

Nanoparticle-Based Adjuvants and Delivery Systems for Modern Vaccines
 , , , and
, , , and
Abstract
:
1. Introduction
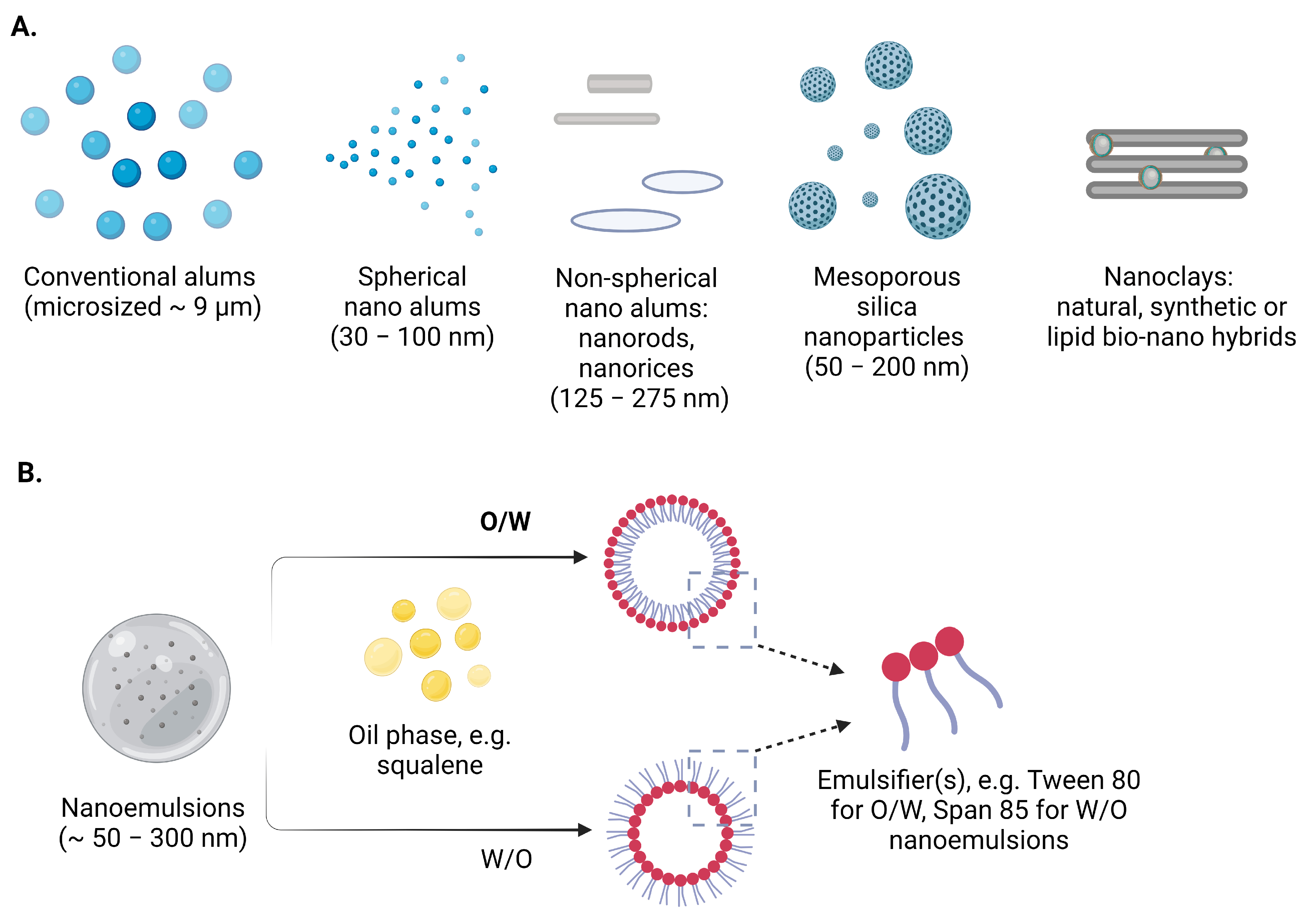
2. Nanoadjuvants
2.1. Inorganic Nanomaterials as Vaccine Adjuvants and/or Carriers
2.1.1. Conventional Alum Adjuvants
2.1.2. Nano Alum Adjuvants
2.1.3. Silica Nanoparticles
2.1.4. Clay Nanoparticles
3. Nanoemulsions
4. Virus-Based Nanocarriers as Vaccine Delivery Systems
4.1. Viral-Vectored-Based Delivery Systems
4.1.1. Adenoviral Vector-Based Delivery System
4.1.2. Adeno-Associated Viral Vector as a Delivery System for Vaccines
4.1.3. Vesicular Stomatitis Viral Vector for Vaccines
4.1.4. Lentiviral Vector for Vaccines
4.2. Virus-like Particles and Virosomes
4.2.1. Virus-like Particles (VLPs)
Bacteriophage-Based VLPs
4.2.2. Virosomes
5. Conclusions

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Nanoadjuvant/Nanoparticulate Delivery System | Effect | Safety | Immune Target | Examples of Clinical Use | References | |
|---|---|---|---|---|---|---|
| Inorganic nanomaterials | Nano-alum | NLRP3 inflammasome activation | Generally deemed safe Local adverse reactions (subcutaneous granuloma and other local inflammatory reactions) | Th1 | / | [34,41,42,43] |
| Silica nanoparticles | Delivery system with controlled antigen release | Safety is strongly dependent on preparation technique, physicochemical properties and administration route | Th1/Th2 Th17 | / | [51,52,53,54,55,56,57,58] | |
| Clay nanoparticles | Depot effect | High immunogenicity; much higher compared to that of the FDA-approved adjuvants (e.g., Alum) | Th1 | / | [64,67] | |
| Lipid-based nanoadjuvants | Nanoemulsions | Sustained antigen release TLR4 ligand | O/W nanoemulsions (e.g., MF59®, AS03® and AF03®) are less reactogenic and less toxic compared to W/O (e.g., Montanide®) nanoemulsions Narcolepsy (ASO3® ) | Th1/Th2 | Vaccines against seasonal influenza, pandemic influenza and avian influenza; therapeutic vaccines against lung cancer and melanoma | [18,76,77,78,79,80,82,83,84,91] |
| Lipid nanoparticles | Antigen delivery system (especially for mRNA) Antigen protection from degradation | Highly dependent on lipid components Local reactions and the injection site | Th1/Th2 CD8+ T cells | Vaccines against SARS-CoV-2 | [107,108,109,169] | |
| Virus-based nanoparticles | Adenoviral vector | Antigen delivery system | Local reactions and the injection site Fatigue, fever and headache Rare cases of thrombotic thrombocytopaenia | Th1/ Th2 CD8+ T cells | Vaccines against SARS-CoV-2 and Ebola | [118,121,129] |
| Adeno-associated viral vector | Delivery system for antigens as well as for monoclonal antibodies and derivatives of immunoglobulins for passive immunization | Generally safe, but with low immunogenicity Transient and asymptomatic hepatitis | Th1/ Th2 CD8+ T cells | / | [118,133,134,170] | |
| Vesicular stomatitis viral vector | Delivery system for antigens | Fatigue, fever and headache | Th1/ Th2 CD8+ T cells | Vaccine against Ebola virus | [135,136,137,171] | |
| Lentiviral vector | Delivery system for antigens | Safety is still under evaluation | Th1/ Th2 CD8+ T cells | / | [138,172] | |
| Virus-like particles (VLPs) | Delivery system for antigens | Improved safety compared to viral-vectored vaccines Generally recognized as safe | Th1/ Th2 CD8+ T cells | Vaccines against hepatitis B virus, vaccines against human papilloma viruses (HPV), vaccine against hepatitis E virus and vaccine against malaria | [120,155,156] | |
| Virosomes | Delivery system for antigens | Improved safety profile compared to VLPs | Th1/Th2 CD8+ T cells | Vaccines against influenza and hepatitis A viruses | [167,168] | |
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Lemoine, C.; Thakur, A.; Krajišnik, D.; Guyon, R.; Longet, S.; Razim, A.; Górska, S.; Pantelic, I.; Ilic, T.; Nikolic, I.; et al. Technological Approaches for Improving Vaccination Compliance and Coverage. Vaccines 2020, 8, 304. [Google Scholar] [CrossRef] [PubMed]
- Candela, F.; Quarta, E.; Buttini, F.; Ancona, A.; Bettini, R.; Sonvico, F. Recent Patents on Nasal Vaccines Containing Nanoadjuvants. Recent. Adv. Drug. Deliv. 2022, 16, 103–121. [Google Scholar] [CrossRef]
- Zhang, N.; Li, M.; Hou, Z.; Ma, L.; Younas, A.; Wang, Z.; Jiang, X.; Gao, J. From vaccines to nanovaccines: A promising strategy to revolutionize rheumatoid arthritis treatment. J. Control. Release 2022, 350, 107–121. [Google Scholar] [CrossRef] [PubMed]
- Etter, E.; Mei, K.-C.; Nguyen, J. Delivering more for less: Nanosized, minimal-carrier and pharmacoactive drug delivery systems. Adv. Drug. Deliv. Rev. 2021, 179, 113994. [Google Scholar] [CrossRef]
- Yuba, E.; Kado, Y.; Kasho, N.; Harada, A. Cationic lipid potentiated the adjuvanticity of polysaccharide derivative-modified liposome vaccines. J. Control. Release, 2022; Online ahead of print. [Google Scholar] [CrossRef]
- Bi, S.; Li, M.; Liang, Z.; Li, G.; Yu, G.; Zhang, J.; Chen, C.; Yang, C.; Xue, C.; Zuo, Y.Y.; et al. Self-assembled aluminum oxyhydroxide nanorices with superior suspension stability for vaccine adjuvant. J. Colloid. Interface Sci. 2022, 627, 238–246. [Google Scholar] [CrossRef]
- Carmona-Ribeiro, A.M. Supramolecular Nanostructures for Vaccines. Biomimetics 2021, 7, 6. [Google Scholar] [CrossRef]
- Wack, A.; Rappuoli, R. Vaccinology at the beginning of the 21st century. Curr. Opin. Immunol. 2005, 17, 411–418. [Google Scholar] [CrossRef]
- Reed, S.G.; Bertholet, S.; Coler, R.N.; Friede, M. New horizons in adjuvants for vaccine development. Trends Immunol. 2009, 30, 23–32. [Google Scholar] [CrossRef]
- Ruterbusch, M.; Pruner, K.B.; Shehata, L.; Pepper, M. In Vivo CD4+ T Cell Differentiation and Function: Revisiting the Th1/Th2 Paradigm. Annu. Rev. Immunol. 2020, 38, 705–725. [Google Scholar] [CrossRef] [PubMed]
- Rapaka, R.R.; Cross, A.S.; McArthur, M.A. Using Adjuvants to Drive T Cell Responses for Next-Generation Infectious Disease Vaccines. Vaccines 2021, 9, 820. [Google Scholar] [CrossRef]
- Verma, S.K.; Mahajan, P.; Singh, N.K.; Gupta, A.; Aggarwal, R.; Rappuoli, R.; Johri, A.K. New-age vaccine adjuvants, their development, and future perspective. Front. Immunol. 2023, 14, 1043109. [Google Scholar] [CrossRef]
- Pulendran, B.S.; Arunachalam, P.; O’Hagan, D.T. Emerging concepts in the science of vaccine adjuvants. Nat. Rev. Drug. Discov. 2021, 20, 454–475. [Google Scholar] [CrossRef] [PubMed]
- Ojha, S.K.; Pattnaik, R.; Singh, P.K.; Dixit, S.; Mishra, S.; Pal, S.; Kumar, S. Virus as a Nanocarrier for Drug Delivery Redefining Medical Therapeutics—A Status Report. Comb. Chem. High. Throughput Screen 2022, 25, 1619–1629. [Google Scholar] [CrossRef] [PubMed]
- Mamo, T.; Poland, G.A. Nanovaccinology: The next generation of vaccines meets 21st century materials science and engineering. Vaccine 2012, 30, 6609–6611. [Google Scholar] [CrossRef]
- Yasamineh, S.; Kalajahi, H.G.; Yasamineh, P.; Yazdani, Y.; Gholizadeh, O.; Tabatabaie, R.; Afkhami, H.; Davodabadi, F.; Farkhad, A.K.; Pahlevan, D.; et al. An overview on nanoparticle-based strategies to fight viral infections with a focus on COVID-19. J. Nanobiotechnol. 2022, 20, 440. [Google Scholar] [CrossRef] [PubMed]
- Lindblad, E.B.; Duroux, L. Immunopotentiators in Modern Vaccines; Schijns, V., O’Hagan, D., Eds.; Academic Press: Cambridge, MA, USA, 2017; Chapter 18; p. 347. [Google Scholar]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- Hem, S.L.; Hogen Esch, H. Vaccine Adjuvants and Delivery Systems; Singh, M., Ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2007; pp. 81–114. [Google Scholar]
- Hem, S.L.; Hogenesch, H. Relationship between physical and chemical properties of aluminum-containing adjuvants and immunopotentiation. Expert. Rev. Vaccines 2007, 6, 685–698. [Google Scholar] [CrossRef]
- Sheskey, P.J.; Cook, W.G.; Cable, C.T. Handbook of Pharmaceutical Excipients, 8th ed.; Pharmaceutical Press: London, UK, 2017; p. 67. [Google Scholar]
- Garçon, N.; Friede, M. Plotkin’s Vaccines; Plotkin, S.A., Orenstein, W.A., Offit, P.A., Edwards, K.M., Eds.; Elsevier: Philadelphia, PA, USA, 2018; Section 1; Chapter 6; pp. 61–74. [Google Scholar]
- Caulfield, M.J.; Shi, L.; Wang, S.; Wang, B.; Tobery, T.W.; Mach, H.; Ahl, P.L.; Cannon, J.L.; Cook, J.C.; Heinrichs, J.H.; et al. Effect of alternative aluminum adjuvants on the absorption and immunogenicity of HPV16 L1 VLPs in mice. Hum. Vaccines 2007, 3, 139–145. [Google Scholar] [CrossRef]
- HogenEsch, H.; O’Hagan, D.T.; Fox, C.B. Optimizing the utilization of aluminum adjuvants in vaccines: You might just get what you want. NJP Vaccines 2018, 3, 51. [Google Scholar] [CrossRef] [Green Version]
- Morefield, G.L. A rational, systematic approach for the development of vaccine formulations. AAPS J. 2011, 13, 191–200. [Google Scholar] [CrossRef] [Green Version]
- Krajišnik, D.; Iliƒá, T.; Nikoliƒá, I.; Saviƒá, S. Established and advanced adjuvants in vaccines’ formulation: Mineral adsorbents, nanoparticulate carriers and microneedle delivery systems. Arh. Farm. 2019, 69, 420–451. [Google Scholar] [CrossRef]
- Galazka, A.M.; Milstien, J.B.; Zaffran, M. Thermostability of Vaccines; World Health Organization: Geneva, Switzerland, 1998; Available online: https://apps.who.int/iris/handle/10665/64980 (accessed on 20 April 2023).
- Maa, Y.F.; Zhao, L.; Payne, L.G.; Chen, D. Stabilization of alum-adjuvanted vaccine dry powder formulations: Mechanism and application. J. Pharm. Sci. 2003, 92, 319–332. [Google Scholar] [CrossRef] [PubMed]
- Clapp, T.; Siebert, P.; Chen, D.; Jones Braun, L. Vaccines with aluminum-containing adjuvants: Optimizing vaccine efficacy and thermal stability. J. Pharm. Sci. 2011, 100, 388–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tonnis, W.F.; Amorij, J.P.; Vreeman, M.A.; Frijlink, H.W.; Kersten, G.F.; Hinrichs, W.L. Improved storage stability and immunogenicity of hepatitis B vaccine after spray-freeze drying in presence of sugars. Eur. J. Pharm. Sci. 2014, 55, 36–45. [Google Scholar] [CrossRef]
- Li, X.; Thakkar, S.G.; Ruwona, T.B.; Williams, R.O., 3rd; Cui, Z. A method of lyophilizing vaccines containing aluminum salts into a dry powder without causing particle aggregation or decreasing the immunogenicity following reconstitution. J. Control. Release 2015, 204, 38–50. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Aldayel, A.M.; Cui, Z. Aluminum hydroxide nanoparticles show a stronger vaccine adjuvant activity than traditional aluminum hydroxide microparticles. J. Control. Release 2014, 173, 148–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hogenesch, H. Mechanism of immunopotentiation and safety of aluminum adjuvants. Front. Immunol. 2013, 3, 406. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Hum. Vaccines Immunother. 2015, 11, 477–488. [Google Scholar] [CrossRef]
- Quandt, D.; Rothe, K.; Baerwald, C.; Rossol, M. GPRC6A mediates Alum-induced Nlrp3 inflammasome activation but limits Th2 type antibody responses. Sci. Rep. 2015, 5, 16719. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, T.C.; Casella, C.R. No pain no gain? Adjuvant effects of alum and monophosphoryl lipid A in pertussis and HPV vac-cines. Curr. Opin. Immunol. 2017, 47, 17–25. [Google Scholar] [CrossRef]
- Baylor, N.W.; Egan, W.; Richman, P. Aluminum salts in vaccines--US perspective. Vaccine 2002, 20, S18–S23. [Google Scholar] [CrossRef] [PubMed]
- Nanishi, E.; Dowling, D.J.; Levy, O. Toward precision adjuvants: Optimizing science and safety. Curr. Opin. Pediatr. 2020, 32, 125–138. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hess, K.L.; Medintz, I.L.; Jewell, C.M. Designing inorganic nanomaterials for vaccines and immunotherapies. Nano Today 2019, 27, 73–98. [Google Scholar] [CrossRef]
- Li, X.; Wang, X.; Ito, A. Tailoring inorganic nanoadjuvants towards next-generation vaccines. Chem. Soc. Rev. 2018, 47, 4954–4980. [Google Scholar] [CrossRef]
- Sun, B.; Ji, Z.; Liao, Y.P.; Wang, M.; Wang, X.; Dong, J.; Chang, C.H.; Li, R.; Zhang, H.; Nel, A.E.; et al. Engineering an effective immune adjuvant by designed control of shape and crystallinity of aluminum oxyhydroxide nanoparticles. ACS Nano 2013, 7, 10834–10849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thakkar, S.G.; Xu, H.; Li, X.; Cui, Z. Uric acid and the vaccine adjuvant activity of aluminum (oxy)hydroxide nanoparticles. J. Drug Target. 2018, 26, 474–480. [Google Scholar] [CrossRef]
- Amini, Y.; Moradi, B.; Fasihi-Ramandi, M. Aluminum hydroxide nanoparticles show strong activity to stimulate Th-1 immune response against tuberculosis. Artif. Cells Nanomed. Biotechnol. 2017, 45, 1331–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, Z.; Wang, X.; Yu, G.; Li, M.; Shi, S.; Bao, H.; Chen, C.; Fu, D.; Ma, W.; Xue, C.; et al. Mechanistic understanding of the aspect ratio-dependent adjuvanticity of engineered aluminum oxyhydroxide nanorods in prophylactic vaccines. Nano Today 2022, 43, 101445. [Google Scholar] [CrossRef]
- Wang, T.; Zhen, Y.; Ma, X.; Wei, B.; Wang, N. Phospholipid bilayer-coated aluminum nanoparticles as an effective vaccine adjuvant-delivery system. ACS Appl. Mater. Interfaces 2015, 7, 6391–6396. [Google Scholar] [CrossRef]
- Liang, Z.; Yang, Y.; Yu, G.; Zhu, H.; Xia, X.; Chen, C.; Fu, D.; Li, M.; Cheng, G.; Xue, C.; et al. Engineering aluminum hydroxyphosphate nanoparticles with well-controlled surface property to enhance humoral immune responses as vaccine adjuvants. Biomaterials 2021, 275, 120960. [Google Scholar] [CrossRef]
- Sun, B.; Ji, Z.; Liao, Y.P.; Chang, C.H.; Wang, X.; Ku, J.; Xue, C.; Mirshafiee, V.; Xia, T. Enhanced Immune Adjuvant Activity of Aluminum Oxyhydroxide Nanorods through Cationic Surface Functionalization. ACS Appl. Mater. Interfaces 2017, 9, 21697–21705. [Google Scholar] [CrossRef] [PubMed]
- Ren, H.; Mou, Y.; Lin, L.; Wang, L.; Hu, H. Efficient antigen cross-presentation through coating conventional aluminum adjuvant particles with PEI. Am. J. Transl. Res. 2021, 13, 4092–4102. [Google Scholar] [PubMed]
- Ghasemi, M.; Bakhshi, B.; Khashei, R.; Soudi, S. Mesoporous silica nano-adjuvant triggers pro-inflammatory responses in Caco-2/peripheral blood mononuclear cell (PBMC) co-cultures. Nanobiomedicine 2022, 9, 18495435221088374. [Google Scholar] [CrossRef]
- Jafari, S.; Derakhshankhah, H.; Alaei, L.; Fattahi, A.; Varnamkhasti, B.S.; Saboury, A.A. Mesoporous silica nanoparticles for therapeutic/diagnostic applications. Biomed. Pharm. 2019, 109, 1100–1111. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, H.; Shi, J. In vivo bio-safety evaluations and diagnostic/therapeutic applications of chemically designed mesoporous silica nanoparticles. Adv. Mater. 2013, 25, 3144–3176. [Google Scholar] [CrossRef] [PubMed]
- Tang, F.; Li, L.; Chen, D. Mesoporous silica nanoparticles: Synthesis, biocompatibility and drug delivery. Adv. Mater. 2012, 24, 1504–1534. [Google Scholar] [CrossRef]
- Zhang, L.; Yang, W.; Hu, C.; Wang, Q.; Wu, Y. Properties and applications of nanoparticle/microparticle conveyors with adjuvant characteristics suitable for oral vaccination. Int. J. Nanomed. 2018, 13, 2973–2987. [Google Scholar] [CrossRef] [Green Version]
- Mahony, D.; Cavallaro, A.S.; Stahr, F.; Mahony, T.J.; Qiao, S.Z.; Mitter, N. Mesoporous silica nanoparticles act as a self-adjuvant for ovalbumin model antigen in mice. Small 2013, 9, 3138–3146. [Google Scholar] [CrossRef]
- Mody, K.T.; Popat, A.; Mahony, D.; Cavallaro, A.S.; Yu, C.; Mitter, N. Mesoporous silica nanoparticles as antigen carriers and adjuvants for vaccine delivery. Nanoscale 2013, 5, 5167–5179. [Google Scholar] [CrossRef]
- Carvalho, L.V.; Ruiz Rde, C.; Scaramuzzi, K.; Marengo, E.B.; Matos, J.R.; Tambourgi, D.V.; Fantini, M.C.; Sant’Anna, O.A. Immunological parameters related to the adjuvant effect of the ordered mesoporous silica SBA-15. Vaccine 2010, 28, 7829–7836. [Google Scholar] [CrossRef]
- Wang, T.; Jiang, H.; Zhao, Q.; Wang, S.; Zou, M.; Cheng, G. Enhanced mucosal and systemic immune responses obtained by porous silica nanoparticles used as an oral vaccine adjuvant: Effect of silica architecture on immunological properties. Int. J. Pharm. 2012, 436, 351–358. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Yoshino, S. Enhancement of ovalbumin-specific Th1, Th2, and Th17 immune responses by amorphous silica nanoparticles. Int. J. Immunopathol. Pharm. 2016, 29, 408–420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, M.K.; Kardjilov, N.; Oliveira, C.L.P.; Watts, B.; Villanova, J.; Botosso, V.F.; Sant’Anna, O.A.; Fantini, M.C.A.; Bordallo, H.N. 3D visualisation of hepatitis B vaccine in the oral delivery vehicle SBA-15. Sci. Rep. 2019, 9, 6106. [Google Scholar] [CrossRef] [Green Version]
- Rasmussen, M.K.; Bordallo, H.N.; Bordenalli, M.A.; Akamatsu, M.A.; Trezena, A.G.; Tino-De-Franco, M.; Sant’Anna, O.A.; da Silva Martins, T.; de Souza Lopes, H.; Fantini, M.C.; et al. Assessing the efficiency of SBA-15 as a nanocarrier for diphtheria anatoxin. Microporous Mesoporous Mater. 2021, 312, 110763. [Google Scholar] [CrossRef]
- Scaramuzzi, K.; Tanaka, G.D.; Neto, F.M.; Garcia, P.R.; Gabrili, J.J.; Oliveira, D.C.; Tambourgi, D.V.; Mussalem, J.S.; Paixao-Cavalcante, D.; D’Azeredo Orlando, M.T.; et al. Nanostructured SBA-15 silica: An effective protective vehicle to oral hepatitis B vaccine immunization. Nanomedicine 2016, 12, 2241–2250. [Google Scholar] [CrossRef]
- Xu, C.; Lei, C.; Yu, C. Mesoporous silica nanoparticles for protein protection and delivery. Front. Chem. 2019, 7, 290. [Google Scholar] [CrossRef] [Green Version]
- Zhou, S.; Zhong, Q.; Wang, Y.; Hua, P.; Zhong, W.; Huang, C.-B.; Yu, Z.-Q.; Ding, C.-D.; Liu, H.; Fu, J. Chemically engineered mesoporous silica nanoparticles-based intelligent delivery systems for theranostic applications in multiple cancerous/non-cancerous diseases. Coord. Chem. Rev. 2022, 452, 214309. [Google Scholar] [CrossRef]
- Chen, W.; Zuo, H.; Li, B.; Duan, C.; Rolfe, B.; Zhang, B.; Mahony, T.J.; Xu, Z.P. Clay nanoparticles elicit long-term immune responses by forming biodegradable depots for sustained antigen stimulation. Small 2018, 14, e1704465. [Google Scholar] [CrossRef]
- Govea-Alonso, D.O.; García-Soto, M.J.; Betancourt-Mendiola, L.; Padilla-Ortega, E.; Rosales-Mendoza, S.; González-Ortega, O. Nanoclays: Promising Materials for Vaccinology. Vaccines 2022, 10, 1549. [Google Scholar] [CrossRef]
- Pilar, A.; Darder, M.; Wicklein, B.; Rytwo, G.; Ruiz-Hitzky, E. Hybrid Organic–Inorganic Interfaces: Towards Advanced Functional Materials; Delville, M.H., Taubert, A., Eds.; Wiley-VCH Verlag GmbH & Co. KgaA: Weinheim, Germany, 2017; pp. 1–84. [Google Scholar]
- Wicklein, B.; Martín del Burgo, M.Á.; Yuste, M.; Darder, M.; Llavata, E.C.; Aranda, P.; Ortín, J.; del Real, G.; Ruiz-Hitzky, E. Lipid-based bio-nanohybrids for functional stabilisation of influenza vaccines. Eur. J. Inorg. Chem. 2012, 32, 5186–5191. [Google Scholar] [CrossRef]
- Chauhan, N.; Tiwari, S.; Iype, T.; Jain, U. An overview of adjuvants utilized in prophylactic vaccine formulation as immunomodulators. Expert. Rev. Vaccines 2017, 16, 491–502. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ren, H.; Zhang, Y. Metal-based nano-vaccines for cancer immunotherapy. Coord. Chem. Rev. 2022, 455, 214345. [Google Scholar] [CrossRef]
- Tadros, T.; Izquierdo, P.; Esquena, J.; Solans, C. Formation and stability of nano-emulsions. Adv. Colloid. Int. Sci. 2004, 108–109, 303–318. [Google Scholar] [CrossRef]
- Bonferoni, M.C.; Rossi, S.; Sandri, G.; Ferrari, F.; Gavini, E.; Rassu, G.; Giunchedi, P. Nanoemulsions for “Nose-to-Brain” Drug Delivery. Pharmaceutic 2019, 11, 84. [Google Scholar] [CrossRef] [Green Version]
- Đorđević, S.M.; Santrač, A.; Cekić, N.D.; Marković, B.D.; Divović, B.; Ilić, T.M.; Savić, M.; Savić, S.D. Parenteral nanoemulsions of risperidone for enhanced brain delivery in acute psychosis: Physicochemical and in vivo performances. Int. J. Pharm. 2017, 533, 421–430. [Google Scholar] [CrossRef]
- Nikolic, I.; Mitsou, E.; Pantelic, I.; Randjelovic, D.; Markovic, B.; Papadimitriou, V.; Xenakis, A.; Lunter, D.; Zugic, A.; Savic, S. Microstructure and biopharmaceutical performances of curcumin-loaded low-energy nanoemulsions containing eucalyptol and pinene: Terpenes’ role overcome penetration enhancement effect? Eur. J. Pharm. Sci. 2020, 142, 105135. [Google Scholar] [CrossRef]
- Tayeb, H.H.; Sainsbury, F. Nanoemulsions in drug delivery: Formulation to medical application. Nanomedicine 2018, 13, 2507–2525. [Google Scholar] [CrossRef]
- The European Medicines Agency, Committee for Medicinal Products for Human Use. Guideline on Adjuvants in Vaccines for Human Use. 2005. Available online: https://www.ema.europa.eu/en/adjuvants-vaccines-human-use (accessed on 11 March 2023).
- O’Hagan, D.T.; Ott, G.S.; Nest, G.V.; Rappuoli, R.; Giudice, G.D. The history of MF59® adjuvant: A phoenix that arose from the ashes. Expert. Rev. Vaccines 2013, 12, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Leroux-Roels, G. Unmet needs in modern vaccinology: Adjuvants to improve the immune response. Vaccine 2010, 28, C25–C36. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, G.; Rappuoli, R.; Didierlaurent, A.M. Correlates of adjuvanticity: A review on adjuvants in licensed vaccines. Semin. Immunol. 2018, 39, 14–21. [Google Scholar] [CrossRef]
- Oyewumi, M.O.; Kumar, A.; Cui, Z. Nano-microparticles as immune adjuvants: Correlating particle sizes and the resultant immune responses. Expert. Rev. Vaccines 2010, 9, 1095–1107. [Google Scholar] [CrossRef] [Green Version]
- Wallis, J.; Shenton, D.P.; Carlisle, R.C. Novel approaches for the design, delivery and administration of vaccine technologies. Clin. Exp. Immunol. 2019, 196, 189–204. [Google Scholar] [CrossRef] [Green Version]
- Aucouturier, J.; Dupuis, L.; Ganne, V. Adjuvants designed for veterinary and human vaccines. Vaccine 2001, 19, 2666–2672. [Google Scholar] [CrossRef]
- Klucker, M.F.; Dalençon, F.; Probeck, P.; Haensler, J. AF03, an alternative squalene emulsion-based vaccine adjuvant prepared by a phase inversion temperature method. J. Pharm. Sci. 2012, 101, 4490–4500. [Google Scholar] [CrossRef]
- Vogel, F.R.; Caillet, C.; Kusters, I.C.; Haensler, J. Emulsion-based adjuvants for influenza vaccines. Expert. Rev. Vaccine 2009, 8, 483–492. [Google Scholar] [CrossRef]
- Allison, A.C. Squalene and squalane emulsions as adjuvants. Methods 1999, 19, 87–93. [Google Scholar] [CrossRef]
- Fox, C. Squalene emulsions for parenteral vaccine and drug delivery. Molecules 2009, 14, 3286–3312. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, D.T.; Podda, A. Influenza Vaccines for the Future; Rappuoli, R., Del Giudice, G., Eds.; Birkhauser Basel: Siena, Italy, 2008; Chapter 10; pp. 221–244. [Google Scholar] [CrossRef]
- Seubert, A.; Monaci, E.; Pizza, M.; O’Hagan, D.T.; Wack, A. The adjuvants aluminum hydroxide and MF59 induce monocyte and granulocyte chemoattractants and enhance monocyte differentiation toward dendritic cells. J. Immunol. 2008, 180, 5402–5412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ozao-Choy, J.; Lee, D.J.; Faries, M.B. Melanoma vaccines: Mixed past, promising future. Surg. Clin. 2014, 94, 1017–1030. [Google Scholar] [CrossRef] [Green Version]
- Sosman, J.A.; Sondak, V.K. Melacine®: An allogeneic melanoma tumor cell lysate vaccine. Expert. Rev. Vaccines 2003, 2, 353–368. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Didierlaurent, A.; Bourguignon, P.; Delhaye, S.; Baras, B.; Jacob, V.; Planty, C.; Elouahabi, A.; Harvengt, P.; Carlsen, H.; et al. Adjuvant System AS03 containing α-tocopherol modulates innate immune response and leads to improved adaptive immunity. Vaccine 2011, 29, 2461–2473. [Google Scholar] [CrossRef] [PubMed]
- Dye, T.J.; Simakajornboon, N. Narcolepsy in Children: Sleep disorders in children, A rapidly evolving field seeking consensus. Pediatr. Pulmonol. 2022, 57, 1952–1962. [Google Scholar] [CrossRef] [PubMed]
- Aucouturier, J.; Dupuis, L.; Deville, S.; Ascarateil, S.; Ganne, V. Montanide ISA 720 and 51: A new generation of water in oil emulsions as adjuvants for human vaccines. Expert. Rev. Vaccines 2002, 1, 111–118. [Google Scholar] [CrossRef]
- Mirzaee, M.; Haghighat, S.; Golkaran, B.; Asgarhalvaei, F.; Mirzaee, R.; Taghizadeh, M.; Savoji, M.A.; Esfandiari, B.; Mahdavi, M. Montanide ISA-720 and Naloxone in HBsAg Vaccine Formulation: Cytokine Profiling and Monitoring of Long-Lasting Humoral Immune Responses. Biomed. Env. Sci. 2022, 35, 792–803. [Google Scholar] [CrossRef]
- Properties of MontanideTM Range of Ingredients. Available online: https://www.seppic.com/en/montanide_pharma (accessed on 15 March 2023).
- Bodewes, R.; Kreijtz, J.H.C.M.; Van Amerongen, G.; Geelhoed-Mieras, M.M.; Verburgh, R.J.; Heldens, J.G.; Bendwell, J.; Van Der Brand, J.M.; Kuiken, T.; Van Baalen, C.A.; et al. A single immunization with CoVaccine HT-adjuvanted H5N1 influenza virus vaccine induces protective cellular and humoral immune responses in ferrets. J. Virol. 2010, 84, 7943–7952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lai, C.Y.; To, A.; Ann, S.; Wong, T.; Lieberman, M.M.; Clements, D.E.; Senda, J.T.; Ball, A.H.; Pessaint, L.; Andersen, H.; et al. Recombinant protein subunit SARS-CoV-2 vaccines formulated with CoVaccine HT adjuvant induce broad, Th1 biased, humoral and cellular immune responses in mice [published online ahead of print, 2021 Nov 5]. Vaccine X 2021, 9, 100126. [Google Scholar] [CrossRef]
- Anamur, C.; Winter, G.; Engert, J. Stability of collapse lyophilized influenza vaccine formulations. Int. J. Pharm. 2015, 483, 131–141. [Google Scholar] [CrossRef]
- Do Vale Morais, A.R.; Do Nascimento Alencar, É.; Júnior, F.H.X.; De Oliveira, C.M.; Marcelino, H.R.; Barratt, G.; Fessi, H.; Do Egito, E.S.T.; Elaissari, A. Freeze-drying of emulsified systems: A review. Int. J. Pharm. 2016, 503, 102–114. [Google Scholar] [CrossRef]
- Yuan, L.; Gao, X.D.; Xia, Y. Optimising the oil phases of aluminium hydrogel-stabilised emulsions for stable, safe and efficient vaccine adjuvant. Front. Chem. Sci. Eng. 2022, 16, 973–984. [Google Scholar] [CrossRef]
- Schultze, V.D.; Agosto, V.; Wack, A.; Novicki, D.; Zorn, J.; Hennig, R. Safety of MF59 adjuvant. Vaccine 2008, 26, 3209–3222. [Google Scholar] [CrossRef]
- Fox, C.B.; Baldwin, S.L.; Duthie, M.S.; Reed, S.G.; Vedvick, T.S. Immunomodulatory and physical effects of oil composition in vaccine adjuvant emulsions. Vaccine 2011, 29, 9563–9572. [Google Scholar] [CrossRef] [Green Version]
- Assis, B.R.D.; da Silva, C.D.; Santiago, M.G.; Ferreira, L.A.M.; Goulart, G.A.C. Nanotechnology in adjuvants and vaccine development: What should we know? Nanomedicine 2021, 16, 2565–2568. [Google Scholar] [CrossRef]
- O’Hagan, D.T.; van der Most, R.; Lodaya, R.N.; Coccia, M.; Lofano, G. “World in motion”—Emulsion adjuvants rising to meet the pandemic challenges. NPJ Vaccines 2021, 6, 158. [Google Scholar] [CrossRef]
- Thanh Le, T.; Andreadakis, Z.; Kumar, A.; Gómez Román, R.; Tollefsen, S.; Saville, M.; Mayhew, S. The COVID-19 vaccine development landscape. Nat. Rev. Drug. Discov. 2020, 19, 305–306. [Google Scholar] [CrossRef] [PubMed]
- European Medicines Agency, VidPrevtyn Beta Product Information File. 2022. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/vidprevtyn-beta#product-information-section (accessed on 4 April 2023).
- Glaxo Smith Kline. Press. Release. 2022. Available online: https://www.gsk.com/en-gb/media/press-releases/sanofi-and-gsk-s-next-generation-covid-19-booster-vaccine-vidprevtyn-beta-approved-by-ec/ (accessed on 4 April 2023).
- Schoenmaker, L.; Witzigmann, D.; Kulkarni, J.A.; Verbeke, R.; Kersten, G.; Jiskoot, W.; Crommelin, D.J.A. mRNA-lipid nanoparticle COVID-19 vaccines: Structure and stability. Int. J. Pharm. 2021, 601, 120586. [Google Scholar] [CrossRef] [PubMed]
- Alameh, M.G.; Tombácz, I.; Bettini, E.; Lederer, K.; Sittplangkoon, C.; Wilmore, J.R.; Gaudette, B.T.; Soliman, O.Y.; Pine, M.; Hicks, P.; et al. Lipid nanoparticles enhance the efficacy of mRNA and protein subunit vaccines by inducing robust T follicular helper cell and humoral responses. Immunity 2021, 54, 2877–2892.e7. [Google Scholar] [CrossRef] [PubMed]
- Kobiyama, K.; Ishii, K.J. Making innate sense of mRNA vaccine adjuvanticity. Nat. Immunol. 2022, 23, 474–476. [Google Scholar] [CrossRef] [PubMed]
- Iyer, V.; Cayatte, C.; Guzman, B.; Schneider-Ohrum, K.; Matuszak, R.; Snell, A.; Rajani, G.M.; McCarthy, M.P.; Muralidhara, B. Impact of formulation and particle size on stability and immunogenicity of oil-in-water emulsion adjuvants. Hum. Vaccin. Immunother. 2015, 11, 1853–1864. [Google Scholar] [CrossRef] [Green Version]
- Niikura, K.; Matsunaga, T.; Suzuki, T.; Kobayashi, S.; Yamaguchi, H.; Orba, Y.; Kawaguchi, A.; Hasegawa, H.; Kajino, K.; Ninomiya, T.; et al. Gold nanoparticles as a vaccine platform: Influence of size and shape on immunological responses in vitro and in vivo. ACS Nano 2013, 7, 3926–3938. [Google Scholar] [CrossRef]
- Shah, R.R.; Taccone, M.; Monaci, E.; Brito, L.A.; Bonci, A.; O’Hagan, D.T.; Amiji, M.M.; Seubert, A. The droplet size of emulsion adjuvants has significant impact on their potency, due to differences in immune cell-recruitment and-activation. Sci. Rep. 2019, 9, 11520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shah, R.R.; O’Hagan, D.T.; Amiji, M.M.; Brito, L.A. The impact of size on particulate vaccine adjuvants. Nanomedicine 2014, 9, 2671–2681. [Google Scholar] [CrossRef]
- Somiya, M.; Kuroda, S. Development of a virus-mimicking nanocarrier for drug delivery systems: The bio-nanocapsule. Adv. Drug. Deliv. Rev. 2015, 95, 77–89. [Google Scholar] [CrossRef]
- Chung, Y.H.; Cai, H.; Steinmetz, N.F. Viral nanoparticles for drug delivery, imaging, immunotherapy, and theranostic applications. Adv. Drug. Deliv. Rev. 2020, 156, 214–235. [Google Scholar] [CrossRef]
- Fan, J.; Jin, S.; Gilmartin, L.; Toth, I.; Hussein, W.M.; Stephenson, R.J. Advances in Infectious Disease Vaccine Adjuvants. Vaccines 2022, 10, 1120. [Google Scholar] [CrossRef]
- Petkar, K.C.; Patil, S.M.; Chavhan, S.S.; Kaneko, K.; Sawant, K.K.; Kunda, N.K.; Saleem, I.Y. An Overview of Nanocarrier-Based Adjuvants for Vaccine Delivery. Pharmaceutics 2021, 27, 455. [Google Scholar] [CrossRef]
- Travieso, T.; Li, J.; Mahesh, S.; Mello, J.D.F.R.E.; Blasi, M. The use of viral vectors in vaccine development. NPJ Vaccines 2022, 7, 75. [Google Scholar] [CrossRef]
- Luiz, M.T.; Dutra, J.A.P.; Tofani, L.B.; de Araújo, J.T.C.; Di Filippo, L.D.; Marchetti, J.M.; Chorilli, M. Targeted Liposomes: A Nonviral Gene Delivery System for Cancer Therapy. Pharmaceutics 2022, 14, 821. [Google Scholar] [CrossRef] [PubMed]
- Plummer, E.M.; Manchester, M. Viral nanoparticles and virus-like particles: Platforms for contemporary vaccine design. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol 2011, 3, 174–196. [Google Scholar] [CrossRef]
- Suleman, M.; Galea, S.; Gavard, F.; Merillon, N.; Klonjkowski, B.; Tartour, E.; Richardson, J. Antigen encoded by vaccine vectors derived from human adenovirus serotype 5 is preferentially presented to CD8+ T lymphocytes by the CD8α+ dendritic cell subset. Vaccine 2011, 29, 5892–5903. [Google Scholar] [CrossRef] [PubMed]
- Buchbinder, S.P.; Mehrotra, D.V.; Duerr, A.; Fitzgerald, D.W.; Mogg, R.; Li, D.; Gilbert, P.B.; Lama, J.R.; Marmor, M.; Del Rio, C.; et al. Step Study Protocol Team. Efficacy assessment of a cell-mediated immunity HIV-1 vaccine (the Step Study): A double-blind, randomised, placebo-controlled, test-of-concept trial. Lancet 2008, 372, 1881–1893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Zhong, G.; Zhang, J.; Shuai, L.; Zhang, Z.; Wen, Z.; Wang, B.; Zhao, Z.; Song, X.; Chen, Y.; et al. A single dose of an adenovirus-vectored vaccine provides protection against SARS-CoV-2 challenge. Nat. Commun. 2020, 11, 4081. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.C.; Guan, X.H.; Li, Y.H.; Huang, J.Y.; Jiang, T.; Hou, L.H.; Li, J.X.; Yang, B.F.; Wang, L.; Wang, W.J.; et al. Immunogenicity and safety of a recombinant adenovirus type-5-vectored COVID-19 vaccine in healthy adults aged 18 years or older: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Zhu, F.-C.; Li, Y.-H.; Guan, X.-H.; Hou, L.-H.; Wang, W.-J.; Li, J.-X.; Wu, S.-P.; Wang, B.-S.; Wang, Z.; Wang, L.; et al. Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: A dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 2020, 395, 1845–1854. [Google Scholar] [CrossRef] [PubMed]
- Milligan, I.D.; Gibani, M.M.; Sewell, R.; Clutterbuck, E.A.; Campbell, D.; Plested, E.; Nuthall, E.; Voysey, M.; Silva-Reyes, L.; McElrath, M.J.; et al. Safety and Immunogenicity of Novel Adenovirus Type 26- and Modified Vaccinia Ankara-Vectored Ebola Vaccines: A Randomized Clinical Trial. JAMA 2016, 315, 1610–1623. [Google Scholar] [CrossRef] [PubMed]
- Sadoff, J.; Le Gars, M.; Shukarev, G.; Heerwegh, D.; Truyers, C.; de Groot, A.M.; Stoop, J.; Tete, S.; Van Damme, W.; Leroux-Roels, I.; et al. Interim Results of a Phase 1-2a Trial of Ad26.COV2.S Covid-19 Vaccine. N. Engl. J. Med. 2021, 384, 1824–1835. [Google Scholar] [CrossRef] [PubMed]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Oxford COVID Vaccine Trial Group. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Baker, A.T.; Boyd, R.J.; Sarkar, D.; Teijeira-Crespo, A.; Chan, C.K.; Bates, E.; Waraich, K.; Vant, J.; Wilson, E.; Truong, C.D. ChAdOx1 interacts with CAR and PF4 with implications for thrombosis with thrombocytopenia syndrome. Sci. Adv. 2021, 7, eabl8213. [Google Scholar] [CrossRef]
- Manning, W.C.; Paliard, X.; Zhou, S.; Pat Bland, M.; Lee, A.Y.; Hong, K.; Walker, C.M.; Escobedo, J.A.; Dwarki, V. Genetic immunization with adeno-associated virus vectors expressing herpes simplex virus type 2 glycoproteins B and D. J. Virol. 1997, 71, 7960–7962. [Google Scholar] [CrossRef] [Green Version]
- Kuck, D.; Lau, T.; Leuchs, B.; Kern, A.; Müller, M.; Gissmann, L.; Kleinschmidt, J.A. Intranasal vaccination with recombinant adeno-associated virus type 5 against human papillomavirus type 16 L1. J. Virol. 2006, 80, 2621–2630. [Google Scholar] [CrossRef] [Green Version]
- Ploquin, A.; Szécsi, J.; Mathieu, C.; Guillaume, V.; Barateau, V.; Ong, K.C.; Wong, K.T.; Cosset, F.L.; Horvat, B.; Salvetti, A. Protection against henipavirus infection by use of recombinant adeno-associated virus-vector vaccines. J. Infect. Dis. 2013, 207, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Zhan, W.; Muhuri, M.; Tai, P.W.L.; Gao, G. Vectored Immunotherapeutics for Infectious Diseases: Can rAAVs Be The Game Changers for Fighting Transmissible Pathogens? Front. Immunol. 2021, 2021, 673699. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Navio, J.M.; Fuchs, S.P.; Pantry, S.N.; Lauer, W.A.; Duggan, N.N.; Keele, B.F.; Rakasz, E.G.; Gao, G.; Lifson, J.D.; Desrosiers, R.C. Adeno-Associated Virus Delivery of Anti-HIV Monoclonal Antibodies Can Drive Long-Term Virologic Suppression. Immunity 2019, 50, 567–575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fathi, A.; Dahlke, C.; Addo, M.M. Recombinant vesicular stomatitis virus vector vaccines for WHO blueprint priority pathogens. Hum. Vaccines Immunother. 2019, 15, 2269–2285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurana, S.; Fuentes, S.; Coyle, E.M.; Ravichandran, S.; Davey, R.T., Jr.; Beigel, J.H. Human antibody repertoire after VSV-Ebola vaccination identifies novel targets and virus-neutralizing IgM antibodies. Nat. Med. 2016, 22, 1439–1447. [Google Scholar] [CrossRef]
- Case, J.B.; Rothlauf, P.W.; Chen, R.E.; Kafai, N.M.; Fox, J.M.; Smith, B.K.; Shrihari, S.; McCune, B.T.; Harvey, I.B.; Keeler, S.P.; et al. Replication-Competent Vesicular Stomatitis Virus Vaccine Vector Protects against SARS-CoV-2-Mediated Pathogenesis in Mice. Cell Host Microbe 2020, 28, 465–474. [Google Scholar] [CrossRef]
- Ku, M.W.; Charneau, P.; Majlessi, L. Use of lentiviral vectors in vaccination. Expert. Rev. Vaccines 2021, 20, 1571–1586. [Google Scholar] [CrossRef]
- Ku, M.W.; Bourgine, M.; Authié, P.; Lopez, J.; Nemirov, K.; Moncoq, F.; Noirat, A.; Vesin, B.; Nevo, F.; Blanc, C.; et al. Intranasal vaccination with a lentiviral vector protects against SARS-CoV-2 in preclinical animal models. Cell Host Microbe 2021, 29, 236–249. [Google Scholar] [CrossRef]
- Blasi, M.; Wescott, E.C.; Baker, E.J.; Mildenberg, B.; LaBranche, C.; Rountree, W.; Haynes, B.F.; Saunders, K.O.; Moody, M.A.; Negri, D.; et al. Therapeutic vaccination with IDLV-SIV-Gag results in durable viremia control in chronically SHIV-infected macaques. NPJ Vaccines 2020, 5, 36. [Google Scholar] [CrossRef]
- Coutant, F.; Sanchez David, R.Y.; Félix, T.; Boulay, A.; Caleechurn, L.; Souque, P.; Thouvenot, C.; Bourgouin, C.; Beignon, A.S.; Charneau, P. A nonintegrative lentiviral vector-based vaccine provides long-term sterile protection against malaria. PLoS ONE 2012, 7, e48644. [Google Scholar] [CrossRef] [Green Version]
- Ku, M.W.; Anna, F.; Souque, P.; Petres, S.; Prot, M.; Simon-Loriere, E.; Charneau, P.; Bourgine, M.A. Single Dose of NILV-Based Vaccine Provides Rapid and Durable Protection against Zika Virus. Molecules 2020, 28, 1772–1782. [Google Scholar] [CrossRef]
- Yang, C.; Huang, K. Clinical applications of virus-like particles: Opportunities and challenges. Curr. Protein Pept. Sci. 2019, 20, 488–489. [Google Scholar] [CrossRef] [PubMed]
- Nooraei, S.; Bahrulolum, H.; Hoseini, Z.S.; Katalani, C.; Hajizade, A.; Easton, A.J.; Ahmadian, G. Virus-like particles: Preparation, immunogenicity and their roles as nanovaccines and drug nanocarriers. J. Nanobiotechnology 2021, 19, 59. [Google Scholar] [CrossRef] [PubMed]
- Mejía-Méndez, J.L.; Vazquez-Duhalt, R.; Hernández, L.R.; Sánchez-Arreola, E.; Bach, H. Virus-like Particles: Fundamentals and Biomedical Applications. Int. J. Mol. Sci. 2022, 23, 8579. [Google Scholar] [CrossRef] [PubMed]
- Qian, C.; Liu, X.; Xu, Q.; Wang, Z.; Chen, J.; Li, T.; Zheng, Q.; Yu, H.; Gu, Y.; Li, S.; et al. Recent Progress on the Versatility of Virus-Like Particles. Vaccines 2020, 8, 139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bayer, M.E.; Blumberg, B.S.; Werner, B. Particles associated with Australia antigen in the sera of patients with leukaemia, Down’s Syndrome and hepatitis. Nature 1968, 218, 1057–1059. [Google Scholar] [CrossRef]
- Thomas, D.L.; Astemborski, J.; Rai, R.M.; Anania, F.A.; Schaeffer, M.; Galai, N.; Nolt, K.; Nelson, K.E.; Strathdee, S.A.; Johnson, L. The natural history of hepatitis C virus infection: Host, viral, and environmental factors. JAMA 2000, 284, 450–456. [Google Scholar] [CrossRef] [Green Version]
- Metz, S.W.; Thomas, A.; White, L. Dengue virus-like particles mimic the antigenic properties of the infectious dengue virus envelope. Virol. J. 2018, 15, 60. [Google Scholar] [CrossRef]
- Perotti, M.; Perez, L. Virus-Like Particles and Nanoparticles for Vaccine Development against HCMV. Viruses 2019, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Mohsen, M.O.; Gomes, A.C.; Vogel, M. Interaction of Viral Capsid-Derived Virus-Like Particles (VLPs) with the Innate Immune System. Vaccines 2018, 6, 37. [Google Scholar] [CrossRef] [Green Version]
- Anzaghe, M.; Schülke, S.; Scheurer, S. Virus-like particles as carrier systems to enhance immunomodulation in allergen immunotherapy. Curr. Allergy Asthma Rep. 2018, 18, 71. [Google Scholar] [CrossRef]
- Zepeda-Cervantes, J.; Ramírez-Jarquín, J.O.; Vaca, L. Interaction Between Virus-Like Particles (VLPs) and Pattern Recognition Receptors (PRRs) From Dendritic Cells (DCs): Toward Better Engineering of VLPs. Front. Immunol. 2020, 11, 1100. [Google Scholar] [CrossRef] [PubMed]
- Tariq, H.; Batool, S.; Asif, S.; Ali, M.; Abbasi, B.H. Virus-Like Particles: Revolutionary Platforms for Developing Vaccines Against Emerging Infectious Diseases. Front. Microbiol. 2022, 12, 790121. [Google Scholar] [CrossRef]
- Dai, S.; Wang, H.; Deng, F. Advances and challenges in enveloped virus-like particle (VLP)-based vaccines. J. Immunol. Sci. 2018, 2, 36–41. [Google Scholar]
- Mohsen, M.O.; Zha, L.; Cabral-Miranda, G.; Bachmann, M.F. Major findings and recent advances in virus–like particle (VLP)-based vaccines. Semin. Immunol. 2017, 34, 123–132. [Google Scholar] [CrossRef] [PubMed]
- Peabody, D.S.; Peabody, J.; Bradfute, S.B.; Chackerian, B. RNA Phage VLP-Based Vaccine Platforms. Pharmaceuticals 2021, 4, 764. [Google Scholar] [CrossRef]
- Tars, K. ssRNA Phages: Life Cycle, Structure and Applications. Biocommunication Phages 2020, 30, 261–292. [Google Scholar] [CrossRef]
- Liekniņa, I.; Černova, D.; Rūmnieks, J.; Tārs, K. Novel ssRNA phage VLP platform for displaying foreign epitopes by genetic fusion. Vaccine 2020, 27, 6019–6026. [Google Scholar] [CrossRef]
- Palma, M. Aspects of Phage-Based Vaccines for Protein and Epitope Immunization. Vaccines 2023, 11, 436. [Google Scholar] [CrossRef]
- Tumban, E.; Peabody, J.; Peabody, D.S.; Chackerian, B. A pan-HPV vaccine based on bacteriophage PP7 VLPs displaying broadly cross-neutralizing epitopes from the HPV minor capsid protein, L2. PLoS ONE 2011, 6, e23310. [Google Scholar] [CrossRef]
- Guo, Y.; Guo, R.; Ma, Y.; Chang, W.; Ming, S.; Yang, G.; Guo, Y. Chimeric Virus-like Particles of Universal Antigen Epitopes of Coronavirus and Phage Qβ Coat Protein Trigger the Production of Neutralizing Antibodies. Curr. Top. Med. Chem. 2021, 5, 1235–1250. [Google Scholar] [CrossRef]
- Kamel, M.; El-Sayed, A.; Castañeda Vazquez, H. Foot-and-mouth disease vaccines: Recent updates and future perspectives. Arch. Virol. 2019, 164, 1501–1513. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, N.; Beerli, R.R.; Bauer, M.; Jegerlehner, A.; Dietmeier, K.; Maudrich, M.; Pumpens, P.; Saudan, P.; Bachmann, M.F. Universal vaccine against influenza virus: Linking TLR signaling to anti-viral protection. Eur. J. Immunol. 2012, 42, 863–869. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Chang, X.; Rothen, D.; Derveni, M.; Krenger, P.; Roongta, S.; Wright, E.; Vogel, M.; Tars, K.; Mohsen, M.O.; et al. AP205 VLPs Based on Dimerized Capsid Proteins Accommodate RBM Domain of SARS-CoV-2 and Serve as an Attractive Vaccine Candidate. Vaccines 2021, 19, 403. [Google Scholar] [CrossRef] [PubMed]
- Klimek, L.; Kündig, T.; Kramer, M.F.; Guethoff, S.; Jensen-Jarolim, E.; Schmidt-Weber, C.B.; Palomares, O.; Mohsen, M.O.; Jakob, T.; Bachmann, M. Virus-like particles (VLP) in prophylaxis and immunotherapy of allergic diseases. Allergo J. Int. 2018, 27, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asadi, K.; Gholami, A. Virosome-based nanovaccines; a promising bioinspiration and biomimetic approach for preventing viral diseases: A review. Int. J. Biol. Macromol. 2021, 182, 648–658. [Google Scholar] [CrossRef] [PubMed]
- Huckriede, A.; Bungener, L.; Stegmann, T.; Daemen, T.; Medema, J.; Palache, A.M.; Wilschut, J. The virosome concept for influenza vaccines. Vaccine 2005, 23, S26–S38. [Google Scholar] [CrossRef]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef]
- Nidetz, N.F.; McGee, M.C.; Tse, L.V.; Li, C.; Cong, L.; Li, Y.; Huang, W. Adeno-associated viral vector-mediated immune responses: Understanding barriers to gene delivery. Pharm. 2020, 207, 107453. [Google Scholar] [CrossRef]
- Pinski, A.N.; Messaoudi, I. To B or Not to B: Mechanisms of Protection Conferred by rVSV-EBOV-GP and the Roles of Innate and Adaptive Immunity. Microorganisms 2020, 25, 1473. [Google Scholar] [CrossRef]
- Nemirov, K.; Bourgine, M.; Anna, F.; Wei, Y.; Charneau, P.; Majlessi, L. Lentiviral Vectors as a Vaccine Platform against Infectious Diseases. Pharmaceutics 2023, 15, 846. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filipić, B.; Pantelić, I.; Nikolić, I.; Majhen, D.; Stojić-Vukanić, Z.; Savić, S.; Krajišnik, D. Nanoparticle-Based Adjuvants and Delivery Systems for Modern Vaccines. Vaccines 2023, 11, 1172. https://doi.org/10.3390/vaccines11071172
Filipić B, Pantelić I, Nikolić I, Majhen D, Stojić-Vukanić Z, Savić S, Krajišnik D. Nanoparticle-Based Adjuvants and Delivery Systems for Modern Vaccines. Vaccines. 2023; 11(7):1172. https://doi.org/10.3390/vaccines11071172
Chicago/Turabian StyleFilipić, Brankica, Ivana Pantelić, Ines Nikolić, Dragomira Majhen, Zorica Stojić-Vukanić, Snežana Savić, and Danina Krajišnik. 2023. "Nanoparticle-Based Adjuvants and Delivery Systems for Modern Vaccines" Vaccines 11, no. 7: 1172. https://doi.org/10.3390/vaccines11071172