
Liposome and QS-21 Combined Adjuvant Induces theHumoral and Cellular Responses of Acellular Pertussis Vaccine in a Mice Model

Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. The Preparation of aP Vaccine
2.3. Mouse Immunization
2.4. Serological Assays
2.4.1. Pertussis Antigen-Specific Antibodies Titer Detected by ELISA
2.4.2. PT Neutralization Assay
2.4.3. IgG Subtyping
2.5. Detection of Respiratory Tissue-Resident T Cells
2.6. Flow Cytometry Analysis
2.7. Statistical Analysis
3. Results
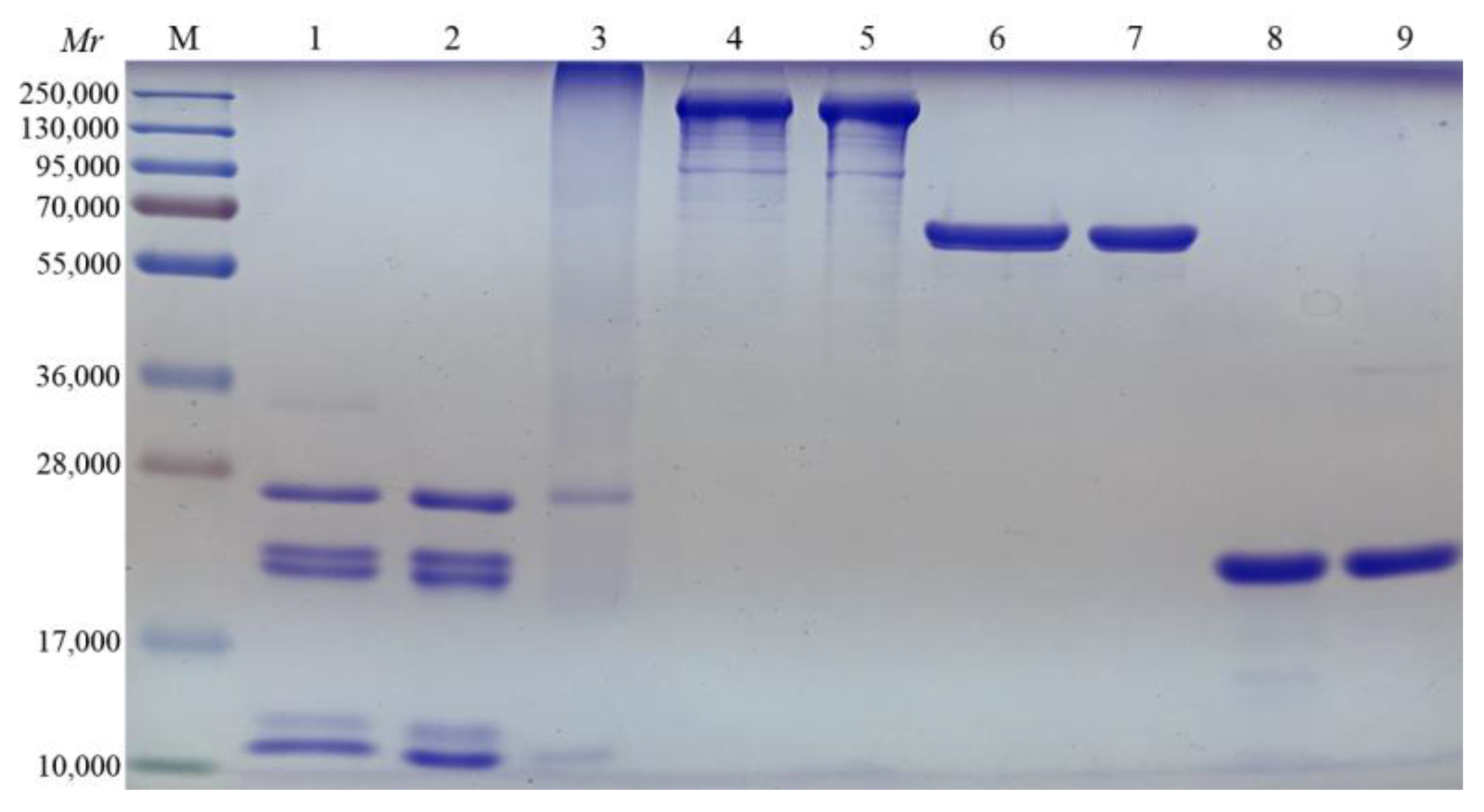
3.1. Pertussis Antigen and Adjuvant Formulations Characterization
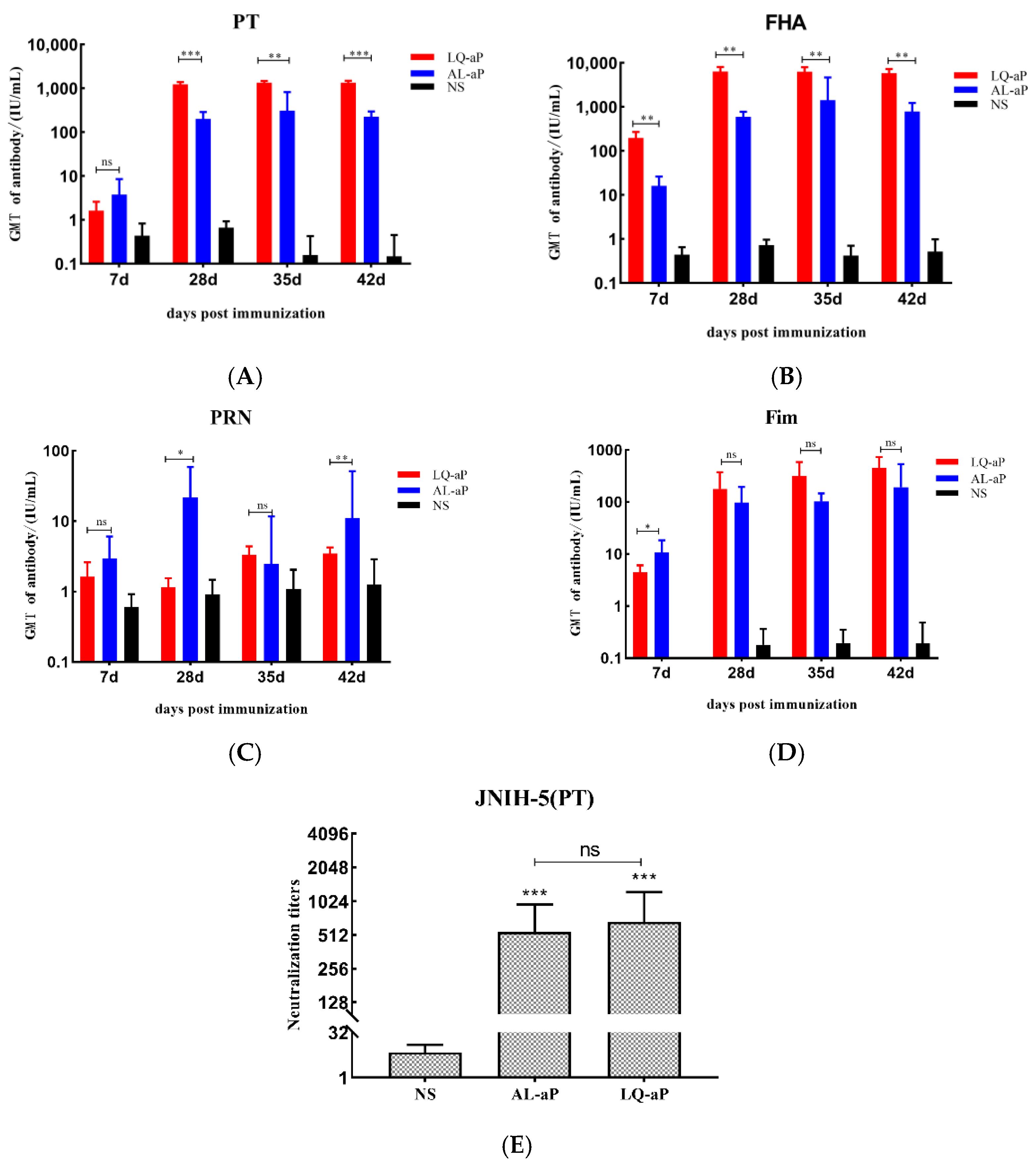
3.2. Serum IgG Responses to Pertussis Vaccines
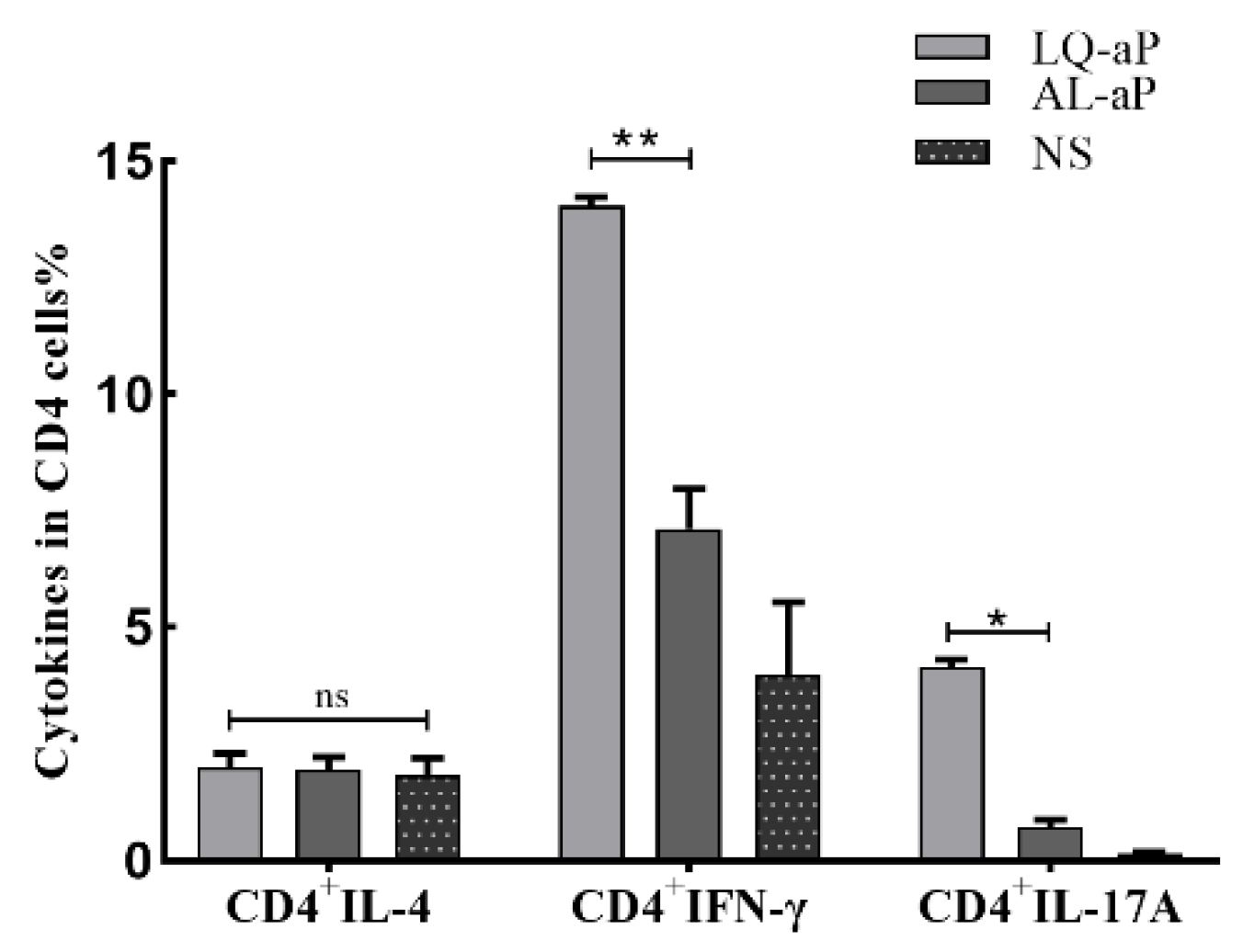
3.3. The Proportion of CD4+ Cells in T Lymphocytes after LQ-aP Immunization
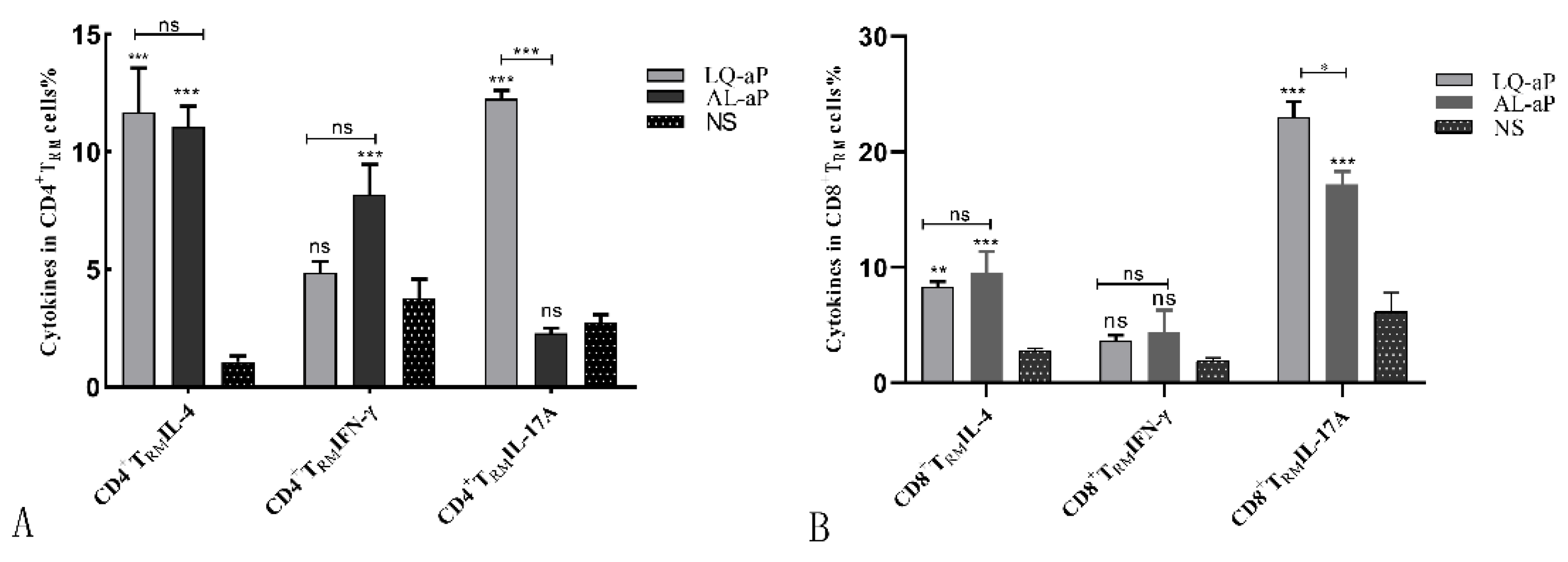
3.4. CD4 +TRM Cells Induced in the Lung Tissue by the LQ-ap Vaccination Group Potently Secrete IFN-γ
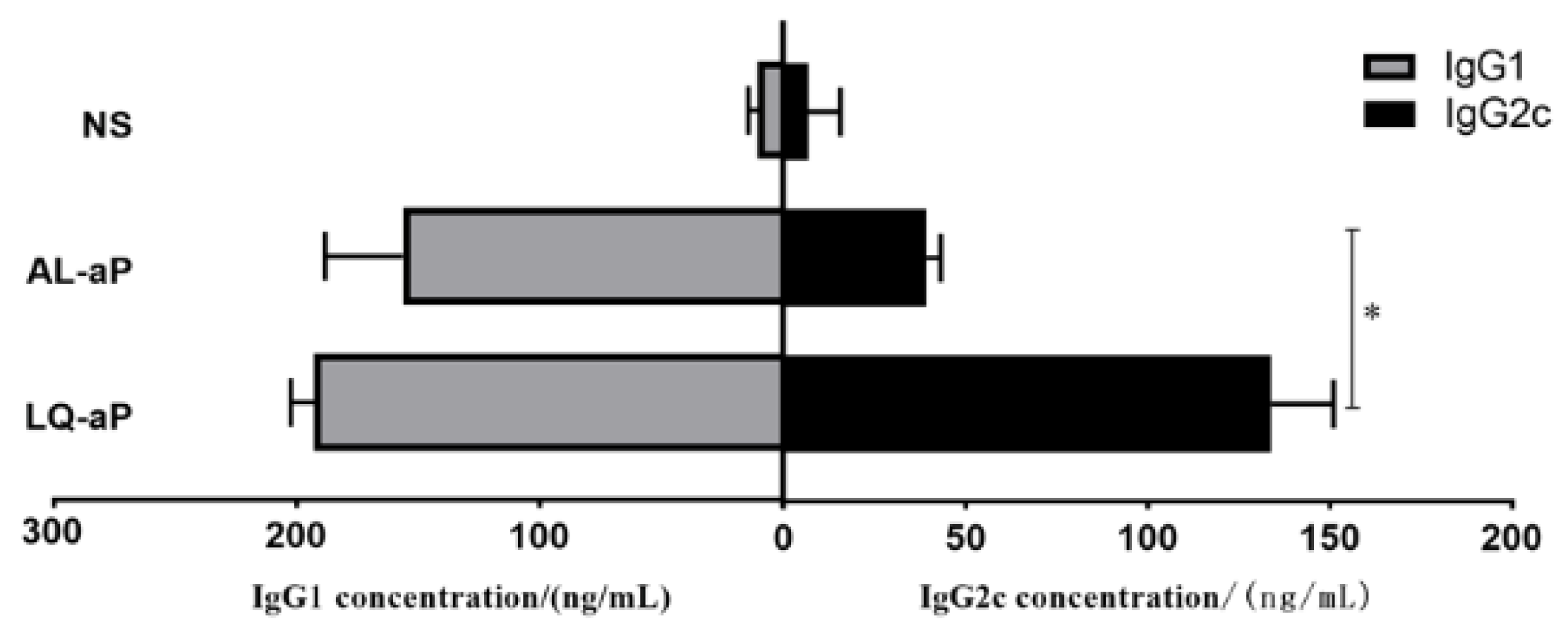
3.5. Shift in the IgG1/IgG2c Profile
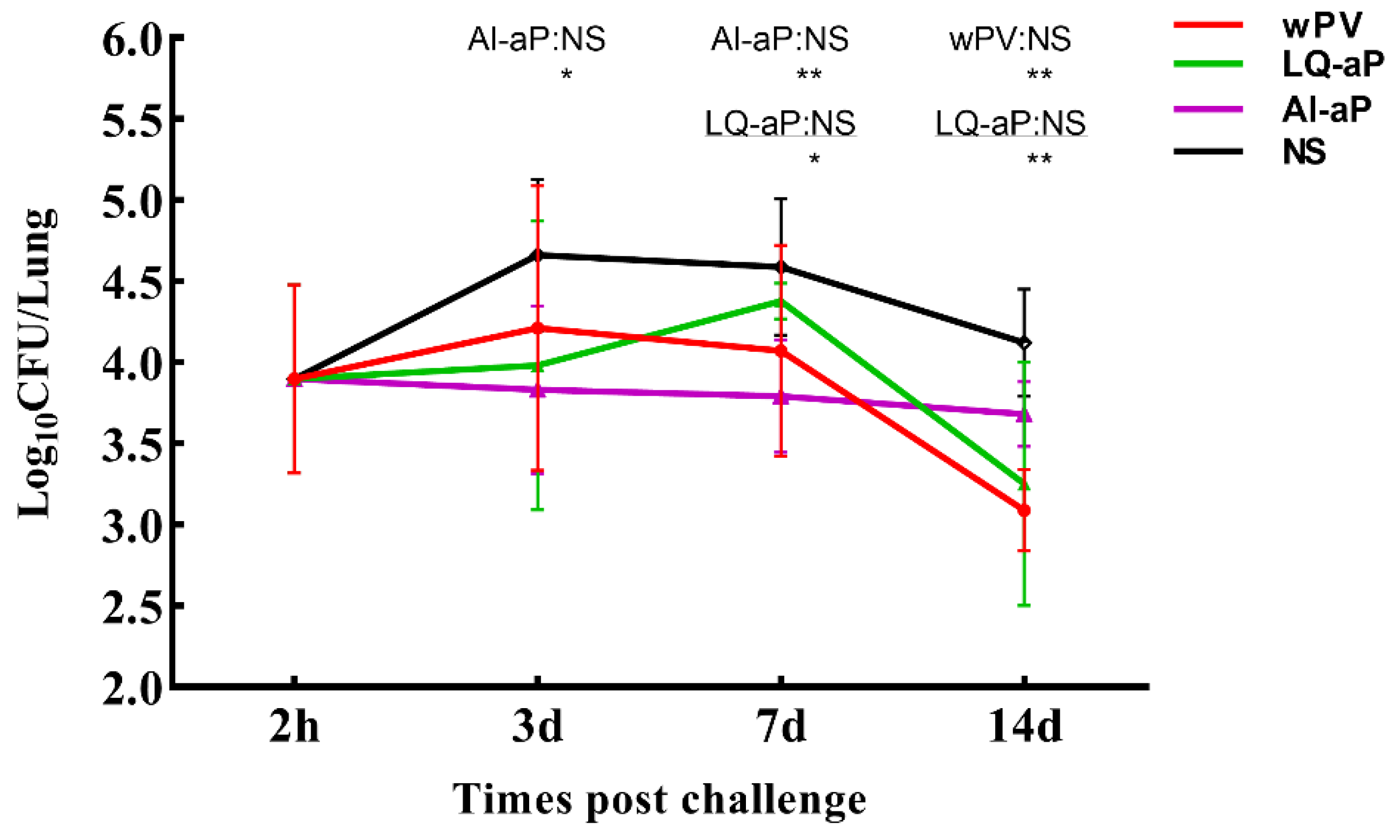
3.6. Results of Intranasal Challenge Protection against Pertussis in Mice
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| aP | Acellular pertussis |
| CD | Cluster of Differentiation |
| CFU | Colony forming unit |
| CpG | Cytosine phosphoguanine |
| FHA | Filamentous hemagglutinin |
| Fim2/3 | Fimbriae type 2 and type 3 |
| IFN-γ | Interferon γ |
| PRN | Pertussis Pertactin |
| PT | Pertussis toxin |
| SD | Standard deviation |
| TRM | Tissue-resident memory T |
| wP | Whole-cell pertussis |
References
- Jia, J.H.; Guo, Q.; Wan, C.M. Resurgence and vaccine strategies of pertussis. Zhonghua Er Ke Za Zhi 2020, 58, 686–689. [Google Scholar] [PubMed]
- Winter, K.; Glaser, C.; Watt, J.; Harriman, K. Pertussis epidemic—California, 2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 1129–1132. [Google Scholar] [PubMed]
- Hara, M.; Fukuoka, M.; Tashiro, K.; Ozaki, I.; Ohfuji, S.; Okada, K.; Nakano, T.; Fukushima, W.; Hirota, Y. Pertussis outbreak in university students and evaluation of acellular pertussis vaccine effectiveness in Japan. BMC Infect. Dis. 2015, 15, 45. [Google Scholar] [CrossRef]
- Menzies, R.; Mcintyre, P.; Beard, F. Vaccine preventable diseases and vaccination coverage in Aboriginal and Torres Strait Islander people, Australia, 1999 to 2002. Commun. Dis. Intell. Q. Rep. 2004, 28, 127–159. [Google Scholar] [PubMed]
- Liu, T.C.; Zhang, J.; Liu, S.Q.; Yin, A.T.; Ruan, S.M. Evaluation of immunisation strategies for pertussis vaccines in Jinan, China—An interrupted time-series study. Epidemiol. Infect. 2020, 148, e26. [Google Scholar] [CrossRef]
- Klein, N.P.; Bartlett, J.; Rowhani-Rahbar, A.; Fireman, B.; Baxter, R. Waning protection after fifth dose of acellular pertussis vaccine in children. N. Engl. J. Med. 2012, 367, 1012–1019. [Google Scholar] [CrossRef]
- Safarchi, A.; Octavia, S.; Luu, L.D.W.; Tay, C.Y.; Sintchenko, V.; Wood, N.; Marshall, H.; McIntyre, P.; Lan, R. Pertactin negative Bordetella pertussis demonstrates higher fitness under vaccine selection pressure in a mixed infection model. Vaccine 2015, 33, 6277–6281. [Google Scholar] [CrossRef]
- Dorji, D.; Mooi, F.; Yantorno, O.; Deora, R.; Graham, R.M.; Mukkur, T.K. Bordetella pertussis virulence factors in the continuing evolution of whooping cough vaccines for improved performance. Med. Microbiol. Immunol. 2018, 207, 3–26. [Google Scholar]
- Ausiello, C.M.; Urbani, F.; La Sala, A.; Lande, R.; Cassone, A. Vaccine- and antigen-dependent type 1 and type 2 cytokine induction after primary vaccination of infants with whole-cell or acellular pertussis vaccines. Infect. Immun. 1997, 65, 2168–2174. [Google Scholar] [CrossRef]
- Mcloughlin, R.; Lavelle, E.; Mills, K.; Sutton, C.; Higgins, S.; Allen, A. Relative contribution of Th1 and Th17 cells in adaptive immunity to Bordetella pertussis: Towards the rational design of an improved acellular pertussis vaccine. PLoS Pathog. 2013, 9, e1003264. [Google Scholar]
- Mahon, B.P.; Sheahan, B.J.; Griffin, F.; Murphy, G.; Mills, K.H. Atypical disease after Bordetella pertussis respiratory infection of mice with targeted disruptions of interferon-gamma receptor or immunoglobulin mu chain genes. J. Exp. Med. 1997, 186, 1843–1851. [Google Scholar] [CrossRef] [PubMed]
- Warfel, J.M.; Zimmerman, L.I.; Merkel, T.J. Acellular pertussis vaccines protect against disease but fail to prevent infection and transmission in a nonhuman primate model. Proc. Natl. Acad. Sci. USA 2014, 111, 787–792. [Google Scholar] [CrossRef] [PubMed]
- Pizzolla, A.; Nguyen, T.H.; Smith, J.M.; Brooks, A.G.; Kedzierska, K.; Heath, W.R.; Reading, P.C.; Wakim, L.M. Resident memory CD8+ T cells in the upper respiratory tract prevent pulmonary influenza virus infection. Sci. Immunol. 2017, 2, eaam6970. [Google Scholar] [CrossRef] [PubMed]
- Wilk, M.M.; Borkner, L.; Misiak, A.; Curham, L.; Allen, A.C.; Mills, K.H. Immunization with whole cell but not acellular pertussis vaccines primes CD4 TRM cells that sustain protective immunity against nasal colonization with Bordetella pertussis. Emerg. Microbes Infect. 2019, 8, 169–185. [Google Scholar] [CrossRef] [PubMed]
- Higgs, R.; Higgins, S.C.; Ross, P.J.; Mills, K.H.G. Immunity to the respiratory pathogen Bordetella pertussis. Mucosal. Immunol. 2012, 5, 485–500. [Google Scholar] [PubMed]
- Asokanathan, C.; Corbel, M.; Xing, D. A CpG-containing oligodeoxynucleotide adjuvant for acellular pertussis vaccine improves the protective response against Bordetella pertussis. Hum. Vaccines Immunother. 2013, 9, 325–331. [Google Scholar] [CrossRef] [PubMed]
- Queenan, A.M.; Dowling, D.J.; Cheng, W.K.; Faé, K.; Fernandez, J.; Flynn, P.J.; Joshi, S.; Brightman, S.E.; Ramirez, J.; Serroyen, J.; et al. Increasing FIM2/3 antigen-content improves efficacy of Bordetella pertussis vaccines in mice in vivo without altering vaccine-induced human reactogenicity biomarkers in vitro. Vaccine 2019, 37, 80–89. [Google Scholar] [CrossRef]
- Dunne, A.; Mielke, L.A.; Allen, A.C.; Sutton, C.E.; Higgs, R.; Cunningham, C.C.; Higgins, S.C.; Mills, K.H.G. A novel TLR2 agonist from Bordetella pertussis is a potent adjuvant that promotes protective immunity with an acellular pertussis vaccine. Mucosal Immunol. 2015, 8, 607–617. [Google Scholar] [CrossRef]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine adjuvants: Understanding the structure and mechanism of adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- GB/T 35892-2018; Laboratory Animal-Guideline for Ethical Review of Animal Welfare. Standardization Administration of China: Beijing, China, 2018.
- Guiso, N.; Capiau, C.; Carletti, G.; Poolman, J.; Hauser, P. Intranasal murine model of Bordetella pertussis infection. I. Prediction of protection in human infants by acellular vaccines. Vaccine 1999, 17, 2366–2376. [Google Scholar] [CrossRef]
- Alexander, F.; Matheson, M.; Fry, N.K.; Labram, B.; Gorringe, A.R. Antibody responses to individual Bordetella pertussis fimbrial antigen Fim2 or Fim3 following immunization with the five-component acellular pertussis vaccine or to pertussis disease. Clin. Vaccine Immunol. 2012, 19, 1776–1783. [Google Scholar] [CrossRef] [PubMed]
- Østergaard, E.; Sørensen, C.; Nielsen, L.; Stawski, G. Elimination of interfering activity in serum samples in the Chinese hamster ovary pertussis serology assay. Clin. Vaccine Immunol. 2008, 15, 1244–1247. [Google Scholar] [CrossRef] [PubMed]
- Anderson, K.G.; Mayer-Barber, K.; Sung, H.; Beura, L.; James, B.R.; Taylor, J.J.; Qunaj, L.; Griffith, T.S.; Vezys, V.; Barber, D.L.; et al. Intravascular staining for discrimination of vascular and tissue leukocytes. Nat. Protoc. 2014, 9, 209–222. [Google Scholar] [CrossRef] [PubMed]
- Agnolon, V.; Bruno, C.; Leuzzi, R.; Galletti, B.; D’Oro, U.; Pizza, M.; Seubert, A.; O’Hagan, D.T.; Baudner, B.C. The potential of adjuvants to improve immune responses against TdaP vaccines: A preclinical evaluation of MF59 and monophosphoryl lipid A. Int. J. Pharm. 2015, 492, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopoeia Commission. Pharmacopoeia of the People’s Republic of China; The Medicine Science and Technology Press of China: Beijing, China, 2020; Volume 3.
- Shapiro, E.D. Acellular vaccines and resurgence of pertussis. JAMA 2012, 308, 2149–2150. [Google Scholar] [CrossRef]
- Diavatopoulos, D.A.; Mills, K.H.; Kester, K.E.; Kampmann, B.; Silerova, M.; Heininger, U.; van Dongen, J.J.; van der Most, R.G.; Huijnen, M.A.; Siena, E.; et al. PERISCOPE: Road towards effective control of pertussis. Lancet Infect. Dis. 2019, 19, e179–e186. [Google Scholar] [CrossRef]
- Dewan, K.K.; Linz, B.; DeRocco, S.E.; Harvill, E.T. Acellular Pertussis Vaccine Components: Today and Tomorrow. Vaccines 2020, 8, 217. [Google Scholar] [CrossRef]
- Brito, L.A.; Malyala, P.; O’Hagan, D.T. Vaccine adjuvant formulations: A pharmaceutical perspective. Semin. Immunol. 2013, 25, 130–145. [Google Scholar] [CrossRef]
- Clapp, T.; Siebert, P.; Chen, D.; Braun, L.J. Vaccines with aluminum-containing adjuvants: Optimizing vaccine efficacy and thermal stability. J. Pharm. Sci. 2011, 100, 388–401. [Google Scholar] [CrossRef]
- Han, S.B.; Kang, K.R.; Huh, D.H.; Lee, H.C.; Lee, S.Y.; Kim, J.H.; Hur, J.K.; Kang, J.H. Preliminary study on the immunogenicity of a newly developed GCC Tdap vaccine and its protection efficacy against Bordetella pertussis in a murine intranasal challenge model. Clin. Exp. Vaccine Res. 2015, 4, 75–82. [Google Scholar] [CrossRef]
- Hogenesch, H. Mechanism of immunopotentiation and safety of aluminum adjuvants. Front. Immunol. 2012, 3, 406. [Google Scholar] [CrossRef] [PubMed]
- Mills, K.H.; Ryan, M.; Ryan, E.; Mahon, B.P. A murine model in which protection correlates with pertussis vaccine efficacy in children reveals complementary roles for humoral and cell-mediated immunity in protection against Bordetella pertussis. Infect. Immun. 1998, 66, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Wilk, M.M.; Mills, K.H. CD4 TRM Cells Following Infection and Immunization: Implications for More Effective Vaccine Design. Front. Immunol. 2018, 9, 1860. [Google Scholar] [CrossRef]
- Wilk, M.M.; Misiak, A.; McManus, R.M.; Allen, A.C.; Lynch, M.A.; Mills, K.H. Lung CD4 Tissue-Resident Memory T Cells Mediate Adaptive Immunity Induced by Previous Infection of Mice with Bordetella pertussis. J. Immunol. 2017, 199, 233–243. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Adjuvant | Antigens | Particle Size (mean ± SD) | Zeta Potential (mean± SD) | Antigen Adsorption (%) |
|---|---|---|---|---|
| Liposome + QS-21 (LQ) | - | 141.80 ± 1.04 nm | −11.57 ± 0.50 mV | n.a |
| + | 148.27 ± 2.25 nm | −3.39 ± 0.32 mV | n.a | |
| Alum | - | 2.61 ± 0.01 μm | 22.53 ± 1.40 mV | n.a |
| + | 6.16 ± 0.15 μm | 10.03 ± 0.56 mV | 97.0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, B.; Zhu, D.; Zhou, Y.; Gong, B.; Hu, Y.; Zhang, J.; Huang, S.; Nian, X.; Li, X.; Li, X.; et al. Liposome and QS-21 Combined Adjuvant Induces theHumoral and Cellular Responses of Acellular Pertussis Vaccine in a Mice Model. Vaccines 2023, 11, 914. https://doi.org/10.3390/vaccines11050914
Yang B, Zhu D, Zhou Y, Gong B, Hu Y, Zhang J, Huang S, Nian X, Li X, Li X, et al. Liposome and QS-21 Combined Adjuvant Induces theHumoral and Cellular Responses of Acellular Pertussis Vaccine in a Mice Model. Vaccines. 2023; 11(5):914. https://doi.org/10.3390/vaccines11050914
Chicago/Turabian StyleYang, Baifeng, Dewu Zhu, Yisi Zhou, Beizhe Gong, Yuan Hu, Jiayou Zhang, Shihe Huang, Xuanxuan Nian, Xinghang Li, Xinguo Li, and et al. 2023. "Liposome and QS-21 Combined Adjuvant Induces theHumoral and Cellular Responses of Acellular Pertussis Vaccine in a Mice Model" Vaccines 11, no. 5: 914. https://doi.org/10.3390/vaccines11050914