K.; Kochetkov, S.N. Hepatitis C virus helicase/ntpase: An efficient expression system and new inhibitors. Biochemistry (Moscow) 2008, 73, 660–668. [Google Scholar] [CrossRef]
- Ivanov, A.V.; Korovina, A.N.; Tunitskaya, V.L.; Kostyuk, D.A.; Rechinsky, V.O.; Kukhanova, M.K.; Kochetkov, S.N. Development of the system ensuring a high-level expression of hepatitis C virus nonstructural ns5b and ns5a proteins. Protein Expr. Purif. 2006, 48, 14–23. [Google Scholar] [CrossRef]
- Ulanova, T.I.; Puzyrev, V.F.; Burkov, A.N.; Obriadina, A.P. Impact of the heterogenicity of amino acid sequence on the immunoreactivity of an antigenic epitopic complex localized within amino acids 1192-1456 of protein ns3 protein of hepatitis c virus. Vopr. Virusol. 2006, 51, 28–30. [Google Scholar]
- Babaiants, A.A.; Manakhova, L.S.; Karganova, G.G.; Porkhovatyi, S.; Kondratenko, I.V. Leukocyte interferon reaction in patients with primary immunodeficiencies. Vopr. Virusol. 1985, 30, 714–717. [Google Scholar]
- Vlaspolder, F.; Donkers, E.; Harmsen, T.; Kraaijeveld, C.A.; Snippe, H. Rapid bioassay of human interferon by direct enzyme immunoassay of encephalomyocarditis virus in hep-2 cell monolayers after a single cycle of infection. J. Virol. Methods 1989, 24, 153–158. [Google Scholar] [CrossRef]
- Fournier, P.; Wilden, H.; Schirrmacher, V. Importance of retinoic acid-inducible gene I and of receptor for type I interferon for cellular resistance to infection by newcastle disease virus. Int. J. Oncol. 2012, 40, 287–298. [Google Scholar] [PubMed] [Green Version]
- Stewart, W.E., II. The Interferon System; Springer-Verlag: New York, NY, USA, 1979. [Google Scholar]
- Hasegawa, H.; Suzuki, K.; Nakaji, S.; Sugawara, K. Analysis and assessment of the capacity of neutrophils to produce reactive oxygen species in a 96-well microplate format using lucigenin- and luminol-dependent chemiluminescence. J. Immunol. Methods 1997, 210, 1–10. [Google Scholar] [CrossRef]
- Dominici, M.; Le Blanc, K.; Mueller, I.; Slaper-Cortenbach, I.; Marini, F.; Krause, D.; Deans, R.; Keating, A.; Prockop, D.; Horwitz, E. Minimal criteria for defining multipotent mesenchymal stromal cells. The international society for cellular therapy position statement. Cytotherapy 2006, 8, 315–317. [Google Scholar] [CrossRef]
- Zhao, L.; Chen, S.; Yang, P.; Cao, H.; Li, L. The role of mesenchymal stem cells in hematopoietic stem cell transplantation: Prevention and treatment of graft-versus-host disease. Stem Cell Res. Ther. 2019, 10, 182. [Google Scholar] [CrossRef]
- Rivera-Cruz, C.M.; Shearer, J.J.; Figueiredo Neto, M.; Figueiredo, M.L. The immunomodulatory effects of mesenchymal stem cell polarization within the tumor microenvironment niche. Stem Cells Int. 2017, 2017, 4015039. [Google Scholar] [CrossRef] [Green Version]
- Volarevic, V.; Gazdic, M.; Simovic Markovic, B.; Jovicic, N.; Djonov, V.; Arsenijevic, N. Mesenchymal stem cell-derived factors: Immuno-modulatory effects and therapeutic potential. Biofactors 2017, 43, 633–644. [Google Scholar] [CrossRef]
- Crop, M.J.; Baan, C.C.; Korevaar, S.S.; Ijzermans, J.N.; Weimar, W.; Hoogduijn, M.J. Human adipose tissue-derived mesenchymal stem cells induce explosive T-cell proliferation. Stem Cells Dev. 2010, 19, 1843–1853. [Google Scholar] [CrossRef]
- Bolhassani, A.; Shahbazi, S.; Agi, E.; Haghighipour, N.; Hadi, A.; Asgari, F. Modified DCs and MSCs with HPV e7 antigen and small Hsps: Which one is the most potent strategy for eradication of tumors? Mol. Immunol. 2019, 108, 102–110. [Google Scholar] [CrossRef]
- Ank, N.; West, H.; Bartholdy, C.; Eriksson, K.; Thomsen, A.R.; Paludan, S.R. Lambda interferon (ifn-lambda), a type III IFN, is induced by viruses and IFNs and displays potent antiviral activity against select virus infections in vivo. J. Virol. 2006, 80, 4501–4509. [Google Scholar] [CrossRef] [Green Version]
- Jin, J.; Yang, J.Y.; Liu, J.; Kong, Y.Y.; Wang, Y.; Li, G.D. DNA immunization with fusion genes encoding different regions of hepatitis C virus e2 fused to the gene for hepatitis b surface antigen elicits immune responses to both hcv and hbv. World J. Gastroenterol. 2002, 8, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Chmielewska, A.M.; Naddeo, M.; Capone, S.; Ammendola, V.; Hu, K.; Meredith, L.; Verhoye, L.; Rychlowska, M.; Rappuoli, R.; Ulmer, J.B.; et al. Combined adenovirus vector and hepatitis c virus envelope protein prime-boost regimen elicits t cell and neutralizing antibody immune responses. J. Virol. 2014, 88, 5502–5510. [Google Scholar] [CrossRef] [Green Version]
- Rasmusson, I.; Le Blanc, K.; Sundberg, B.; Ringden, O. Mesenchymal stem cells stimulate antibody secretion in human B cells. Scand. J. Immunol. 2007, 65, 336–343. [Google Scholar] [CrossRef] [PubMed]
- Splawski, J.B.; McAnally, L.M.; Lipsky, P.E. Il-2 dependence of the promotion of human B cell differentiation by il-6 (bsf-2). J. Immunol. 1990, 144, 562–569. [Google Scholar] [PubMed]
- Borgia, G.; Maraolo, A.E.; Nappa, S.; Gentile, I.; Buonomo, A.R. Ns5b polymerase inhibitors in phase II clinical trials for HCV infection. Expert Opin Investig Drugs 2018, 27, 243–250. [Google Scholar] [CrossRef]
- Rao, X.; Hoof, I.; van Baarle, D.; Kesmir, C.; Textor, J. HLA preferences for conserved epitopes: A potential mechanism for hepatitis c clearance. Front. Immunol. 2015, 6, 552. [Google Scholar] [CrossRef] [Green Version]
- O’Connor, M.A.; Rastad, J.L.; Green, W.R. The role of myeloid-derived suppressor cells in viral infection. Viral. Immunol. 2017, 30, 82–97. [Google Scholar] [CrossRef]
- Cai, W.; Qin, A.; Guo, P.; Yan, D.; Hu, F.; Yang, Q.; Xu, M.; Fu, Y.; Zhou, J.; Tang, X. Clinical significance and functional studies of myeloid-derived suppressor cells in chronic hepatitis C patients. J. Clin. Immunol. 2013, 33, 798–808. [Google Scholar] [CrossRef]
- Goh, C.C.; Roggerson, K.M.; Lee, H.C.; Golden-Mason, L.; Rosen, H.R.; Hahn, Y.S. Hepatitis c virus-induced myeloid-derived suppressor cells suppress NK cell IFN-gamma production by altering cellular metabolism via arginase-1. J. Immunol. 2016, 196, 2283–2292. [Google Scholar] [CrossRef] [Green Version]
- Qin, A.; Cai, W.; Pan, T.; Wu, K.; Yang, Q.; Wang, N.; Liu, Y.; Yan, D.; Hu, F.; Guo, P.; et al. Expansion of monocytic myeloid-derived suppressor cells dampens T cell function in hiv-1-seropositive individuals. J. Virol. 2013, 87, 1477–1490. [Google Scholar] [CrossRef] [Green Version]
- Pallett, L.J.; Gill, U.S.; Quaglia, A.; Sinclair, L.V.; Jover-Cobos, M.; Schurich, A.; Singh, K.P.; Thomas, N.; Das, A.; Chen, A.; et al. Metabolic regulation of hepatitis b immunopathology by myeloid-derived suppressor cells. Nat. Med. 2015, 21, 591–600. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bianchi, G.; Borgonovo, G.; Pistoia, V.; Raffaghello, L. Immunosuppressive cells and tumour microenvironment: Focus on mesenchymal stem cells and myeloid derived suppressor cells. Histol. Histopathol. 2011, 26, 941–951. [Google Scholar] [PubMed]
- Yen, B.L.; Yen, M.L.; Hsu, P.J.; Liu, K.J.; Wang, C.J.; Bai, C.H.; Sytwu, H.K. Multipotent human mesenchymal stromal cells mediate expansion of myeloid-derived suppressor cells via hepatocyte growth factor/C-met and STAT3. Stem Cell Rep. 2013, 1, 139–151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.W.; Chen, H.Y.; Wang, L.T.; Wang, F.H.; Fang, L.W.; Lai, H.Y.; Chen, H.H.; Lu, J.; Hung, M.S.; Cheng, Y.; et al. Mesenchymal stem cells tune the development of monocyte-derived dendritic cells toward a myeloid-derived suppressive phenotype through growth-regulated oncogene chemokines. J. Immunol. 2013, 190, 5065–5077. [Google Scholar] [CrossRef] [Green Version]
- Zheng, H.; Zou, W.; Shen, J.; Xu, L.; Wang, S.; Fu, Y.X.; Fan, W. Opposite effects of coinjection and distant injection of mesenchymal stem cells on breast tumor cell growth. Stem Cells Transl. Med. 2016, 5, 1216–1228. [Google Scholar] [CrossRef] [Green Version]
- Svobodova, E.; Krulova, M.; Zajicova, A.; Pokorna, K.; Prochazkova, J.; Trosan, P.; Holan, V. The role of mouse mesenchymal stem cells in differentiation of naive t-cells into anti-inflammatory regulatory T-cell or proinflammatory helper T-cell 17 population. Stem Cells Dev. 2012, 21, 901–910. [Google Scholar] [CrossRef] [Green Version]
- Kim, M.; Rhee, J.K.; Choi, H.; Kwon, A.; Kim, J.; Lee, G.D.; Jekarl, D.W.; Lee, S.; Kim, Y.; Kim, T.M. Passage-dependent accumulation of somatic mutations in mesenchymal stromal cells during in vitro culture revealed by whole genome sequencing. Sci. Rep. 2017, 7, 14508. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.Y.; He, G.; Li, C.Y.; Tang, M.; Grivennikov, S.; Tsai, W.T.; Wu, M.S.; Hsu, C.W.; Tsai, Y.; Wang, L.H.; et al. Hepatic expression of hcv RNA-dependent RNA polymerase triggers innate immune signaling and cytokine production. Mol. Cell. 2012, 48, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Nishitsuji, H.; Funami, K.; Shimizu, Y.; Ujino, S.; Sugiyama, K.; Seya, T.; Takaku, H.; Shimotohno, K. Hepatitis C virus infection induces inflammatory cytokines and chemokines mediated by the cross talk between hepatocytes and stellate cells. J. Virol. 2013, 87, 8169–8178. [Google Scholar] [CrossRef] [Green Version]
- Masalova, O.V.; Lesnova, E.I.; Permyakova, K.Y.; Samokhvalov, E.I.; Ivanov, A.V.; Kochetkov, S.N.; Kushch, A.A. Effect of hepatitis C virus proteins on the production of proinflammatory and profibrotic cytokines in huh7.5 human hepatoma cells. Mol. Biol. (Mosk) 2016, 50, 422–430. [Google Scholar] [CrossRef]
- Francois, M.; Romieu-Mourez, R.; Stock-Martineau, S.; Boivin, M.N.; Bramson, J.L.; Galipeau, J. Mesenchymal stromal cells cross-present soluble exogenous antigens as part of their antigen-presenting cell properties. Blood 2009, 114, 2632–2638. [Google Scholar] [CrossRef] [Green Version]
- Lin, T.; Pajarinen, J.; Kohno, Y.; Huang, J.F.; Maruyama, M.; Romero-Lopez, M.; Nathan, K.; Yao, Z.; Goodman, S.B. Trained murine mesenchymal stem cells have anti-inflammatory effect on macrophages, but defective regulation on T-cell proliferation. FASEB J. 2019, 33, 4203–4211. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Chen, Y.; Li, F.; You, M.; Zhong, L.; Li, W.; Zhang, B.; Chen, Q. The biological changes of umbilical cord mesenchymal stem cells in inflammatory environment induced by different cytokines. Mol. Cell Biochem. 2018, 446, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Hartnell, F.; Brown, A.; Capone, S.; Kopycinski, J.; Bliss, C.; Makvandi-Nejad, S.; Swadling, L.; Ghaffari, E.; Cicconi, P.; Del Sorbo, M.; et al. A novel vaccine strategy employing serologically different chimpanzee adenoviral vectors for the prevention of HIV-1 and hcv coinfection. Front. Immunol. 2018, 9, 3175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, C.; Swadling, L.; Brown, A.; Capone, S.; Folgori, A.; Salio, M.; Klenerman, P.; Barnes, E. Cross-reactivity of hepatitis C virus specific vaccine-induced T cells at immunodominant epitopes. Eur. J. Immunol. 2015, 45, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Callendret, B.; Eccleston, H.B.; Satterfield, W.; Capone, S.; Folgori, A.; Cortese, R.; Nicosia, A.; Walker, C.M. Persistent hepatitis C viral replication despite priming of functional CD8+ T cells by combined therapy with a vaccine and a direct-acting antiviral. Hepatology 2016, 63, 1442–1454. [Google Scholar] [CrossRef] [Green Version]
- Di Bisceglie, A.M.; Janczweska-Kazek, E.; Habersetzer, F.; Mazur, W.; Stanciu, C.; Carreno, V.; Tanasescu, C.; Flisiak, R.; Romero-Gomez, M.; Fich, A.; et al. Efficacy of immunotherapy with tg4040, peg-interferon, and ribavirin in a phase 2 study of patients with chronic HCV infection. Gastroenterology 2014, 147, 119–131.e3. [Google Scholar] [CrossRef]
- Echevarria, D.; Gutfraind, A.; Boodram, B.; Layden, J.; Ozik, J.; Page, K.; Cotler, S.J.; Major, M.; Dahari, H. Modeling indicates efficient vaccine-based interventions for the elimination of hepatitis C virus among persons who inject drugs in metropolitan chicago. Vaccine 2019, 37, 2608–2616. [Google Scholar] [CrossRef]
- Scott, N.; McBryde, E.; Vickerman, P.; Martin, N.K.; Stone, J.; Drummer, H.; Hellard, M. The role of a hepatitis C virus vaccine: Modelling the benefits alongside direct-acting antiviral treatments. BMC Med. 2015, 13, 198. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HCV Proteins | Group 1 mMSC | Group 2 MSC | Group 3 pcNS3-NS5B | Group 4 Control | ||||
|---|---|---|---|---|---|---|---|---|
| IgG1 | IgG2a | IgG1 | IgG2a | IgG1 | IgG2a | IgG1 | IgG2a | |
| NS3 | 200 ± 34 # | 410 ± 27 # | 20 ± 14 | <10 | <10 | 10 | <10 | <10 |
| (4/10) | (10/10) | (2/10) | (0/10) | (0/10) | (0/10) | (0/10) | (0/10) | |
| NS4 | <10 | 1600 ± 24 # | 80 ± 63 | <10 | 90 ± 64 | 40 ± 12 * | <10 | <10 |
| (0/10) | (10/10) | (4/10) | (0/10) | (2/10) | (9/10) | (0/10) | (0/10) | |
| NS5A | <10 | 40 ± 7 # | 10 ± 15 | <10 | <10 | <10 | <10 | <10 |
| (0/10) | (9/10) | (1/10) | (0/10) | (0/10) | (0/10) | (0/10) | (0/10) | |
| NS5B | <10 | 80 ± 12 * | 60 ± 42 | <10 | <10 | 249 ± 61 # | <10 | <10 |
| (0/10) | (10/10) | (4/10) | (0/10) | (0/10) | (10/10) | (0/10) | (0/10) | |
| Assay | Stimulant | Group 1 mMSC | Group 2 MSC | Group 3 pcNS3-NS5B | Group 4 Control |
|---|---|---|---|---|---|
| T-cell proliferation (SI) | ConA | 320 ± 22 | 311 ± 26 | 348 ± 79 | 240 ± 17 |
| T-cell proliferation (c.p.m.) | medium | 164 ± 46 * | 273 ± 84 * | 73 ± 15 | 65 ± 9 |
| ConA | 52406 ± 10014 * | 84933 ± 15010 * | 25432 ± 12031 | 15602 ± 7025 | |
| ELISA, IFN-γ secretion (pg/mL) | medium | 5 ± 2 | 4 ± 2 | 4 ± 2 | 2 ± 2 |
| ConA | 1108 ± 242 | 1336 ± 380 | 954 ± 314 | 902 ± 143 | |
| ELISpot, IFN-γ synthesis (SFC/106 cells) | medium | 23.0 ± 3.1* | 23.5 ± 3.8* | 12.4 ± 2.4 | 10.6 ± 2.2 |
| ConA | 2400 ± 350* | 2250 ± 407* | 1200 ± 210 | 750 ± 156 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masalova, O.V.; Lesnova, E.I.; Klimova, R.R.; Momotyuk, E.D.; Kozlov, V.V.; Ivanova, A.M.; Payushina, O.V.; Butorina, N.N.; Zakirova, N.F.; Narovlyansky, A.N.; et al. Genetically Modified Mouse Mesenchymal Stem Cells Expressing Non-Structural Proteins of Hepatitis C Virus Induce Effective Immune Response. Vaccines 2020, 8, 62. https://doi.org/10.3390/vaccines8010062
Masalova OV, Lesnova EI, Klimova RR, Momotyuk ED, Kozlov VV, Ivanova AM, Payushina OV, Butorina NN, Zakirova NF, Narovlyansky AN, et al. Genetically Modified Mouse Mesenchymal Stem Cells Expressing Non-Structural Proteins of Hepatitis C Virus Induce Effective Immune Response. Vaccines. 2020; 8(1):62. https://doi.org/10.3390/vaccines8010062
Chicago/Turabian StyleMasalova, Olga V., Ekaterina I. Lesnova, Regina R. Klimova, Ekaterina D. Momotyuk, Vyacheslav V. Kozlov, Alla M. Ivanova, Olga V. Payushina, Nina N. Butorina, Natalia F. Zakirova, Alexander N. Narovlyansky, and et al. 2020. "Genetically Modified Mouse Mesenchymal Stem Cells Expressing Non-Structural Proteins of Hepatitis C Virus Induce Effective Immune Response" Vaccines 8, no. 1: 62. https://doi.org/10.3390/vaccines8010062