

Malaria Vaccines: From the Past towards the mRNA Vaccine Era



Abstract
:1. Introduction
2. Vaccines That Target the Sporozoite Form of Plasmodium Parasite
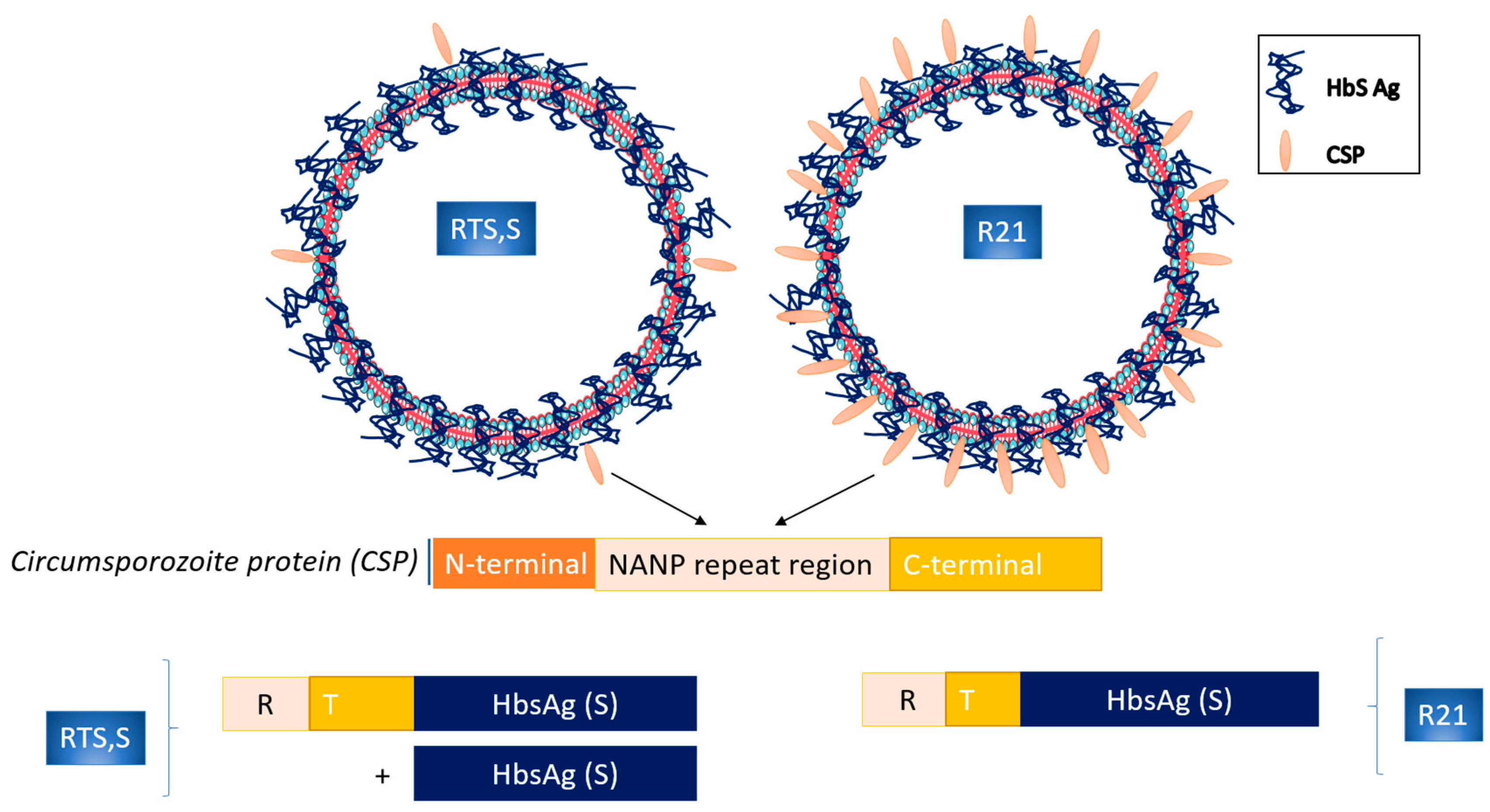
3. Circumsporozoite Protein Subunit Vaccines
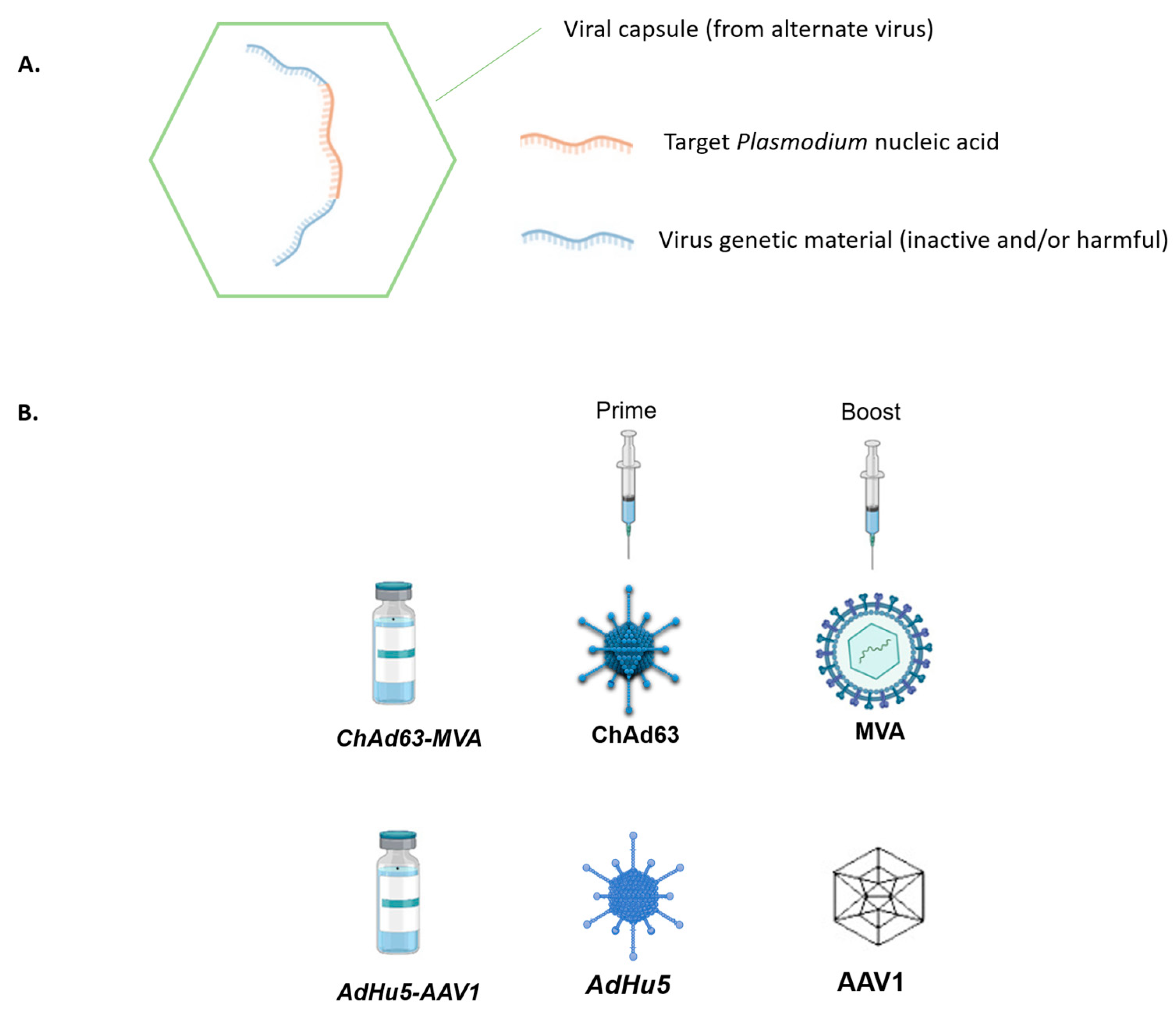
4. Viral-Vector-Vaccines
5. Erythrocytic Vaccines (Blood-Stage Vaccines)
6. Nucleic Acids in Malaria Vaccine Development
6.1. DNA Vaccines
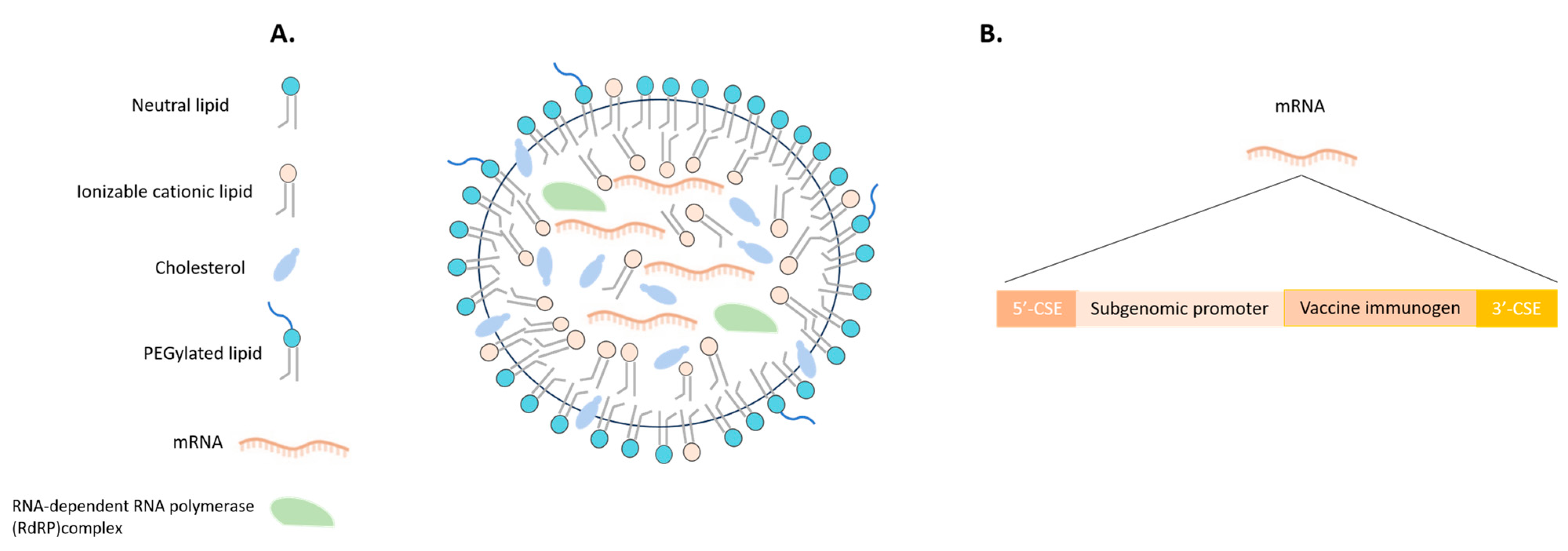
6.2. Malaria Vaccines in mRNA Era
7. Challenges and Future Perspectives in Malaria Vaccine Development
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Malaria. 2023. Available online: https://www.who.int/news-room/fact-sheets/detail/malaria (accessed on 21 July 2023).
- Centers for Disease Control and Prevention (CDC). Malaria. 2023. Available online: https://www.cdc.gov/malaria/about/faqs.html (accessed on 21 July 2023).
- Hoffman, S.L.; Vekemans, J.; Richie, T.L.; Duffy, P.E. The March toward Malaria Vaccines. Am. J. Prev. Med. 2015, 49, S319–S333. [Google Scholar] [CrossRef]
- Waghela, I.N.; Mallory, K.L.; Taylor, J.A.; Schneider, C.G.; Savransky, T.; Janse, C.J.; Lin, P.J.C.; Tam, Y.K.; Weissman, D.; Angov, E. Exploring in vitro expression and immune potency in mice using mRNA encoding the Plasmodium falciparum malaria antigen, CelTOS. Front. Immunol. 2022, 13, 1026052. [Google Scholar] [CrossRef]
- Tuteja, R. DNA vaccine against malaria: A long way to go. Crit. Rev. Biochem. Mol. Biol. 2002, 37, 29–54. [Google Scholar] [CrossRef]
- Laurens, M.B. Novel malaria vaccines. Hum. Vaccines Immunother. 2021, 17, 4549–4552. [Google Scholar] [CrossRef]
- Kurtovic, L.; Reiling, L.; Opi, D.H.; Beeson, J.G. Recent clinical trials inform the future for malaria vaccines. Commun. Med. 2021, 1, 26. [Google Scholar] [CrossRef]
- Richards, W.H.G. Active immunization of chicks against Plasmodium gallinaceum by inactivated homologous sporozoites and erythrocytic parasites. Nature 1966, 212, 1492–1494. [Google Scholar] [CrossRef]
- Hoffman, S.L.; Goh, L.M.L.; Luke, T.C.; Schneider, I.; Le, T.P.; Doolan, D.L.; Sacci, J.; De la Vega, P.; Dowler, M.; Paul, C.; et al. Protection of humans against malaria by immunization with radiation-attenuated Plasmodium falciparum sporozoites. J. Infect. Dis. 2002, 185, 1155–1164. [Google Scholar] [CrossRef]
- Epstein, J.E.; Paolino, K.M.; Richie, T.L.; Sedegah, M.; Singer, A.; Ruben, A.J.; Chakravarty, S.; Stafford, A.; Ruck, R.C.; Eappen, A.G.; et al. Protection against Plasmodium falciparum malaria by PfSPZ Vaccine. JCI Insight 2017, 2, e89154. [Google Scholar] [CrossRef]
- Seder, R.A.; Chang, L.J.; Enama, M.E.; Zephir, K.L.; Sarwar, U.N.; Gordon, I.J.; Holman, L.S.A.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; et al. Protection against malaria by intravenous immunization with a nonreplicating sporozoite vaccine. Science 2013, 341, 1359–1365. [Google Scholar] [CrossRef]
- Ishizuka, A.S.; Lyke, K.E.; DeZure, A.; Berry, A.A.; Richie, T.L.; Mendoza, F.H.; Enama, M.E.; Gordon, I.J.; Chang, L.J.; Sarwar, U.N.; et al. Protection against malaria at 1 year and immune correlates following PfSPZ vaccination. Nat. Med. 2016, 22, 614–623. [Google Scholar] [CrossRef]
- Oneko, M.; Steinhardt, L.C.; Yego, R.; Wiegand, R.E.; Swanson, P.A.; Kc, N.; Akach, D.; Sang, T.; Gutman, J.R.; Nzuu, E.L.; et al. Safety, immunogenicity and efficacy of PfSPZ Vaccine against malaria in infants in western Kenya: A double-blind, randomized, placebo-controlled phase 2 trial. Nat. Med. 2021, 27, 1636–1645. [Google Scholar] [CrossRef]
- Nkumama, I.N.; Osier, F.H.A. Malaria vaccine roller coaster. Nat. Microbiol. 2021, 6, 1345–1346. [Google Scholar] [CrossRef]
- Sissoko, M.S.; Healy, S.A.; Katile, A.; Omaswa, F.; Zaidi, I.; Gabriel, E.E.; Kamate, B.; Samake, Y.; Guindo, M.A.; Dolo, A.; et al. Safety and efficacy of PfSPZ Vaccine against Plasmodium falciparum via direct venous inoculation in healthy malaria-exposed adults in Mali: A randomised, double-blind phase 1 trial. Lancet Infect. Dis. 2017, 17, 498–509. [Google Scholar] [CrossRef]
- Jongo, S.A.; Preston Church, L.W.; Mtoro, A.T.; Schindler, T.; Chakravarty, S.; Ruben, A.J.; Swanson, P.A.; Kassim, K.R.; Mpina, M.; Tumbo, A.M.; et al. Increase of dose associated with decrease in protection against controlled human malaria infection by PfSPZ vaccine in Tanzanian Adults. Clin. Infect. Dis. 2020, 71, 2849–2857. [Google Scholar] [CrossRef]
- Sirima, S.B.; Ouédraogo, A.; Tiono, A.B.; Kaboré, J.M.; Bougouma, E.C.; Ouattara, M.S.; Kargougou, D.; Diarra, A.; Henry, N.; Ouédraogo, I.N.; et al. A randomized controlled trial showing safety and efficacy of a whole sporozoite vaccine against endemic malaria. Sci. Transl. Med. 2022, 14, eabj3776. [Google Scholar] [CrossRef]
- Bijker, E.M.; Bastiaens, G.J.H.; Teirlinck, A.C.; Van Gemert, G.J.; Graumans, W.; Van De Vegte-Bolmer, M.; Siebelink-Stoter, R.; Arens, T.; Teelen, K.; Nahrendorf, W.; et al. Protection against malaria after immunization by chloroquine prophylaxis and sporozoites is mediated by preerythrocytic immunity. Proc. Natl. Acad. Sci. USA 2013, 110, 7862–7867. [Google Scholar] [CrossRef]
- Lobel, H.O.; Campbell, C.C.; Hightower, A.H.; Eng, T.; Miani, M.; Eng, T.; Bernard, K.W. Long-term malaria prophylaxis with weekly mefloquine. Lancet 1993, 341, 848–851. [Google Scholar] [CrossRef]
- Coulibaly, D.; Kone, A.K.; Traore, K.; Niangaly, A.; Kouriba, B.; Arama, C.; Zeguime, A.; Dolo, A.; Lyke, K.E.; Plowe, C.V.; et al. PfSPZ-CVac malaria vaccine demonstrates safety among malaria-experienced adults: A randomized, controlled phase 1 trial. eClinicalMedicine 2022, 52, 101579. [Google Scholar] [CrossRef]
- Richie, T.; Billingsley, P.; Epstein, J.E.; Lyke, K.E.; Mordmüller, B.; Alonso, P.; Kremsner, P.G.; Seder, R.A.; Hoffman, S.L. Progress with P. falciparum sporozoite based malaria vaccines. Vaccine 2015, 33, 7452–7461. [Google Scholar] [CrossRef]
- Ibanez, J.; Fendel, R.; Lorenz, F.R.; Granados-Bayon, P.; Brückner, S.; Esen, M.; Sulyok, M.; Sulyok, Z.; Borrmann, S.; Bacher, P.; et al. Efficacy, T cell activation and antibody responses in accelerated Plasmodium falciparum sporozoite chemoprophylaxis vaccine regimens. NPJ Vaccines 2022, 7, 59. [Google Scholar] [CrossRef]
- Vaughan, A.M.; Kappe, S.H.I. Genetically attenuated malaria parasites as vaccines. Expert Rev. Vaccines 2017, 16, 765–767. [Google Scholar] [CrossRef] [PubMed]
- Kublin, J.G.; Mikolajczak, S.A.; Sack, B.K.; Fishbaugher, M.E.; Seilie, A.; Shelton, L.; VonGoedert, T.; Firat, M.; Magee, S.; Fritzen, E.; et al. Complete attenuation of genetically engineered Plasmodium falciparum sporozoites in human subjects. Sci. Transl. Med. 2017, 9, eaad9099. [Google Scholar] [CrossRef] [PubMed]
- Murphy, S.C.; Vaughan, A.M.; Kublin, J.G.; Fishbauger, M.; Seilie, A.M.; Cruz, K.P.; Mankowski, T.; Firat, M.; Magee, S.; Betz, W.; et al. A genetically-engineered Plasmodium falciparum parasite vaccine provides protection from controlled human malaria infection. Sci. Transl. Med. 2022, 14, eabn9709. [Google Scholar] [CrossRef]
- Roestenberg, M.; Walk, J.; Van Der Boor, S.C.; Langenberg, M.C.C.; Hoogerwerf, M.A.; Janse, J.J.; Manurung, M.; Yap, X.Z.; García, A.F.; Koopman, J.P.R.; et al. A double-blind, placebo-controlled phase 1/2a trial of the genetically attenuated malaria vaccine PfSPZ-GA1. Sci. Transl. Med. 2020, 12, eaaz5629. [Google Scholar] [CrossRef]
- El-Moamly, A.A.; El-Sweify, M.A. Malaria vaccines: The 60-year journey of hope and final success—lessons learned and future prospects. Trop. Med. Health 2023, 51, 29. [Google Scholar] [CrossRef]
- Cockburn, I.A.; Seder, R.A. Malaria prevention: From immunological concepts to effective vaccines and protective antibodies. Nat. Immunol. 2018, 19, 1199–1211. [Google Scholar] [CrossRef] [PubMed]
- Mandala, W.L.; Harawa, V.; Dzinjalamala, F.; Tembo, D. The role of different components of the immune system against Plasmodium falciparum malaria: Possible contribution towards malaria vaccine development. Mol. Biochem. Parasitol. 2021, 246, 111425. [Google Scholar] [CrossRef] [PubMed]
- The RTS,S Clinical Trials Partnership. First Results of Phase 3 Trial of RTS,S/AS01 Malaria Vaccine in African Children. N. Engl. J. Med. 2011, 365, 1863–1875. [Google Scholar] [CrossRef]
- Gross, M. Magazine New beginnings for malaria research. Curr. Biol. 2023, 33, R203–R205. [Google Scholar] [CrossRef]
- Samuels, A.M.; Ansong, D.; Kariuki, S.K.; Adjei, S.; Bollaerts, A.; Ockenhouse, C.; Westercamp, N.; Lee, C.K.; Schuerman, L.; Bii, D.K.; et al. Efficacy of RTS,S/AS01E malaria vaccine administered according to different full, fractional, and delayed third or early fourth dose regimens in children aged 5–17 months in Ghana and Kenya: An open-label, phase 2b, randomised controlled trial. Lancet Infect. Dis. 2022, 22, 1329–1342. [Google Scholar] [CrossRef]
- Collins, K.A.; Snaith, R.; Cottingham, M.G.; Gilbert, S.C.; Hill, A.V.S. Enhancing protective immunity to malaria with a highly immunogenic virus-like particle vaccine. Sci. Rep. 2017, 7, srep46621. [Google Scholar] [CrossRef] [PubMed]
- Movin Abeywickrema, A.L.G. COVID-19 vaccine results might inform malaria vaccine strategies. Lancet Infect. Dis. 2022, 22, 440–441. [Google Scholar] [CrossRef]
- Datoo, M.S.; Natama, M.H.; Somé, A.; Traoré, O.; Rouamba, T.; Bellamy, D.; Yameogo, P.; Valia, D.; Tegneri, M.; Ouedraogo, F.; et al. Efficacy of a low-dose candidate malaria vaccine, R21 in adjuvant Matrix-M, with seasonal administration to children in Burkina Faso: A randomised controlled trial. Lancet 2021, 397, 1809–1818. [Google Scholar] [CrossRef] [PubMed]
- Datoo, M.S.; Natama, H.M.; Somé, A.; Bellamy, D.; Traoré, O.; Rouamba, T.; Tahita, M.C.; Ido, N.F.A.; Yameogo, P.; Valia, D.; et al. Efficacy and immunogenicity of R21/Matrix-M vaccine against clinical malaria after 2 years’ follow-up in children in Burkina Faso: A phase 1/2b randomised controlled trial. Lancet Infect. Dis. 2022, 22, 1728–1736. [Google Scholar] [CrossRef] [PubMed]
- Hill, A.V.S. Vaccines against malaria. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 2806–2814. [Google Scholar] [CrossRef]
- Ogwang, C.; Kimani, D.; Edwards, N.J.; Roberts, R.; Mwacharo, J.; Bowyer, G.; Bliss, C.; Hodgson, S.H.; Njuguna, P.; Viebig, N.K.; et al. Prime-boost vaccination with chimpanzee adenovirus and modified vaccinia Ankara encoding TRAP provides partial protection against Plasmodium falciparum infection in Kenyan adults. Sci. Transl. Med. 2015, 7, 286re5. [Google Scholar] [CrossRef]
- Yusuf, Y.; Yoshii, T.; Iyori, M.; Mizukami, H.; Fukumoto, S.; Yamamoto, D.S.; Bin Emran, T.; Amelia, F.; Islam, A.; Syafira, I.; et al. A viral-vectored multi-stage malaria vaccine regimen with protective and transmission-blocking efficacies. Front. Immunol. 2019, 10, 2412. [Google Scholar] [CrossRef]
- Moorthy, V.S.; Good, M.F.; Hill, A.V.S. Malaria vaccine developments. Lancet 2004, 363, 150–156. [Google Scholar] [CrossRef]
- Palacpac, N.M.Q.; Ntege, E.; Yeka, A.; Balikagala, B.; Suzuki, N.; Shirai, H.; Yagi, M.; Ito, K.; Fukushima, W.; Hirota, Y.; et al. Phase 1b Randomized Trial and Follow-Up Study in Uganda of the Blood-Stage Malaria Vaccine Candidate BK-SE36. PLoS ONE 2013, 8, e64073. [Google Scholar] [CrossRef]
- Bougouma, E.C.; Palacpac, N.M.Q.; Tiono, A.B.; Nebie, I.; Ouédraogo, A.; Houard, S.; Yagi, M.; Coulibaly, S.A.; Diarra, A.; Tougan, T.; et al. Safety and immunogenicity of BK-SE36 in a blinded, randomized, controlled, age de-escalating phase Ib clinical trial in Burkinabe children. Front. Immunol. 2022, 13, 978591. [Google Scholar] [CrossRef]
- Pirahmadi, S.; Afzali, S.; Zargar, M.; Zakeri, S.; Mehrizi, A.A. How can we develop an effective subunit vaccine to achieve successful malaria eradication? Microb. Pathog. 2021, 160, 105203. [Google Scholar] [CrossRef] [PubMed]
- Dolgin, E. How COVID unlocked the power of RNA vaccines. Nature 2021, 589, 189–191. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, J.B.; Mansoura, M.K.; Geall, A.J. Vaccines “on demand”: Science fiction or a future reality. Expert Opin. Drug Discov. 2015, 10, 101–106. [Google Scholar] [CrossRef] [PubMed]
- De Groot, A.S.; Einck, L.; Moise, L.; Chambers, M.; Ballantyne, J.; Malone, R.W.; Ardito, M.; Martin, W. Making vaccines “on demand”. Hum. Vaccin. Immunother. 2013, 9, 1877–1884. [Google Scholar] [CrossRef] [PubMed]
- Bacon, D.J.; Sedegah, M. Reduced production of RNA transcripts from individual DNA plasmids given in a multivalent DNA vaccine formula. Hum. Vaccin. 2007, 3, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Dobaño, C.; Sedegah, M.; Rogers, W.O.; Kumar, S.; Zheng, H.; Hoffman, S.L.; Doolan, D.L. Plasmodium: Mammalian codon optimization of malaria plasmid DNA vaccines enhances antibody responses but not T cell responses nor protective immunity. Exp. Parasitol. 2009, 122, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Nagata, T.; Uchijima, M.; Yoshida, A.; Kawashima, M.; Koide, Y. Codon optimization effect on translational efficiency of DNA vaccine in mammalian cells: Analysis of plasmid DNA encoding a CTL epitope derived from microorganisms. Biochem. Biophys. Res. Commun. 1999, 261, 445–451. [Google Scholar] [CrossRef]
- Reddy, G.R.; Chakrabarti, D.; Schuster, S.M.; Ferl, R.J.; Almira, E.C.; Dame, J.B. Gene sequence tags from Plasmodium falciparum genomic DNA fragments prepared by the “genease” activity of mung bean nuclease. Proc. Natl. Acad. Sci. USA 1993, 90, 9867–9871. [Google Scholar] [CrossRef]
- Wang, R.; Epstein, J.; Charoenvit, Y.; Baraceros, F.M.; Rahardjo, N.; Gay, T.; Banania, J.-G.; Chattopadhyay, R.; de la Vega, P.; Richie, T.L.; et al. Induction in Humans of CD8+ and CD4+ T Cell and Antibody Responses by Sequential Immunization with Malaria DNA and Recombinant Protein. J. Immunol. 2004, 172, 5561–5569. [Google Scholar] [CrossRef]
- Yoshida, S.; Kashiwamura, S.I.; Hosoya, Y.; Luo, E.; Matsuoka, H.; Ishii, A.; Fujimura, A.; Kobayashi, E. Direct immunization of malaria DNA vaccine into the liver by gene gun protects against lethal challenge of Plasmodium berghei sporozoite. Biochem. Biophys. Res. Commun. 2000, 271, 107–115. [Google Scholar] [CrossRef]
- Wang, L.; Geng, C.; Jiang, L.; Gong, D.; Liu, D.; Yoshimura, H.; Zhong, L. The anti-atherosclerotic effect of olive leaf extract is related to suppressed inflammatory response in rabbits with experimental atherosclerosis. Eur. J. Nutr. 2008, 47, 235–243. [Google Scholar] [CrossRef] [PubMed]
- Datta, D.; Bansal, G.P.; Kumar, R.; Ellefsen, B.; Hannaman, D.; Kumar, N. Evaluation of the impact of codon optimization and N-linked glycosylation on functional immunogenicity of Pfs25 DNA vaccines delivered by in vivo electroporation in preclinical Studies in mice. Clin. Vaccine Immunol. 2015, 22, 1013–1019. [Google Scholar] [CrossRef] [PubMed]
- Shuaibu, M.N.; Kikuchi, M.; Cherif, M.S.; Helegbe, G.K.; Yanagi, T.; Hirayama, K. Selection and identification of malaria vaccine target molecule using bioinformatics and DNA vaccination. Vaccine 2010, 28, 6868–6875. [Google Scholar] [CrossRef]
- Fotoran, W.L.; da Silva, J.R.; Glitz, C.; Ferreira, L.C.d.S.; Wunderlich, G. Establishment of an Antiplasmodial Vaccine Based on PfRH5-Encoding RNA Replicons Stabilized by Cationic Liposomes. Pharmaceutics 2023, 15, 1223. [Google Scholar] [CrossRef] [PubMed]
- Kaslow, D.C. A potential disruptive technology in vaccine development: Gene-based vaccines and their application to infectious diseases. Trans. R. Soc. Trop. Med. Hyg. 2004, 98, 593–601. [Google Scholar] [CrossRef] [PubMed]
- Smerdou, C.; Liljeström, P. Non-viral amplification systems for gene transfer: Vectors based on alphaviruses. Curr. Opin. Mol. Ther. 1999, 1, 244–251. [Google Scholar]
- Mallory, K.L.; Taylor, J.A.; Zou, X.; Waghela, I.N.; Schneider, C.G.; Sibilo, M.Q.; Punde, N.M.; Perazzo, L.C.; Savransky, T.; Sedegah, M.; et al. Messenger RNA expressing PfCSP induces functional, protective immune responses against malaria in mice. NPJ Vaccines 2021, 6, 84. [Google Scholar] [CrossRef]
- Garcia, A.B.; Siu, E.; Sun, T.; Exler, V.; Brito, L.; Hekele, A.; Otten, G.; Augustijn, K.; Janse, C.J.; Ulmer, J.B.; et al. Neutralization of the Plasmodium-encoded MIF ortholog confers protective immunity against malaria infection. Nat. Commun. 2018, 9, 2714. [Google Scholar] [CrossRef]
- Hayashi, C.T.H.; Cao, Y.; Clark, L.C.; Tripathi, A.K.; Zavala, F.; Dwivedi, G.; Knox, J.; Alameh, M.G.; Lin, P.J.C.; Tam, Y.K.; et al. mRNA-LNP expressing PfCSP and Pfs25 vaccine candidates targeting infection and transmission of Plasmodium falciparum. NPJ Vaccines 2022, 7, 155. [Google Scholar] [CrossRef]
- Bergmann-Leitner, E.S.; Chaudhury, S.; Steers, N.J.; Sabato, M.; Delvecchio, V.; Wallqvist, A.S.; Ockenhouse, C.F.; Angov, E. Computational and Experimental Validation of B and T-Cell Epitopes of the In Vivo Immune Response to a Novel Malarial Antigen. PLoS ONE 2013, 8, e71610. [Google Scholar] [CrossRef]
- Sedegah, M.; Hollingdale, M.R.; Ganeshan, H.; Belmonte, M.; Huang, J.; Belmonte, A.; Inoue, S.; Velasco, R.; Hickey, B.; Teneza-Mora, N.; et al. IMRAS-Immunization with radiationattenuated Plasmodium falciparum sporozoites by mosquito bite: Cellular immunity to sporozoites, CSP, AMA1, TRAP and CelTOS. PLoS ONE 2021, 16, e0256396. [Google Scholar] [CrossRef] [PubMed]
- Bergmann-Leitner, E.S.; Mease, R.M.; de la Vega, P.; Savranskaya, T.; Polhemus, M.; Ockenhouse, C.; Angov, E. Immunization with pre-erythrocytic antigen CelTOS from plasmodium falciparum elicits cross-species protection against heterologous challenge with Plasmodium berghei. PLoS ONE 2010, 5, e12294. [Google Scholar] [CrossRef] [PubMed]
- Bergmann-Leitner, E.S.; Legler, P.M.; Savranskaya, T.; Ockenhouse, C.F.; Angov, E. Cellular and humoral immune effector mechanisms required for sterile protection against sporozoite challenge induced with the novel malaria vaccine candidate CelTOS. Vaccine 2011, 29, 5940–5949. [Google Scholar] [CrossRef]
- Espinosa, D.A.; Vega-Rodriguez, J.; Flores-Garcia, Y.; Noe, A.R.; Muñoz, C.; Coleman, R.; Bruck, T.; Haney, K.; Stevens, A.; Retallack, D.; et al. The Plasmodium falciparum cell-traversal protein for ookinetes and sporozoites as a candidate for preerythrocytic and transmission-blocking vaccines. Infect. Immun. 2017, 85, e00498-16. [Google Scholar] [CrossRef]
- Chuang, Y.M.; Alameh, M.G.; Abouneameh, S.; Raduwan, H.; Ledizet, M.; Weissman, D.; Fikrig, E. A mosquito AgTRIO mRNA vaccine contributes to immunity against malaria. NPJ Vaccines 2023, 8, 88. [Google Scholar] [CrossRef] [PubMed]
- Ganley, M.; Holz, L.E.; Minnell, J.J.; de Menezes, M.N.; Burn, O.K.; Poa, K.C.Y.; Draper, S.L.; English, K.; Chan, S.T.S.; Anderson, R.J.; et al. mRNA vaccine against malaria tailored for liver-resident memory T cells. Nat. Immunol. 2023, 24, 1487–1498. [Google Scholar] [CrossRef]
- Raj, D.K.; Das Mohapatra, A.; Jnawali, A.; Zuromski, J.; Jha, A.; Cham-Kpu, G.; Sherman, B.; Rudlaff, R.M.; Nixon, C.E.; Hilton, N.; et al. Anti-PfGARP activates programmed cell death of parasites and reduces severe malaria. Nature 2020, 582, 104–108. [Google Scholar] [CrossRef]
- Chen, W. Will the mRNA vaccine platform be the panacea for the development of vaccines against antimicrobial resistant (AMR) pathogens? Expert Rev. Vaccines 2022, 21, 155–157. [Google Scholar] [CrossRef]
- Chen, J.; Chen, J.; Xu, Q. Current Developments and Challenges of mRNA Vaccines. Annu. Rev. Biomed. Eng. 2022, 24, 85–109. [Google Scholar] [CrossRef]
- Chung, S.; Lee, C.M.; Zhang, M. Advances in nanoparticle-based mRNA delivery for liver cancer and liver-associated infectious diseases. Nanoscale Horizons 2022, 8, 10–28. [Google Scholar] [CrossRef]
- Chaudhary, N.; Weissman, D.; Whitehead, K.A. mRNA vaccines for infectious diseases: Principles, delivery and clinical translation. Nat. Rev. Drug Discov. 2021, 20, 817–838. [Google Scholar] [CrossRef] [PubMed]
- Safety and Immune Responses after Vaccination With an Investigational RNA-based Vaccine Against Malaria. ClinicalTrials.gov, Identifier: NCT05581641. Available online: https://classic.clinicaltrials.gov/ct2/show/NCT05581641?term=BNT165-01&draw=2&rank=1 (accessed on 18 August 2023).




{kind=link}
{kind=link}
{kind=link}
| Vaccine | Company | Target | Source of Peptide Antigens | Adjuvant | Immune Response | Clinical Status |
|---|---|---|---|---|---|---|
| RTS,S/AS01Ε | GlaxoSmithKline Biologicals SA | Inhibition of sporozoite infection | Circumsporozoite protein | AS01 | Protective humoral/cellular | WHO recommended |
| R21 | Serum Institute of India (Pune, India) | Inhibition of sporozoite infection | Circumsporozoite protein | Matrix-M | Protective humoral | Phase III clinical trial |
| PfSPZ | Sanaria Inc. | Killing of infected hepatocytes | Whole sporozoite | None | Protective humoral/cellular | Phase II clinical trial |
| PfSPZ-CVac (CVac = Chemoprophylaxis Vaccine) | Sanaria Inc. | Killing of infected hepatocytes | Whole sporozoite | None | Protective humoral/cellular | Phase II clinical trial |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsoumani, M.E.; Voyiatzaki, C.; Efstathiou, A. Malaria Vaccines: From the Past towards the mRNA Vaccine Era. Vaccines 2023, 11, 1452. https://doi.org/10.3390/vaccines11091452
Tsoumani ME, Voyiatzaki C, Efstathiou A. Malaria Vaccines: From the Past towards the mRNA Vaccine Era. Vaccines. 2023; 11(9):1452. https://doi.org/10.3390/vaccines11091452
Chicago/Turabian StyleTsoumani, Maria E., Chrysa Voyiatzaki, and Antonia Efstathiou. 2023. "Malaria Vaccines: From the Past towards the mRNA Vaccine Era" Vaccines 11, no. 9: 1452. https://doi.org/10.3390/vaccines11091452