
Mycobacterium avium subsp. paratuberculosis Candidate Vaccine Strains Are Pro-apoptotic in RAW 264.7 Murine Macrophages
 , , and
, , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
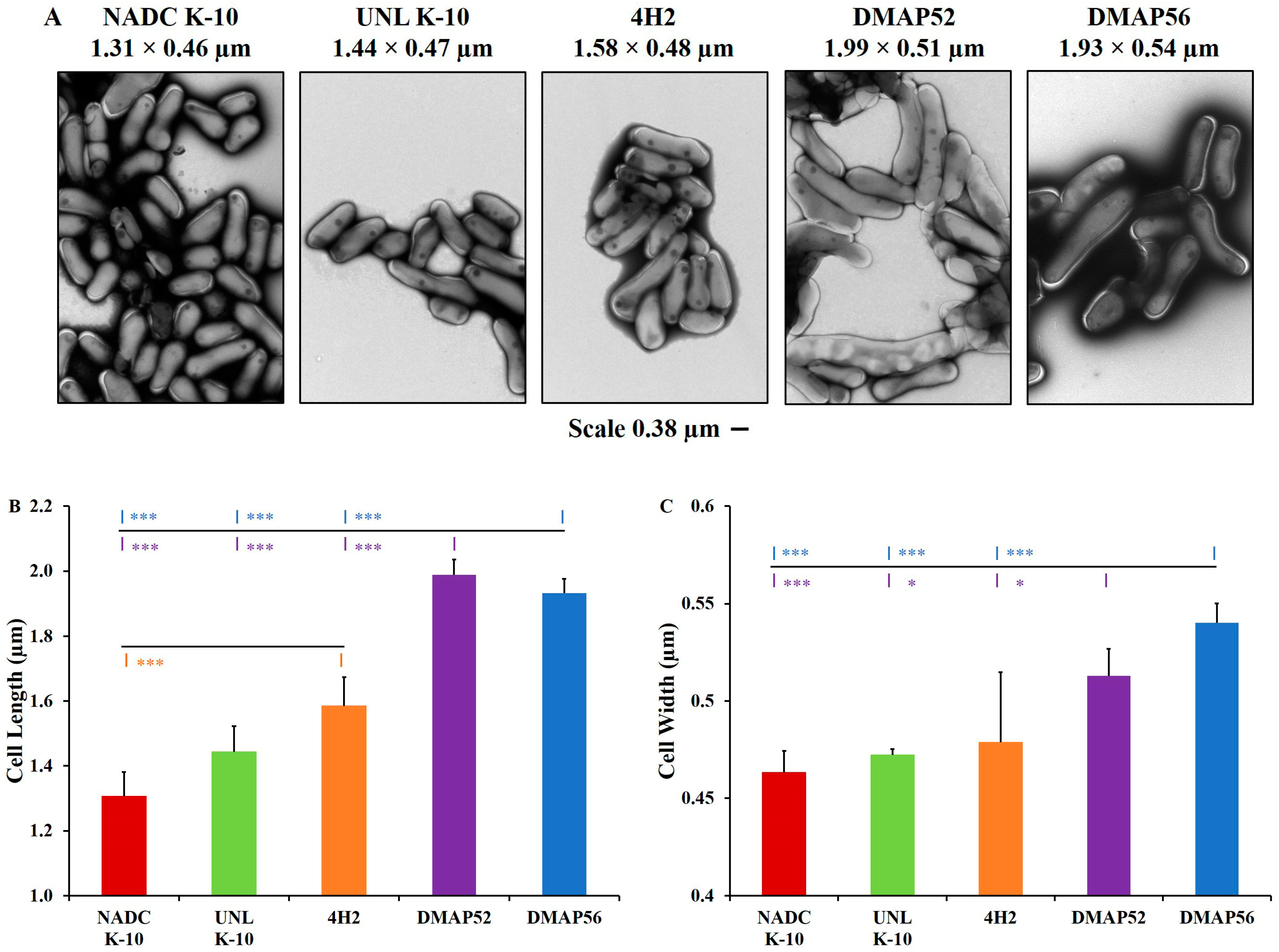
2.1. Bacterial Strains, Growth Curves, and Cell Morphology
2.2. RAW 264.7 Macrophage Cells and MAP Infection Protocol
2.3. Real-Time Apoptosis and Necrosis Assays
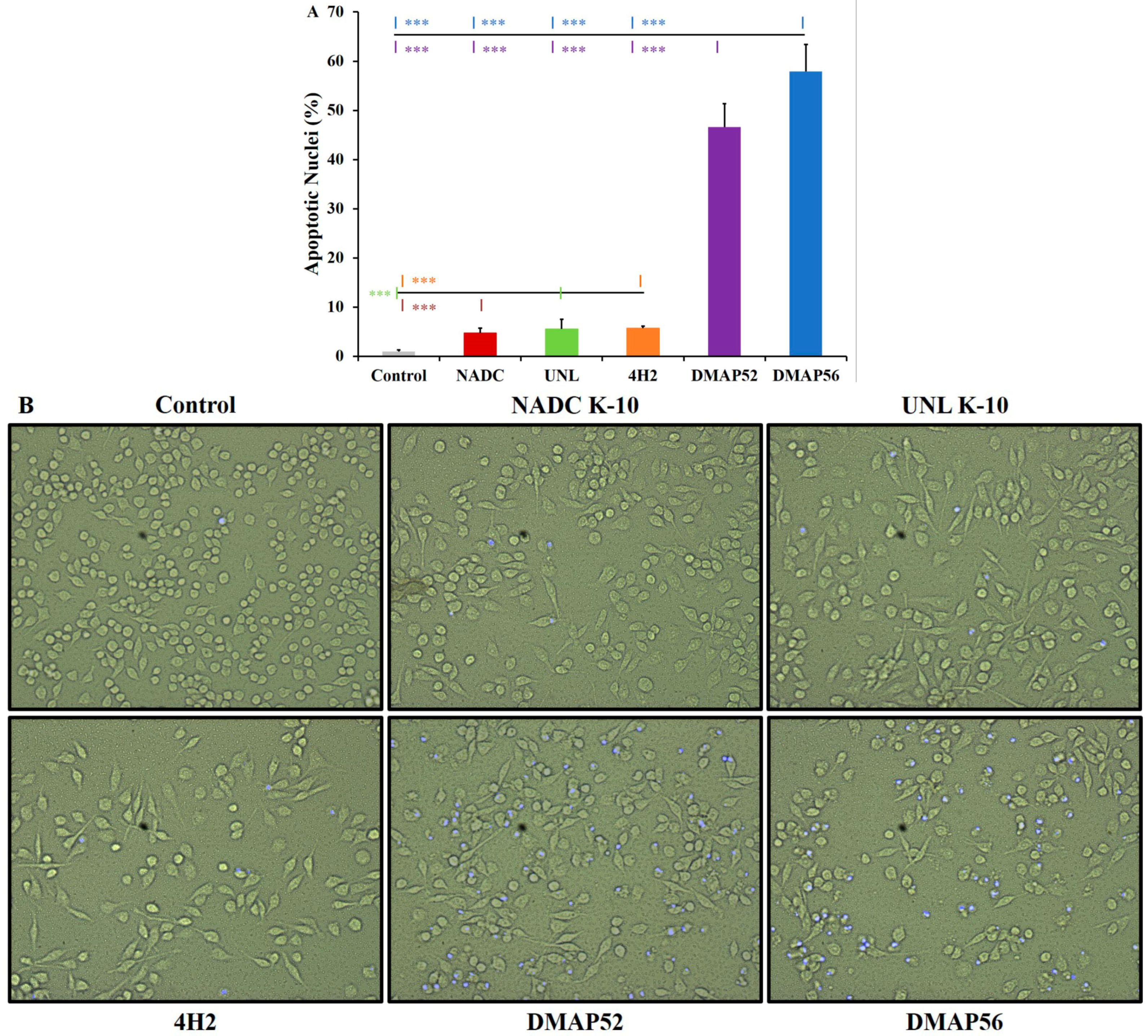
2.4. Apoptosis Assay via DAPI Staining
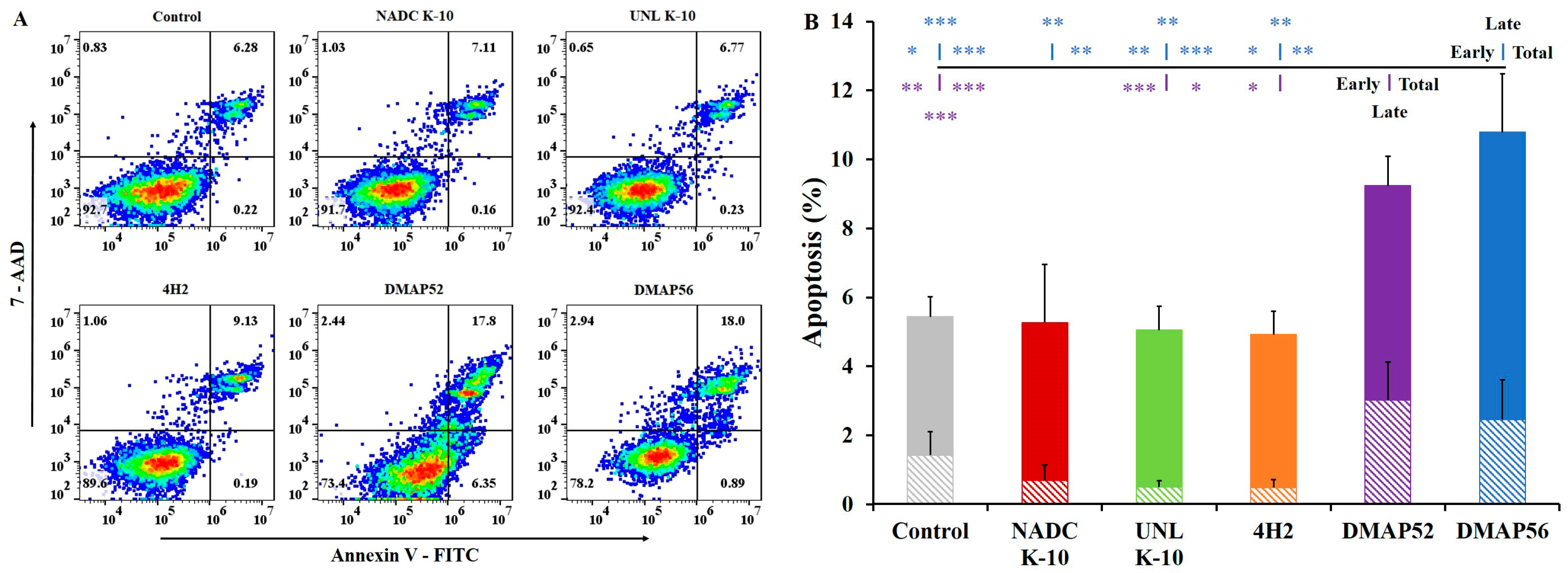
2.5. Apoptosis Assay via Flow Cytometry
2.6. Statistics
3. Results
3.1. Growth Curves and Cell Morphology
3.2. Real-Time Apoptosis and Necrosis Assays
3.3. Apoptosis Assay via DAPI Staining
3.4. Apoptosis Assay via Flow Cytometry
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rathnaiah, G.; Zinniel, D.K.; Bannantine, J.P.; Stabel, J.R.; Grohn, Y.T.; Collins, M.T.; Barletta, R.G. Pathogenesis, molecular genetics, and genomics of Mycobacterium avium subsp. paratuberculosis, the etiologic agent of Johne’s disease. Front. Vet. Sci. 2017, 4, 187. [Google Scholar] [CrossRef] [Green Version]
- Harris, N.B.; Barletta, R.G. Mycobacterium avium subsp. paratuberculosis in veterinary medicine. Clin. Microbiol. Rev. 2001, 14, 489–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bastida, F.; Juste, R.A. Paratuberculosis control: A review with a focus on vaccination. J. Immune Based Ther. Vaccines 2011, 9, 8. [Google Scholar] [CrossRef] [Green Version]
- Juste, R.A.; Alonso-Hearn, M.; Molina, E.; Geijo, M.; Vazquez, P.; Sevilla, I.A.; Garrido, J.M. Significant reduction in bacterial shedding and improvement in milk production in dairy farms after the use of a new inactivated paratuberculosis vaccine in a field trial. BMC Res. Notes 2009, 2. [Google Scholar] [CrossRef] [Green Version]
- Tewari, D.; Hovingh, E.; Linscott, R.; Martel, E.; Lawrence, J.; Wolfgang, D.; Griswold, D. Mycobacterium avium subsp. paratuberculosis antibody response, fecal shedding, and antibody cross-reactivity to Mycobacterium bovis in M. avium subsp. paratuberculosis-infected cattle herds vaccinated against Johne’s disease. Clin. Vaccine Immunol. 2014, 21, 698–703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sechi, L.A.; Dow, C.T. Mycobacterium avium ss. paratuberculosis zoonosis–the hundred year war—Beyond Crohn’s disease. Front. Immunol. 2015, 6, 96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Galluzzi, L.; Maiuri, M.C.; Vitale, I.; Zischka, H.; Castedo, M.; Zitvogel, L.; Kroemer, G. Cell death modalities: Classification and pathophysiological implications. Cell Death Differ. 2007, 14, 1237–1243. [Google Scholar] [CrossRef]
- Finlay, B.B.; McFadden, G. Anti-immunology: Evasion of the host immune system by bacterial and viral pathogens. Cell 2006, 124, 767–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostetter, J.; Steadham, E.; Haynes, J.; Bailey, T.; Cheville, N. Phagosomal maturation and intracellular survival of Mycobacterium avium subspecies paratuberculosis in J774 cells. Comp. Immunol. Microbiol. Infect. Dis. 2003, 26, 269–283. [Google Scholar] [CrossRef]
- Tanaka, S.; Sato, M.; Onitsuka, T.; Kamata, H.; Yokomizo, Y. Inflammatory cytokine gene expression in different types of granulomatous lesions during asymptomatic stages of bovine paratuberculosis. Vet. Pathol. 2005, 42, 579–588. [Google Scholar] [CrossRef]
- Faherty, C.S.; Maurelli, A.T. Staying alive: Bacterial inhibition of apoptosis during infection. Trends Microbiol. 2008, 16, 173–180. [Google Scholar] [CrossRef] [Green Version]
- Lam, A.; Prabhu, R.; Gross, C.M.; Riesenberg, L.A.; Singh, V.; Aggarwal, S. Role of apoptosis and autophagy in tuberculosis. Am. J. Physiol.-Lung Cell Mol. Physiol. 2017, 313, L218–L229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kabara, E.; Coussens, P.M. Infection of primary bovine macrophages with Mycobacterium avium subspecies paratuberculosis suppresses host cell apoptosis. Front. Microbiol. 2012, 3, 215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucena, A.N.; Garza-Cuartero, L.; McAloon, C.; Mulcahy, G.; Zintl, A.; Perez, J.; Wolfe, A. Apoptosis levels in bovine Johne’s disease ileal lesions and association with bacterial numbers. Vet. Pathol. 2021, 58, 1086–1090. [Google Scholar] [CrossRef] [PubMed]
- Aporta, A.; Arbues, A.; Aguilo, J.I.; Monzon, M.; Badiola, J.J.; de Martino, A.; Ferrer, N.; Marinova, D.; Anel, A.; Martin, C.; et al. Attenuated Mycobacterium tuberculosis SO2 vaccine candidate is unable to induce cell death. PLoS ONE 2012, 7, e45213. [Google Scholar] [CrossRef]
- Early, J.; Fischer, K.; Bermudez, L.E. Mycobacterium avium uses apoptotic macrophages as tools for spreading. Microb. Pathog. 2011, 50, 132–139. [Google Scholar] [CrossRef] [Green Version]
- Woo, S.R.; Sotos, J.; Hart, A.P.; Barletta, R.G.; Czuprynski, C.J. Bovine monocytes and a macrophage cell line differ in their ability to phagocytose and support the intracellular survival of Mycobacterium avium subsp. paratuberculosis. Vet. Immunol. Immunopathol. 2006, 110, 109–120. [Google Scholar] [CrossRef] [PubMed]
- Adams, J.L.; Czuprynski, C.J. Mycobacterial cell wall components induce the production of TNF-alpha, IL-1, and IL-6 by bovine monocytes and the murine macrophage cell line RAW 264.7. Microb. Pathog. 1994, 16, 401–411. [Google Scholar] [CrossRef]
- Wang, Z.; Kong, L.C.; Jia, B.Y.; Chen, J.R.; Dong, Y.; Jiang, X.Y.; Ma, H.X. Analysis of the microRNA expression profile of bovine monocyte-derived macrophages infected with Mycobacterium avium subsp. paratuberculosis reveals that miR-150 suppresses cell apoptosis by targeting PDCD4. Int. J. Mol. Sci. 2019, 20, 2708. [Google Scholar] [CrossRef] [Green Version]
- Roupie, V.; Rosseels, V.; Piersoel, V.; Zinniel, D.K.; Barletta, R.G.; Huygen, K. Genetic resistance of mice to Mycobacterium paratuberculosis is influenced by Slc11a1 at the early but not at the late stage of infection. Infect. Immun. 2008, 76, 2099–2105. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.J.; Wu, C.W.; Steinberg, H.; Talaat, A.M. Identification of novel virulence determinants in Mycobacterium paratuberculosis by screening a library of insertional mutants. Infect. Immun. 2006, 74, 3825–3833. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.W.; Schmoller, S.K.; Shin, S.J.; Talaat, A.M. Defining the stressome of Mycobacterium avium subsp. paratuberculosis in vitro and in naturally infected cows. J. Bacteriol. 2007, 189, 7877–7886. [Google Scholar] [CrossRef] [Green Version]
- Hanafy, M.; Hansen, C.; Phanse, Y.; Wu, C.W.; Nelson, K.; Aschenbroich, S.A.; Talaat, A.M. Characterization of early immune responses elicited by live and inactivated vaccines against Johne’s disease in goats. Front. Vet. Sci. 2022, 9. [Google Scholar] [CrossRef]
- Walker, K.B.; Brennan, M.J.; Ho, M.M.; Eskola, J.; Thiry, G.; Sadoff, J.; Dobbelaer, R.; Grode, L.; Liu, M.A.; Fruth, U.; et al. The second Geneva consensus: Recommendations for novel live TB vaccines. Vaccine 2010, 28, 2259–2270. [Google Scholar] [CrossRef]
- Rathnaiah, G.; Lamont, E.A.; Harris, N.B.; Fenton, R.J.; Zinniel, D.K.; Liu, X.; Sotos, J.; Feng, Z.; Livneh-Kol, A.; Shpigel, N.Y.; et al. Generation and screening of a comprehensive Mycobacterium avium subsp. paratuberculosis transposon mutant bank. Front. Cell Infect. Microbiol. 2014, 4, 144. [Google Scholar] [CrossRef]
- Rathnaiah, G.; Bannantine, J.P.; Bayles, D.O.; Zinniel, D.K.; Stabel, J.R.; Grohn, Y.T.; Barletta, R.G. Analysis of Mycobacterium avium subsp. paratuberculosis mutant libraries reveals loci-dependent transposition biases and strategies for novel mutant discovery. Microbiology 2016, 162, 633–641. [Google Scholar] [CrossRef]
- Bannantine, J.P.; Zinniel, D.K.; Barletta, R.G. Transposon mutagenesis in Mycobacterium avium subspecies paratuberculosis. Methods Mol. Biol. 2019, 2016, 117–125. [Google Scholar] [CrossRef] [Green Version]
- Bannantine, J.P.; Paulson, A.L.; Chacon, O.; Fenton, R.J.; Zinniel, D.K.; McVey, D.S.; Smith, D.R.; Czuprynski, C.J.; Barletta, R.G. Immunogenicity and reactivity of novel Mycobacterium avium subsp. paratuberculosis PPE MAP1152 and conserved MAP1156 proteins with sera from experimentally and naturally infected animals. Clin. Vaccine Immunol. 2011, 18, 105–112. [Google Scholar] [CrossRef] [Green Version]
- Barletta, R.G.; Lamont, E.A.; Stabel, J.R.; Fenton, R.J.; Rathnaiah, G.; Zinniel, D.K.; Posey, J.E.; McGarvey, J.A.; Bannantine, J.P.; Sreevatsan, S. Mycobacterium avium subsp. paratuberculosis mutants in ORFs MAP_1152 and MAP_1156 are attenuated and immunogenic in primary bovine macrophages. In Proceedings of the American Society of Microbiology Microbe, San Francisco, CA, USA, 23 June 2019. [Google Scholar]
- Foley-Thomas, E.M.; Whipple, D.L.; Bermudez, L.E.; Barletta, R.G. Phage infection, transfection and transformation of Mycobacterium avium complex and Mycobacterium paratuberculosis. Microbiology 1995, 141, 1173–1181. [Google Scholar] [CrossRef] [Green Version]
- Stabel, J.R.; Palmer, M.V.; Harris, B.; Plattner, B.; Hostetter, J.; Robbe-Austerman, S. Pathogenesis of Mycobacterium avium subsp. paratuberculosis in neonatal calves after oral or intraperitoneal experimental infection. Vet. Microbiol. 2009, 136, 306–313. [Google Scholar] [CrossRef]
- Bardarov, S.; Bardarov, S.; Pavelka, M.S.; Sambandamurthy, V.; Larsen, M.; Tufariello, J.; Chan, J.; Hatfull, G.; Jacobs, W.R. Specialized transduction: An efficient method for generating marked and unmarked targeted gene disruptions in Mycobacterium tuberculosis, M. bovis BCG and M. smegmatis. Microbiology 2002, 148, 3007–3017. [Google Scholar] [CrossRef] [Green Version]
- Zanetti, S.; Molicotti, P.; Cannas, S.; Ortu, S.; Ahmed, N.; Sechi, L.A. “In vitro” activities of antimycobacterial agents against Mycobacterium avium subsp. paratuberculosis linked to Crohn’s disease and paratuberculosis. Ann. Clin. Microbiol. Antimicrob. 2006, 5. [Google Scholar] [CrossRef] [Green Version]
- Nazarova, E.V.; Russell, D.G. Growing and handling of Mycobacterium tuberculosis for macrophage infection assays. Methods Mol. Biol. 2017, 1519, 325–331. [Google Scholar] [CrossRef] [Green Version]
- Mira, P.; Yeh, P.; Hall, B.G. Estimating microbial population data from optical density. PLoS ONE 2022, 17. [Google Scholar] [CrossRef]
- USDA. Johne’s Disease. Available online: https://www.aphis.usda.gov/aphis/ourfocus/animalhealth/nvap/NVAP-Reference-Guide/Control-and-Eradication/Johnes-Disease (accessed on 21 April 2023).
- Carter, M.A. Prevalence and prevention of paratuberculosis in North America. Jpn J. Vet. Res. 2012, 60, S9–S18. [Google Scholar]
- Bannantine, J.P.; Talaat, A.M. Controlling Johne’s disease: Vaccination is the way forward. Front. Cell Infect. Microbiol. 2015, 5, 2. [Google Scholar] [CrossRef]
- Coad, M.; Clifford, D.J.; Vordermeier, H.M.; Whelan, A.O. The consequences of vaccination with the Johne’s disease vaccine, Gudair, on diagnosis of bovine tuberculosis. Vet. Rec. 2013, 172, 266. [Google Scholar] [CrossRef]
- Hines, M.E., 2nd; Turnquist, S.E.; Ilha, M.R.; Rajeev, S.; Jones, A.L.; Whittington, L.; Bannantine, J.P.; Barletta, R.G.; Grohn, Y.T.; Katani, R.; et al. Evaluation of novel oral vaccine candidates and validation of a caprine model of Johne’s disease. Front. Cell Infect. Microbiol. 2014, 4, 26. [Google Scholar] [CrossRef] [Green Version]
- Cirillo, J.D.; Stover, C.K.; Bloom, B.R.; Jacobs, W.R., Jr.; Barletta, R.G. Bacterial vaccine vectors and bacillus Calmette-Guerin. Clin. Infect. Dis. 1995, 20, 1001–1009. [Google Scholar] [CrossRef]
- Bannantine, J.P.; Everman, J.L.; Rose, S.J.; Babrak, L.; Katani, R.; Barletta, R.G.; Talaat, A.M.; Grohn, Y.T.; Chang, Y.F.; Kapur, V.; et al. Evaluation of eight live attenuated vaccine candidates for protection against challenge with virulent Mycobacterium avium subspecies paratuberculosis in mice. Front. Cell Infect. Microbiol. 2014, 4, 88. [Google Scholar] [CrossRef] [Green Version]
- Eshraghisamani, R.; Arrazuria, R.; Luo, L.; De Buck, J. Evaluation of Mycobacterium avium subsp. paratuberculosis isocitrate lyase (IcL) and ABC transporter (BacA) knockout mutants as vaccine candidates. Front. Cell Infect. Microbiol. 2023, 13. [Google Scholar] [CrossRef]
- Kabara, E.; Kloss, C.C.; Wilson, M.; Tempelman, R.J.; Sreevatsan, S.; Janagama, H.; Coussens, P.M. A large-scale study of differential gene expression in monocyte-derived macrophages infected with several strains of Mycobacterium avium subspecies paratuberculosis. Brief. Funct. Genomics 2010, 9, 220–237. [Google Scholar] [CrossRef] [Green Version]
- Baranov, M.V.; Kumar, M.; Sacanna, S.; Thutupalli, S.; van den Bogaart, G. Modulation of Immune Responses by Particle Size and Shape. Front. Immunol. 2020, 11, 607945. [Google Scholar] [CrossRef]
- Doshi, N.; Mitragotri, S. Macrophages recognize size and shape of their targets. PLoS ONE 2010, 5, e10051. [Google Scholar] [CrossRef] [Green Version]
- van Genderen, H.O.; Kenis, H.; Hofstra, L.; Narula, J.; Reutelingsperger, C.P. Extracellular annexin A5: Functions of phosphatidylserine-binding and two-dimensional crystallization. Biochim Biophys Acta-Mol. Cell Res. 2008, 1783, 953–963. [Google Scholar] [CrossRef] [Green Version]
- Martin, S.J.; Reutelingsperger, C.P.; McGahon, A.J.; Rader, J.A.; van Schie, R.C.; LaFace, D.M.; Green, D.R. Early redistribution of plasma membrane phosphatidylserine is a general feature of apoptosis regardless of the initiating stimulus: Inhibition by overexpression of Bcl-2 and Abl. J. Exp. Med. 1995, 182, 1545–1556. [Google Scholar] [CrossRef] [Green Version]
- Cooley, R.; Kara, N.; Hui, N.S.; Tart, J.; Roustan, C.; George, R.; Hancock, D.C.; Binkowski, B.F.; Wood, K.V.; Ismail, M.; et al. Development of a cell-free split-luciferase biochemical assay as a tool for screening for inhibitors of challenging protein-protein interaction targets. Wellcome Open Res. 2020, 5. [Google Scholar] [CrossRef]
- Niles, A.; Riss, T. Multiplexed viability, cytotoxicity, and caspase activity assays. Methods Mol. Biol. 2014, 1219, 21–33. [Google Scholar] [CrossRef]
- Atale, N.; Gupta, S.; Yadav, U.C.; Rani, V. Cell-death assessment by fluorescent and nonfluorescent cytosolic and nuclear staining techniques. J. Microsc. 2014, 255, 7–19. [Google Scholar] [CrossRef]
- Telford, W.G. Multiparametric analysis of apoptosis by flow cytometry. Methods Mol. Biol. 2018, 1678, 167–202. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Barletta, R.G.; Bannantine, J.P.; Stabel, J.R.; Muthukrishnan, E.; Anderson, D.K.; Dutta, E.; Manthena, V.; Hanafy, M.; Zinniel, D.K. Mycobacterium avium subsp. paratuberculosis Candidate Vaccine Strains Are Pro-apoptotic in RAW 264.7 Murine Macrophages. Vaccines 2023, 11, 1085. https://doi.org/10.3390/vaccines11061085
Barletta RG, Bannantine JP, Stabel JR, Muthukrishnan E, Anderson DK, Dutta E, Manthena V, Hanafy M, Zinniel DK. Mycobacterium avium subsp. paratuberculosis Candidate Vaccine Strains Are Pro-apoptotic in RAW 264.7 Murine Macrophages. Vaccines. 2023; 11(6):1085. https://doi.org/10.3390/vaccines11061085
Chicago/Turabian StyleBarletta, Raul G., John P. Bannantine, Judith R. Stabel, Ezhumalai Muthukrishnan, Dirk K. Anderson, Enakshy Dutta, Vamsi Manthena, Mostafa Hanafy, and Denise K. Zinniel. 2023. "Mycobacterium avium subsp. paratuberculosis Candidate Vaccine Strains Are Pro-apoptotic in RAW 264.7 Murine Macrophages" Vaccines 11, no. 6: 1085. https://doi.org/10.3390/vaccines11061085