
A COVID-19 DNA Vaccine Candidate Elicits Broadly Neutralizing Antibodies against Multiple SARS-CoV-2 Variants including the Currently Circulating Omicron BA.5, BF.7, BQ.1 and XBB

 and
and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. DNA Vaccine Construction
2.2. MAP-1016 Preparation
2.3. Inactivated Prototype SARS-CoV-2 Virus Vaccine
2.4. Western Blot
2.5. qRT-PCR
2.6. Animal Immunization
2.7. Enzyme-Linked Immunosorbent Assay
2.8. Neutralization Antibody Detection
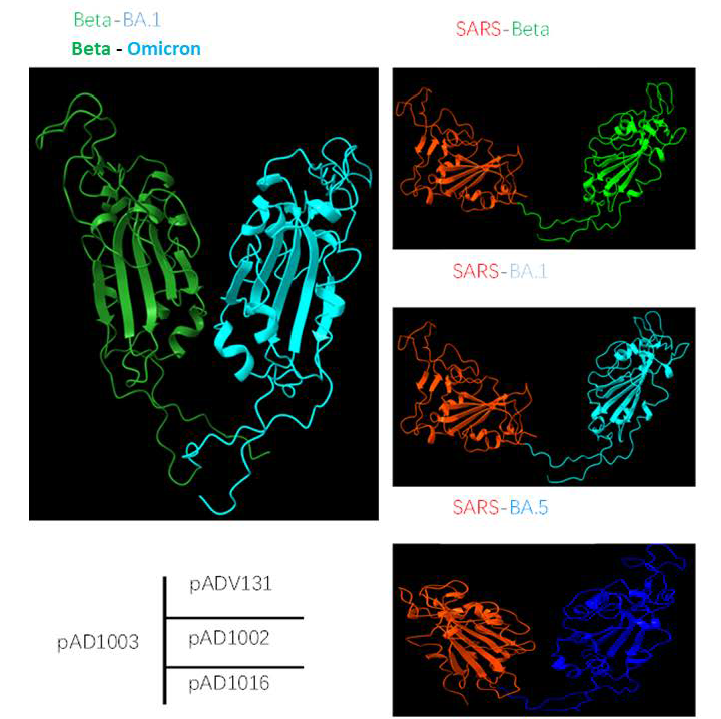
2.9. Molecular Structure AI Modeling
2.10. ELISpots
2.11. Statistics
3. Results
3.1. Preparation of DNA Construct Encoding RBD Chimera of SARS-CoV-1 and SARS-CoV-2 Omicron BA.4/5
3.2. Cellular Responses Elicited by pAD1016 in Mice
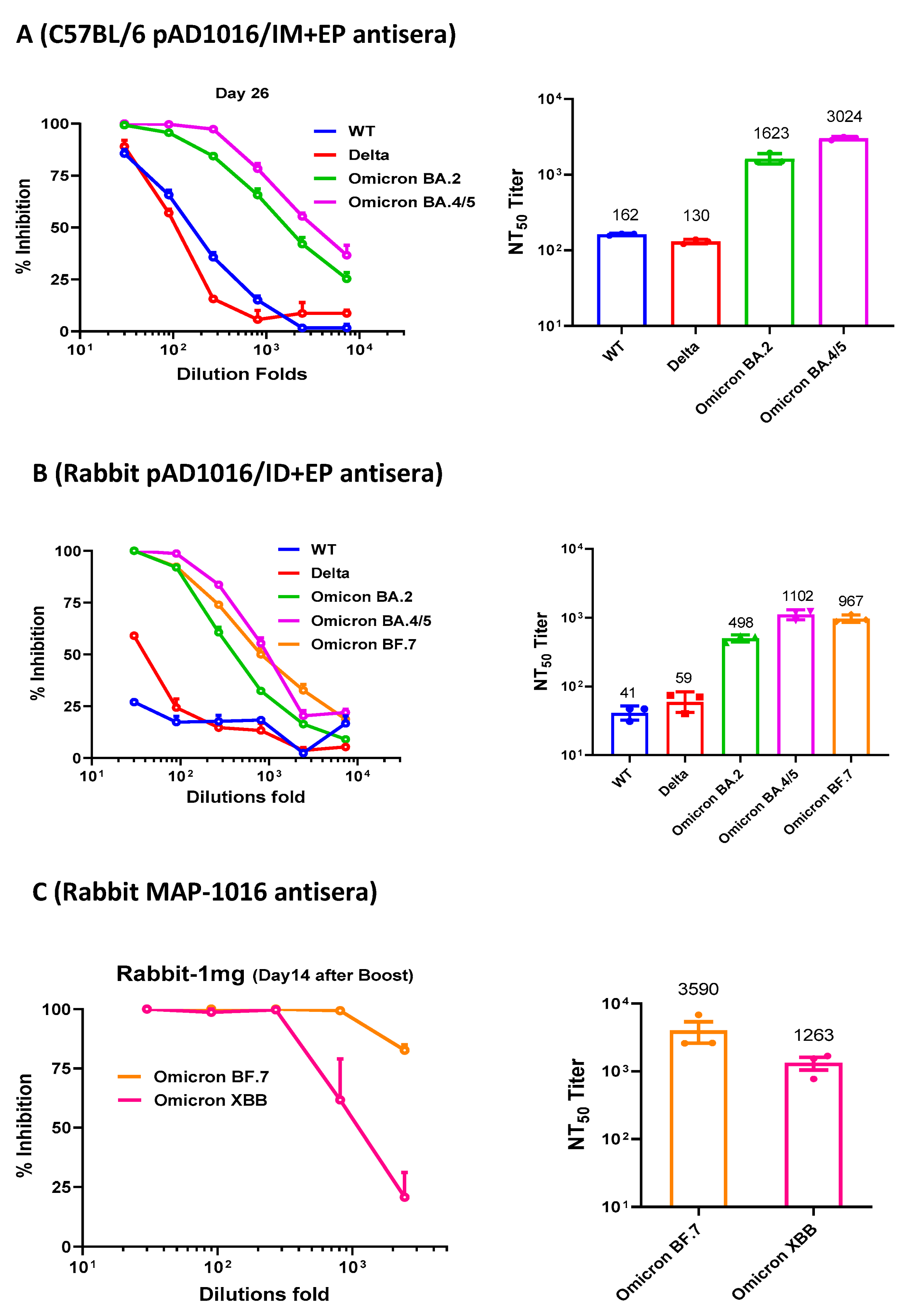
3.3. Neutralizing Abs induced by pAD1016 in Mice and Rabbits
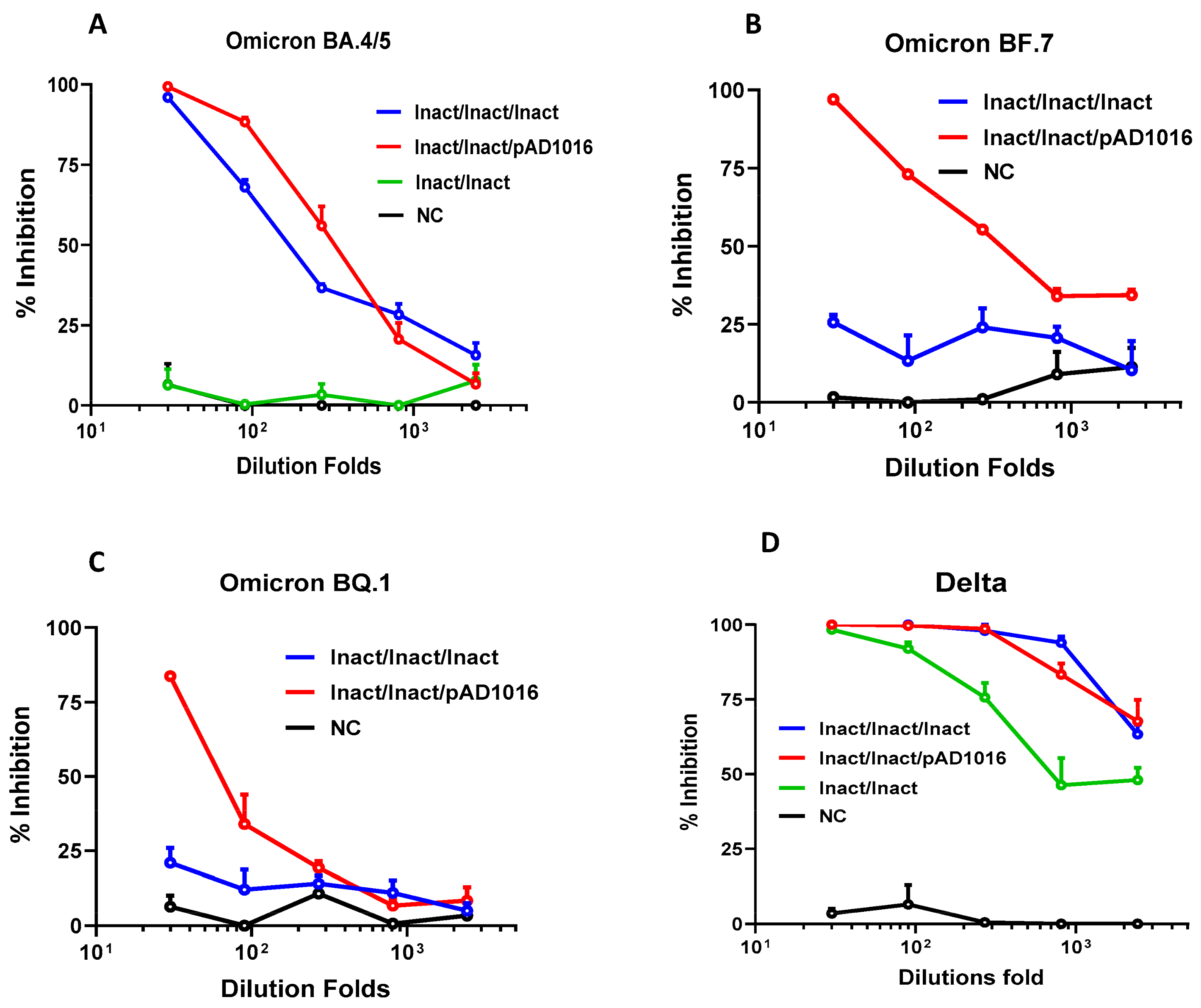
3.4. Plasmid pAD1016 as a Booster Dose to Inactivated Prototype SARS-CoV-2 Virus Pre-Vaccination
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO. Interim Statement on COVID-19 Vaccines in the Context of the Circulation of the Omicron SARS-CoV-2 Variant from the WHO Technical Advisory Group on COVID-19 Vaccine Composition (TAG-CO-VAC). 2022. Available online: https://www.who.int/news/item/11-01-2022-interim-statement-on-covid-2019-vaccines-in-the-context-of-the-circulation-of-the-omicron-sars-cov-2022-variant-from-the-who-technical-advisory-group-on-covid-2019-vaccine-composition (accessed on 16 June 2022).
- Xu, K.; Fan, C.; Han, Y.; Dai, L.; Gao, G.F. Immunogenicity, efficacy and safety of COVID-19 vaccines: An update of data published by 31 December 2021. Int. Immunol. 2022, 34, 595–607. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raddad, L.J.; Chemaitelly, H.; Butt, A.A.; National Study Group for COVID-19 Vaccination. Effectiveness of the BNT162b2 Covid-19 vaccine against the B.1.1.7 and B.1.351 variants. N Engl. J. Med. 2021, 385, 187–189. [Google Scholar] [CrossRef] [PubMed]
- Krause, P.R.; Fleming, T.R.; Longini, I.M.; Peto, R.; Briand, S.; Heymann, D.L.; Beral, V.; Snape, M.D.; Rees, H.; Ropero, A.M.; et al. SARS-CoV-2 variants and vaccines. N Engl. J. Med. 2021, 385, 179–186. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Wang, J.; Jian, F.; Xiao, T.; Song, W.; Yisimayi, A.; Huang, W.; Li, Q.; Wang, P.; An, R.; et al. Omicron escapes the majority of existing SARS-CoV-2 neutralizing antibodies. Nature 2021, 602, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Cele, S.; Jackson, L.; Khoury, D.S.; Khan, K.; Moyo-Gwete, T.; Tegally, H.; San, J.E.; Cromer, D.; Scheepers, C.; Amoako, D.; et al. Omicron extensively but incompletely escapes Pfizer BNT162b2 neutralization. Nature 2021, 602, 654–656. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Huo, J.; Zhou, D.; Zahradník, J.; Supasa, P.; Liu, C.; Duyvesteyn, H.M.E.; Ginn, H.M.; Mentzer, A.J.; Tuekprakhon, A.; et al. SARS-CoV-2 Omicron-B.1.1.529 leads to widespread escape from neutralizing antibody responses. Cell 2022, 185, 467–484.e415. [Google Scholar] [CrossRef] [PubMed]
- Karim, S.S.A.; Karim, Q.A. Omicron SARS-CoV-2 variant: A new chapter in the COVID-19 pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef]
- Garcia-Beltran, W.F.; Lam, E.C.; St Denis, K.; Nitido, A.D.; Garcia, Z.H.; Hauser, B.M.; Feldman, J.; Pavlovic, M.N.; Gregory, D.J.; Poznansky, M.C.; et al. Multiple SARS-CoV-2 variants escape neutralization by vaccine-induced humoral immunity. Cell 2021, 184, 2372–2383.e2379. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kruger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2021, 185, 447–456.e11. [Google Scholar] [CrossRef]
- Qu, P.; Evans, J.P.; Faraone, J.; Zheng, Y.M.; Carlin, C.; Anghelina, M.; Stevens, P.; Fernandez, S.; Jones, D.; Lozanski, G.; et al. Distinct neutralizing antibody escape of SARS-CoV-2 omicron subvariants BQ.1, BQ.1.1, BA.4.6, BF.7 and BA.2.75.2. Biorxiv 2022. [Google Scholar] [CrossRef]
- Li, Q.; Wu, J.; Nie, J.; Zhang, L.; Hao, H.; Liu, S.; Zhao, C.; Zhang, Q.; Liu, H.; Nie, L.; et al. The impact of mutations in SARS-CoV-2 spike on viral infectivity and antigenicity. Cell 2020, 182, 1284–1294.e1289. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.Y.; et al. Structural and functional basis of SARS-CoV-2 entry by using human ACE2. Cell 2020, 181, 894–904.e899. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Liu, M.A. DNA vaccines: An historical perspective and view to the future. Immunol. Rev. 2011, 239, 62–84. [Google Scholar] [CrossRef]
- Gurunathan, S.; Klinman, D.M.; Seder, R.A. DNA vaccines: Immunology, application, and optimization. Annu. Rev. Immunol 2000, 18, 927–974. [Google Scholar] [CrossRef] [Green Version]
- Zhao, G.; Zhang, Z.; Ding, Y.; Hou, J.; Liu, Y.; Zhang, M.; Sui, C.; Wang, L.; Xu, X.; Gao, X.; et al. A DNA vaccine encoding the full-length spike protein of Beta variant elicited broader cross-reactive immune responses against SARS-CoV-2 variants. Vaccines 2023, 11, 513. [Google Scholar] [CrossRef] [PubMed]
- Conforti, A.; Marra, E.; Palombo, F.; Roscilli, G.; Ravà, M.; Fumagalli, V.; Muzi, A.; Maffei, M.; Luberto, L.; Lione, L.; et al. COVID-eVax, an electroporated DNA vaccine candidate encoding the SARS-CoV-2 RBD, elicits protective responses in animal models. Mol. Ther. 2022, 30, 311–326. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.R.F.; Patel, A.; Ramos, S.; Elwood, D.; Zhu, X.; Yan, J.; Gary, E.N.; Walker, S.N.; Schultheis, K.; Purwar, M.; et al. Immunogenicity of a DNA vaccine candidate for COVID-19. Nat. Commun. 2020, 11, 2601. [Google Scholar] [CrossRef] [PubMed]
- Mallapaty, S. India’s DNA COVID vaccine is a world first-more are coming. Nat 2021, 597, 161–162. [Google Scholar] [CrossRef]
- Fan, F.; Zhang, X.; Zhang, Z.; Ding, Y.; Wang, L.; Xu, X.; Pan, Y.; Gong, F.Y.; Jiang, L.; Kang, L.; et al. Potent immunogenicity and broad-spectrum protection potential of microneedle array patch-based COVID-19 DNA vaccine candidates encoding dimeric RBD chimera of SARS-CoV and SARS-CoV-2 variants. Emerg. Microbes. Infect 2023. Revised. [Google Scholar]
- Moss, P. The T cell immune response against SARS-CoV-2. Nat. Immunol. 2022, 23, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Gao, P.; Liu, S.; Lu, S.; Lei, W.; Zheng, T.; Liu, X.; Xie, Y.; Zhao, Z.; Guo, S.; et al. Protective prototype-Beta and Delta-Omicron chimeric RBD-dimmer vaccines against SARS-CoV-2. Cell 2022, 185, 2265–2278. [Google Scholar] [CrossRef] [PubMed]
- An, Y.; Li, S.; Jin, X.; Han, J.B.; Xu, K.; Xu, S.; Han, Y.; Liu, C.; Zheng, T.; Liu, M.; et al. A tandem-repeat dimeric RBD protein-based COVID-19 vaccine ZF2001 protects mice and nonhuman primates. Emerg. Microbes. Infect. 2022, 11, 1058–1071. [Google Scholar] [CrossRef]
- Tan, C.W.; Chia, W.N.; Young, B.E.; Zhu, F.; Lim, B.L.; Sia, W.R.; Thein, T.L.; Chen, M.I.; Leo, Y.S.; Lye, D.C.; et al. Pan-sarbecovirus neutralizing antibodies in BNT162b2-immunized SARS-CoV-1 survivors. N Engl. J. Med. 2021, 10, 385. [Google Scholar] [CrossRef]
- Nagao, K.; Ginhoux, F.; Leitner, W.W.; Motegi, S.I.; Bennett, C.L.; Clausen, B.E.; Merad, M.; Udey, M.C. Murine epidermal Langerhans cells and langerin-expressing dermal dendritic cells are unrelated and exhibit distinct functions. Proc. Natl. Acad. Sci. USA 2009, 106, 3312–3317. [Google Scholar] [CrossRef] [Green Version]
- Seneschal, J.; Clark, R.A.; Gehad, A.; Baecher-Allan, C.M.; Kupper, T.S. Human epidermal Langerhans cells maintain immune homeostasis in skin by activating skin resident regulatory T cells. Immun. 2012, 36, 873–884. [Google Scholar] [CrossRef] [Green Version]
- Kabashima, K.; Honda, T.; Ginhoux, F.; Egawa, G. The immunological anatomy of the skin. Nat. Rev. Immunol. 2019, 19, 19–30. [Google Scholar] [CrossRef]
- Hirobea, S.; Susaia, R.; Takeuchia, H.; Eguchi, R.; Ito, S.; Quan, Y.-S.; Kamiyama, F.; Okada, N. Characteristics of immune induction by transcutaneous vaccination using dissolving microneedle patches in mice. Internat. J. Pharmac. 2021, 601, 120563. [Google Scholar]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, Y.; Fan, F.; Xu, X.; Zhao, G.; Zhang, X.; Zhao, H.; Wang, L.; Wang, B.; Gao, X.-M. A COVID-19 DNA Vaccine Candidate Elicits Broadly Neutralizing Antibodies against Multiple SARS-CoV-2 Variants including the Currently Circulating Omicron BA.5, BF.7, BQ.1 and XBB. Vaccines 2023, 11, 778. https://doi.org/10.3390/vaccines11040778
Ding Y, Fan F, Xu X, Zhao G, Zhang X, Zhao H, Wang L, Wang B, Gao X-M. A COVID-19 DNA Vaccine Candidate Elicits Broadly Neutralizing Antibodies against Multiple SARS-CoV-2 Variants including the Currently Circulating Omicron BA.5, BF.7, BQ.1 and XBB. Vaccines. 2023; 11(4):778. https://doi.org/10.3390/vaccines11040778
Chicago/Turabian StyleDing, Yuan, Feng Fan, Xin Xu, Gan Zhao, Xin Zhang, Huiyun Zhao, Limei Wang, Bin Wang, and Xiao-Ming Gao. 2023. "A COVID-19 DNA Vaccine Candidate Elicits Broadly Neutralizing Antibodies against Multiple SARS-CoV-2 Variants including the Currently Circulating Omicron BA.5, BF.7, BQ.1 and XBB" Vaccines 11, no. 4: 778. https://doi.org/10.3390/vaccines11040778