
A Perspective on Oral Immunotherapeutic Tools and Strategies for Autoimmune Disorders



{kind=link}
Abstract
:1. Introduction
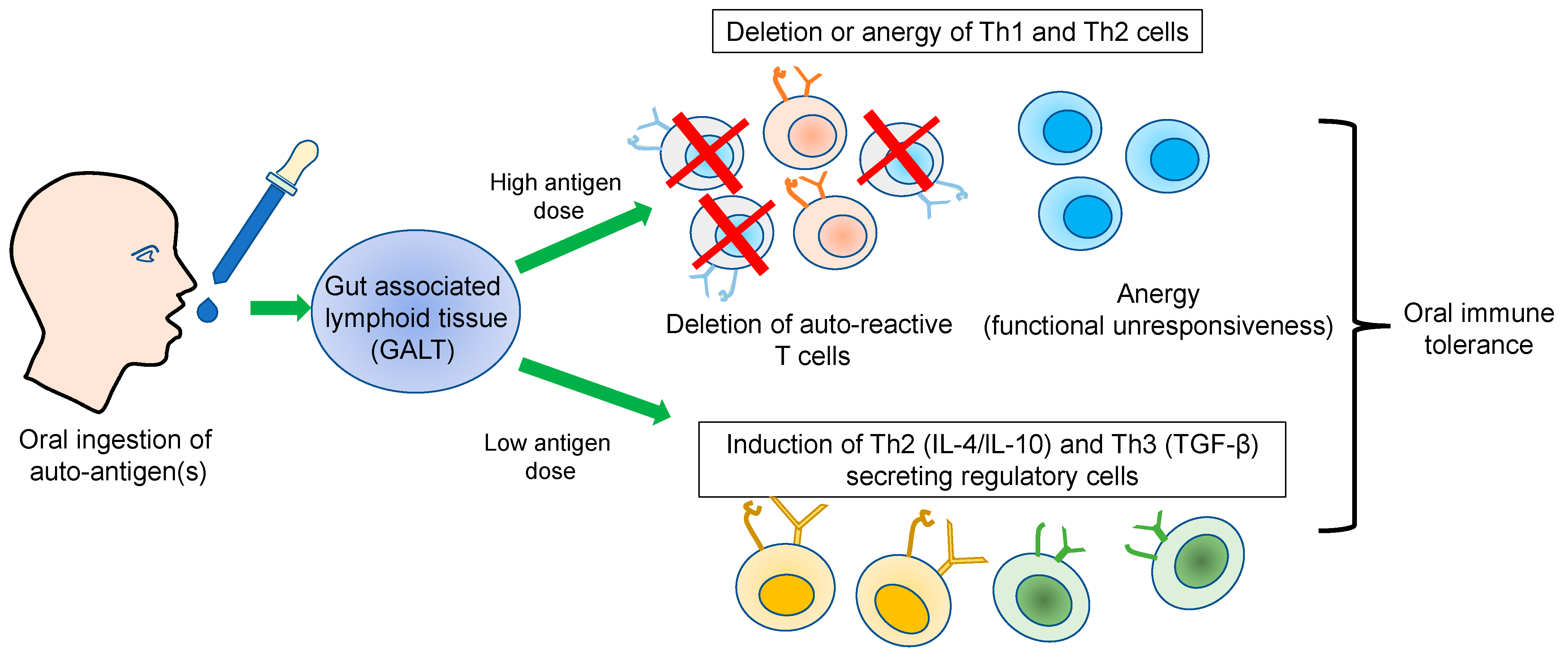
2. Oral Immune Tolerance
3. Immunological Basis of Oral Tolerance
3.1. Dose Effect
3.2. Bystander Effect
4. Tools and Approaches for Antigen-Specific Oral Immune Tolerance
4.1. Synthetic Nanoparticles
4.2. Plant-Based Antigen Delivery
5. Antigen Nonspecific Oral Immune Tolerance
5.1. Therapeutics
5.2. Probiotics
5.3. Plant-Based Products
6. Conclusions and Future Prospects
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, L.; Wang, F.S.; Gershwin, M.E. Human autoimmune diseases: A comprehensive update. J. Intern. Med. 2015, 278, 369–395. [Google Scholar] [CrossRef]
- Aaron Lerner, P.J.; Matthias, T. The World Incidence and Prevalence of Autoimmune Diseases is Increasing. Int. J. Celiac Dis. 2015, 3, 151–155. [Google Scholar] [CrossRef]
- Meier, F.M.; Frerix, M.; Hermann, W.; Muller-Ladner, U. Current immunotherapy in rheumatoid arthritis. Immunotherapy 2013, 5, 955–974. [Google Scholar] [CrossRef]
- Fujita, H.; Soyka, M.B.; Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy. Clin. Transl. Allergy 2012, 2, 2. [Google Scholar] [CrossRef] [PubMed]
- Nelson, H.S. Allergen immunotherapy now and in the future. Allergy Asthma Proc. 2016, 37, 268–272. [Google Scholar] [CrossRef]
- Anderson, R.P.; Jabri, B. Vaccine against autoimmune disease: Antigen-specific immunotherapy. Curr. Opin. Immunol. 2013, 25, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Gao, W.; Awasthi, A.; Jager, A.; Strom, T.B.; Oukka, M.; Kuchroo, V.K. IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells. Nature 2007, 448, 484–487. [Google Scholar] [CrossRef]
- Schaffert, H.; Pelz, A.; Saxena, A.; Losen, M.; Meisel, A.; Thiel, A.; Kohler, S. IL-17-producing CD4(+) T cells contribute to the loss of B-cell tolerance in experimental autoimmune myasthenia gravis. Eur. J. Immunol. 2015, 45, 1339–1347. [Google Scholar] [CrossRef]
- Nemazee, D. Mechanisms of central tolerance for B cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef]
- Mostarica-Stojkovic, M. Mechanisms of the induction of autoimmunity. Srp. Arh. Celok. Lek. 2005, 133 (Suppl. 1), 9–15. [Google Scholar] [CrossRef]
- Fang, Q.; Li, T.; Chen, P.; Wu, Y.; Wang, T.; Mo, L.; Ou, J.; Nandakumar, K.S. Comparative Analysis on Abnormal Methylome of Differentially Expressed Genes and Disease Pathways in the Immune Cells of RA and SLE. Front. Immunol. 2021, 12, 668007. [Google Scholar] [CrossRef] [PubMed]
- Harrison, L.C.; Hafler, D.A. Antigen-specific therapy for autoimmune disease. Curr. Opin. Immunol. 2000, 12, 704–711. [Google Scholar] [CrossRef] [PubMed]
- Peakman, M.; Dayan, C.M. Antigen-specific immunotherapy for autoimmune disease: Fighting fire with fire? Immunology 2001, 104, 361–366. [Google Scholar] [CrossRef]
- Levich, J.D.; Parks, D.E.; Weigle, W.O. Tolerance induction in antigen-specific helper T cell clones and lines in vitro. J. Immunol. 1985, 135, 873–878. [Google Scholar] [CrossRef]
- Falb, D.; Briner, T.J.; Sunshine, G.H.; Bourque, C.R.; Luqman, M.; Gefter, M.L.; Kamradt, T. Peripheral tolerance in T cell receptor-transgenic mice: Evidence for T cell anergy. Eur. J. Immunol. 1996, 26, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Auphan, N.; Schonrich, G.; Malissen, M.; Barad, M.; Hammerling, G.; Arnold, B.; Malissen, B.; Schmitt-Verhulst, A.M. Influence of antigen density on degree of clonal deletion in T cell receptor transgenic mice. Int. Immunol. 1992, 4, 541–547. [Google Scholar] [CrossRef] [PubMed]
- Liblau, R.S.; Tisch, R.; Shokat, K.; Yang, X.; Dumont, N.; Goodnow, C.C.; McDevitt, H.O. Intravenous injection of soluble antigen induces thymic and peripheral T-cells apoptosis. Proc. Natl. Acad. Sci. USA 1996, 93, 3031–3036. [Google Scholar] [CrossRef]
- Sytwu, H.K.; Liblau, R.S.; McDevitt, H.O. The roles of Fas/APO-1 (CD95) and TNF in antigen-induced programmed cell death in T cell receptor transgenic mice. Immunity 1996, 5, 17–30. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Wing, K.; Miyara, M. Regulatory T cells-a brief history and perspective. Eur. J. Immunol. 2007, 37 (Suppl. 1), S116–S123. [Google Scholar] [CrossRef]
- Miyara, M.; Yoshioka, Y.; Kitoh, A.; Shima, T.; Wing, K.; Niwa, A.; Parizot, C.; Taflin, C.; Heike, T.; Valeyre, D.; et al. Functional delineation and differentiation dynamics of human CD4+ T cells expressing the FoxP3 transcription factor. Immunity 2009, 30, 899–911. [Google Scholar] [CrossRef]
- Miyara, M.; Wing, K.; Sakaguchi, S. Therapeutic approaches to allergy and autoimmunity based on FoxP3+ regulatory T-cell activation and expansion. J. Allergy Clin. Immunol. 2009, 123, 749–755. [Google Scholar] [CrossRef]
- Bettini, M.; Vignali, D.A. Regulatory T cells and inhibitory cytokines in autoimmunity. Curr. Opin. Immunol. 2009, 21, 612–618. [Google Scholar] [CrossRef]
- Luczynski, W.; Stasiak-Barmuta, A.; Urban, R.; Urban, M.; Florys, B.; Hryszko, M. Lower percentages of T regulatory cells in children with type 1 diabetes-preliminary report. Pediatr. Endocrinol. Diabetes Metab. 2009, 15, 34–38. [Google Scholar]
- Michel, L.; Berthelot, L.; Pettre, S.; Wiertlewski, S.; Lefrere, F.; Braudeau, C.; Brouard, S.; Soulillou, J.P.; Laplaud, D.A. Patients with relapsing-remitting multiple sclerosis have normal Treg function when cells expressing IL-7 receptor alpha-chain are excluded from the analysis. J. Clin. Investig. 2008, 118, 3411–3419. [Google Scholar] [CrossRef] [PubMed]
- Wing, K.; Sakaguchi, S. Regulatory T cells exert checks and balances on self tolerance and autoimmunity. Nat. Immunol. 2010, 11, 7–13. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T. Regulatory T cells: How do they suppress immune responses? Int. Immunol. 2009, 21, 1105–1111. [Google Scholar] [CrossRef]
- Wing, K.; Onishi, Y.; Prieto-Martin, P.; Yamaguchi, T.; Miyara, M.; Fehervari, Z.; Nomura, T.; Sakaguchi, S. CTLA-4 control over Foxp3+ regulatory T cell function. Science 2008, 322, 271–275. [Google Scholar] [CrossRef]
- Mustafa, S.; Hussain, M.F.; Latif, M.; Ijaz, M.; Asif, M.; Hassan, M.; Faisal, M.; Iqbal, F. A Missense Mutation (c.1037 G > C, p. R346P) in PAPSS2 Gene Results in Autosomal Recessive form of Brachyolmia Type 1 (Hobaek Form) in A Consanguineous Family. Genes 2022, 13, 2096. [Google Scholar] [CrossRef] [PubMed]
- Mangan, P.R.; Harrington, L.E.; O’Quinn, D.B.; Helms, W.S.; Bullard, D.C.; Elson, C.O.; Hatton, R.D.; Wahl, S.M.; Schoeb, T.R.; Weaver, C.T. Transforming growth factor-beta induces development of the T(H)17 lineage. Nature 2006, 441, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S. The role of transforming growth factor beta in T helper 17 differentiation. Immunology 2018, 155, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Kehrl, J.H.; Wakefield, L.M.; Roberts, A.B.; Jakowlew, S.; Alvarez-Mon, M.; Derynck, R.; Sporn, M.B.; Fauci, A.S. Production of transforming growth factor beta by human T lymphocytes and its potential role in the regulation of T cell growth. J. Exp. Med. 1986, 163, 1037–1050. [Google Scholar] [CrossRef]
- Chen, Y.; Kuchroo, V.K.; Inobe, J.; Hafler, D.A.; Weiner, H.L. Regulatory T cell clones induced by oral tolerance: Suppression of autoimmune encephalomyelitis. Science 1994, 265, 1237–1240. [Google Scholar] [CrossRef]
- Kaufman, D.L.; Clare-Salzler, M.; Tian, J.; Forsthuber, T.; Ting, G.S.; Robinson, P.; Atkinson, M.A.; Sercarz, E.E.; Tobin, A.J.; Lehmann, P.V. Spontaneous loss of T-cell tolerance to glutamic acid decarboxylase in murine insulin-dependent diabetes. Nature 1993, 366, 69–72. [Google Scholar] [CrossRef]
- Carambia, A.; Freund, B.; Schwinge, D.; Heine, M.; Laschtowitz, A.; Huber, S.; Wraith, D.C.; Korn, T.; Schramm, C.; Lohse, A.W.; et al. TGF-beta-dependent induction of CD4(+)CD25(+)Foxp3(+) Tregs by liver sinusoidal endothelial cells. J. Hepatol. 2014, 61, 594–599. [Google Scholar] [CrossRef]
- Umeshappa, C.S.; Singha, S.; Blanco, J.; Shao, K.; Nanjundappa, R.H.; Yamanouchi, J.; Pares, A.; Serra, P.; Yang, Y.; Santamaria, P. Suppression of a broad spectrum of liver autoimmune pathologies by single peptide-MHC-based nanomedicines. Nat. Commun. 2019, 10, 2150. [Google Scholar] [CrossRef]
- Groschwitz, K.R.; Hogan, S.P. Intestinal barrier function: Molecular regulation and disease pathogenesis. J. Allergy Clin. Immunol. 2009, 124, 3–20. [Google Scholar] [CrossRef]
- Zhu, G.; Xu, Y.; Cen, X.; Nandakumar, K.S.; Liu, S.; Cheng, K. Targeting pattern-recognition receptors to discover new small molecule immune modulators. Eur. J. Med. Chem. 2018, 144, 82–92. [Google Scholar] [CrossRef] [PubMed]
- Gottschalk, R.A.; Martins, A.J.; Angermann, B.R.; Dutta, B.; Ng, C.E.; Uderhardt, S.; Tsang, J.S.; Fraser, I.D.; Meier-Schellersheim, M.; Germain, R.N. Distinct NF-kappaB and MAPK Activation Thresholds Uncouple Steady-State Microbe Sensing from Anti-pathogen Inflammatory Responses. Cell. Syst. 2016, 2, 378–390. [Google Scholar] [CrossRef]
- Clark, M.A.; Jepson, M.A.; Hirst, B.H. Exploiting M cells for drug and vaccine delivery. Adv. Drug. Deliv. Rev. 2001, 50, 81–106. [Google Scholar] [CrossRef] [PubMed]
- Bland, P.W.; Warren, L.G. Antigen presentation by epithelial cells of the rat small intestine. II. Selective induction of suppressor T cells. Immunology 1986, 58, 9–14. [Google Scholar] [PubMed]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2001, 2, 361–367. [Google Scholar] [CrossRef]
- Campbell, D.J.; Debes, G.F.; Johnston, B.; Wilson, E.; Butcher, E.C. Targeting T cell responses by selective chemokine receptor expression. Semin. Immunol. 2003, 15, 277–286. [Google Scholar] [CrossRef]
- Miller, A.; Lider, O.; Roberts, A.B.; Sporn, M.B.; Weiner, H.L. Suppressor T cells generated by oral tolerization to myelin basic protein suppress both in vitro and in vivo immune responses by the release of transforming growth factor beta after antigen-specific triggering. Proc. Natl. Acad. Sci. USA 1992, 89, 421–425. [Google Scholar] [CrossRef]
- Whitacre, C.C.; Gienapp, I.E.; Orosz, C.G.; Bitar, D.M. Oral tolerance in experimental autoimmune encephalomyelitis. III. Evidence for clonal anergy. J. Immunol. 1991, 147, 2155–2163. [Google Scholar] [CrossRef]
- Chambers, C.A.; Sullivan, T.J.; Truong, T.; Allison, J.P. Secondary but not primary T cell responses are enhanced in CTLA-4-deficient CD8+ T cells. Eur. J. Immunol. 1998, 28, 3137–3143. [Google Scholar] [CrossRef]
- Lider, O.; Santos, L.M.; Lee, C.S.; Higgins, P.J.; Weiner, H.L. Suppression of experimental autoimmune encephalomyelitis by oral administration of myelin basic protein. II. Suppression of disease and in vitro immune responses is mediated by antigen-specific CD8+ T lymphocytes. J. Immunol. 1989, 142, 748–752. [Google Scholar] [CrossRef]
- Barone, K.S.; Herms, B.; Karlosky, L.; Murray, S.; Qualls, J. Effect of in vivo administration of anti-CTLA-4 monoclonal antibody and IL-12 on the induction of low-dose oral tolerance. Clin. Exp. Immunol. 2002, 130, 196–203. [Google Scholar]
- Wilkes, D.S.; Chew, T.; Flaherty, K.R.; Frye, S.; Gibson, K.F.; Kaminski, N.; Klemsz, M.J.; Lange, W.; Noth, I.; Rothhaar, K. Oral immunotherapy with type V collagen in idiopathic pulmonary fibrosis. Eur. Respir. J. 2015, 45, 1393–1402. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.; Lider, O.; Weiner, H.L. Antigen-driven bystander suppression after oral administration of antigens. J. Exp. Med. 1991, 174, 791–798. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jin, W.; Cook, M.; Weiner, H.L.; Wahl, S.M. Oral delivery of group A streptococcal cell walls augments circulating TGF-beta and suppresses streptococcal cell wall arthritis. J. Immunol. 1998, 161, 6297–6304. [Google Scholar] [CrossRef] [PubMed]
- Pham, M.N.; Gibson, C.; Ryden, A.K.; Perdue, N.; Boursalian, T.E.; Pagni, P.P.; Coppieters, K.; Skonberg, C.; Porsgaard, T.; von Herrath, M.; et al. Oral insulin (human, murine, or porcine) does not prevent diabetes in the non-obese diabetic mouse. Clin. Immunol. 2016, 164, 28–33. [Google Scholar] [CrossRef]
- Kim, W.U.; Lee, W.K.; Ryoo, J.W.; Kim, S.H.; Kim, J.; Youn, J.; Min, S.Y.; Bae, E.Y.; Hwang, S.Y.; Park, S.H.; et al. Suppression of collagen-induced arthritis by single administration of poly(lactic-co-glycolic acid) nanoparticles entrapping type II collagen: A novel treatment strategy for induction of oral tolerance. Arthritis Rheum. 2002, 46, 1109–1120. [Google Scholar] [CrossRef]
- Haggag, Y.; Abdel-Wahab, Y.; Ojo, O.; Osman, M.; El-Gizawy, S.; El-Tanani, M.; Faheem, A.; McCarron, P. Preparation and in vivo evaluation of insulin-loaded biodegradable nanoparticles prepared from diblock copolymers of PLGA and PEG. Int. J. Pharm. 2016, 499, 236–246. [Google Scholar] [CrossRef]
- Salvioni, L.; Fiandra, L.; Del Curto, M.D.; Mazzucchelli, S.; Allevi, R.; Truffi, M.; Sorrentino, L.; Santini, B.; Cerea, M.; Palugan, L.; et al. Oral delivery of insulin via polyethylene imine-based nanoparticles for colonic release allows glycemic control in diabetic rats. Pharmacol. Res. 2016, 110, 122–130. [Google Scholar] [CrossRef]
- Jabri, T.R.T.; Razzak, A.; Aziz, S.; Imran, M.; Sikandar, B.; Elhissi, A.; Shafiullah Aslam, S.M.; RazaShah, M. Fabrication of hesperidin hybrid lecithin-folic acid silver nanoparticles and its evaluation as anti-arthritis formulation in autoimmune arthritic rat model. J. Mol. Struct. 2023, 1276, 134722. [Google Scholar] [CrossRef]
- Raimondi, G.Z.Y.; Hou, S.; Wang, J.; Calderon-Colon, X.; Tiburzi, O.; Igleasias Lozano, M.; Patrone, J. Prevention of Type 1 Diabetes via a Lipid Nanoparticle-Based Oral Immunotherapy that Targets Selective Lymphoid Tissues. J. Immunol. 2022, 208, 174.12. [Google Scholar] [CrossRef]
- Ma, S.; Huang, Y.; Yin, Z.; Menassa, R.; Brandle, J.E.; Jevnikar, A.M. Induction of oral tolerance to prevent diabetes with transgenic plants requires glutamic acid decarboxylase (GAD) and IL-4. Proc. Natl. Acad. Sci. USA 2004, 101, 5680–5685. [Google Scholar] [CrossRef]
- Ma, S.W.; Zhao, D.L.; Yin, Z.Q.; Mukherjee, R.; Singh, B.; Qin, H.Y.; Stiller, C.R.; Jevnikar, A.M. Transgenic plants expressing autoantigens fed to mice to induce oral immune tolerance. Nat. Med. 1997, 3, 793–796. [Google Scholar] [CrossRef] [PubMed]
- Ruhlman, T.; Ahangari, R.; Devine, A.; Samsam, M.; Daniell, H. Expression of cholera toxin B-proinsulin fusion protein in lettuce and tobacco chloroplasts--oral administration protects against development of insulitis in non-obese diabetic mice. Plant. Biotechnol. J. 2007, 5, 495–510. [Google Scholar] [CrossRef]
- Iizuka, M.; Wakasa, Y.; Tsuboi, H.; Asashima, H.; Hirota, T.; Kondo, Y.; Matsumoto, I.; Sumida, T.; Takaiwa, F. Prophylactic effect of the oral administration of transgenic rice seeds containing altered peptide ligands of type II collagen on rheumatoid arthritis. Biosci. Biotechnol. Biochem. 2014, 78, 1662–1668. [Google Scholar] [CrossRef] [PubMed]
- Hansson, C.; Schon, K.; Kalbina, I.; Strid, A.; Andersson, S.; Bokarewa, M.I.; Lycke, N.Y. Feeding transgenic plants that express a tolerogenic fusion protein effectively protects against arthritis. Plant. Biotechnol. J. 2016, 14, 1106–1115. [Google Scholar] [CrossRef]
- Dittel, L.J.; Dittel, B.N.; Brod, S.A. Ingested ACTH blocks Th17 production by inhibiting GALT IL-6. J. Neurol. Sci. 2020, 409, 116602. [Google Scholar] [CrossRef]
- Dittel, L.J.; Dittel, B.N.; Brod, S.A. Ingested (Oral) Adrenocorticotropic Hormone Inhibits IL-17 in the Central Nervous System in the Mouse Model of Multiple Sclerosis and Experimental Autoimmune Encephalomyelitis. Immunohorizons 2022, 6, 497–506. [Google Scholar] [CrossRef]
- Brod, S.A.; Bauer, V.L. Ingested (oral) tocilizumab inhibits EAE. Cytokine 2014, 68, 86–93. [Google Scholar] [CrossRef]
- Brod, S.A. Ingested (oral) anti-IL-12/23 inhibits EAE. J. Neurol. Sci. 2016, 361, 19–25. [Google Scholar] [CrossRef]
- Fang, X.; Liu, C.; Zhang, K.; Yang, W.; Wu, Z.; Shen, S.; Ma, Y.; Lu, X.; Chen, Y.; Lu, T.; et al. Discovery of orally active 1,4,5,6,8-pentaazaacenaphthylens as novel, selective, and potent covalent BTK inhibitors for the treatment of rheumatoid arthritis. Eur. J. Med. Chem. 2023, 246, 114940. [Google Scholar] [CrossRef] [PubMed]
- Salehipour, Z.; Haghmorad, D.; Sankian, M.; Rastin, M.; Nosratabadi, R.; Soltan Dallal, M.M.; Tabasi, N.; Khazaee, M.; Nasiraii, L.R.; Mahmoudi, M. Bifidobacterium animalis in combination with human origin of Lactobacillus plantarum ameliorate neuroinflammation in experimental model of multiple sclerosis by altering CD4+ T cell subset balance. Biomed. Pharmacother. 2017, 95, 1535–1548. [Google Scholar] [CrossRef]
- Ibrahim, H.I.M.; Sheikh, A.; Khalil, H.E.; Khalifa, A. Bacillus amyloliquifaciens-Supplemented Camel Milk Suppresses Neuroinflammation of Autoimmune Encephalomyelitis in a Mouse Model by Regulating Inflammatory Markers. Nutrients 2023, 15, 550. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhu, D.; Ortiz-Velez, L.C.; Perry, J.L.; Pennington, M.W.; Hyser, J.M.; Britton, R.A.; Beeton, C. A bioengineered probiotic for the oral delivery of a peptide Kv1.3 channel blocker to treat rheumatoid arthritis. Proc. Natl. Acad. Sci. USA 2023, 120, e2211977120. [Google Scholar] [CrossRef] [PubMed]
- Guimaraes, M.A.F.; Pinheiro-Rosa, N.; Oliveira, R.P.; Aguiar, S.L.F.; Miranda, M.C.G.; Lemos, L.; Souza, A.L.; Dos Reis, D.S.; Medeiros, S.R.; Goncalves, W.A.; et al. Hsp65-producing Lactococcus lactis inhibits experimental autoimmune encephalomyelitis by preventing cell migration into spinal cord. Cell. Immunol. 2023, 384, 104661. [Google Scholar] [CrossRef]
- Ying, S.; Yang, H.; Gu, Q.; Wu, Z.; Zou, N.; Wang, C.Z.; Wan, C.; Yuan, C.S. The Small-Molecule compound baicalein alleviates experimental autoimmune encephalomyelitis by suppressing pathogenetic CXCR6(+) CD4 cells. Int. Immunopharmacol. 2023, 114, 109562. [Google Scholar] [CrossRef] [PubMed]
- Que, W.; Lin, H.; Li, X.; Zhang, B.; Liu, M.; Hu, X.; Fu, J.; Cheng, Y.; Qiu, H. Koumine ameliorates concanavalin A-induced autoimmune hepatitis in mice: Involvement of the Nrf2, NF-kappaB pathways, and gut microbiota. Int. Immunopharmacol. 2023, 114, 109573. [Google Scholar] [CrossRef] [PubMed]
- Nevius, E.; Srivastava, P.K.; Basu, S. Oral ingestion of Capsaicin, the pungent component of chili pepper, enhances a discreet population of macrophages and confers protection from autoimmune diabetes. Mucosal Immunol. 2012, 5, 76–86. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shakya, A.K.; Mallick, B.; Nandakumar, K.S. A Perspective on Oral Immunotherapeutic Tools and Strategies for Autoimmune Disorders. Vaccines 2023, 11, 1031. https://doi.org/10.3390/vaccines11061031
Shakya AK, Mallick B, Nandakumar KS. A Perspective on Oral Immunotherapeutic Tools and Strategies for Autoimmune Disorders. Vaccines. 2023; 11(6):1031. https://doi.org/10.3390/vaccines11061031
Chicago/Turabian StyleShakya, Akhilesh Kumar, Buddhadev Mallick, and Kutty Selva Nandakumar. 2023. "A Perspective on Oral Immunotherapeutic Tools and Strategies for Autoimmune Disorders" Vaccines 11, no. 6: 1031. https://doi.org/10.3390/vaccines11061031