The Immunomodulatory and Antimicrobial Properties of the Vertebrate Ribonuclease A Superfamily


Abstract
:1. Introduction
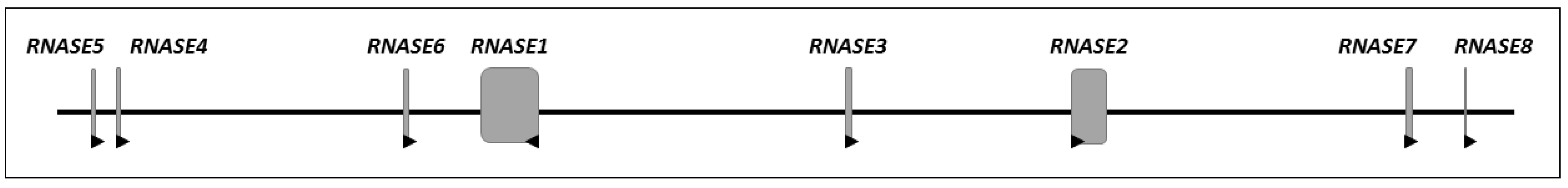
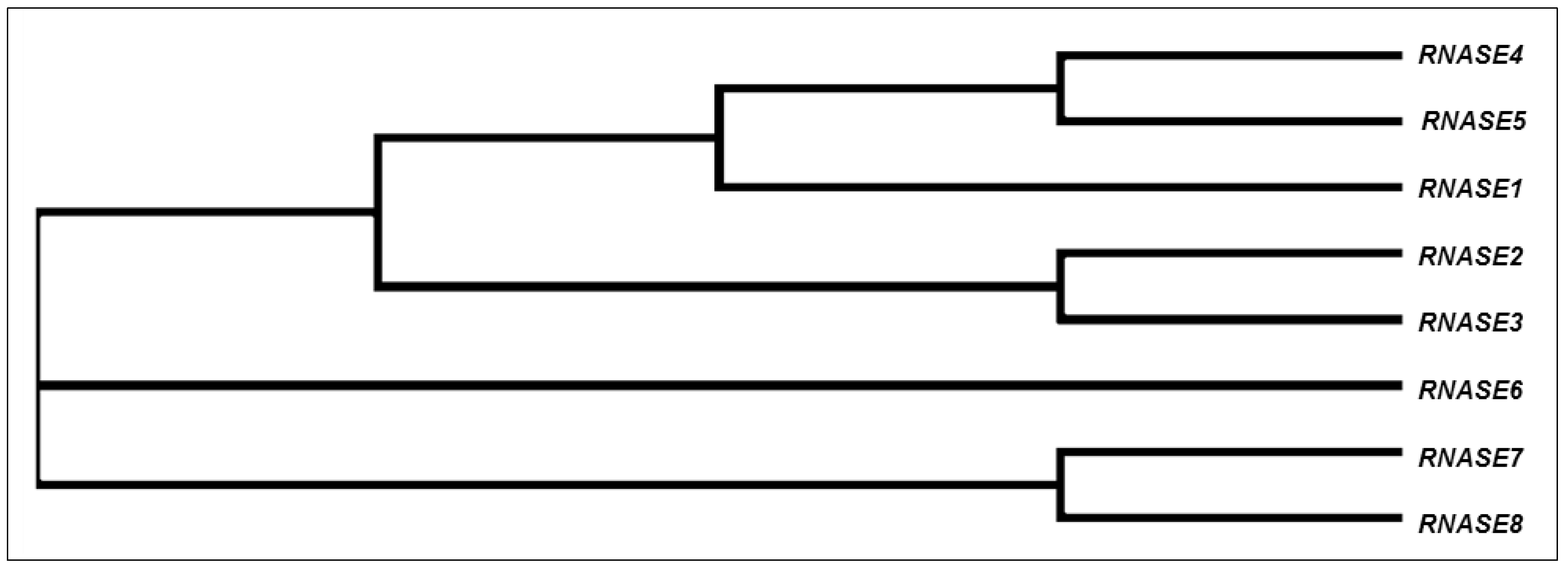
2. Evolution of the Ribonuclease A Superfamily
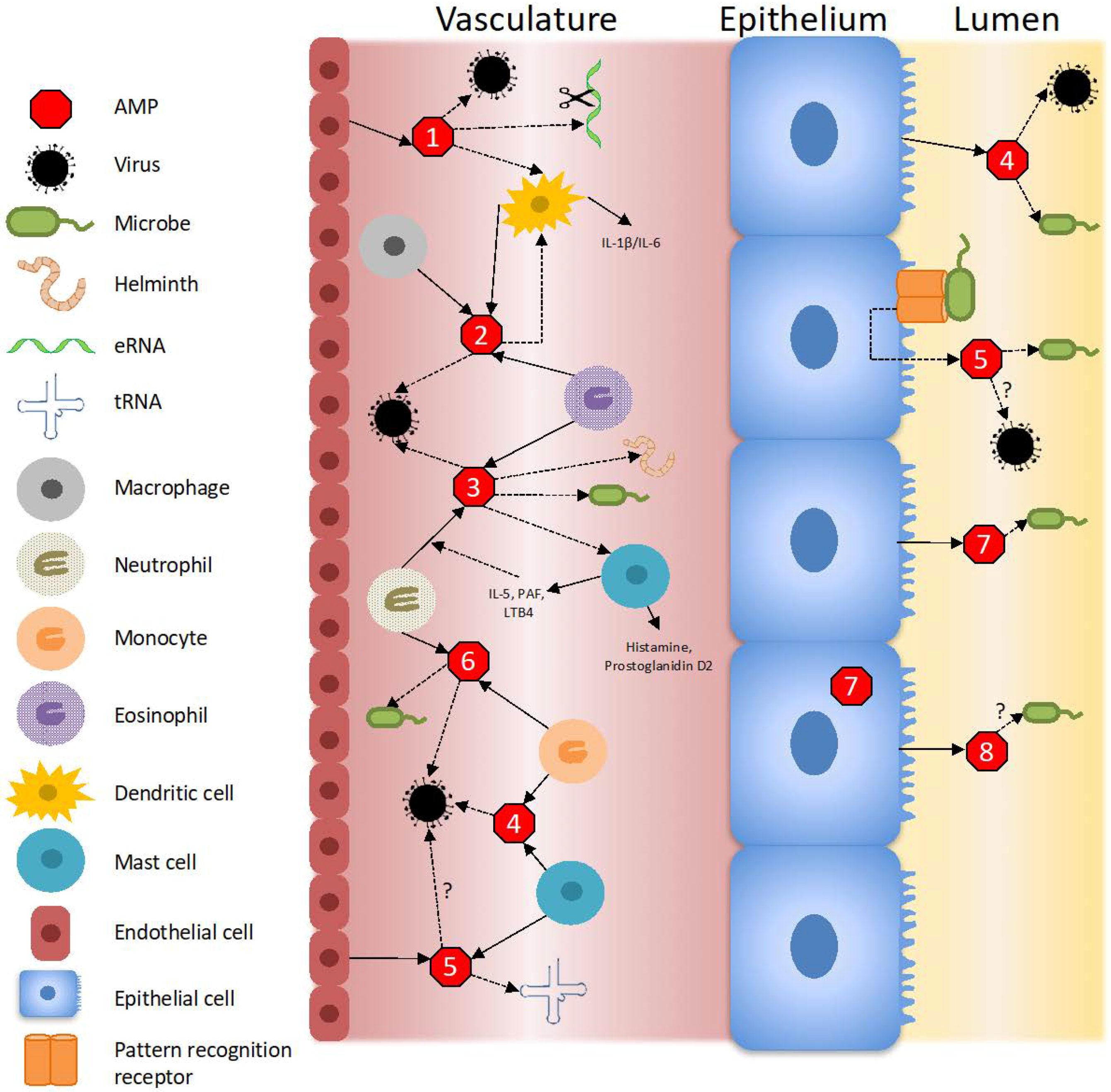
3. The Antimicrobial and Immunomodulatory Properties of the Ribonuclease A Superfamily
4. Ribonuclease 1
5. The Eosinophilic Ribonucleases—Ribonuclease 2 and 3
6. Ribonuclease 4
7. Ribonuclease 5 (Angiogenin)
8. Ribonuclease 6
9. Ribonuclease 7
10. Ribonuclease 8
11. The Non-Canonical Ribonucleases—Ribonuclease 9–13
12. Other Immunomodulatory Ribonucleases
13. Clinical Implications in Health and Disease
14. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cho, S.; Beintema, J.J.; Zhang, J. The ribonuclease A superfamily of mammals and birds: Identifying new members and tracing evolutionary histories. Genomics 2005, 85, 208–220. [Google Scholar] [CrossRef] [PubMed]
- Goo, S.M.; Cho, S. The expansion and functional diversification of the mammalian ribonuclease a superfamily epitomizes the efficiency of multigene families at generating biological novelty. Genome Biol. Evol. 2013, 5, 2124–2140. [Google Scholar] [CrossRef] [PubMed]
- Boix, E.; Nogues, M.V. Mammalian antimicrobial proteins and peptides: Overview on the RNase A superfamily members involved in innate host defence. Mol. Biosyst. 2007, 3, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.K.; Haigh, B.J.; Griffin, F.J.; Wheeler, T.T. The mammalian secreted RNases: Mechanisms of action in host defence. Innate Immun. 2013, 19, 86–97. [Google Scholar] [CrossRef] [PubMed]
- Dyer, K.D.; Rosenberg, H.F. The RNase a superfamily: Generation of diversity and innate host defense. Mol. Divers. 2006, 10, 585–597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Nei, M.; Gu, X.; Sitnikova, T. Evolution by the birth-and-death process in multigene families of the vertebrate immune system. Proc. Natl. Acad. Sci. USA 1997, 94, 7799–7806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simanski, M.; Koten, B.; Schroder, J.M.; Glaser, R.; Harder, J. Antimicrobial RNases in cutaneous defense. J. Innate Immun. 2012, 4, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Schwaderer, A.; Hains, D.S.; Spencer, J.D. Amplifying renal immunity: The role of antimicrobial peptides in pyelonephritis. Nat. Rev. Nephrol. 2015, 11, 642–655. [Google Scholar] [CrossRef] [PubMed]
- Koczera, P.; Martin, L.; Marx, G.; Schuerholz, T. The Ribonuclease A Superfamily in Humans: Canonical RNases as the Buttress of Innate Immunity. Int. J. Mol. Sci. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Torrent, M.; Pulido, D.; Valle, J.; Nogues, M.V.; Andreu, D.; Boix, E. Ribonucleases as a host-defence family: Evidence of evolutionarily conserved antimicrobial activity at the N-terminus. Biochem. J. 2013, 456, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Hamann, K.J.; Barker, R.L.; Loegering, D.A.; Gleich, G.J. Comparative toxicity of purified human eosinophil granule proteins for newborn larvae of Trichinella spiralis. J. Parasitol. 1987, 73, 523–529. [Google Scholar] [CrossRef] [PubMed]
- Lehrer, R.I.; Szklarek, D.; Barton, A.; Ganz, T.; Hamann, K.J.; Gleich, G.J. Antibacterial properties of eosinophil major basic protein and eosinophil cationic protein. J. Immunol. 1989, 142, 4428–4434. [Google Scholar] [PubMed]
- Eriksson, J.; Reimert, C.M.; Kabatereine, N.B.; Kazibwe, F.; Ireri, E.; Kadzo, H.; Eltahir, H.B.; Mohamed, A.O.; Vennervald, B.J.; Venge, P. The 434(G>C) polymorphism within the coding sequence of Eosinophil Cationic Protein (ECP) correlates with the natural course of Schistosoma mansoni infection. Int. J. Parasitol. 2007, 37, 1359–1366. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Schroder, J.M. RNase 7, a novel innate immune defense antimicrobial protein of healthy human skin. J. Biol. Chem. 2002, 277, 46779–46784. [Google Scholar] [CrossRef] [PubMed]
- Domachowske, J.B.; Dyer, K.D.; Bonville, C.A.; Rosenberg, H.F. Recombinant human eosinophil-derived neurotoxin/RNase 2 functions as an effective antiviral agent against respiratory syncytial virus. J. Infect. Dis. 1998, 177, 1458–1464. [Google Scholar] [CrossRef] [PubMed]
- Rudolph, B.; Podschun, R.; Sahly, H.; Schubert, S.; Schroder, J.M.; Harder, J. Identification of RNase 8 as a novel human antimicrobial protein. Antimicrob. Agents Chemother. 2006, 50, 3194–3196. [Google Scholar] [CrossRef] [PubMed]
- Rugeles, M.T.; Trubey, C.M.; Bedoya, V.I.; Pinto, L.A.; Oppenheim, J.J.; Rybak, S.M.; Shearer, G.M. Ribonuclease is partly responsible for the HIV-1 inhibitory effect activated by HLA alloantigen recognition. AIDS 2003, 17, 481–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorrentino, S. The eight human “canonical” ribonucleases: Molecular diversity, catalytic properties, and special biological actions of the enzyme proteins. FEBS Lett. 2010, 584, 2194–2200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosenberg, H.F. RNase A ribonucleases and host defense: An evolving story. J. Leukoc. Biol. 2008, 83, 1079–1087. [Google Scholar] [CrossRef] [PubMed]
- Lu, L.; Li, J.; Moussaoui, M.; Boix, E. Immune Modulation by Human Secreted RNases at the Extracellular Space. Front. Immunol. 2018, 9, 1012. [Google Scholar] [CrossRef] [PubMed]
- Noll, F.; Behnke, J.; Leiting, S.; Troidl, K.; Alves, G.T.; Muller-Redetzky, H.; Preissner, K.T.; Fischer, S. Self-extracellular RNA acts in synergy with exogenous danger signals to promote inflammation. PLoS ONE 2017, 12, e0190002. [Google Scholar] [CrossRef] [PubMed]
- Zernecke, A.; Preissner, K.T. Extracellular Ribonucleic Acids (RNA) Enter the Stage in Cardiovascular Disease. Circ. Res. 2016, 118, 469–479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Chen, Q.; Rosenberg, H.F.; Rybak, S.M.; Newton, D.L.; Wang, Z.Y.; Fu, Q.; Tchernev, V.T.; Wang, M.; Schweitzer, B.; et al. Human ribonuclease A superfamily members, eosinophil-derived neurotoxin and pancreatic ribonuclease, induce dendritic cell maturation and activation. J. Immunol. 2004, 173, 6134–6142. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.; Koczera, P.; Simons, N.; Zechendorf, E.; Hoeger, J.; Marx, G.; Schuerholz, T. The Human Host Defense Ribonucleases 1, 3 and 7 Are Elevated in Patients with Sepsis after Major Surgery—A Pilot Study. Int. J. Mol. Sci. 2016, 17, 294. [Google Scholar] [CrossRef] [PubMed]
- Fischer, S.; Cabrera-Fuentes, H.A.; Noll, T.; Preissner, K.T. Impact of extracellular RNA on endothelial barrier function. Cell Tissue Res. 2014, 355, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F. Eosinophil-Derived Neurotoxin (EDN/RNase 2) and the Mouse Eosinophil-Associated RNases (mEars): Expanding Roles in Promoting Host Defense. Int. J. Mol. Sci. 2015, 16, 15442–15455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, D.; Chen, Q.; Su, S.B.; Zhang, P.; Kurosaka, K.; Caspi, R.R.; Michalek, S.M.; Rosenberg, H.F.; Zhang, N.; Oppenheim, J.J. Eosinophil-derived neurotoxin acts as an alarmin to activate the TLR2-MyD88 signal pathway in dendritic cells and enhances Th2 immune responses. J. Exp. Med. 2008, 205, 79–90. [Google Scholar] [CrossRef] [PubMed]
- Torrent, M.; Badia, M.; Moussaoui, M.; Sanchez, D.; Nogues, M.V.; Boix, E. Comparison of human RNase 3 and RNase 7 bactericidal action at the Gram-negative and Gram-positive bacterial cell wall. FEBS J. 2010, 277, 1713–1725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulido, D.; Arranz-Trullen, J.; Prats-Ejarque, G.; Velazquez, D.; Torrent, M.; Moussaoui, M.; Boix, E. Insights into the Antimicrobial Mechanism of Action of Human RNase6: Structural Determinants for Bacterial Cell Agglutination and Membrane Permeation. Int. J. Mol. Sci. 2016, 17, 552. [Google Scholar] [CrossRef] [PubMed]
- Weller, P.F.; Spencer, L.A. Functions of tissue-resident eosinophils. Nat. Rev. Immunol. 2017, 17, 746–760. [Google Scholar] [CrossRef] [PubMed]
- Chihara, J.; Urayama, O.; Tsuda, A.; Kakazu, T.; Higashimoto, I.; Yamada, H. Eosinophil cationic protein induces insulin-like growth factor I receptor expression on bronchial epithelial cells. Int. Arch. Allergy Immunol 1996, 111 (Suppl. 1), 43–45. [Google Scholar] [CrossRef] [PubMed]
- Bystrom, J.; Amin, K.; Bishop-Bailey, D. Analysing the eosinophil cationic protein--a clue to the function of the eosinophil granulocyte. Respir. Res. 2011, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Zagai, U.; Dadfar, E.; Lundahl, J.; Venge, P.; Skold, C.M. Eosinophil cationic protein stimulates TGF-beta1 release by human lung fibroblasts in vitro. Inflammation 2007, 30, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Sheng, J.; Hu, J.K.; Yu, W.; Kishikawa, H.; Hu, M.G.; Shima, K.; Wu, D.; Xu, Z.; Xin, W.; et al. Ribonuclease 4 protects neuron degeneration by promoting angiogenesis, neurogenesis, and neuronal survival under stress. Angiogenesis 2013, 16, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Padhi, A.K.; Narain, P.; Dave, U.; Satija, R.; Patir, A.; Gomes, J. Insights into the role of ribonuclease 4 polymorphisms in amyotrophic lateral sclerosis. J. Biomol. Struct. Dyn. 2018, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Cocchi, F.; DeVico, A.L.; Lu, W.; Popovic, M.; Latinovic, O.; Sajadi, M.M.; Redfield, R.R.; Lafferty, M.K.; Galli, M.; Garzino-Demo, A.; et al. Soluble factors from T cells inhibiting X4 strains of HIV are a mixture of beta chemokines and RNases. Proc. Natl. Acad. Sci. USA 2012, 109, 5411–5416. [Google Scholar] [CrossRef] [PubMed]
- Liang, S.; Acharya, K.R. Structural basis of substrate specificity in porcine RNase 4. FEBS J. 2016, 283, 912–928. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Terzyan, S.S.; Peracaula, R.; de Llorens, R.; Tsushima, Y.; Yamada, H.; Seno, M.; Gomis-Ruth, F.X.; Coll, M. The three-dimensional structure of human RNase 4, unliganded and complexed with d(Up), reveals the basis for its uridine selectivity. J. Mol. Biol. 1999, 285, 205–214. [Google Scholar] [CrossRef] [PubMed]
- Harris, P.J.K.; Smolenski, G.; Callaghan, M.; Broadhurst, M.K.; Kim, K.; Wheeler, T.T. Characterisation of the anti-microbial activity of bovine milk ribonuclease4 and ribonuclease5 (angiogenin). Int. Dairy J. 2010, 20, 400–407. [Google Scholar] [CrossRef]
- Di Liddo, R.; Dalzoppo, D.; Baiguera, S.; Conconi, M.T.; Dettin, M.; Parnigotto, P.P.; Grandi, C. In vitro biological activity of bovine milk ribonuclease-4. Mol. Med. Rep. 2010, 3, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Miyake, M.; Goodison, S.; Lawton, A.; Gomes-Giacoia, E.; Rosser, C.J. Angiogenin promotes tumoral growth and angiogenesis by regulating matrix metallopeptidase-2 expression via the ERK1/2 pathway. Oncogene 2015, 34, 890–901. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Xu, Z. Three decades of research on angiogenin: A review and perspective. Acta Biochim. Biophys. Sin. (Shanghai) 2016, 48, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, K.A.; Silberstein, L.; Li, S.; Severe, N.; Hu, M.G.; Yang, H.; Scadden, D.T.; Hu, G.F. Angiogenin Promotes Hematopoietic Regeneration by Dichotomously Regulating Quiescence of Stem and Progenitor Cells. Cell 2016, 166, 894–906. [Google Scholar] [CrossRef] [PubMed]
- Tschesche, H.; Kopp, C.; Horl, W.H.; Hempelmann, U. Inhibition of degranulation of polymorphonuclear leukocytes by angiogenin and its tryptic fragment. J. Biol. Chem. 1994, 269, 30274–30280. [Google Scholar] [PubMed]
- Shcheglovitova, O.N.; Maksyanina, E.V.; Ionova, I.I.; Rustam’yan, Y.L.; Komolova, G.S. Cow milk angiogenin induces cytokine production in human blood leukocytes. Bull. Exp. Biol. Med. 2003, 135, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Hooper, L.V.; Stappenbeck, T.S.; Hong, C.V.; Gordon, J.I. Angiogenins: A new class of microbicidal proteins involved in innate immunity. Nat. Immunol. 2003, 4, 269–273. [Google Scholar] [CrossRef] [PubMed]
- Abtin, A.; Eckhart, L.; Mildner, M.; Ghannadan, M.; Harder, J.; Schroder, J.M.; Tschachler, E. Degradation by stratum corneum proteases prevents endogenous RNase inhibitor from blocking antimicrobial activities of RNase 5 and RNase 7. J. Investig. Dermatol. 2009, 129, 2193–2201. [Google Scholar] [CrossRef] [PubMed]
- Avdeeva, S.V.; Chernukha, M.U.; Shaginyan, I.A.; Tarantul, V.Z.; Naroditsky, B.S. Human angiogenin lacks specific antimicrobial activity. Curr. Microbiol. 2006, 53, 477–478. [Google Scholar] [CrossRef] [PubMed]
- Saxena, S.K.; Rybak, S.M.; Davey, R.T., Jr.; Youle, R.J.; Ackerman, E.J. Angiogenin is a cytotoxic, tRNA-specific ribonuclease in the RNase A superfamily. J. Biol. Chem. 1992, 267, 21982–21986. [Google Scholar] [PubMed]
- Tsuji, T.; Sun, Y.; Kishimoto, K.; Olson, K.A.; Liu, S.; Hirukawa, S.; Hu, G.F. Angiogenin is translocated to the nucleus of HeLa cells and is involved in ribosomal RNA transcription and cell proliferation. Cancer Res. 2005, 65, 1352–1360. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, S.; Ivanov, P.; Hu, G.F.; Anderson, P. Angiogenin cleaves tRNA and promotes stress-induced translational repression. J. Cell Biol. 2009, 185, 35–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Hu, G.F. Emerging role of angiogenin in stress response and cell survival under adverse conditions. J. Cell. Physiol. 2012, 227, 2822–2826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Yu, W.; Hu, G.F. Angiogenin inhibits nuclear translocation of apoptosis inducing factor in a Bcl-2-dependent manner. J. Cell. Physiol. 2012, 227, 1639–1644. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Eichler, T.E.; Beceiro, S.; Li, B.; Easterling, R.S.; Carpenter, A.R.; James, C.L.; McHugh, K.M.; Hains, D.S.; Partida-Sanchez, S.; et al. Ribonucleases 6 and 7 have antimicrobial function in the human and murine urinary tract. Kidney Int. 2015, 87, 151–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, L.J.; McClellan, S.; Hansen, P.J. Differentiation of the endometrial macrophage during pregnancy in the cow. PLoS ONE 2010, 5, e13213. [Google Scholar] [CrossRef] [PubMed]
- Christensen-Quick, A.; Lafferty, M.; Sun, L.; Marchionni, L.; DeVico, A.; Garzino-Demo, A. Human Th17 Cells Lack HIV-Inhibitory RNases and Are Highly Permissive to Productive HIV Infection. J. Virol. 2016, 90, 7833–7847. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kopfnagel, V.; Wagenknecht, S.; Harder, J.; Hofmann, K.; Kleine, M.; Buch, A.; Sodeik, B.; Werfel, T. RNase 7 Strongly Promotes TLR9-Mediated DNA Sensing by Human Plasmacytoid Dendritic Cells. J. Investig. Dermatol. 2018, 138, 872–881. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.D.; Schwaderer, A.L.; Dirosario, J.D.; McHugh, K.M.; McGillivary, G.; Justice, S.S.; Carpenter, A.R.; Baker, P.B.; Harder, J.; Hains, D.S. Ribonuclease 7 is a potent antimicrobial peptide within the human urinary tract. Kidney Int. 2011, 80, 174–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Y.C.; Lin, Y.M.; Chang, T.W.; Wu, S.J.; Lee, Y.S.; Chang, M.D.; Chen, C.; Wu, S.H.; Liao, Y.D. The flexible and clustered lysine residues of human ribonuclease 7 are critical for membrane permeability and antimicrobial activity. J. Biol. Chem. 2007, 282, 4626–4633. [Google Scholar] [CrossRef] [PubMed]
- Spencer, J.D.; Schwaderer, A.L.; Wang, H.; Bartz, J.; Kline, J.; Eichler, T.; DeSouza, K.R.; Sims-Lucas, S.; Baker, P.; Hains, D.S. Ribonuclease 7, an antimicrobial peptide upregulated during infection, contributes to microbial defense of the human urinary tract. Kidney Int. 2013, 83, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Kopfnagel, V.; Wagenknecht, S.; Brand, L.; Zeitvogel, J.; Harder, J.; Hofmann, K.; Kleine, M.; Werfel, T. RNase 7 downregulates TH2 cytokine production by activated human T cells. Allergy 2017, 72, 1694–1703. [Google Scholar] [CrossRef] [PubMed]
- Bernard, J.J.; Gallo, R.L. Protecting the boundary: The sentinel role of host defense peptides in the skin. Cell. Mol. Life Sci. 2011, 68, 2189–2199. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dyer, K.D.; Rosenberg, H.F. RNase 8, a novel RNase A superfamily ribonuclease expressed uniquely in placenta. Nucleic Acids Res. 2002, 30, 1169–1175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennedy, R.B.; Ovsyannikova, I.G.; Haralambieva, I.H.; Lambert, N.D.; Pankratz, V.S.; Poland, G.A. Genome-wide SNP associations with rubella-specific cytokine responses in measles-mumps-rubella vaccine recipients. Immunogenetics 2014, 66, 493–499. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, G.Z.; Li, J.Y.; Li, F.; Wang, H.Y.; Shi, G.X. Human ribonuclease 9, a member of ribonuclease A superfamily, specifically expressed in epididymis, is a novel sperm-binding protein. Asian J. Androl. 2009, 11, 240–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gansler, J.; Preissner, K.T.; Fischer, S. Influence of proinflammatory stimuli on the expression of vascular ribonuclease 1 in endothelial cells. FASEB J. 2014, 28, 752–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cabrera-Fuentes, H.A.; Ruiz-Meana, M.; Simsekyilmaz, S.; Kostin, S.; Inserte, J.; Saffarzadeh, M.; Galuska, S.P.; Vijayan, V.; Barba, I.; Barreto, G.; et al. RNase1 prevents the damaging interplay between extracellular RNA and tumour necrosis factor-alpha in cardiac ischaemia/reperfusion injury. Thromb. Haemost. 2014, 112, 1110–1119. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewicz, D.; Bergmann, S.; Didiasova, M.; Giaimo, B.D.; Borggrefe, T.; Mieth, M.; Hocke, A.C.; Lochnit, G.; Schaefer, L.; Hammerschmidt, S.; et al. Host-derived extracellular RNA promotes adhesion of Streptococcus pneumoniae to endothelial and epithelial cells. Sci. Rep. 2016, 6, 37758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann-Geller, B.; Koppert, S.; Fischer, S.; Cabrera-Fuentes, H.A.; Lefevre, S.; Rickert, M.; Steinmeyer, J.; Rehart, S.; Umscheid, T.; Schonburg, M.; et al. Influence of Extracellular RNAs, Released by Rheumatoid Arthritis Synovial Fibroblasts, on Their Adhesive and Invasive Properties. J. Immunol. 2016, 197, 2589–2597. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedoya, V.I.; Boasso, A.; Hardy, A.W.; Rybak, S.; Shearer, G.M.; Rugeles, M.T. Ribonucleases in HIV type 1 inhibition: Effect of recombinant RNases on infection of primary T cells and immune activation-induced RNase gene and protein expression. AIDS Res. Hum. Retroviruses 2006, 22, 897–907. [Google Scholar] [CrossRef] [PubMed]
- Durack, D.T.; Sumi, S.M.; Klebanoff, S.J. Neurotoxicity of human eosinophils. Proc. Natl. Acad. Sci. USA 1979, 76, 1443–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu-Ghazaleh, R.I.; Dunnette, S.L.; Loegering, D.A.; Checkel, J.L.; Kita, H.; Thomas, L.L.; Gleich, G.J. Eosinophil granule proteins in peripheral blood granulocytes. J. Leukoc. Biol. 1992, 52, 611–618. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.K.; Seo, J.K.; Ban, S.H.; Fujisawa, T.; Kim, D.W.; Callaway, Z. Eosinophil-derived neurotoxin levels at 3 months post-respiratory syncytial virus bronchiolitis are a predictive biomarker of recurrent wheezing. Biomarkers 2013, 18, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Phipps, S.; Lam, C.E.; Mahalingam, S.; Newhouse, M.; Ramirez, R.; Rosenberg, H.F.; Foster, P.S.; Matthaei, K.I. Eosinophils contribute to innate antiviral immunity and promote clearance of respiratory syncytial virus. Blood 2007, 110, 1578–1586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, Z.; Dyer, K.D.; Xie, Z.; Radinger, M.; Rosenberg, H.F. GATA transcription factors regulate the expression of the human eosinophil-derived neurotoxin (RNase 2) gene. J. Biol. Chem. 2009, 284, 13099–13109. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D.; Tiffany, H.L.; Gonzalez, M. Rapid evolution of a unique family of primate ribonuclease genes. Nat. Genet. 1995, 10, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Neves, J.S.; Weller, P.F. Functional extracellular eosinophil granules: Novel implications in eosinophil immunobiology. Curr. Opin. Immunol. 2009, 21, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Venge, P.; Bystrom, J.; Carlson, M.; Hakansson, L.; Karawacjzyk, M.; Peterson, C.; Seveus, L.; Trulson, A. Eosinophil cationic protein (ECP): Molecular and biological properties and the use of ECP as a marker of eosinophil activation in disease. Clin. Exp. Allergy 1999, 29, 1172–1186. [Google Scholar] [CrossRef] [PubMed]
- Robinson, D.S.; Assoufi, B.; Durham, S.R.; Kay, A.B. Eosinophil cationic protein (ECP) and eosinophil protein X (EPX) concentrations in serum and bronchial lavage fluid in asthma. Effect of prednisolone treatment. Clin. Exp. Allergy 1995, 25, 1118–1127. [Google Scholar] [CrossRef] [PubMed]
- Koh, G.C.; Shek, L.P.; Goh, D.Y.; Van Bever, H.; Koh, D.S. Eosinophil cationic protein: Is it useful in asthma? A systematic review. Respir. Med. 2007, 101, 696–705. [Google Scholar] [CrossRef] [PubMed]
- Hamann, K.J.; Ten, R.M.; Loegering, D.A.; Jenkins, R.B.; Heise, M.T.; Schad, C.R.; Pease, L.R.; Gleich, G.J.; Barker, R.L. Structure and chromosome localization of the human eosinophil-derived neurotoxin and eosinophil cationic protein genes: Evidence for intronless coding sequences in the ribonuclease gene superfamily. Genomics 1990, 7, 535–546. [Google Scholar] [CrossRef]
- Boix, E.; Torrent, M.; Sanchez, D.; Nogues, M.V. The antipathogen activities of eosinophil cationic protein. Curr. Pharm. Biotechnol. 2008, 9, 141–152. [Google Scholar] [CrossRef] [PubMed]
- Carreras, E.; Boix, E.; Rosenberg, H.F.; Cuchillo, C.M.; Nogues, M.V. Both aromatic and cationic residues contribute to the membrane-lytic and bactericidal activity of eosinophil cationic protein. Biochemistry 2003, 42, 6636–6644. [Google Scholar] [CrossRef] [PubMed]
- Pulido, D.; Moussaoui, M.; Andreu, D.; Nogues, M.V.; Torrent, M.; Boix, E. Antimicrobial action and cell agglutination by the eosinophil cationic protein are modulated by the cell wall lipopolysaccharide structure. Antimicrob. Agents Chemother. 2012, 56, 2378–2385. [Google Scholar] [CrossRef] [PubMed]
- Young, J.D.; Peterson, C.G.; Venge, P.; Cohn, Z.A. Mechanism of membrane damage mediated by human eosinophil cationic protein. Nature 1986, 321, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Torrent, M.; Sanchez, D.; Buzon, V.; Nogues, M.V.; Cladera, J.; Boix, E. Comparison of the membrane interaction mechanism of two antimicrobial RNases: RNase 3/ECP and RNase 7. Biochim. Biophys. Acta 2009, 1788, 1116–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, K.R.; Ackerman, S.J. Eosinophil granule proteins: Form and function. J. Biol. Chem. 2014, 289, 17406–17415. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Domachowske, J.B. Eosinophils, eosinophil ribonucleases, and their role in host defense against respiratory virus pathogens. J. Leukoc. Biol. 2001, 70, 691–698. [Google Scholar] [PubMed]
- Moreau, J.M.; Dyer, K.D.; Bonville, C.A.; Nitto, T.; Vasquez, N.L.; Easton, A.J.; Domachowske, J.B.; Rosenberg, H.F. Diminished expression of an antiviral ribonuclease in response to pneumovirus infection in vivo. Antivir. Res. 2003, 59, 181–191. [Google Scholar] [CrossRef]
- McDevitt, A.L.; Deming, M.S.; Rosenberg, H.F.; Dyer, K.D. Gene structure and enzymatic activity of mouse eosinophil-associated ribonuclease 2. Gene 2001, 267, 23–30. [Google Scholar] [CrossRef]
- Nittoh, T.; Hirakata, M.; Mue, S.; Ohuchi, K. Identification of cDNA encoding rat eosinophil cationic protein/eosinophil-associated ribonuclease. Biochim. Biophys. Acta 1997, 1351, 42–46. [Google Scholar] [CrossRef]
- Singhania, N.A.; Dyer, K.D.; Zhang, J.; Deming, M.S.; Bonville, C.A.; Domachowske, J.B.; Rosenberg, H.F. Rapid evolution of the ribonuclease A superfamily: Adaptive expansion of independent gene clusters in rats and mice. J. Mol. Evol. 1999, 49, 721–728. [Google Scholar] [CrossRef] [PubMed]
- Larson, K.A.; Olson, E.V.; Madden, B.J.; Gleich, G.J.; Lee, N.A.; Lee, J.J. Two highly homologous ribonuclease genes expressed in mouse eosinophils identify a larger subgroup of the mammalian ribonuclease superfamily. Proc. Natl. Acad. Sci. USA 1996, 93, 12370–12375. [Google Scholar] [CrossRef] [PubMed]
- Cormier, S.A.; Larson, K.A.; Yuan, S.; Mitchell, T.L.; Lindenberger, K.; Carrigan, P.; Lee, N.A.; Lee, J.J. Mouse eosinophil-associated ribonucleases: A unique subfamily expressed during hematopoiesis. Mamm. Genome 2001, 12, 352–361. [Google Scholar] [CrossRef] [PubMed]
- Cormier, S.A.; Yuan, S.; Crosby, J.R.; Protheroe, C.A.; Dimina, D.M.; Hines, E.M.; Lee, N.A.; Lee, J.J. T(H)2-mediated pulmonary inflammation leads to the differential expression of ribonuclease genes by alveolar macrophages. Am. J. Respir. Cell Mol. Biol. 2002, 27, 678–687. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dyer, K.D.; Rosenberg, H.F. Evolution of the rodent eosinophil-associated RNase gene family by rapid gene sorting and positive selection. Proc. Natl. Acad. Sci. USA 2000, 97, 4701–4706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, K.J.; Barker, T.; Dyer, K.D.; Rice, T.A.; Percopo, C.M.; Garcia-Crespo, K.E.; Cho, S.; Lee, J.J.; Druey, K.M.; Rosenberg, H.F. Eosinophil-associated ribonuclease 11 is a macrophage chemoattractant. J. Biol. Chem. 2015, 290, 8863–8875. [Google Scholar] [CrossRef] [PubMed]
- Sheng, J.; Luo, C.; Jiang, Y.; Hinds, P.W.; Xu, Z.; Hu, G.F. Transcription of angiogenin and ribonuclease 4 is regulated by RNA polymerase III elements and a CCCTC binding factor (CTCF)-dependent intragenic chromatin loop. J. Biol. Chem. 2014, 289, 12520–12534. [Google Scholar] [CrossRef] [PubMed]
- Dyer, K.D.; Rosenberg, H.F. The mouse RNase 4 and RNase 5/ang 1 locus utilizes dual promoters for tissue-specific expression. Nucleic Acids Res. 2005, 33, 1077–1086. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hofsteenge, J.; Vicentini, A.; Zelenko, O. Ribonuclease 4, an evolutionarily highly conserved member of the superfamily. Cell. Mol. Life Sci. 1998, 54, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D. Human ribonuclease 4 (RNase 4): Coding sequence, chromosomal localization and identification of two distinct transcripts in human somatic tissues. Nucleic Acids Res. 1995, 23, 4290–4295. [Google Scholar] [CrossRef] [PubMed]
- Seno, M.; Futami, J.; Tsushima, Y.; Akutagawa, K.; Kosaka, M.; Tada, H.; Yamada, H. Molecular cloning and expression of human ribonuclease 4 cDNA. Biochim. Biophys. Acta 1995, 1261, 424–426. [Google Scholar] [CrossRef]
- Dong, X.; Cong, S. Identification of differentially expressed genes and regulatory relationships in Huntington’s disease by bioinformatics analysis. Mol. Med. Rep. 2018, 17, 4317–4326. [Google Scholar] [CrossRef] [PubMed]
- Rybak, S.M.; Vallee, B.L. Base cleavage specificity of angiogenin with Saccharomyces cerevisiae and Escherichia coli 5S RNAs. Biochemistry 1988, 27, 2288–2294. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Xu, C.; Riordan, J.F. Human angiogenin is rapidly translocated to the nucleus of human umbilical vein endothelial cells and binds to DNA. J. Cell. Biochem. 2000, 76, 452–462. [Google Scholar] [CrossRef]
- Rendl, M.; Mayer, C.; Weninger, W.; Tschachler, E. Topically applied lactic acid increases spontaneous secretion of vascular endothelial growth factor by human reconstructed epidermis. Br. J. Dermatol. 2001, 145, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, H.F.; Dyer, K.D. Molecular cloning and characterization of a novel human ribonuclease (RNase k6): Increasing diversity in the enlarging ribonuclease gene family. Nucleic Acids Res. 1996, 24, 3507–3513. [Google Scholar] [CrossRef] [PubMed]
- Prats-Ejarque, G.; Blanco, J.A.; Salazar, V.A.; Nogues, V.M.; Moussaoui, M.; Boix, E. Characterization of an RNase with two catalytic centers. Human RNase6 catalytic and phosphate-binding site arrangement favors the endonuclease cleavage of polymeric substrates. Biochim. Biophys. Acta Gen. Subj. 2018. [Google Scholar] [CrossRef] [PubMed]
- Dyer, K.D.; Rosenberg, H.F.; Zhang, J. Isolation, characterization, and evolutionary divergence of mouse RNase 6: Evidence for unusual evolution in rodents. J. Mol. Evol. 2004, 59, 657–665. [Google Scholar] [CrossRef] [PubMed]
- Amatngalim, G.D.; van Wijck, Y.; de Mooij-Eijk, Y.; Verhoosel, R.M.; Harder, J.; Lekkerkerker, A.N.; Janssen, R.A.; Hiemstra, P.S. Basal cells contribute to innate immunity of the airway epithelium through production of the antimicrobial protein RNase 7. J. Immunol. 2015, 194, 3340–3350. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, I.; Yeung, A.; Abedin, A.; Hopkinson, A.; Dua, H.S. Signalling pathways involved in ribonuclease-7 expression. Cell. Mol. Life Sci. 2011, 68, 1941–1952. [Google Scholar] [CrossRef] [PubMed]
- Simanski, M.; Dressel, S.; Glaser, R.; Harder, J. RNase 7 protects healthy skin from Staphylococcus aureus colonization. J. Investig. Dermatol. 2010, 130, 2836–2838. [Google Scholar] [CrossRef] [PubMed]
- Simanski, M.; Rademacher, F.; Schroder, L.; Schumacher, H.M.; Glaser, R.; Harder, J. IL-17A and IFN-gamma synergistically induce RNase 7 expression via STAT3 in primary keratinocytes. PLoS ONE 2013, 8, e59531. [Google Scholar] [CrossRef] [PubMed]
- Wanke, I.; Steffen, H.; Christ, C.; Krismer, B.; Gotz, F.; Peschel, A.; Schaller, M.; Schittek, B. Skin commensals amplify the innate immune response to pathogens by activation of distinct signaling pathways. J. Investig. Dermatol. 2011, 131, 382–390. [Google Scholar] [CrossRef] [PubMed]
- Burgey, C.; Kern, W.V.; Romer, W.; Sakinc, T.; Rieg, S. The innate defense antimicrobial peptides hBD3 and RNase7 are induced in human umbilical vein endothelial cells by classical inflammatory cytokines but not Th17 cytokines. Microbes Infect. 2015, 17, 353–359. [Google Scholar] [CrossRef] [PubMed]
- Firat, Y.H.; Simanski, M.; Rademacher, F.; Schroder, L.; Brasch, J.; Harder, J. Infection of keratinocytes with Trichophytum rubrum induces epidermal growth factor-dependent RNase 7 and human beta-defensin-3 expression. PLoS ONE 2014, 9, e93941. [Google Scholar] [CrossRef] [PubMed]
- Reithmayer, K.; Meyer, K.C.; Kleditzsch, P.; Tiede, S.; Uppalapati, S.K.; Glaser, R.; Harder, J.; Schroder, J.M.; Paus, R. Human hair follicle epithelium has an antimicrobial defence system that includes the inducible antimicrobial peptide psoriasin (S100A7) and RNase 7. Br. J. Dermatol. 2009, 161, 78–89. [Google Scholar] [CrossRef] [PubMed]
- Eichler, T.E.; Becknell, B.; Easterling, R.S.; Ingraham, S.E.; Cohen, D.M.; Schwaderer, A.L.; Hains, D.S.; Li, B.; Cohen, A.; Metheny, J.; et al. Insulin and the phosphatidylinositol 3-kinase signaling pathway regulate Ribonuclease 7 expression in the human urinary tract. Kidney Int. 2016, 90, 568–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambichler, T.; Skrygan, M.; Tomi, N.S.; Othlinghaus, N.; Brockmeyer, N.H.; Altmeyer, P.; Kreuter, A. Differential mRNA expression of antimicrobial peptides and proteins in atopic dermatitis as compared to psoriasis vulgaris and healthy skin. Int. Arch. Allergy Immunol. 2008, 147, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Harder, J.; Dressel, S.; Wittersheim, M.; Cordes, J.; Meyer-Hoffert, U.; Mrowietz, U.; Folster-Holst, R.; Proksch, E.; Schroder, J.M.; Schwarz, T.; et al. Enhanced expression and secretion of antimicrobial peptides in atopic dermatitis and after superficial skin injury. J. Investig. Dermatol. 2010, 130, 1355–1364. [Google Scholar] [CrossRef] [PubMed]
- Rademacher, F.; Simanski, M.; Harder, J. RNase 7 in Cutaneous Defense. Int. J. Mol. Sci. 2016, 17, 560. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Dyer, K.D.; Rosenberg, H.F. Human RNase 7: A new cationic ribonuclease of the RNase A superfamily. Nucleic Acids Res. 2003, 31, 602–607. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.M.; Wu, S.J.; Chang, T.W.; Wang, C.F.; Suen, C.S.; Hwang, M.J.; Chang, M.D.; Chen, Y.T.; Liao, Y.D. Outer membrane protein I of Pseudomonas aeruginosa is a target of cationic antimicrobial peptide/protein. J. Biol. Chem. 2010, 285, 8985–8994. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Schwaderer, A.L.; Kline, J.; Spencer, J.D.; Kline, D.; Hains, D.S. Contribution of structural domains to the activity of ribonuclease 7 against uropathogenic bacteria. Antimicrob. Agents Chemother. 2013, 57, 766–774. [Google Scholar] [CrossRef] [PubMed]
- Becknell, B.; Spencer, J.D. A Review of Ribonuclease 7’s Structure, Regulation, and Contributions to Host Defense. Int. J. Mol. Sci. 2016, 17, 423. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.C.; Moser, J.M.; Dyer, K.D.; Percopo, C.M.; Rosenberg, H.F. Genetic diversity of human RNase 8. BMC Genom. 2012, 13, 40. [Google Scholar] [CrossRef] [PubMed]
- Zhu, C.F.; Liu, Q.; Zhang, L.; Yuan, H.X.; Zhen, W.; Zhang, J.S.; Chen, Z.J.; Hall, S.H.; French, F.S.; Zhang, Y.L. RNase9, an androgen-dependent member of the RNase A family, is specifically expressed in the rat epididymis. Biol. Reprod. 2007, 76, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Li, J.; Wang, H.; Zhang, C.; Li, N.; Lin, Y.; Liu, J.; Wang, W. Cloning, expression and location of RNase9 in human epididymis. BMC Res. Notes 2008, 1, 111. [Google Scholar] [CrossRef] [PubMed]
- Penttinen, J.; Pujianto, D.A.; Sipila, P.; Huhtaniemi, I.; Poutanen, M. Discovery in silico and characterization in vitro of novel genes exclusively expressed in the mouse epididymis. Mol. Endocrinol. 2003, 17, 2138–2151. [Google Scholar] [CrossRef] [PubMed]
- Westmuckett, A.D.; Nguyen, E.B.; Herlea-Pana, O.M.; Alvau, A.; Salicioni, A.M.; Moore, K.L. Impaired sperm maturation in RNASE9 knockout mice. Biol. Reprod. 2014, 90, 120. [Google Scholar] [CrossRef] [PubMed]
- Devor, E.J.; Moffat-Wilson, K.A.; Galbraith, J.J. LOC 390443 (RNase 9) on chromosome 14q11.2 is related to the RNase A superfamily and contains a unique amino-terminal preproteinlike sequence. Hum. Biol. 2004, 76, 921–935. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.C.; Tang, C.H.; Wang, J.J. Caspase activation in response to cytotoxic Rana catesbeiana ribonuclease in MCF-7 cells. FEBS Lett. 2001, 503, 65–68. [Google Scholar] [CrossRef] [Green Version]
- Ardelt, W.; Shogen, K.; Darzynkiewicz, Z. Onconase and amphinase, the antitumor ribonucleases from Rana pipiens oocytes. Curr. Pharm. Biotechnol. 2008, 9, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Vert, A.; Castro, J.; Ribo, M.; Benito, A.; Vilanova, M. Activating transcription factor 3 is crucial for antitumor activity and to strengthen the antiviral properties of Onconase. Oncotarget 2017, 8, 11692–11707. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Raines, R.T. Ribonucleases as novel chemotherapeutics: The ranpirnase example. BioDrugs 2008, 22, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Luhtala, N.; Parker, R. T2 Family ribonucleases: Ancient enzymes with diverse roles. Trends Biochem. Sci. 2010, 35, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.K.; Roth, J.; Oppenheim, J.J.; Tracey, K.J.; Vogl, T.; Feldmann, M.; Horwood, N.; Nanchahal, J. Alarmins: Awaiting a clinical response. J. Clin. Investig. 2012, 122, 2711–2719. [Google Scholar] [CrossRef] [PubMed]
- Baranzini, N.; Pedrini, E.; Girardello, R.; Tettamanti, G.; de Eguileor, M.; Taramelli, R.; Acquati, F.; Grimaldi, A. Human recombinant RNASET2-induced inflammatory response and connective tissue remodeling in the medicinal leech. Cell Tissue Res. 2017, 368, 337–351. [Google Scholar] [CrossRef] [PubMed]
- Acquati, F.; Possati, L.; Ferrante, L.; Campomenosi, P.; Talevi, S.; Bardelli, S.; Margiotta, C.; Russo, A.; Bortoletto, E.; Rocchetti, R.; et al. Tumor and metastasis suppression by the human RNASET2 gene. Int. J. Oncol. 2005, 26, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Henneke, M.; Diekmann, S.; Ohlenbusch, A.; Kaiser, J.; Engelbrecht, V.; Kohlschutter, A.; Kratzner, R.; Madruga-Garrido, M.; Mayer, M.; Opitz, L.; et al. RNASET2-deficient cystic leukoencephalopathy resembles congenital cytomegalovirus brain infection. Nat. Genet. 2009, 41, 773–775. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, P.; Roiz, L.; Angelkovitch, B.; Schwartz, B.; Shoseyov, O. A recombinant human RNASET2 glycoprotein with antitumorigenic and antiangiogenic characteristics: Expression, purification, and characterization. Cancer 2006, 107, 2760–2769. [Google Scholar] [CrossRef] [PubMed]
- Steinfelder, S.; Andersen, J.F.; Cannons, J.L.; Feng, C.G.; Joshi, M.; Dwyer, D.; Caspar, P.; Schwartzberg, P.L.; Sher, A.; Jankovic, D. The major component in schistosome eggs responsible for conditioning dendritic cells for Th2 polarization is a T2 ribonuclease (omega-1). J. Exp. Med. 2009, 206, 1681–1690. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tischendorf, F.W.; Brattig, N.W.; Lintzel, M.; Buttner, D.W.; Burchard, G.D.; Bork, K.; Muller, M. Eosinophil granule proteins in serum and urine of patients with helminth infections and atopic dermatitis. Trop. Med. Int. Health 2000, 5, 898–905. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woschnagg, C.; Rubin, J.; Venge, P. Eosinophil cationic protein (ECP) is processed during secretion. J. Immunol. 2009, 183, 3949–3954. [Google Scholar] [CrossRef] [PubMed]
- Kjolvmark, C.; Akesson, P.; Pahlman, L.I. Urine concentrations of human beta-defensins and ribonuclease 7 in urinary tract infection and asymptomatic bacteriuria. Diagn. Microbiol. Infect. Dis. 2017, 89, 58–60. [Google Scholar] [CrossRef] [PubMed]
- Rucksaken, R.; Pairojkul, C.; Pinlaor, P.; Khuntikeo, N.; Roytrakul, S.; Selmi, C.; Pinlaor, S. Plasma autoantibodies against heat shock protein 70, enolase 1 and ribonuclease/angiogenin inhibitor 1 as potential biomarkers for cholangiocarcinoma. PLoS ONE 2014, 9, e103259. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.N.; Lee, H.H.; Chou, C.K.; Yang, W.H.; Wei, Y.; Chen, C.T.; Yao, J.; Hsu, J.L.; Zhu, C.; Ying, H.; et al. Angiogenin/Ribonuclease 5 Is an EGFR Ligand and a Serum Biomarker for Erlotinib Sensitivity in Pancreatic Cancer. Cancer Cell 2018, 33, 752–769. [Google Scholar] [CrossRef] [PubMed]
- Leland, P.A.; Raines, R.T. Cancer chemotherapy–ribonucleases to the rescue. Chem. Biol. 2001, 8, 405–413. [Google Scholar] [CrossRef]
- Goparaju, C.M.; Blasberg, J.D.; Volinia, S.; Palatini, J.; Ivanov, S.; Donington, J.S.; Croce, C.; Carbone, M.; Yang, H.; Pass, H.I. Onconase mediated NFKbeta downregulation in malignant pleural mesothelioma. Oncogene 2011, 30, 2767–2777. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.; Chen, C.; Jiang, H.; Qiu, Y.; Li, Y.; Li, X.; Zhang, X.; Liu, J.; Zhu, T. Ribonuclease attenuates hepatic ischemia reperfusion induced cognitive impairment through the inhibition of inflammatory cytokines in aged mice. Biomed. Pharmacother. 2017, 90, 62–68. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ribonuclease | Expression a | Immunomodulatory Functions and Activities |
|---|---|---|
| RNase 1 | pancreas, brain, gastrointestinal tract, endothelium, male reproductive tract, placenta, adipose tissue | Degrades extracellular RNA [21,22,23], stimulates dendritic cell maturation [24,25], antiviral in context of HIV-1 infection [21,26] |
| RNase 2 (EDN) | eosinophils, macrophages, dendritic cells, liver, lung, spleen | Alarmin [24,27,28], broad spectrum antiviral [16,18,27], dendritic cell chemotaxis and activation [24,28] |
| RNase 3 (ECP) | neutrophils, eosinophils | Antimicrobial [11], promotes bacterial cell agglutination [29,30], tissue remodeling [31,32], chemotaxis of fibroblasts [32,33,34] |
| RNase 4 | liver, lungs, monocytes, mast cells | Angiogenesis [35,36], antiviral [37], sequence specific RNA cleavage [38,39], antimicrobial [40,41], neuroprotective [35,36] |
| RNase 5 (angiogenin) | liver, mast cells, endothelium | Angiogenesis [42,43], hematopoiesis [44], inhibits neutrophil degranulation [45], modulates inflammatory response [46], possible antimicrobial activity [43,47,48,49], cleaves tRNA and upregulates rRNA transcription [50,51,52], induces tiRNA fragments with stress [53], neuroprotective [54], tissue remodeling [48] |
| RNase 6 | monocytes, neutrophils, lung, spleen | Antimicrobial [55,56], promotes bacterial cell agglutination [55], antiviral [57] |
| RNase 7 | skin, urinary tract, respiratory tract | Alarmin [58], antimicrobial [15,59,60,61], promotes inflammation and chemotaxis of immune cells [48,58,62], tissue remodeling [48,63] |
| RNase 8 | placenta, lung, liver, testes | Possible antimicrobial activity [17,63], possible role in amniotic cavity sterility [10,64] |
| RNase 9–13 | male reproductive tract | Modulates cytokine secretion [65], antimicrobial [66] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schwartz, L.; Cohen, A.; Thomas, J.; Spencer, J.D. The Immunomodulatory and Antimicrobial Properties of the Vertebrate Ribonuclease A Superfamily. Vaccines 2018, 6, 76. https://doi.org/10.3390/vaccines6040076
Schwartz L, Cohen A, Thomas J, Spencer JD. The Immunomodulatory and Antimicrobial Properties of the Vertebrate Ribonuclease A Superfamily. Vaccines. 2018; 6(4):76. https://doi.org/10.3390/vaccines6040076
Chicago/Turabian StyleSchwartz, Laura, Ariel Cohen, Jason Thomas, and John David Spencer. 2018. "The Immunomodulatory and Antimicrobial Properties of the Vertebrate Ribonuclease A Superfamily" Vaccines 6, no. 4: 76. https://doi.org/10.3390/vaccines6040076