

Establishment and Application of Indirect ELISAs for Detecting Antibodies against Goose Astrovirus Genotype 1 and 2
Abstract
:1. Introduction
2. Materials and Methods
2.1. Virus, Cells and Serum Samples
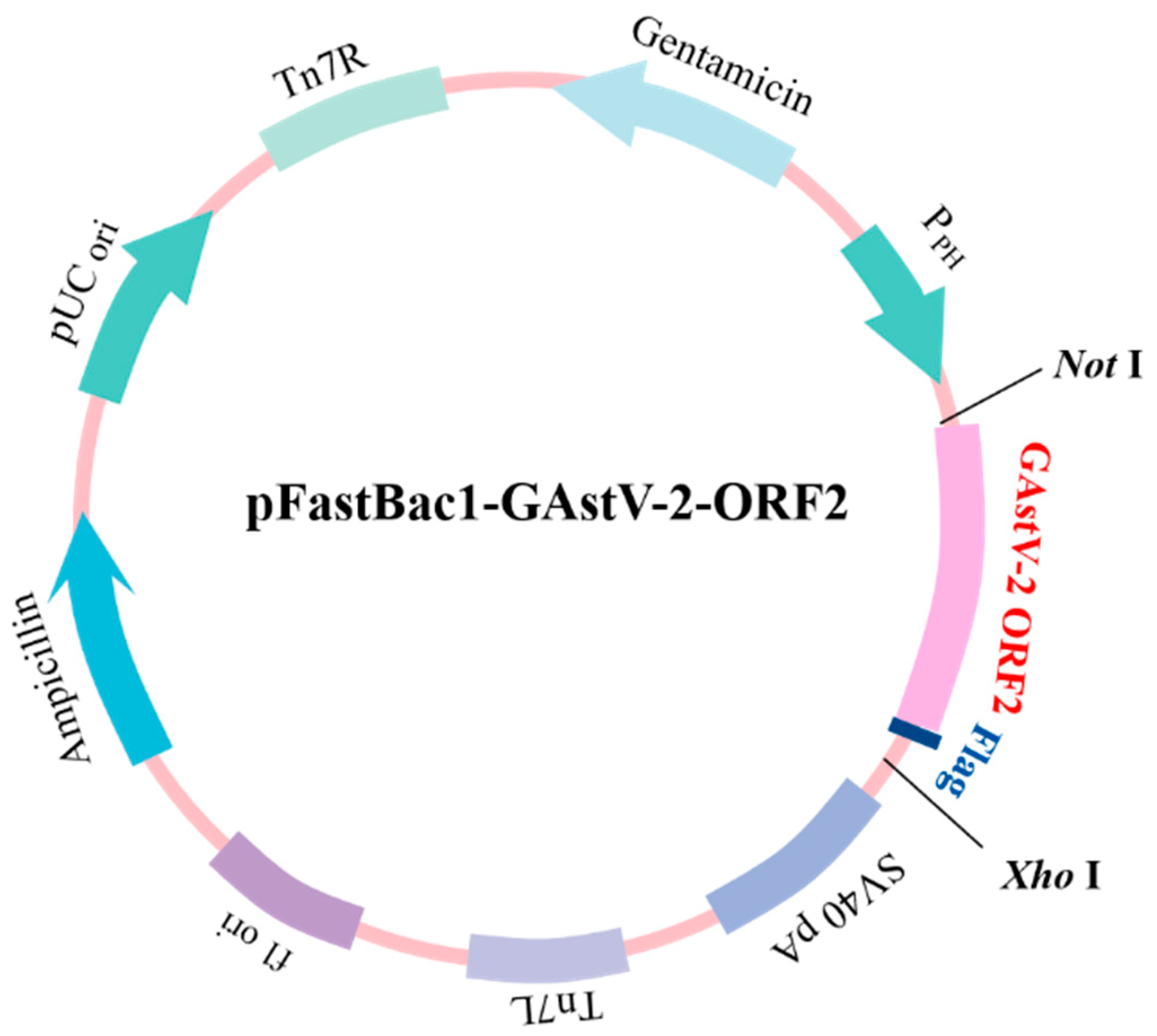
2.2. Expression and Purification of GAstV-2 ORF2 Protein
2.3. Preparation and Purification of GAstV-1
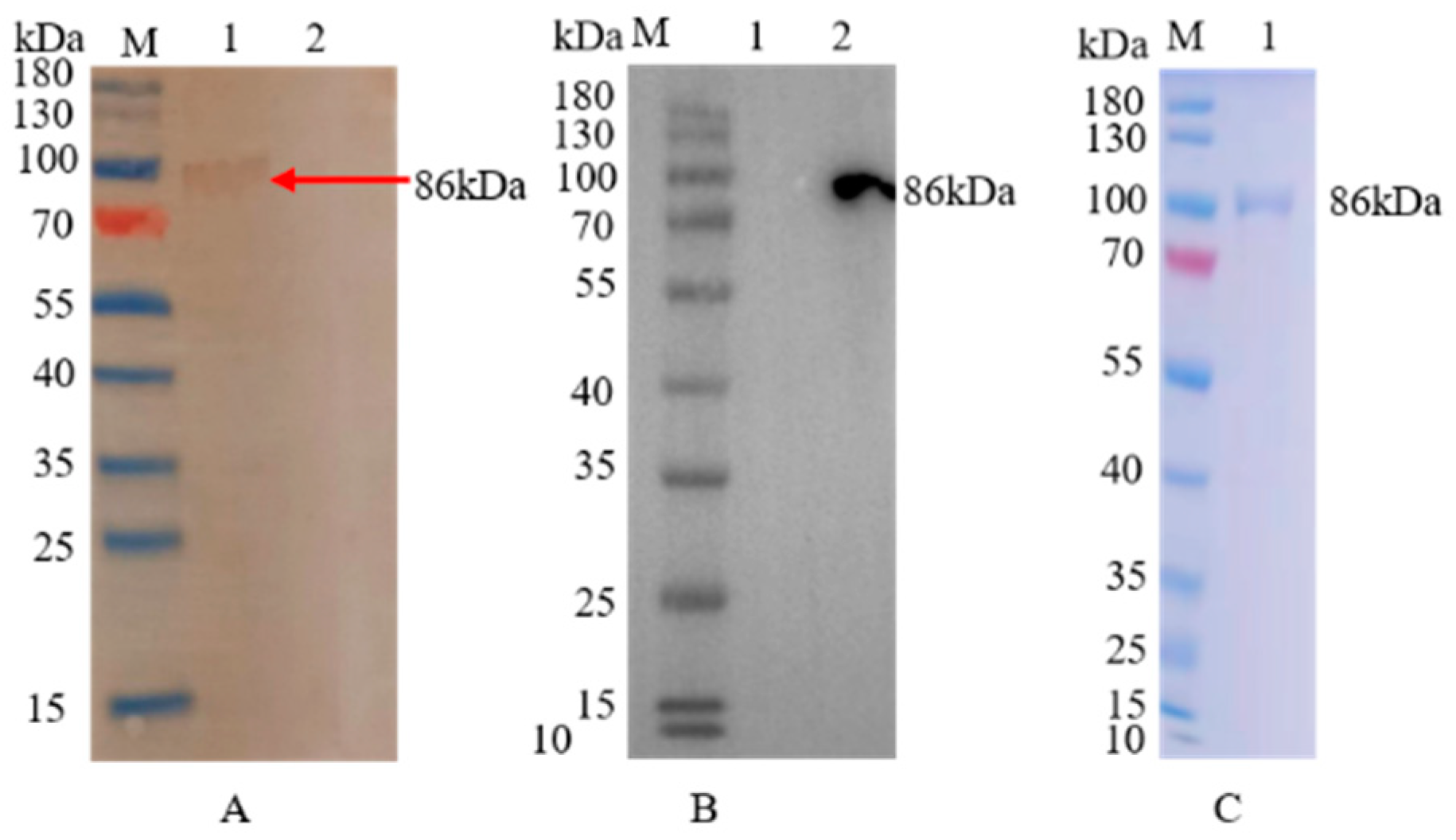
2.4. SDS-PAGE and Western Blotting Analysis
2.5. Development of GAstV-1-ELISA and GAstV-2-Cap-ELISA
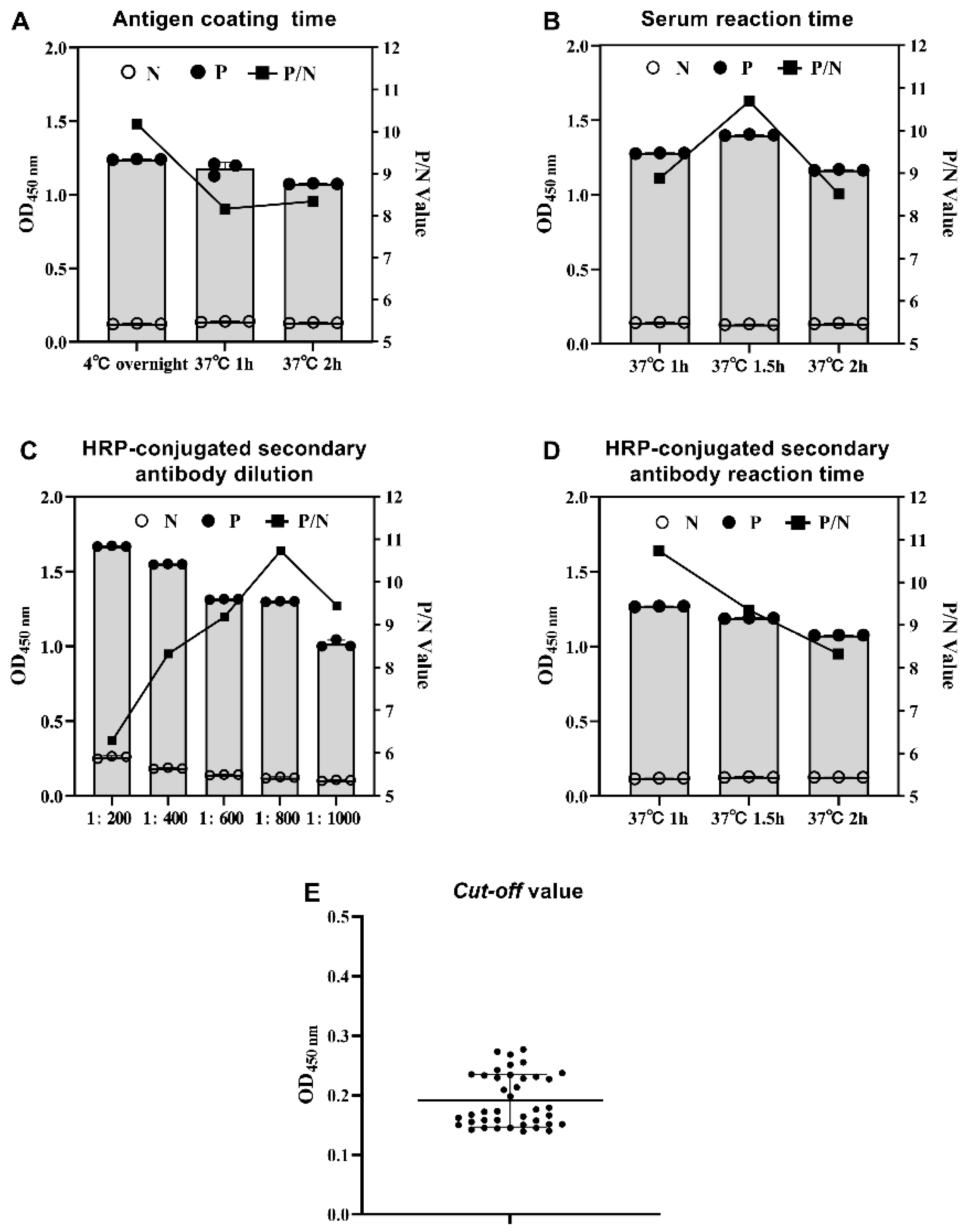
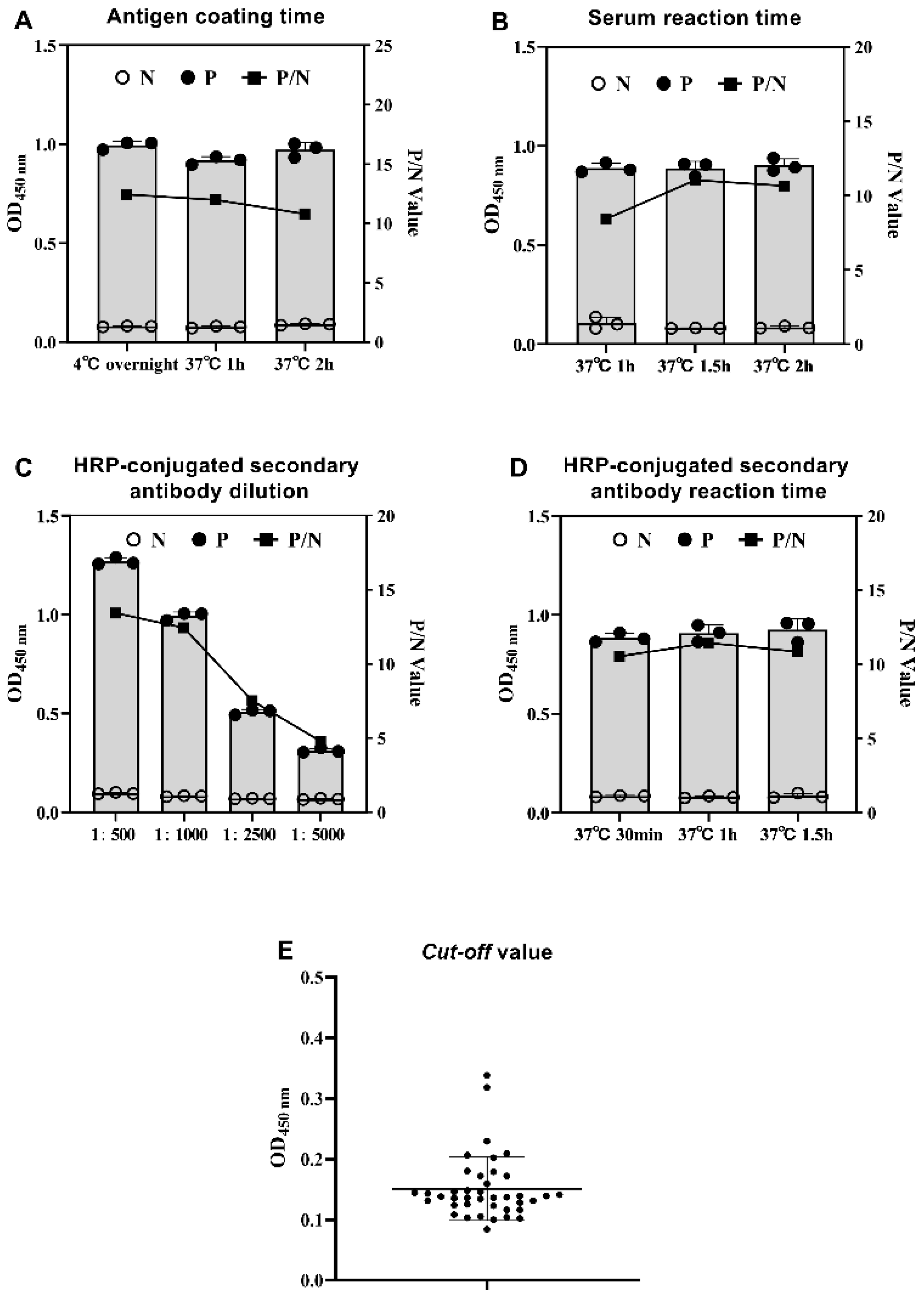
2.6. Determination of the Cut-off Value for GAstV-1-ELISA and GAstV-2-Cap-ELISA
2.7. Determination of the Specificity, Sensitivity, and Repeatability of GAstV-1-ELISA and GAstV-2-Cap-ELISA
2.8. Comparison of Indirect ELISAs with AGID Assay
2.9. Application of GAstV-1-ELISA and GAstV-2-Cap-ELISA in Clinical Samples
2.10. Statistical Analysis
3. Results
3.1. Expression and Purification of the Recombinant Protein
3.2. Establishment of Indirect ELISAs
3.2.1. Establishment of Indirect ELISA for GAstV-1 (GAstV-1-ELISA)
3.2.2. Establishment of Indirect ELISA for GAstV-2 (GAstV-2-Cap-ELISA)
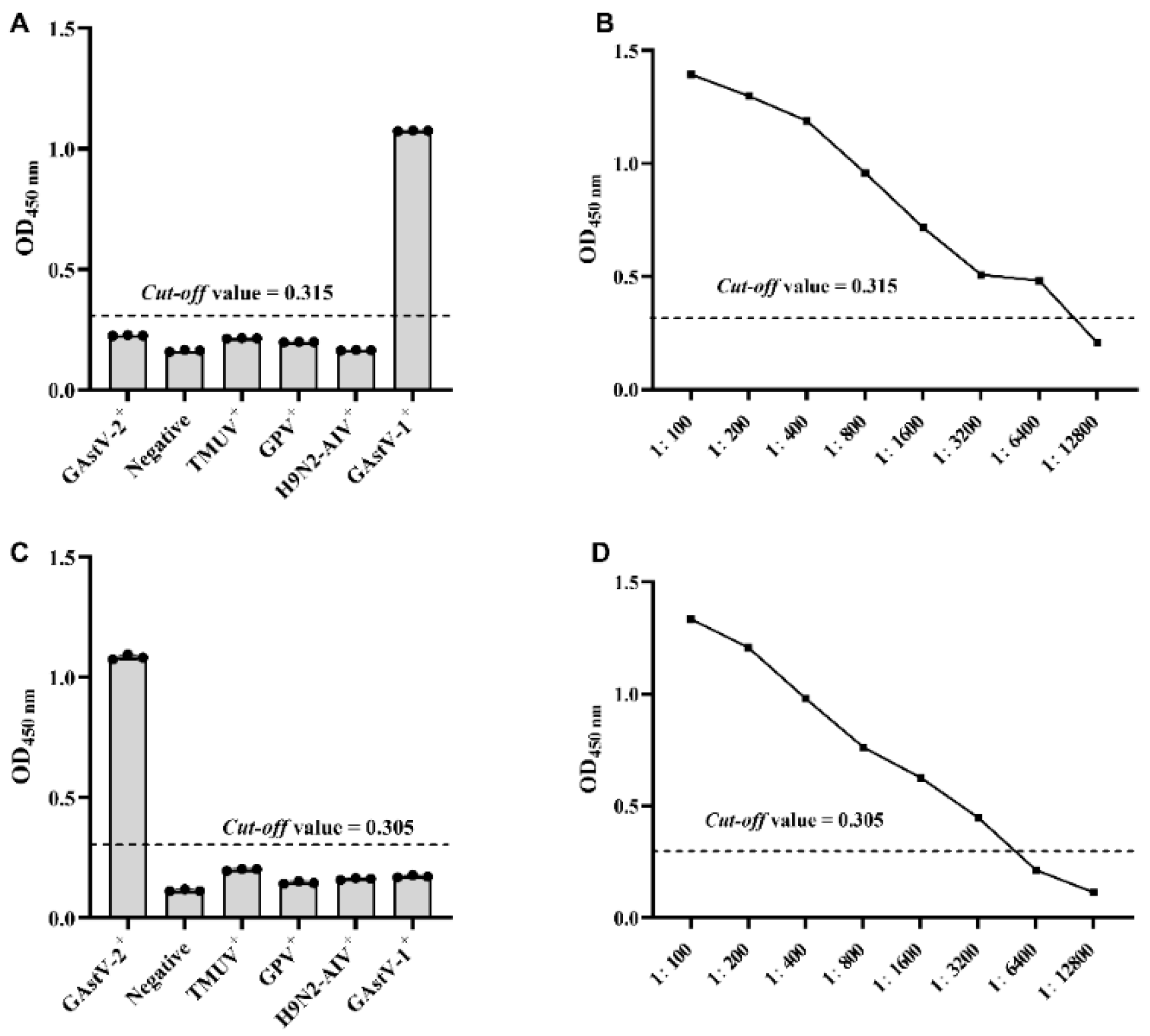
3.3. Determination of the Specificity, Sensitivity and Repeatability of GAstV-1-ELISA and GAstV-2-Cap-ELISA
3.4. Comparison of Indirect ELISAs and the AGID Assay
3.5. Application of GAstv-1-ELISA and GAstV-2-Cap-ELISA in Detecting Antibodies
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, H.; Zhang, B.; Yan, M.; Diao, Y.; Tang, Y. First report of a novel goose astrovirus outbreak in Cherry Valley ducklings in China. Transbound. Emerg. Dis. 2020, 67, 1019–1024. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.; Yang, J.; He, D.; Diao, Y.; Tang, Y. Evidence of vertical transmission of novel astrovirus virus in goose. Vet. Microbiol. 2020, 244, 108657. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Cao, Y.; Wang, J.; Fu, G.; Sun, M.; Zhang, L.; Meng, L.; Cui, G.; Huang, Y.; Hu, X.; et al. Isolation and characterization of an astrovirus causing fatal visceral gout in domestic goslings. Emerg. Microbes Infect. 2018, 7, 71. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Wang, F.; Liu, N.; Yang, L.; Zhang, D. Complete genome sequence of a novel avastrovirus in goose. Arch. Virol. 2017, 162, 2135–2139. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Xu, X.; Yu, Z.; Sui, C.; Zuo, K.; Zhi, G.; Ji, J.; Yao, L.; Kan, Y.; Bi, Y.; et al. Characterization and genomic analysis of emerging astroviruses causing fatal gout in goslings. Transbound. Emerg. Dis. 2020, 67, 865–876. [Google Scholar] [CrossRef]
- Arias, C.F.; DuBois, R.M. The Astrovirus Capsid: A Review. Viruses 2017, 9, 15. [Google Scholar] [CrossRef] [Green Version]
- Espinosa, R.; López, T.; Bogdanoff, W.A.; Espinoza, M.A.; López, S.; DuBois, R.M.; Arias, C.F. Isolation of Neutralizing Monoclonal Antibodies to Human Astrovirus and Characterization of Virus Variants That Escape Neutralization. J. Virol. 2019, 93, e01465-18. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Q.; Sun, D. Goose Astrovirus in China: A Comprehensive Review. Viruses 2022, 14, 1759. [Google Scholar] [CrossRef]
- Lin, Y.; Jiang, X.; Zahng, J.; You, W.; Hu, Q. Isolation of Novel Goose Astrovirus, FJ-NP. Fujian J. Agric. Sci. 2021, 36, 141–147. [Google Scholar]
- Shen, Q.; Zhuang, Z.; Lu, J.; Qian, L.; Li, G.; Kanton, A.G.; Yang, S.; Wang, X.; Wang, H.; Yin, J.; et al. Genome Analysis of Goose-Origin Astroviruses Causing Fatal Gout in Shanghai, China Reveals One of Them Belonging to a Novel Type Is a Recombinant Strain. Front. Vet. Sci. 2022, 9, 878441. [Google Scholar] [CrossRef]
- Tan, Z.; Zhai, H.; Sun, R.; Sun, Z.; Xie, R.; Zhang, L.; Li, Z.; Huang, J. Complete genome sequence and phylogenetic analysis of a goose astrovirus isolate in China. Braz. J. Microbiol. [Publ. Braz. Soc. Microbiol.] 2023, 54, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Liu, L.; Zhang, S.; Ye, W.; Zheng, T.; Xie, J.; Wu, S.; Wu, Z.; Feng, Q.; Dong, H.; et al. Development of a duplex real-time reverse transcription-polymerase chain reaction assay for the simultaneous detection of goose astrovirus genotypes 1 and 2. J. Virol. Methods 2022, 310, 114612. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Bai, C.; Zhang, D.; Yang, K.; Yu, Z.; Jiang, S.; Ge, K.; Li, Y. Genomic and phylogenetic characteristics of a novel goose astrovirus in Anhui Province, Central-Eastern China. Gene 2020, 756, 144898. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Zhang, D.; Yang, K.; Bai, C.; Li, Y.; Li, J.; Jiang, S.; Wang, Y. A simple and rapid diagnostic method to detect new goose astrovirus using reverse-transcription loop-mediated isothermal amplification. 3 Biotech 2020, 10, 20. [Google Scholar] [CrossRef] [PubMed]
- Wan, C.; Chen, C.; Cheng, L.; Fu, G.; Shi, S.; Liu, R.; Chen, H.; Fu, Q.; Huang, Y. Specific detection of the novel goose astrovirus using a TaqMan real-time RT-PCR technology. Microb. Pathog. 2019, 137, 103766. [Google Scholar] [CrossRef]
- Yi, Z.; Ding, R.; Cao, R.; Sun, W.; Sun, M.; Dong, Y.; Rehana, B.; Bao, E.; Lv, Y. Development of a duplex TaqMan real-time RT-PCR assay for simultaneous detection of goose astrovirus genotypes 1 and 2. J. Virol. Methods 2022, 306, 114542. [Google Scholar] [CrossRef]
- Wang, Z.; Chen, H.; Gao, S.; Song, M.; Shi, Z.; Peng, Z.; Jin, Q.; Zhao, L.; Qiao, H.; Bian, C.; et al. Core antigenic advantage domain-based ELISA to detect antibody against novel goose astrovirus in breeding geese. Appl. Microbiol. Biotechnol. 2022, 106, 2053–2062. [Google Scholar] [CrossRef]
- Ren, D.; Zhang, X.; Zhang, W.; Lian, M.; Meng, X.; Li, T.; Xie, Q.; Shao, H.; Wan, Z.; Qin, A.; et al. A peptide-based ELISA for detection of antibodies against novel goose astrovirus type 1. J. Virol. Methods 2023, 312, 114646. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, L.; Yang, J.; Zhao, D.; Han, K.; Huang, X.; Liu, Q.; Xiao, Y.; Gu, Y.; Li, Y. An IgY Effectively Prevents Goslings from Virulent GAstV Infection. Vaccines 2022, 10, 2090. [Google Scholar] [CrossRef]
- He, D.; Sun, M.; Jiang, X.; Zhang, S.; Wei, F.; Wu, B.; Diao, Y.; Tang, Y. Development of an indirect competitive ELISA method based on ORF2 detecting the antibodies of novel goose astrovirus. J. Virol. Methods 2023, 311, 114643. [Google Scholar] [CrossRef]
- Ren, D.; Li, T.; Zhang, W.; Zhang, X.; Zhang, X.; Xie, Q.; Zhang, J.; Shao, H.; Wan, Z.; Qin, A.; et al. Identification of three novel B cell epitopes in ORF2 protein of the emerging goose astrovirus and their application. Appl. Microbiol. Biotechnol. 2022, 106, 855–863. [Google Scholar] [CrossRef]
- Yuan, X.; Meng, K.; Zhang, Y.; Yu, Z.; Ai, W.; Wang, Y. Genome analysis of newly emerging goose-origin nephrotic astrovirus in China reveals it belongs to a novel genetically distinct astrovirus. Infect. Genet. Evol. J. Mol. Epidemiol. Evol. Genet. Infect. Dis. 2019, 67, 1–6. [Google Scholar] [CrossRef] [PubMed]
- An, D.; Zhang, J.; Yang, J.; Tang, Y.; Diao, Y. Novel goose-origin astrovirus infection in geese: The effect of age at infection. Poult. Sci. 2020, 99, 4323–4333. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Ren, D.; Li, T.; Zhou, H.; Liu, X.; Wang, X.; Lu, H.; Gao, W.; Wang, Y.; Zou, X.; et al. An emerging novel goose astrovirus associated with gosling gout disease, China. Emerg. Microbes Infect. 2018, 7, 152. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhu, Q.; Chen, L.; Lan, S.; Feng, W.; Li, D.; Huang, B.; Shen, S.; Shi, T. Establishment and Primary Application of a Duplex PCR Method for Detection of Goose Astrovirus and Goose Parvovirus. Chin. J. Vet. Drug 2021, 55, 1–5. [Google Scholar]
- Zhang, H.; Tian, Q.; Wang, Z.; Huang, X.; Zou, Y.; Li, X.; Lv, M.; Yang, H.; Ma, Z.; Huo, M.; et al. Establishment and application of a triple PCR method for detection of astrovirus, parvovirus and Tembusu virus in goose. Vet. Sci. China 2023, 53, 163–169. [Google Scholar]
- Liu, M.; Zhao, Y.; Hu, D.; Huang, X.; Xiong, H.; Qi, K.; Liu, H. Clinical and Histologic Characterization of Co-infection with Astrovirus and Goose Parvovirus in Goslings. Avian Dis. 2019, 63, 731–736. [Google Scholar] [CrossRef]
- Wang, Z.; Li, L.; Liu, P.; Wang, C.; Lu, Q.; Liu, L.; Yang, Y.; Luo, Q.; Shao, H. Host innate immune responses of geese infected with goose origin nephrotic astrovirus. Microb. Pathog. 2021, 152, 104753. [Google Scholar] [CrossRef]
- Lin, A.V. Indirect ELISA. Methods Mol. Biol. (Clifton N. J.) 2015, 1318, 51–59. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′-3′) |
|---|---|
| ORF2-F1 | ATGGCAGACAGGGCGGTGG |
| ORF2-R1 | TCACTTGTCATCGTCGTCCTTGTAATCCTCATGTCCGCCCTTC a |
| ORF2-F2 | GACGACCTCACTAGTCGCGGCCGCATGGCAGACAGGGCGG b |
| ORF2-F2 | GCTTGGTACCGCATGCCTCGAGTCACTTGTCATCGTCGTC c |
| M13-F | CCCAGTCACGACGTTGTAAACG |
| M13-R | AGCGGATAACAATTTCACACAGG |
| Serum Samples | GAstV-1-ELISA | GAstV-2-Cap-ELISA | ||||||
|---|---|---|---|---|---|---|---|---|
| Intra-Assay | Inter-Assay | Intra-Assay | Inter-Assay | |||||
| X ± SD | CV% | X ± SD | CV% | X ± SD | CV% | X ± SD | CV% | |
| 1 | 0.253 ± 0.006 | 2.4 | 0.249 ± 0.019 | 7.8 | 0.463 ± 0.005 | 1.1 | 0.447 ± 0.023 | 5.2 |
| 2 | 0.493 ± 0.007 | 1.3 | 0.512 ± 0.020 | 3.9 | 0.257 ± 0.010 | 4.0 | 0.217 ± 0.014 | 6.6 |
| 3 | 0.186 ± 0.011 | 5.9 | 0.162 ± 0.014 | 8.7 | 0.117 ± 0.004 | 3.2 | 0.146 ± 0.011 | 7.3 |
| 4 | 0.624 ± 0.008 | 1.3 | 0.626 ± 0.017 | 2.7 | 0.636 ± 0.008 | 1.3 | 0.666 ± 0.036 | 5.3 |
| 5 | 0.942 ± 0.027 | 2.8 | 0.952 ± 0.032 | 3.4 | 1.118 ± 0.024 | 2.2 | 1.114 ± 0.024 | 2.2 |
| 6 | 0.849 ± 0.027 | 3.2 | 0.840 ± 0.020 | 2.4 | 0.439 ± 0.027 | 6.1 | 0.447 ± 0.021 | 4.8 |
| AGID | GAstV-1-ELISA | GAstV-2-Cap-ELISA | ||||||
|---|---|---|---|---|---|---|---|---|
| Positive (n) | Negative (n) | Total (n) | Coincidence Rate (%) | Positive (n) | Negative (n) | Total (n) | Coincidence Rate (%) | |
| Positive | 21 | 0 | 21 | 60 (24/40) | 26 | 0 | 26 | 65 (26/40) |
| negative | 16 | 3 | 19 | 14 | 0 | 14 | ||
| Total | 37 | 3 | 40 | 40 | 0 | 40 | ||
| GAstV-1-ELISA | GAstV-2-Cap-ELISA | Co-Positivity (%) | ||||
|---|---|---|---|---|---|---|
| Samples (n) | Positive (n) | Positive Rate (%) | Positive (n) | Positive Rate (%) | ||
| Suqian | 345 | 8 | 2.3 (8/345) | 203 | 58.8 (203/345) | 2.3 (8/345) |
| Changzhou | 156 | 130 | 83.3 (130/156) | 146 | 93.6 (146/156) | 78.8 (123/156) |
| Zhenjiang | 94 | 60 | 63.8 (60/94) | 76 | 80.9 (76/94) | 57.4 (54/94) |
| Total | 595 | 198 | 33.3 (198/595) | 425 | 71.4 (425/595) | 31.1 (185/595) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, M.; Wei, X.; Qian, J.; Yu, Z.; Liu, X.; Luo, Y.; Zhang, H.; Gu, Y.; Li, Y. Establishment and Application of Indirect ELISAs for Detecting Antibodies against Goose Astrovirus Genotype 1 and 2. Vaccines 2023, 11, 664. https://doi.org/10.3390/vaccines11030664
Zhang M, Wei X, Qian J, Yu Z, Liu X, Luo Y, Zhang H, Gu Y, Li Y. Establishment and Application of Indirect ELISAs for Detecting Antibodies against Goose Astrovirus Genotype 1 and 2. Vaccines. 2023; 11(3):664. https://doi.org/10.3390/vaccines11030664
Chicago/Turabian StyleZhang, Mengran, Xinyu Wei, Jing Qian, Zhengyu Yu, Xin Liu, Yan Luo, Haitao Zhang, Youfang Gu, and Yin Li. 2023. "Establishment and Application of Indirect ELISAs for Detecting Antibodies against Goose Astrovirus Genotype 1 and 2" Vaccines 11, no. 3: 664. https://doi.org/10.3390/vaccines11030664