An Antibody Persistent and Protective Two rSsCLP-Based Subunit Cocktail Vaccine against Sarcoptes scabiei in a Rabbit Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Approval and Consent to Participate
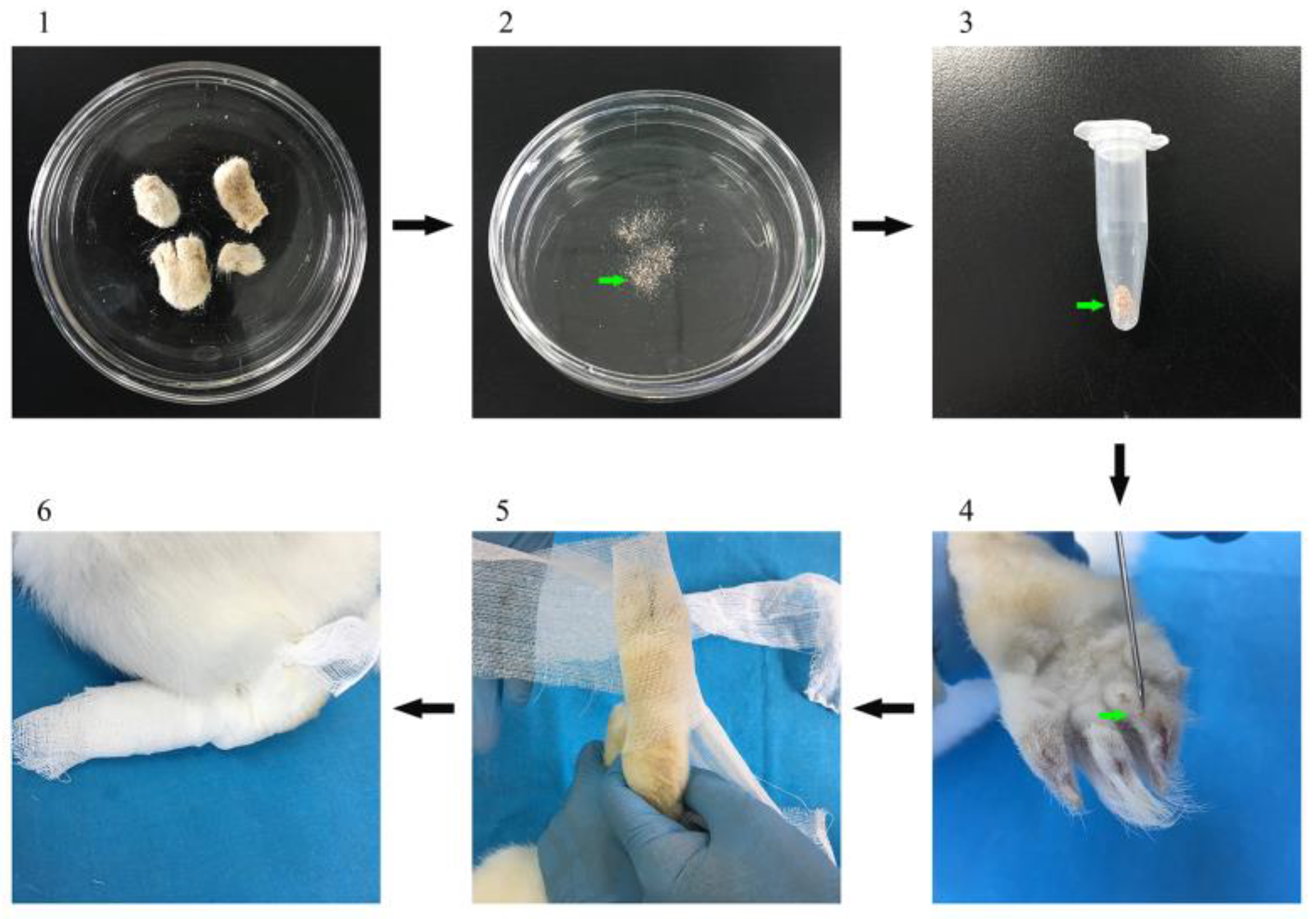
2.2. Source of Mites and Animals
2.3. Preparation of Proteins for Vaccination
2.4. Optimization of a Vaccination Schedule
2.5. Vaccination Trial and Mite Challenge
2.6. Clinical Monitoring and Mite Burden
2.7. Measurement of Antibody and Cytokine Responses
2.8. Histology of Skin Lesions
2.9. Statistical Analyses
3. Results
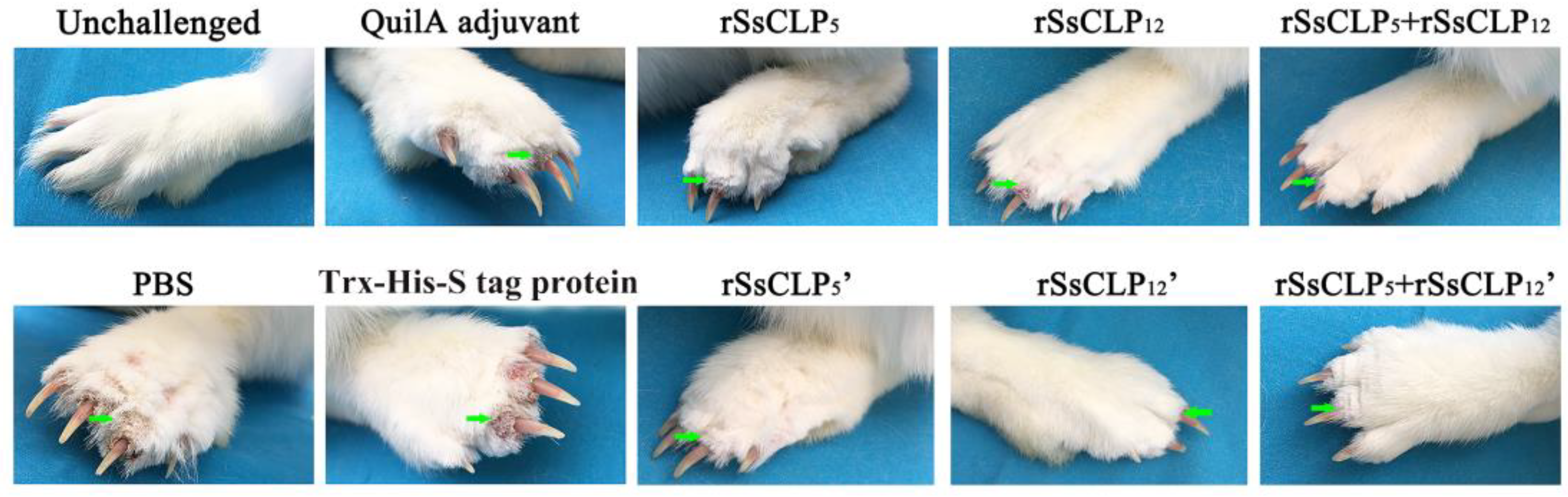
3.1. Vaccination Trial and Clinical Monitoring
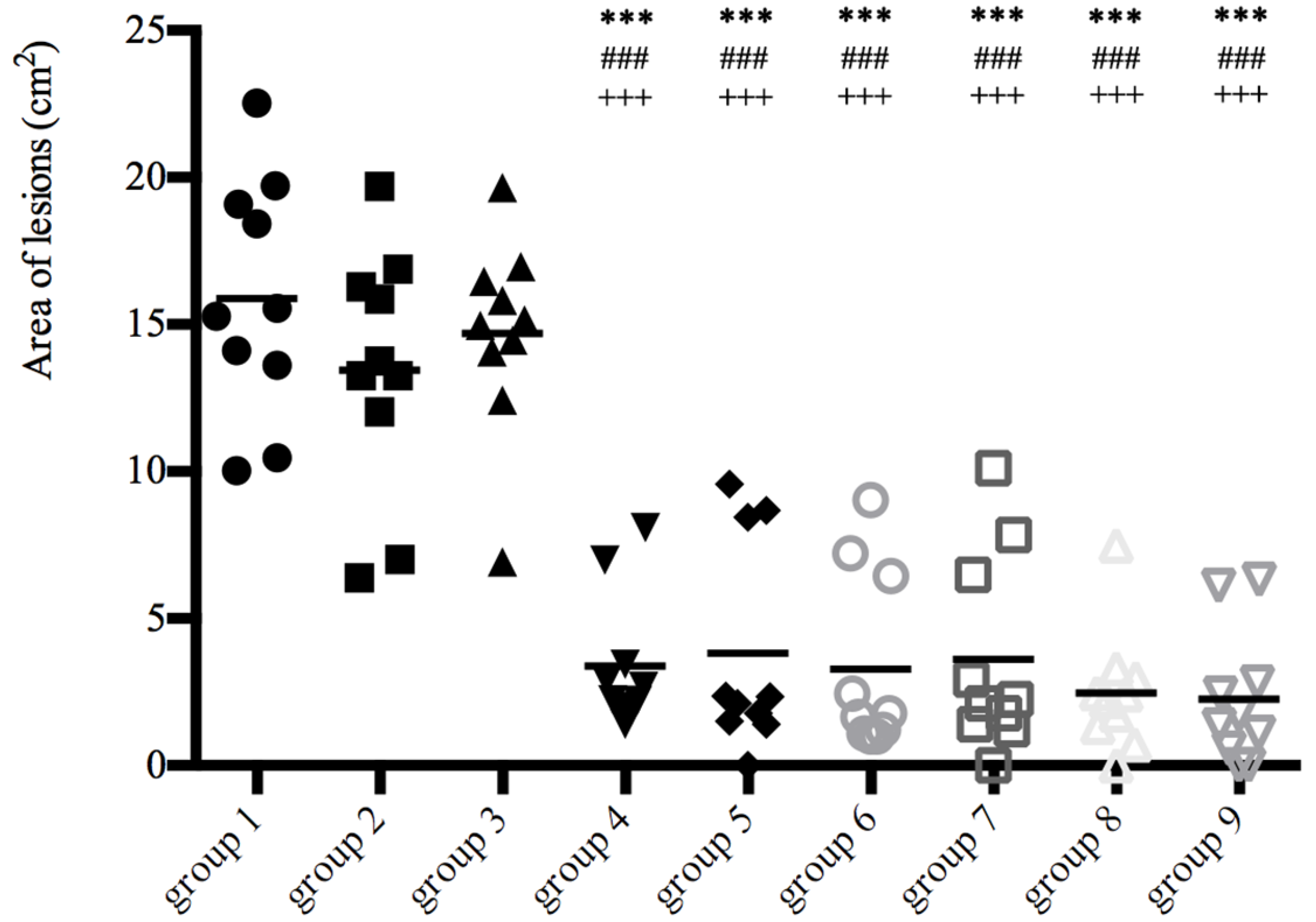
3.2. Lesion Size and Mite Burden
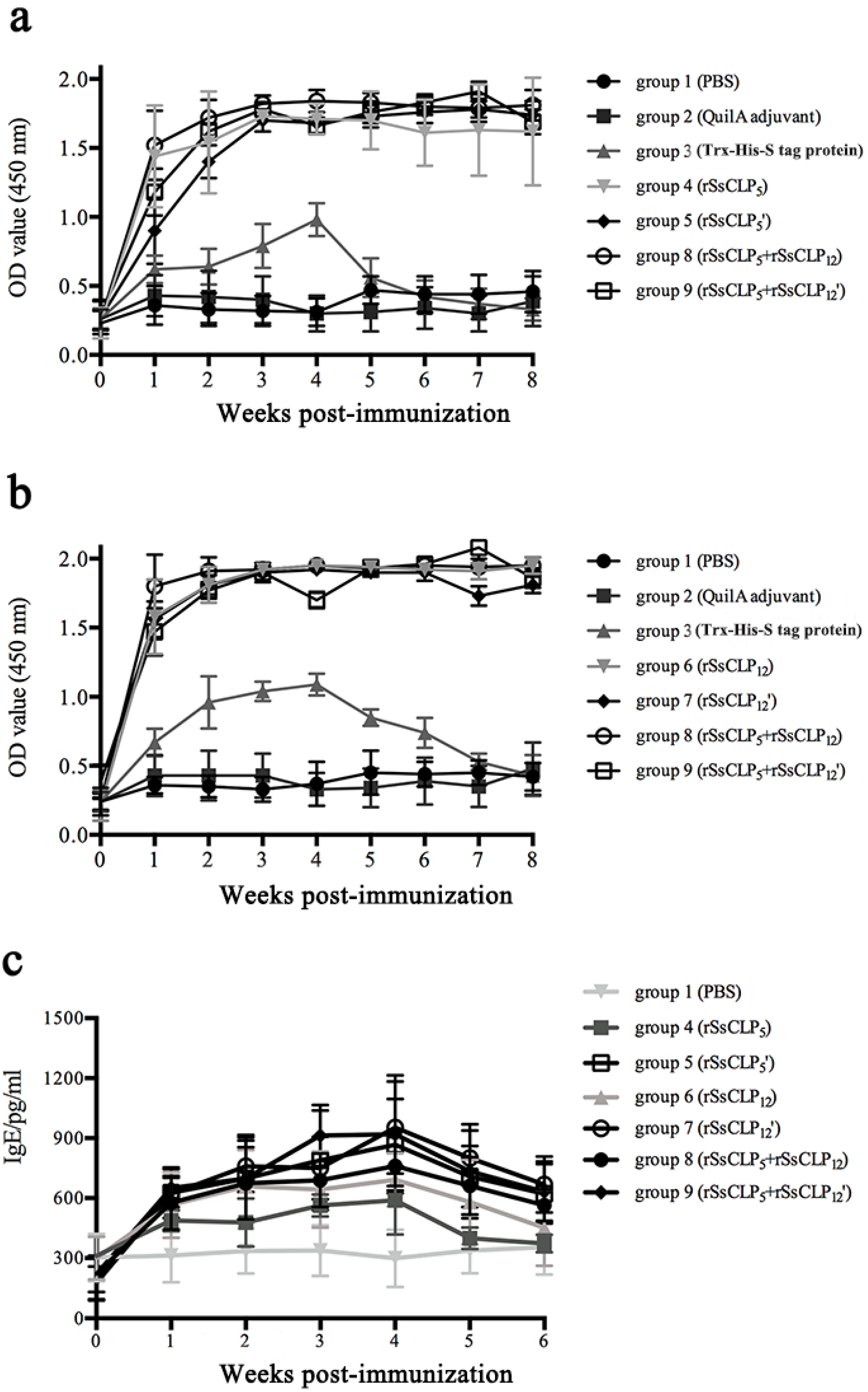
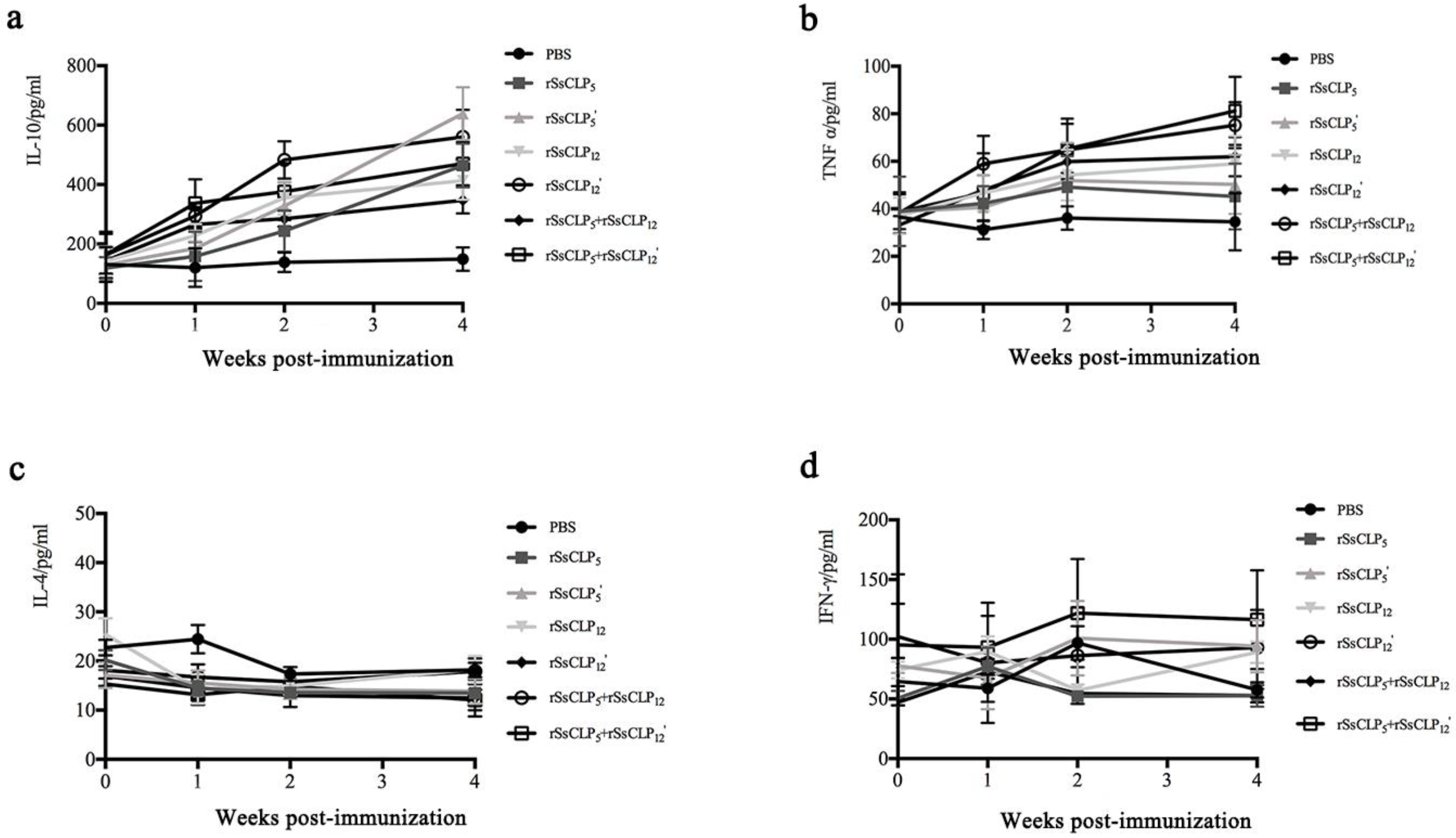
3.3. Antibody and Cytokine Responses
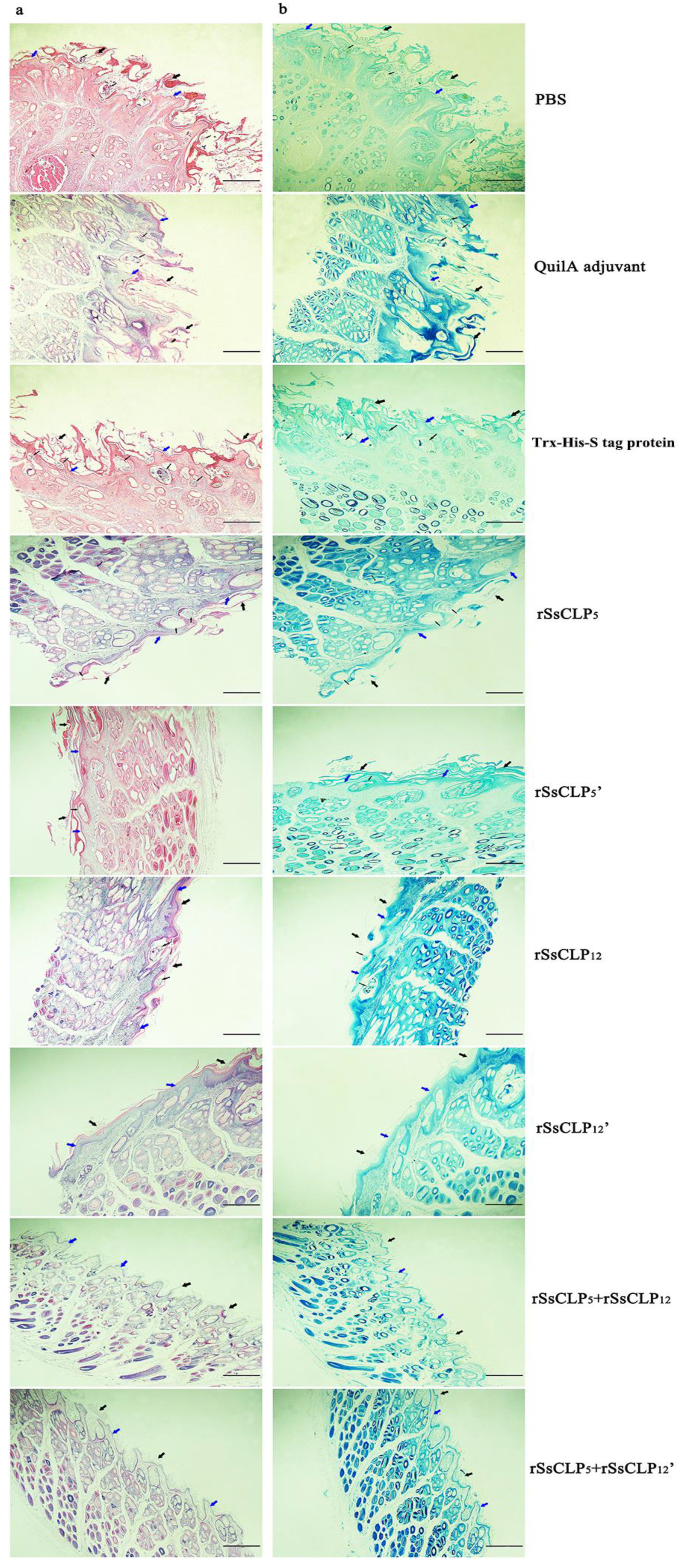
3.4. Histopathology
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Burkhart, C.G.; Burkhart, C.N.; Burkhart, K.M. An epidemiologic and therapeutic reassessment of scabies. Cutis 2000, 65, 233–240. [Google Scholar] [PubMed]
- Dressler, C.; Rosumeck, S.; Sunderkötter, C.; Werner, R.N.; Nast, A. The Treatment of Scabies. Dtsch. Arztebl. Int. 2016, 113, 757–762. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelman, D.; Fuller, L.C.; Steer, A.C.; International Alliance for the Control of Scabies Delphi panel. Consensus criteria for the diagnosis of scabies: A Delphi study of international experts. PLoS Negl. Trop. Dis. 2018, 12, e0006549. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, S.; Mörner, T.; Samuel, W.M. Sarcoptes Scabiei and Sarcoptic Mange; Iowa State University Press: Ames, IA, USA, 2008; pp. 107–119. [Google Scholar]
- Hay, R.J.; Johns, N.E.; Williams, H.C.; Bolliger, I.W.; Dellavalle, R.P.; Margolis, D.J.; Marks, R.; Naldi, L.; Weinstock, M.A.; Wulf, S.K. The global burden of skin disease in 2010: An analysis of the prevalence and impact of skin conditions. J. Investig. Dermatol. 2014, 134, 1527–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terry, B.C.; Kanjah, F.; Sahr, F.; Kortequee, S.; Dukulay, I.; Gbakima, A.A. Sarcoptes scabiei infestation among children in a displacement camp in Sierra Leone. Public Health 2001, 115, 208–211. [Google Scholar] [CrossRef]
- Heukelbach, J.; Feldmeier, H. Scabies. Lancet 2006, 367, 1767–1774. [Google Scholar] [CrossRef]
- Romani, L.; Steer, A.C.; Whitfeld, M.J.; Kaldor, J.M. Prevalence of scabies and impetigo worldwide: A systematic review. Lancet Infect. Dis. 2015, 15, 960–967. [Google Scholar] [CrossRef]
- Andrews, R.M.; Mccarthy, J.; Carapetis, J.R.; Currie, B.J. Skin disorders, including pyoderma, scabies, and tinea infections. Pediatric Clin. N. Am. 2009, 56, 1421–1440. [Google Scholar] [CrossRef]
- Hoy, W.E.; White, A.V.; Dowling, A.; Sharma, S.K.; Bloomfield, H.; Tipiloura, B.T.; Swanson, C.E.; Mathews, J.D.; Mccredie, D.A. Post-streptococcal glomerulonephritis is a strong risk factor for chronic kidney disease in later life. Kidney Int. 2012, 81, 1026–1032. [Google Scholar] [CrossRef] [Green Version]
- Walton, S.F.; Holt, D.C.; Currie, B.J.; Kemp, D.J. Scabies: New Future for a Neglected Disease. Adv. Parasitol. 2004, 57, 309–376. [Google Scholar] [CrossRef]
- Romani, L.; Marks, M.; Sokana, O.; Nasi, T.; Kamoriki, B.; Cordell, B.; Wand, H.; Whitfeld, M.J.; Engelman, D.; Solomon, A.W.; et al. Efficacy of mass drug administration with ivermectin for control of scabies and impetigo, with coadministration of azithromycin: A single-arm community intervention trial. Lancet Infect. Dis. 2019, 19, 510–518. [Google Scholar] [CrossRef] [Green Version]
- Marks, M.; Toloka, H.; Baker, C.; Kositz, C.; Asugeni, J.; Puiahi, E.; Asugeni, R.; Azzopardi, K.; Diau, J.; Kaldor, J.M.; et al. Randomized Trial of Community Treatment With Azithromycin and Ivermectin Mass Drug Administration for Control of Scabies and Impetigo. Clin. Infect. Dis. 2019, 68, 927–933. [Google Scholar] [CrossRef] [PubMed]
- Currie, B.J.; Harumal, P.; Mckinnon, M.; Walton, S.F. First Documentation of in vivo and in vitro Ivermectin Resistance in Sarcoptes scabiei [Abstract]. Clin. Infect. Dis. 2004, 39, 154. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terada, Y.; Murayama, N.; Ikemura, H.; Morita, T.; Nagata, M. Sarcoptes scabiei var. canis refractory to ivermectin treatment in two dogs. Vet. Dermatol. 2010, 21, 608–612. [Google Scholar] [CrossRef]
- Zhang, R.; Jise, Q.; Zheng, W.; Ren, Y.; Xiang, N.; Wu, X.; Gu, X.; Wang, S.; Peng, X.; Lai, S. Characterization and evaluation of a Sarcoptes scabiei allergen as a candidate vaccine. Parasites Vectors 2012, 5, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casais, R.; Granda, V.; Balseiro, A.; Del Cerro, A.; Dalton, K.P.; Gonzalez, R.; Bravo, P.; Prieto, J.M.; Montoya, M. Vaccination of rabbits with immunodominant antigens from Sarcoptes scabiei induced high levels of humoral responses and pro-inflammatory cytokines but confers limited protection. Parasit Vectors 2016, 9, 435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhat, S.A.; Mounsey, K.E.; Liu, X.; Walton, S.F. Host immune responses to the itch mite, Sarcoptes scabiei, in humans. Parasites Vectors 2017, 10, 385. [Google Scholar] [CrossRef] [Green Version]
- Rider, S.D., Jr.; Morgan, M.S.; Arlian, L.G. Draft genome of the scabies mite. Parasit Vectors 2015, 8, 585. [Google Scholar] [CrossRef] [Green Version]
- Morgan, M.S.; Arlian, L.G.; Rider, S.D., Jr.; Grunwald, W.C., Jr.; Cool, D.R. A Proteomic Analysis of Sarcoptes scabiei (Acari: Sarcoptidae). J. Med. Entomol. 2016, 53, 553–561. [Google Scholar] [CrossRef]
- He, R.; Gu, X.; Lai, W.; Peng, X.; Yang, G. Transcriptome-microRNA analysis of Sarcoptes scabiei and host immune response. PLoS ONE 2017, 12, e0177733. [Google Scholar] [CrossRef]
- Tarigan, S.; Huntley, J.F. Failure to protect goats following vaccination with soluble proteins of Sarcoptes scabiei: Evidence for a role for IgE antibody in protection. Vet. Parasitol. 2005, 133, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Harumal, P.; Morgan, M.; Walton, S.F.; Holt, D.C.; Rode, J.; Arlian, L.G.; Currie, B.J.; Kemp, D.J. Identification of a homologue of a house dust mite allergen in a cDNA library from Sarcoptes scabiei var hominis and evaluation of its vaccine potential in a rabbit/S. scabiei var. canis model. Am. J. Trop. Med. Hyg. 2003, 68, 54–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, N.; Zhang, H.; Ren, Y.; He, R.; Xu, J.; Li, C.; Lai, W.; Gu, X.; Xie, Y.; Peng, X. A chitinase-like protein from Sarcoptes scabiei as a candidate anti-mite vaccine that contributes to immune protection in rabbits. Parasites Vectors 2018, 11, 599. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, T.E.; Maizels, R.M.; Allen, J.E. Chitinases and chitinase-like proteins: Potential therapeutic targets for the treatment of T-helper type 2 allergies. Clin. Exp. Allergy 2009, 39, 943–955. [Google Scholar] [CrossRef]
- He, R.; Shen, N.; Zhang, H.; Ren, Y.; He, M.; Xu, J.; Guo, C.; Xie, Y.; Gu, X.; Lai, W. Molecular characteristics and serodiagnostic potential of chitinase-like protein fromSarcoptes scabiei. Oncotarget 2017, 8, 83995–84005. [Google Scholar] [CrossRef] [Green Version]
- ARRIVE guidelines Home Page. Available online: http://www.nc3rs.org.uk/arrive-guidelines (accessed on 14 March 2020).
- Casais, R.; Dalton, K.P.; Millan, J.; Balseiro, A.; Oleaga, A.; Solano, P.; Goyache, F.; Prieto, J.M.; Parra, F. Primary and secondary experimental infestation of rabbits (Oryctolagus cuniculus) with Sarcoptes scabiei from a wild rabbit: Factors determining resistance to reinfestation. Vet. Parasitol. 2014, 203, 173–183. [Google Scholar] [CrossRef]
- Rodríguezcadenas, F.; Carbajalgonzález, M.T.; Fregenedagrandes, J.M.; Allergancedo, J.M.; Rojovázquez, F.A. Clinical evaluation and antibody responses in sheep after primary and secondary experimental challenges with the mange mite Sarcoptes scabiei var. ovis. Vet. Immunol. Immunopathol. 2010, 133, 109–116. [Google Scholar]
- Liu, X.; Walton, S.; Mounsey, K. Vaccine against scabies: Necessity and possibility. Parasitology 2014, 141, 725–732. [Google Scholar] [CrossRef]
- Engelman, D.; Steer, A.C. Diagnosis, treatment, and control of scabies: Can we do better? Lancet Infect. Dis. 2018, 188, 822. [Google Scholar] [CrossRef]
- Wei, W.; Ren, Y.; Shen, N.; Song, H.; Xu, J.; Hua, R.; Zhang, H.; Angel, C.; Gu, X.; Kuang, L.; et al. Comparative analysis of host resistance to Sarcoptes scabiei var. cuniculi in two different rabbit breeds. Parasit Vectors 2019, 12, 530. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, J.S. Scabies: More than just an irritation. Postgrad. Med. J. 2004, 80, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kessler, E.; Matthes, H.F.; Schein, E.; Wendt, M. Detection of antibodies in sera of weaned pigs after contact infection with Sarcoptes scabiei var. suis and after treatment with an antiparasitic agent by three different indirect ELISAs. Vet. Parasitol. 2003, 114, 63–73. [Google Scholar] [CrossRef]
- Anisuzzaman; Islam, M.K.; Alim, M.A.; Miyoshi, T.; Hatta, T.; Yamaji, K.; Matsumoto, Y.; Fujisaki, K.; Tsuji, N. Longistatin is an unconventional serine protease and induces protective immunity against tick infestation. Mol. Biochem. Parasitol. 2012, 182, 45–53. [Google Scholar]
- Kritratanasak, S.; Chiampanichayakul, S.; Kasinrerk, W. Production of IgY anti-mouse IgG antibodies from chicken eggs. Asian Pac. J. Allergy Immunol. 2004, 22, 61–68. [Google Scholar] [PubMed]
- Burgess, S.T.G.; Nunn, F.; Nath, M.; Frew, D.; Wells, B.; Marr, E.J.; Huntley, J.F.; Mcneilly, T.N.; Nisbet, A.J. A recombinant subunit vaccine for the control of ovine psoroptic mange (sheep scab). Vet. Res. 2016, 47, 26. [Google Scholar] [CrossRef] [Green Version]
- Roberts, L.J.; Huffam, S.E.; Walton, S.F.; Currie, B.J. Crusted scabies: Clinical and immunological findings in seventy-eight patients and a review of the literature. J. Infect. 2005, 50, 375–381. [Google Scholar] [CrossRef]
- Arlian, L.G.; Feldmeier, H.; Morgan, M.S. The Potential for a Blood Test for Scabies. PLoS Negl. Trop. Dis. 2015, 9, e0004188. [Google Scholar] [CrossRef] [Green Version]
- Hagan, P.; Blumenthal, U.J.; Dunn, D.; Simpson, A.J.G.; Wilkins, H.A. Human IgE, IgG4 and resistance to reinfection with Schistosomahaematobium. Nature 1991, 349, 243. [Google Scholar] [CrossRef]
- Nyindo, M.; Kariuki, T.M.; Mola, P.W.; Farah, I.O.; Elson, L.; Blanton, R.E.; King, C.L. Role of Adult Worm Antigen-Specific Immunoglobulin E in Acquired Immunity to Schistosoma mansoni Infection in Baboons. Infect. Immun. 1999, 67, 636–642. [Google Scholar] [CrossRef] [Green Version]
- Walton, S.F.; Pizzutto, S.; Slender, A.; Viberg, L.; Holt, D.; Hales, B.J.; Kemp, D.J.; Currie, B.J.; Rolland, J.M.; O’Hehir, R. Increased allergic immune response to Sarcoptes scabiei antigens in crusted versus ordinary scabies. Clin. Vaccine Immunol. 2010, 17, 1428–1438. [Google Scholar] [CrossRef] [Green Version]
- Mounsey, K.E.; Murray, H.C.; Bielefeldtohmann, H.; Pasay, C.; Holt, D.C.; Currie, B.J.; Walton, S.F.; Mccarthy, J.S. Prospective Study in a Porcine Model of Sarcoptes scabiei Indicates the Association of Th2 and Th17 Pathways with the Clinical Severity of Scabies. PLoS Negl. Trop. Dis. 2015, 9, e0003498. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arlian, L.G.; Morgan, M.S.; Neal, J.S. Modulation of cytokine expression in human keratinocytes and fibroblasts by extracts of scabies mites. Am. J. Trop. Med. Hyg. 2003, 69, 652–656. [Google Scholar] [CrossRef] [PubMed]
- Mullins, J.S.; Arlian, L.G.; Morgan, M.S. Extracts of Sarcoptes scabiei De Geer downmodulate secretion of IL-8 by skin keratinocytes and fibroblasts and of GM-CSF by fibroblasts in the presence of proinflammatory cytokines. J. Med. Entomol. 2009, 46, 845–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Prior to Challenge | Week 1 Post-Challenge | Week 2 Post-Challenge | Week 3 Post-Challenge | Week 4 Post-Challenge |
|---|---|---|---|---|---|
| group 1 (PBS) (n = 10) | 0a (0/10) | 2.1 ± 0.88ab (10/0) | 1.7 ± 0.67ab (10/0) | 2.8 ± 1.14ac (10/0) | 4.5 ± 0.53ad (10/0) |
| group 2 (QuilA adjuvant) (n = 10) | 0a (0/10) | 1.7 ± 0.48abcb (10/0) | 1.9 ± 0.74ab (10/0) | 2.5 ± 0.71ac (10/0) | 4.2 ± 0.79ad (10/0) |
| group 3 (Trx-His-S tag protein) (n = 10) | 0a (0/10) | 1.6 ± 0.70abcb (9/1) | 1.7 ± 0.67ab (10/0) | 2.1 ± 0.57ab (10/0) | 4.3 ± 0.67ac (10/0) |
| group 4 (rSsCLP5) (n = 10) | 0a (0/10) | 2.0 ± 0.67abb (10/0) | 1.2 ± 1.03abbc (3/7) | 1.1 ± 0.74bc (4/6) | 1.7 ± 1.25bbc (2/8) |
| group 5 (rSsCLP5’) (n = 10) | 0a (0/10) | 1.4 ± 0.70abcb (10/0) | 1.1 ± 0.74abb (2/8) | 1.2 ± 1.03bb (3/7) | 1.9 ± 1.66bb (3/7) |
| group 6 (rSsCLP12) (n = 10) | 0a (0/10) | 1.6 ± 0.70abcb (10/0) | 1.1 ± 1.10abb (4/6) | 1.0 ± 0.82bb (3/7) | 1.6 ± 1.43bb (3/7) |
| group 7 (rSsCLP12’) (n = 10) | 0a (0/10) | 1.3 ± 0.82bcb (10/0) | 1.1 ± 0.88abb (2/8) | 1.3 ± 1.16bb (3/7) | 1.4 ± 1.71bb (3/7) |
| group 8 (rSsCLP5 + rSsCLP12) (n = 10) | 0a (0/10) | 1.3 ± 0.67bcb (10/0) | 0.4 ± 0.70ba (1/9) | 0.7 ± 1.06bab (1/9) | 1.2 ± 1.03bb (1/9) |
| group 9 (rSsCLP5 + rSsCLP12’) (n = 10) | 0a (0/10) | 1.1 ± 0.88cb (10/0) | 0.7±0.67bab (2/8) | 1.1 ± 0.88bb (2/8) | 1.0 ± 1.05bb (2/8) |
| Group | Number of Mites | Mite Reduction Rate |
|---|---|---|
| group 1 (PBS) (n = 10) | 4719 ± 1072.39a | -- |
| group 2 (QuilA adjuvant) (n = 10) | 4464 ± 1663.22a | 5.41% |
| group 3 (Trx-His-S tag protein) (n = 10) | 4139 ± 742.05a | 12.30% |
| group 4 (rSsCLP5) (n = 10) | 1423 ± 984.64b | 69.85% |
| group 5 (rSsCLP5’) (n = 10) | 1640 ± 1534.27b | 65.24% |
| group 6 (rSsCLP12) (n = 10) | 1280 ± 863.21b | 72.87% |
| group 7 (rSsCLP12’) (n = 10) | 1338 ± 985.67b | 71.66% |
| group 8 (rSsCLP5 + rSsCLP12) (n = 10) | 781 ± 298.54b | 83.45% |
| group 9 (rSsCLP5 + rSsCLP12’) (n = 10) | 637 ± 449.66b | 86.51% |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, N.; Wei, W.; Chen, Y.; Ren, Y.; Xiong, L.; Tao, Y.; Gu, X.; Xie, Y.; Peng, X.; Yang, G. An Antibody Persistent and Protective Two rSsCLP-Based Subunit Cocktail Vaccine against Sarcoptes scabiei in a Rabbit Model. Vaccines 2020, 8, 129. https://doi.org/10.3390/vaccines8010129
Shen N, Wei W, Chen Y, Ren Y, Xiong L, Tao Y, Gu X, Xie Y, Peng X, Yang G. An Antibody Persistent and Protective Two rSsCLP-Based Subunit Cocktail Vaccine against Sarcoptes scabiei in a Rabbit Model. Vaccines. 2020; 8(1):129. https://doi.org/10.3390/vaccines8010129
Chicago/Turabian StyleShen, Nengxing, Wenrui Wei, Yuhang Chen, Yongjun Ren, Lang Xiong, Yuanyuan Tao, Xiaobin Gu, Yue Xie, Xuerong Peng, and Guangyou Yang. 2020. "An Antibody Persistent and Protective Two rSsCLP-Based Subunit Cocktail Vaccine against Sarcoptes scabiei in a Rabbit Model" Vaccines 8, no. 1: 129. https://doi.org/10.3390/vaccines8010129