Mechanisms of Mixed Th1/Th2 Responses in Mice Induced by Albizia julibrissin Saponin Active Fraction by in Silico Analysis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation and Characterization of AJSAF
2.3. Mice
2.4. Immunization
2.5. Measurement of H5Ag-Specific Antibody
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. Microarray Analysis
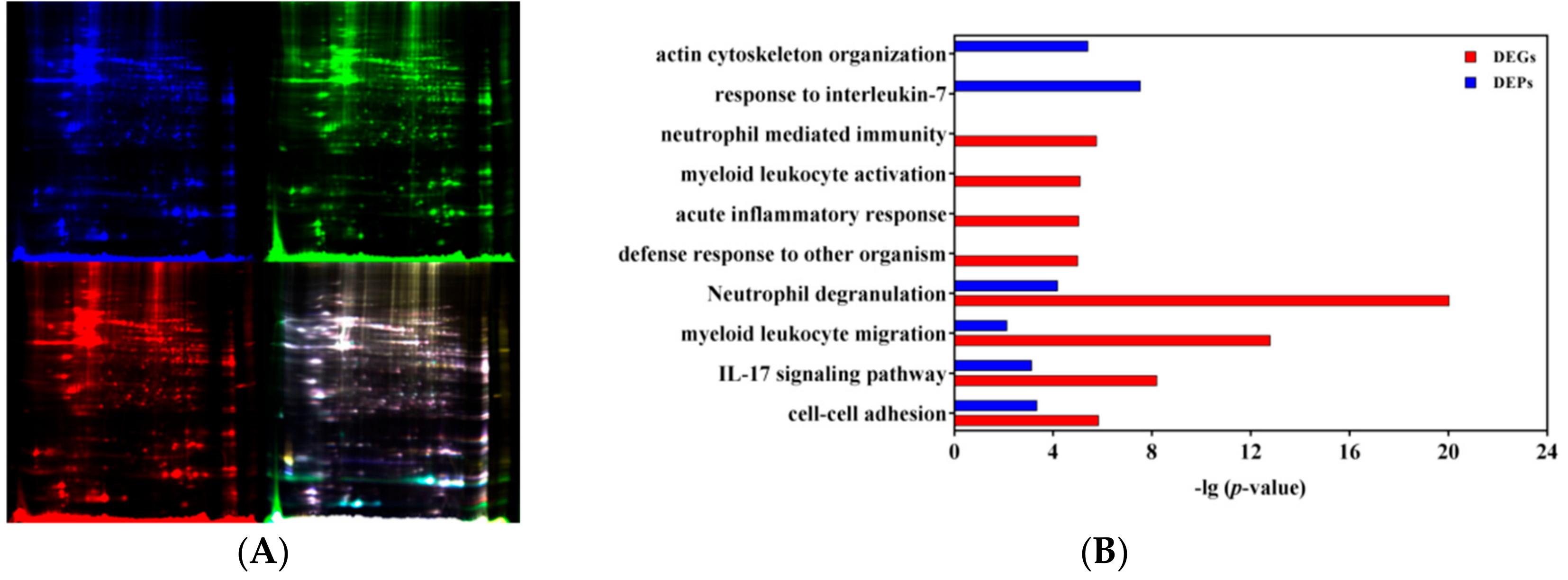
2.8. Two-Dimensional Difference Gel Electrophoresis (2D DIGE)
2.9. Protein Identification by MALDI-TOF-MS
2.10. Statistical Analysis
3. Results
3.1. Comparative Analysis of H5Ag-Specific Serum Antibody Response
3.2. Comparative Analysis of Global Gene Expression
3.3. Pathway and Function Analysis of DEGs
3.4. Integrative Analysis of Transcriptomic and Proteomic Profiles
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Oyston, P.; Robinso, K. The current challenges for vaccine development. J. Med. Microbiol. 2012, 61, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T.R.; Sad, S. The expanding universe of T-cells subsets: Th1, Th2 and more. Immunol. Today 1996, 17, 138–146. [Google Scholar] [CrossRef]
- McKee, A.S.; Munks, M.W.; Marrack, P. How do adjuvants work? Important considerations for new generation adjuvants. Immunity 2007, 27, 687–690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKee, A.S.; Marrack, P. Old and new adjuvants. Curr. Opin. Immunol. 2017, 47, 44–51. [Google Scholar] [CrossRef]
- Reed, S.G.; Bertholet, S.; Coler, R.N.; Friede, M. New horizons in adjuvants for vaccine development. Trends Immunol. 2009, 30, 23–32. [Google Scholar] [CrossRef]
- Kool, M.; Pétrilli, V.; De Smedt, T.; Rolaz, A.; Hammad, H.; van Nimwegen, M.; Bergen, I.M.; Castillo, R.; Lambrecht, B.N.; Tschopp, J. Cutting edge: alum adjuvant stimulates inflammatory dendritic cells through activation of the NALP3 inflammasome. J. Immunol. 2008, 181, 3755–3759. [Google Scholar] [CrossRef] [Green Version]
- Eisenbarth, S.C.; Colegio, O.R.; O’Connor, W.; Sutterwala, F.S.; Flavell, R.A. Crucial role for the Nalp3 inflammasome in the immunostimulatory properties of aluminium adjuvants. Nature 2008, 453, 1122–1126. [Google Scholar] [CrossRef]
- Li, H.; Willingham, S.B.; Ting, J.P.Y.; Re, F. Cutting edge: inflammasome activation by alum and alum’s adjuvant effect are mediated by NLRP3. J. Immunol. 2008, 181, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Kool, M.; Soullié, T.; van Nimwegen, M.; Willart, M.A.; Muskens, F.; Jung, S.; Hoogsteden, H.C.; Hammad, H.; Lambrecht, B.N. Alum adjuvant boosts adaptive immunity by inducing uric acid and activating inflammatory dendritic cells. J. Exp. Med. 2008, 205, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Marichal, T.; Ohata, K.; Bedoret, D.; Mesnil, C.; Sabatel, C.; Kobiyama, K.; Lekeux, P.; Coban, C.; Akira, S.; Ishii, K.J.; et al. DNA released from dying host cells mediates aluminum adjuvant activity. Nat. Med. 2011, 17, 996–1002. [Google Scholar] [CrossRef]
- McKee, A.S.; Burchill, M.A.; Munks, M.W.; Jin, L.; Kappler, J.W.; Friedman, R.S.; Jacobelli, J.; Marrack, P. Host DNA released in response to aluminum adjuvant enhances MHC class II-mediated antigen presentation and prolongs CD4 T-cell interactions with dendritic cells. Proc. Natl. Acad. Sci. USA 2013, 110, E1122–E1131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Franchi, L.; Núñez, G. The Nlrp3 inflammasome is critical for aluminium hydroxide-mediated IL-1β secretion but dispensable for adjuvant activity. Eur. J. Immunol. 2008, 38, 2085–2089. [Google Scholar] [CrossRef] [PubMed]
- McKee, A.S.; Munks, M.W.; MacLeod, M.K.; Fleenor, C.J.; Van Rooijen, N.; Kappler, J.W.; Marrack, P. Alum induces innate immune responses through macrophage and mast cell sensors, but these sensors are not required for alum to act as an adjuvant for specific immunity. J. Immunol. 2009, 183, 4403–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, H.X.; He, S.W.; Shi, M.H. Adjuvant-active fraction from Albizia julibrissin saponins improves immune responses by inducing cytokine and chemokine at the site of injection. Int. Immunopharmacol. 2014, 22, 346–355. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.N.; He, T.Y.; Gao, X.Y.; Shi, M.H.; Sun, H.X. Evaluation and characteristics of immunological adjuvant activity of purified fraction of Albizia julibrissin saponins. Immunol. Invest. 2019, 48, 283–302. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing α-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef]
- He, Y.F.; Liu, Z.Y.; Ye, Y.P.; Sun, H.X. Rapid annotation and structural characterization of saponins in the active fraction of Albizia julibrissin by HPLC coupled with quadrupole time-of-flight mass spectrometry based on accurate mass database. J. Sep. Sci. 2019, 42, 2922–2941. [Google Scholar] [CrossRef]
- Xie, Y.; Sun, H.X.; Li, D. Platycodin D improves the immunogenicity of Newcastle disease virus-based recombinant avian influenza vaccine in mice. Chem. Biodiv. 2010, 7, 677–689. [Google Scholar] [CrossRef]
- Du, J.; Chen, X.F.; Wang, C.Y.; Sun, H.X. Pathway analysis of global gene expression change in dendritic cells induced by the polysaccharide from the roots of Actinidia eriantha. J. Ethnopharmacol. 2018, 214, 141–152. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: a knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Breuer, K.; Foroushani, A.K.; Laird, M.R.; Chen, C.; Sribnaia, A.; Lo, R.; Winsor, G.L.; Hancock, R.E.; Brinkman, F.S.; Lynn, D.J. InnateDB: systems biology of innate immunity and beyond—recent updates and continuing curation. Nucleic Acids Res. 2012, 41, D1228–D1233. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Szabo, S.J.; Kim, S.T.; Costa, G.L.; Zhang, X.; Fathman, C.G.; Glimcher, L.H. A novel transcription factor, T-bet, directs Th1 lineage commitment. Cell 2000, 100, 655–669. [Google Scholar] [CrossRef] [Green Version]
- Jang, E.J.; Park, H.R.; Hong, J.H.; Hwang, E.S. Lysine 313 of T-box is crucial for modulation of protein stability, DNA binding, and threonine phosphorylation of T-bet. J. Immunol. 2013, 190, 5764–5770. [Google Scholar] [CrossRef] [Green Version]
- Lazarevic, V.; Chen, X.; Shim, J.H.; Hwang, E.S.; Jang, E.; Bolm, A.N.; Oukka, M.; Kuchroo, V.K.; Glimcher, L.H. T-bet represses T(H)17 differentiation by preventing Runx1-mediated activation of the gene encoding RORγt. Nat. Immunol. 2011, 12, 96–104. [Google Scholar] [CrossRef] [Green Version]
- De Sousa Abreu, R.; Penalva, L.O.; Marcotte, E.M.; Vogel, C. Global signatures of protein and mRNA expression levels. Mol. Biosyst. 2009, 5, 1512–1526. [Google Scholar] [CrossRef] [Green Version]
- Seyfried, N.T.; Dammer, E.B.; Swarup, V.; Nandakumar, D.; Duong, D.M.; Yin, L.; Deng, Q.; Nguyen, T.; Hales, C.M.; Wingo, T.; et al. A multi-network approach identifies protein-specific co-expression in asymptomatic and symptomatic Alzheimer’s disease. Cell Syst. 2017, 4, 60–72. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Rouphael, N.; Duraisingham, S.; Romero-Steiner, S.; Presnell, S.; Davis, C.; Schmidt, D.S.; Johnson, S.E.; Milton, A.; Rajam, G.; et al. Molecular signatures of antibody responses derived from a systems biology study of five human vaccines. Nat. Immunol. 2014, 15, 195–204. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Y.; Du, J.; Chen, X.F.; Zhu, Y.L.; Sun, H.X. Activation of RAW264. 7 macrophages by active fraction of Albizia julibrissin saponin via Ca2+–ERK1/2–CREB–lncRNA pathways. Int. Immunopharmacol. 2019, 77, 105955. [Google Scholar] [CrossRef]
- Wang, S.; Song, R.; Wang, Z.; Jing, Z.; Wang, S.; Ma, J. S100A8/A9 in Inflammation. Front. Immunol. 2018, 9, 1298. [Google Scholar] [CrossRef] [PubMed]
- Petersen, B.; Wolf, M.; Austermann, J.; van Lent, P.; Foell, D.; Ahlmann, M.; Kupas, V.; Loser, K.; Sorg, C.; Roth, J.; et al. The alarmin Mrp8/14 as regulator of the adaptive immune response during allergic contact dermatitis. EMBO J. 2013, 32, 100–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loser, K.; Vogl, T.; Voskort, M.; Lueken, A.; Kupas, V.; Nacken, W.; Klenner, L.; Kuhn, A.; Foell, D.; Sorokin, L.; et al. The Toll-like receptor 4 ligands Mrp8 and Mrp14 are crucial in the development of autoreactive CD8+ T cells. Nat. Med. 2010, 16, 713–717. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.; Han, Z.; Oppenheim, J.J. Alarmins and immunity. Immunol. Rev. 2017, 280, 41–56. [Google Scholar] [CrossRef]
- Oppenheim, J.J.; Yang, D. Alarmins: Chemotactic activators of immune responses. Curr. Opin. Immunol. 2005, 17, 359–365. [Google Scholar] [CrossRef]
- Wang, Y.; Rahman, D.; Lehner, T. A comparative study of stress-mediated immunological functions with the adjuvanticity of alum. J. Biol. Chem. 2012, 287, 17152–17160. [Google Scholar] [CrossRef] [Green Version]
- Rovere-Querini, P.; Capobianco, A.; Scaffidi, P.; Valentinis, B.; Catalanotti, F.; Giazzon, M.; Dumitriu, I.E.; Müller, S.; Iannacone, M.; Traversari, C.; et al. HMGB1 is an endogenous immune adjuvant released by necrotic cells. EMBO Rep. 2004, 5, 825–830. [Google Scholar] [CrossRef]
- Sugimoto, M.A.; Vago, J.P.; Teixeira, M.M.; Sousa, L.P. Annexin A1 and the resolution of inflammation: Modulation of neutrophil recruitment, apoptosis, and clearance. J. Immunol. Res. 2016, 2016, 8239258. [Google Scholar] [CrossRef] [Green Version]
- Gavins, F.N.; Yona, S.; Kamal, A.M.; Flower, R.J.; Perretti, M. Leukocyte antiadhesive actions of annexin 1: ALXR-and FPR-related anti-inflammatory mechanisms. Blood 2003, 101, 4140–4147. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Cai, L.; Wang, H.; Wu, P.; Gu, W.; Chen, Y.; Hao, H.; Tang, K.; Yi, P.; Liu, M.; et al. Pleiotropic regulation of macrophage polarization and tumorigenesis by formyl peptide receptor-2. Oncogene 2011, 30, 3887–3899. [Google Scholar] [CrossRef] [Green Version]
- Cooray, S.N.; Gobbetti, T.; Montero-Melendez, T.; McArthur, S.; Thompson, D.; Clark, A.J.; Flower, R.J.; Perretti, M. Ligand-specific conformational change of the G-protein–coupled receptor ALX/FPR2 determines proresolving functional responses. Proc. Natl. Acad. Sci. USA 2013, 110, 18232–18237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooley, J.; Takayama, T.K.; Shapiro, S.D.; Schechter, N.M.; Remold-O’Donnell, E. The serpin MNEI inhibits elastase-like and chymotrypsin-like serine proteases through efficient reactions at two active sites. Biochemistry 2001, 40, 15762–15770. [Google Scholar] [CrossRef] [PubMed]
- Fiorentino, D.F.; Zlotnik, A.; Vieira, P.; Mosmann, T.R.; Howard, M.; Moore, K.W.; O’Garra, A. IL-10 acts on the antigen-presenting cell to inhibit cytokine production by Th1 cells. J. Immunol. 1991, 146, 3444–3451. [Google Scholar] [PubMed]
- Khan, A.Q.; Shen, Y.; Wu, Z.Q.; Wynn, T.A.; Snapper, C.M. Endogenous pro-and anti-inflammatory cytokines differentially regulate an in vivo humoral response to Streptococcus pneumoniae. Infect. Immun. 2002, 70, 749–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Coffman, R.L.; Sher, A.; Seder, R.A. Vaccine adjuvants: putting innate immunity to work. Immunity 2010, 33, 492–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burns, S.O.; Zarafov, A.; Thrasher, A.J. Primary immunodeficiencies due to abnormalities of the actin cytoskeleton. Curr. Opin. Hematol. 2017, 24, 16–22. [Google Scholar] [CrossRef]
- Feldt, J.; Schicht, M.; Garreis, F.; Welss, J.; Schneider, U.W.; Paulsen, F. Structure, regulation and related diseases of the actin-binding protein gelsolin. Expert Rev. Mol. Med. 2019, 20, e7. [Google Scholar] [CrossRef]
- Arora, P.D.; Glogauer, M.; Kapus, A.; Kwiatkowski, D.J.; McCulloch, C.A. Gelsolin mediates collagen phagocytosis through a rac-dependent step. Mol. Biol. Cell 2004, 15, 588–599. [Google Scholar] [CrossRef] [Green Version]
- Gonçalves, A.F.; Dias, N.G.; Moransard, M.; Correia, R.; Pereira, J.A.; Witke, W.; Suter, U.; Relvas, J.B. Gelsolin is required for macrophage recruitment during remyelination of the peripheral nervous system. Glia 2010, 58, 706–715. [Google Scholar]
- Langereis, J.D. Neutrophil integrin affinity regulation in adhesion, migration, and bacterial clearance. Cell Adh. Migr. 2013, 7, 486–491. [Google Scholar] [CrossRef] [Green Version]
- Provost, P.; Doucet, J.; Stock, A.; Gerisch, G.; Samuelsson, B.; Rådmark, O. Coactosin-like protein, a human F-actin-binding protein: critical role of lysine-75. Biochem. J. 2001, 359, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Shapiro, M.J.; Bamidele, A.O.; Gurel, P.; Thapa, P.; Higgs, H.N.; Hedin, K.E.; Shapiro, V.S.; Billadeau, D.D. Coactosin-like 1 antagonizes cofilin to promote lamellipodial protrusion at the immune synapse. PLoS ONE 2014, 9, e85090. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.T.; Creed, S.J.; Bear, J.E. Unraveling the enigma: progress towards understanding the coronin family of actin regulators. Trends Cell Biol. 2011, 21, 481–488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yokoyama, K.; Kaji, H.; He, J.; Tanaka, C.; Hazama, R.; Kamigaki, T.; Ku, Y.; Tohyama, K.; Tohyama, Y. Rab27a negatively regulates phagocytosis by prolongation of the actin-coating stage around phagosomes. J. Biol. Chem. 2011, 286, 5375–5382. [Google Scholar] [CrossRef] [Green Version]
- Föger, N.; Rangell, L.; Danilenko, D.M.; Chan, A.C. Requirement for coronin 1 in T lymphocyte trafficking and cellular homeostasis. Science 2006, 313, 839–842. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Harada, T.; Juang, Y.T.; Kyttaris, V.C.; Wang, Y.; Zidanic, M.; Tung, K.; Tsokos, G.C. Phosphorylated ERM is responsible for increased T cell polarization, adhesion, and migration in patients with systemic lupus erythematosus. J. Immunol. 2007, 178, 1938–1947. [Google Scholar] [CrossRef] [Green Version]
- Cullinan, P.; Sperling, A.I.; Burkhardt, J.K. The distal pole complex: A novel membrane domain distal to the immunological synapse. Immunol. Rev. 2002, 189, 111–122. [Google Scholar] [CrossRef]
- Pore, D.; Parameswaran, N.; Matsui, K.; Stone, M.B.; Saotome, I.; McClatchey, A.I.; Veatch, S.L.; Gupta, N. Ezrin tunes the magnitude of humoral immunity. J. Immunol. 2013, 191, 4048–4058. [Google Scholar] [CrossRef] [Green Version]
- Pore, D.; Matsui, K.; Parameswaran, N.; Gupta, N. Cutting edge: ezrin regulates inflammation by limiting B cell IL-10 production. J. Immunol. 2016, 196, 558–562. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spot No. | p-Value | Up/Down | Abbr | Accession No. | ID | MW | PI | Pep Count | Protein Score | Protein/Ion Credibility | Best Ion Score |
|---|---|---|---|---|---|---|---|---|---|---|---|
| L21 | 0.0031 | 3.38 | NGP | IPI00127280 | 18054 | 19661.7 | 5.21 | 18 | 731 | 100/100 | 518 |
| L24 | 0.00054 | 3.91 | S100A8 | IPI00230768 | 20201 | 10345.1 | 5.43 | 5 | 206 | 100/100 | 154 |
| M1 | 0.0085 | 2.67 | 10345.1 | 5.43 | 3 | 279 | 100/100 | 252 | |||
| M23 | 0.0017 | 3.05 | 10345.1 | 5.43 | 5 | 322 | 100/100 | 270 | |||
| M24 | 0.017 | 3.02 | S100A9 | IPI00222556 | 20202 | 13211.3 | 6.64 | 9 | 383 | 100/100 | 311 |
| N2 | 0.034 | 2.34 | 13211.3 | 6.64 | 10 | 426 | 100/100 | 339 | |||
| L15 | 0.0032 | 2.31 | ANXA1 | IPI00230395 | 16952 | 38995.1 | 6.97 | 24 | 826 | 100/100 | 606 |
| M11 | 0.019 | 1.92 | EFHD2 | IPI00112223 | 27984 | 26774.6 | 5.01 | 17 | 438 | 100/100 | 294 |
| N8 | 0.00095 | 1.91 | GM9234 | IPI00987580 | 668548 | 18173.8 | 6.2 | 7 | 414 | 100/100 | 356 |
| N9 | 0.00075 | 1.76 | 18173.8 | 6.2 | 8 | 526 | 100/100 | 456 | |||
| L23 | 0.0096 | 1.8 | CAMP | IPI00875797 | 12796 | 19797.3 | 8.9 | 3 | 159 | 100/100 | 142 |
| M16 | 0.034 | 1.67 | ACTG1 | IPI01027491 | 11465 | 32941.3 | 5.15 | 6 | 107 | 100/100 | 75 |
| M15 | 0.049 | 1.66 | FTH1 | IPI00230145 | 14319 | 21224.3 | 5.53 | 7 | 236 | 100/100 | 187 |
| M6 | 0.037 | 1.63 | ALDH2 | IPI00111218 | 11669 | 57014.9 | 7.53 | 23 | 849 | 100/100 | 647 |
| L14 | 0.013 | 1.56 | CAPZA2 | IPI00111265 | 12343 | 33117.7 | 5.57 | 11 | 347 | 100/100 | 260 |
| M8 | 0.05 | 1.39 | SERPINB1A | IPI00457659 | 66222 | 42718.8 | 5.85 | 22 | 1,130 | 100/100 | 928 |
| L17 | 0.0012 | 1.38 | PNP | IPI00607023 | 18950 | 32527.2 | 5.78 | 16 | 680 | 100/100 | 513 |
| N20 | 0.0053 | 1.28 | SRSF1 | IPI00420807 | 110809 | 27841.9 | 10.37 | 9 | 82 | 99.96/93.89 | 35 |
| M10 | 0.0066 | 1.27 | GSN | IPI00759948 | 227753 | 80997.5 | 5.52 | 7 | 113 | 100/100 | 94 |
| N19 | 0.042 | 1.25 | ECH1 | IPI00130804 | 51798 | 36437.3 | 7.6 | 14 | 603 | 100/100 | 493 |
| M5 | 0.02 | 1.24 | P4HB | IPI00133522 | 18453 | 57421.9 | 4.77 | 31 | 905 | 100/100 | 595 |
| N16 | 0.044 | 1.2 | PGAM1 | IPI00457898 | 18648 | 28927.9 | 6.67 | 15 | 812 | 100/100 | 666 |
| O3 | 0.022 | 1.25 | 28927.9 | 6.67 | 17 | 819 | 100/100 | 643 | |||
| N14 | 0.0042 | −2.12 | BLVRB | IPI00113996 | 233016 | 22297.4 | 6.49 | 8 | 321 | 100/100 | 255 |
| N13 | 0.0079 | −1.83 | IGK-C | IPI00850020 | 16071 | 24434.9 | 7.05 | 8 | 256 | 100/100 | 192 |
| M4 | 0.03 | −1.55 | CORO1A | IPI00323600 | 12721 | 51641.2 | 6.05 | 15 | 447 | 100/100 | 339 |
| M2 | 0.0017 | −1.54 | RDX | IPI00308324 | 19684 | 68614.4 | 5.91 | 15 | 247 | 100/100 | 183 |
| M20 | 0.024 | −1.48 | SNRPF | IPI00943994 | 69878 | 9775.8 | 4.7 | 4 | 110 | 100/100 | 76 |
| L11 | 0.0027 | −1.45 | EZR | IPI00330862 | 22350 | 69477.7 | 5.83 | 10 | 217 | 100/100 | 186 |
| N12 | 0.019 | −1.38 | GUK1 | IPI00986878 | 14923 | 22018.3 | 6.12 | 5 | 168 | 100/100 | 136 |
| L22 | 0.0036 | −1.38 | RGS10 | IPI00132450 | 67865 | 21194.6 | 6.36 | 15 | 500 | 100/100 | 347 |
| N15 | 0.0065 | −1.36 | TPI1 | IPI00988063 | 21991 | 27037.9 | 6.9 | 15 | 643 | 100/100 | 494 |
| M7 | 0.0051 | −1.33 | ENO1 | IPI00462072 | 13806 | 47453.3 | 6.37 | 21 | 628 | 100/100 | 431 |
| N6 | 0.0013 | −1.38 | PPIA | IPI00554989 | 268373 | 18473.1 | 7.74 | 9 | 336 | 100/100 | 255 |
| N7 | 0.0013 | −1.27 | 18473.1 | 7.74 | 9 | 243 | 100/100 | 165 | |||
| N18 | 0.0099 | −1.3 | EIF4H | IPI00124742 | 22384 | 27381.4 | 6.67 | 14 | 350 | 100/100 | 240 |
| N3 | 0.0012 | −1.28 | SOD1 | IPI00130589 | 20655 | 16103.9 | 6.02 | 13 | 645 | 100/100 | 500 |
| N4 | 0.028 | −1.3 | 16103.9 | 6.02 | 8 | 164 | 100/100 | 93 | |||
| N11 | 0.0063 | −1.28 | GPX1 | IPI00319652 | 14775 | 22553.4 | 6.74 | 7 | 457 | 100/100 | 404 |
| M17 | 0.027 | −1.28 | COTL1 | IPI00132575 | 72042 | 16048 | 5.28 | 9 | 318 | 100/100 | 244 |
| M12 | 0.013 | −1.28 | PRDX2 | IPI00117910 | 21672 | 21936.1 | 5.2 | 9 | 593 | 100/100 | 515 |
| M14 | 0.029 | −1.26 | PARK7 | IPI00117264 | 57320 | 20236.5 | 6.32 | 9 | 334 | 100/100 | 261 |
| M3 | 0.011 | −1.25 | PDIA3 | IPI00230108 | 14827 | 57098.9 | 5.88 | 28 | 692 | 100/100 | 426 |
| N17 | 0.052 | −1.24 | HMGB1 | IPI00420261 | 15289 | 25049.2 | 5.62 | 11 | 220 | 100/100 | 144 |
| M19 | 0.016 | −1.24 | SH3BGRL3 | IPI00127358 | 73723 | 10527.3 | 5.02 | 6 | 274 | 100/100 | 214 |
| M13 | 0.00047 | −1.23 | CMPK1 | IPI00331146 | 66588 | 26040.4 | 8.13 | 14 | 511 | 100/100 | 394 |
| N5 | 0.032 | −1.22 | RBM3 | IPI00130883 | 19652 | 16594.7 | 6.84 | 6 | 158 | 100/100 | 114 |
| N10 | 0.021 | −1.2 | ARPC5L | IPI00111117 | 74192 | 17026.8 | 6.32 | 7 | 384 | 100/100 | 326 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Du, J.; Jin, J.; Wang, J.; Sun, H. Mechanisms of Mixed Th1/Th2 Responses in Mice Induced by Albizia julibrissin Saponin Active Fraction by in Silico Analysis. Vaccines 2020, 8, 48. https://doi.org/10.3390/vaccines8010048
Du J, Jin J, Wang J, Sun H. Mechanisms of Mixed Th1/Th2 Responses in Mice Induced by Albizia julibrissin Saponin Active Fraction by in Silico Analysis. Vaccines. 2020; 8(1):48. https://doi.org/10.3390/vaccines8010048
Chicago/Turabian StyleDu, Jing, Junjie Jin, Juanjuan Wang, and Hongxiang Sun. 2020. "Mechanisms of Mixed Th1/Th2 Responses in Mice Induced by Albizia julibrissin Saponin Active Fraction by in Silico Analysis" Vaccines 8, no. 1: 48. https://doi.org/10.3390/vaccines8010048