
Changing Ecotypes of Dengue Virus 2 Serotype in Nigeria and the Emergence of Cosmopolitan and Asian I Lineages, 1966–2019

 , , , and
, , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Data Sources
2.2. Evolutionary History of DENV-2 Serotypes
2.3. Computational Analysis
3. Results
3.1. Prevalence of Dengue Virus (DENV) and (DENV-2) Isolates Characterized in Nigeria
3.2. Phylogenetics of DENV-2 Isolates in Nigeria and across the World
3.3. Identification of Mutations in the DENV-2 Isolates
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brady, O.J.; Gething, P.W.; Bhatt, S.; Messina, J.P.; Brownstein, J.S.; Hoen, A.G.; Moyes, C.L.; Farlow, A.W.; Scott, T.W.; Hay, S.I. Refining the global spatial limits of dengue virus transmission by evidence-based consensus. PLoS Negl. Trop. Dis. 2012, 6, e1760. [Google Scholar] [CrossRef] [PubMed]
- Onoja, A.B.; Omatola, C.A.; Maiga, M.; Gadzama, I.S. Recurrent Episodes of Some Mosquito-Borne Viral Diseases in Nigeria: A Systematic Review and Meta-Analysis. Pathogens 2022, 11, 1162. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Quam, M.B.; Zhang, T.; Sang, S. Global burden for dengue and the evolving pattern in the past 30 years. J. Travel Med. 2021, 28, taab146. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Sakamoto, T.; Sekizuka, T.; Kato, K.; Takasaki, T.; Kuroda, M. DGV: Dengue Genographic Viewer. Front. Microbiol. 2016, 7, 875. [Google Scholar] [CrossRef]
- Omatola, C.A.; Onoja, A.B.; Moses, E.; Mahmud, M.; Mofolorunsho, C.K. Dengue in parts of the Guinea Savannah region of Nigeria and the risk of increased transmission. Int. Health 2021, 13, 248–252. [Google Scholar] [CrossRef]
- Mustafa, M.S.; Rasotgi, V.; Jain, S.; Gupta, V. Discovery of fifth serotype of dengue virus (DENV-5): A new public health dilemma in dengue control. Med. J. Armed Forces India 2015, 71, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Waman, V.P.; Kolekar, P.; Ramtirthkar, M.R.; Kale, M.M.; Kulkarni-Kale, U. Analysis of genotype diversity and evolution of Dengue virus serotype 2 using complete genomes. PeerJ 2016, 24, e2326. [Google Scholar] [CrossRef]
- Maginnis, M.S. Virus-Receptor Interactions: The Key to Cellular Invasion. J. Mol. Biol. 2018, 430, 2590–2611. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Dengue Haemorrhagic Fever: Diagnosis, Treatment, Prevention and Control, 2nd ed.; WHO: Geneva, Switzerland, 1997. [Google Scholar]
- World Health Organization (WHO). Dengue Guidelines for Diagnosis, Treatment, Prevention and Control; NBK143157; WHO: Geneva, Switzerland, 2009. [Google Scholar]
- Mena Lora, A.J.; Fernandez, J.; Morales, A.; Soto, Y.; Feris-Iglesias, J.; Brito, M.O. Disease severity and mortality caused by dengue in a Dominican pediatric population. Am. J. Trop. Med. Hyg. 2014, 90, 169–172. [Google Scholar] [CrossRef]
- Jonduo, M.; Neave, M.J.; Javati, S.; Abala, D.; Bilo, E.; Kini, A.; Kumbu, J.; Laman, M.; Robinson, L.J.; Makita, L.; et al. Genomic Sequencing of Dengue Virus Strains Associated with Papua New Guinean Outbreaks in 2016 Reveals Endemic Circulation of DENV-1 and DENV-2. Am. J. Trop. Med. Hyg. 2022, 5, 1234–1238. [Google Scholar] [CrossRef]
- Fagbami, A.H.; Onoja, A.B. Dengue Haemorrhagic Fever: An emerging disease in Nigeria, West Africa. J. Infect. Public Health 2018, 11, 757–762. [Google Scholar] [CrossRef]
- Gubler, D.J. The changing epidemiology of yellow fever and dengue, 1900 to 2003. Comp Immunol. Microbiol. Infect. Dis. 2004, 27, 319–330. [Google Scholar]
- Ogunlade, S.T.; Meehan, M.T.; Adekunle, A.I.; Rojas, D.P.; Adegboye, O.A.; McBryde, E.S. A review: Aedes-borne arboviral infections, controls and Wolbachia-based strategies. Vaccines 2021, 8, 32. [Google Scholar] [CrossRef] [PubMed]
- Onoja, A.B.; Adeniji, J.A.; Opayele, A.V. Yellow fever vaccination in Nigeria: Focus on Oyo State. Highl. Med. Res. J. 2016, 16, 37–41. [Google Scholar]
- Irikannu, K.C.; Chukwuekezie, O.C. Malaria and Man-Biting Mosquitoes in Tropical Africa; Omniscriptum; Lambert Academic Publishers: Saarbrucken, Germany, 2015; p. 168. ISBN 3659767182. [Google Scholar]
- Chen, R.; Vasilakis, N. Dengue—Quo tu et quo vadis? Viruses 2011, 3, 1562–1608. [Google Scholar] [CrossRef]
- Amarasinghe, A.; Kuritsk, J.N.; Letson, G.W.; Margolis, H.S. Dengue virus infection in Africa. Emerg. Infect. Dis. 2011, 17, 1349–1354. [Google Scholar] [CrossRef] [PubMed]
- Yenamandra, S.P.; Koo, C.; Chiang, S.; Lim HS, J.; Yeo, Z.Y.; Ng, L.C.; Hapuarachchi, H.C. Evolution, heterogeneity and global dispersal of cosmopolitan genotype of Dengue virus type 2. Sci. Rep. 2021, 11, 13496. [Google Scholar] [CrossRef] [PubMed]
- Were, F. The dengue situation in Africa. Paediatr. Int. Child Health 2012, 32 (Suppl. 1), 18–21. [Google Scholar] [CrossRef] [PubMed]
- Raut, C.G.; Rao, N.M.; Sinha, D.P.; Hanumaiah, H.; Manjunatha, M.J. Chikungunya, Dengue, and Malaria Co-Infection after Travel to Nigeria, India. Emerg. Infect. Dis. 2015, 21, 908–909. [Google Scholar] [CrossRef] [PubMed]
- Carey, D.E.; Causey, O.R.; Reddy, S.; Cooke, A.R. Dengue viruses from febrile patients in Nigeria, 1964–1968. Lancet 1971, 1, 105–106. [Google Scholar] [CrossRef] [PubMed]
- Vasilakis, N.; Tesh, R.B.; Weaver, S.C. Sylvatic Dengue Virus Type 2 Activity in Humans, Nigeria, 1966. Emerg. Infect. Dis. 2008, 14, 502–504. [Google Scholar] [CrossRef]
- Onoja, A.B.; Adeniji, J.A.; Olaleye, O.D. High rate of unrecognized dengue virus infection in parts of the rainforest region of Nigeria. Acta Tropica 2016, 160, 39–43. [Google Scholar] [CrossRef]
- Isa, I.; Ndams, I.S.; Aminu, M.; Chechet, G.; Dotzauer, A.; Simon, A.Y. Genetic diversity of Dengue virus serotypes circulating among Aedes mosquitoes in selected regions of northeastern Nigeria. One Health 2021, 10, 100348. [Google Scholar] [CrossRef]
- Tamura, K. Estimation of the number of nucleotide substitutions when there are strong transition-transversion and G + C-content biases. Mol. Biol. Evol. 1992, 9, 678–687. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Fagbami, A.H.; Monath, T.P.; Fabiyi, A. Dengue virus infections in Nigeria: A survey for antibodies in monkeys and humans. Trans. R. Soc. Trop. Med. Hyg. 1997, 71, 60–65. [Google Scholar] [CrossRef] [PubMed]
- Robin, Y.; Cornet, M.; Heme, G.; Le Gonidec, G. Isolement du virus de la dengue au Senegal. Ann. Virol. 1980, 131, 149–154. [Google Scholar] [CrossRef]
- Diallo, M.; Ba, Y.; Sall, A.A.; Diop, O.M.; Ndione, J.A.; Mondo, M.; Girault, L.; Mathiot, C. Amplification of the sylvatic cycle of dengue virus type 2, Senegal, 1999–2000: Entomologic findings and epidemiologic considerations. Emerg. Infect. Dis. 2003, 9, 362–367. [Google Scholar] [CrossRef] [PubMed]
- Idrissa, D.; Niang, S.S.; Mignane, N.M.; Aliou, B.M.; Cheikh, T.; Moufid, M.; Diamilatou, B. Detection of human case of dengue virus 2 belonging to sylvatic genotype during routine surveillance of fever in Senegal, Kolda 2021. Front. Virol. 2022, 2. [Google Scholar] [CrossRef]
- Rudnick, A. Dengue virus ecology in Malaysia. Inst. Med. Res. Malays. Bull. 1986, 23, 51–152. [Google Scholar]
- Saluzzo, J.F.; Cornet, M.; Castagnet, P.; Rey, C.; Digoutte, J.P. Isolation of dengue 2 and dengue 4 viruses from patients in Senegal. Trans. R. Soc. Trop. Med. Hyg. 1986, 80, 5. [Google Scholar] [CrossRef]
- Otu, A.; Ebenso, B.; Etokidem, A.; Chukwuekezie, O. Dengue fever—An update review and implications for Nigeria, and similar countries. Afr. Health Sci. 2019, 19, 2000–2007. [Google Scholar] [CrossRef]
- Diallo, M.; Fernandez, Z.; Coffey, L.L.; Ba, Y.; Weaver, S.; Ortiz, D.; Mathiot, C.; Tesh, R.B.; Moncayo, A.C.; Sall, A.A. Potential role of sylvatic and domestic African mosquito species in dengue emergence. Am. J. Trop. Med. Hyg. 2005, 73, 445–449. [Google Scholar] [CrossRef] [PubMed]
- Diallo, M.; Ba, Y.; Faye, O.; Soumare, M.L.; Dia, I.; Sall, A.A. Vector competence of Aedes aegypti populations from Senegal for sylvatic and epidemic dengue 2 virus isolated in West Africa. Trans. R. Soc. Trop. Med. Hyg. 2008, 102, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Langat, S.K.; Eyase, F.L.; Berry, I.M.; Nyunja, A.; Bulimo, W.; Owaka, S.; Ofula, V.; Limbaso, S.; Lutomiah, J.; Jarman, R.; et al. Origin and evolution of dengue virus type 2 causing outbreaks in Kenya: Evidence of circulation of two cosmopolitan genotype lineages. Virus Evol. 2020, 6, p.veaa026. [Google Scholar] [CrossRef]
- Monath, T.P. Dengue: The risk to developed and developing countries. Proc. Nat. Acad. Sci. USA. 1994, 91, 2395–2400. [Google Scholar] [CrossRef]
- Eldin, C.; Gautret, P.; Nougairede, A.; Sentis, M.; Ninove, L.; Saidani, N.; Million, M.; Brouqui, P.; Charrel, R.; Parola, P. Identification of dengue type 2 virus in febrile travellers returning from Burkina Faso to France, related to an ongoing outbreak, October to November 2016. Euro. Surveill. 2016, 21, 30425. [Google Scholar] [CrossRef]
- Gathii, K.; Nyataya, J.N.; Mutai, B.K.; Awinda, G.; Waitumbi, J.N. Complete Coding Sequences of Dengue Virus Type 2 Strains from Febrile Patients Seen in Malindi District Hospital, Kenya, during the 2017 Dengue Fever Outbreak. Genome Announc. 2018, 6, e00076-18. [Google Scholar] [CrossRef]
- Kamau, E.; Agoti, C.N.; Ngoi, J.M.; de Laurent, Z.R.; Gitonga, J.; Cotten, M.; Phan, M.V.T.; Nokes, D.J.; Delwart, E.; Sanders, E. Complete Genome Sequences of Dengue Virus Type 2 Strains from Kilifi, Kenya. Microbiol. Res. Announc. 2019, 8, e01566-18. [Google Scholar] [CrossRef]
- Masika, M.M.; Korhonen, E.M.; Smura, T.; Uusitalo, R.; Vapalahti, K.; Mwaengo, D.; Jääskeläinen, A.J.; Anzala, O.; Vapalahti, O.; Huhtamo, E. Detection of dengue virus type 2 of Indian origin in acute febrile patients in rural Kenya. PLoS Negl. Trop. Dis. 2020, 14, e0008099. [Google Scholar] [CrossRef] [PubMed]
- Amoako, N.; Duodu, S.; Dennis, F.E.; Bonney, J.H.; Asante, K.P.; Ameh, J.; Mosi, L.; Hayashi, T.; Agbosu, E.E.; Pratt, D.; et al. Detection of Dengue Virus among Children with Suspected Malaria, Accra, Ghana. Emerg. Infect. Dis. 2018, 24, 1544–1547. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.M.; Vorndam, A.V.; Adeniji, J.A.; Diop, O.; Olaleye, D.O. Dengue Virus Infections in Patients Suspected of Malaria/Typhoid in Nigeria. J. Am. Sci. 2009, 5, 129–134. [Google Scholar]
- Agwu, E.J.; Isaak, C.; Igbinosa, B.I. Detection of yellow fever and dengue viruses in mosquitoes between 2014 and 2015 in Bayelsa and Benue states of Nigeria. Acta Entomol. Serbica 2019, 24, 59–78. [Google Scholar]
- Vasilakis, N.; Fokam, E.B.; Hanson, C.T.; Weinberg, E.; Sall, A.A.; Whitehead, S.S.; Hanley, K.A.; Weaver, S.C. Genetic and phenotypic characterization of sylvatic dengue virus type 2 strains. Virology 2008, 377, 296–307. [Google Scholar] [CrossRef] [PubMed]
- Phan, T.T.N.; Hvasta, M.G.; Kudlacek, S.T.; Thiono, D.J.; Tripathy, A.; Nicely, N.I.; de Silva, A.M.; Kuhlman, B. A conserved set of mutations for stabilizing soluble envelope protein dimers from dengue and Zika viruses to advance the development of subunit vaccines. J. Biol Chem. 2022, 298, 102079. [Google Scholar] [CrossRef] [PubMed]





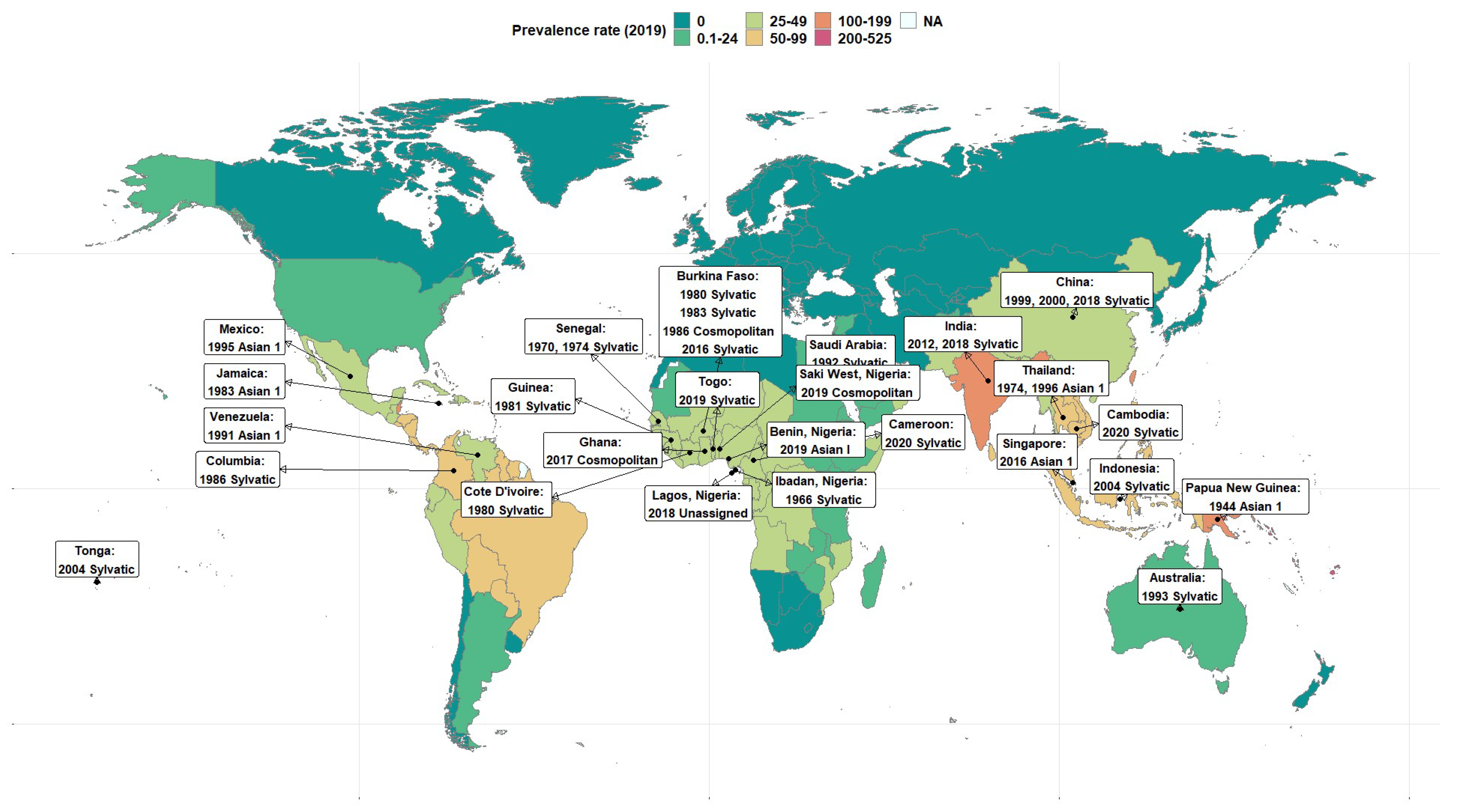{kind=link}
{kind=link}
{kind=link}
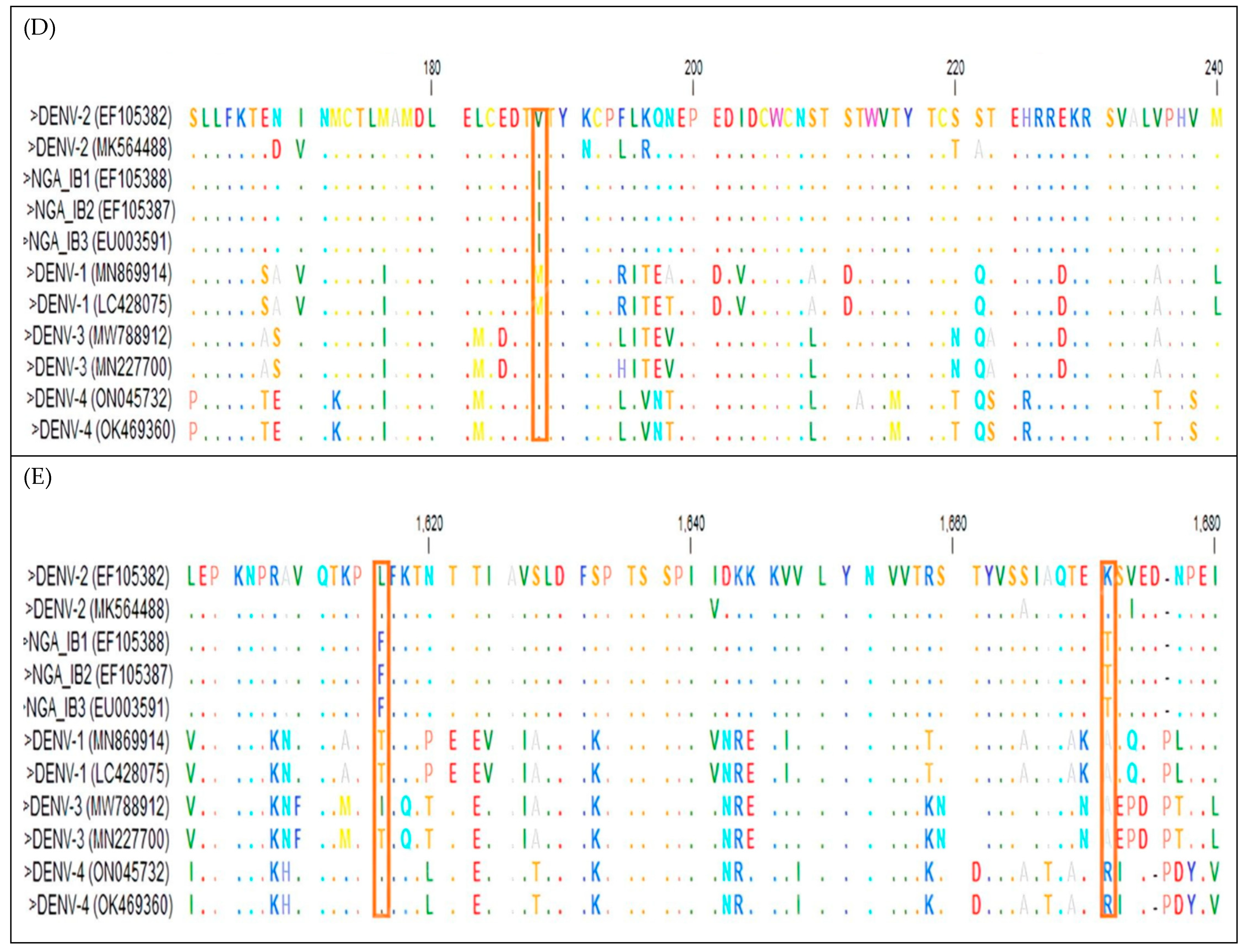{kind=link}
| SN | Accession No. | Genotype | Strain | Year | Location | Vegetation | Source |
|---|---|---|---|---|---|---|---|
| 1 | EF105387.1 | Genotype VI | Sylvatic | 1966 | Ibadan, Nigeria | Derived savannah | Human |
| 2 | DQ341197.1 | Genotype VI | Sylvatic | 1966 | Ibadan, Nigeria | Derived savannah | Human |
| 3 | EF105388.1 | Genotype VI | Sylvatic | 1966 | Ibadan, Nigeria | Derived savannah | Human |
| 4 | EU003591.1 | Genotype VI | Sylvatic | 1966 | Ibadan, Nigeria | Derived savannah | Human |
| 5 | MK045282.1 * | Unassigned | Unassigned | 2018 | Lagos, Nigeria | Mangrove, rainforest, and freshwater swamp | Human |
| 6 | MK045280.1 * | Unassigned | Unassigned | 2018 | Lagos, Nigeria | Mangrove, rainforest, and freshwater swamp | Human |
| 7 | MT012480.1 | Genotype II | Cosmopolitan | 2019 | Saki West, Nigeria | Derived savannah | Human |
| 8 | MW341537.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 9 | MW341538.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 10 | MW341539.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 11 | MW341540.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 12 | MW341541.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 13 | MW341542.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 14 | MW341543.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 15 | MW341544.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 16 | MW341545.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 17 | MW341546.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 18 | MW341547.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
| 19 | MW332260.1 | Genotype V | Asian I | 2019 | Benin, Nigeria | Rainforest | Human |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Onoja, B.A.; Maiga, M.; Adesola, R.O.; Adamu, A.M.; Adegboye, O.A. Changing Ecotypes of Dengue Virus 2 Serotype in Nigeria and the Emergence of Cosmopolitan and Asian I Lineages, 1966–2019. Vaccines 2023, 11, 547. https://doi.org/10.3390/vaccines11030547
Onoja BA, Maiga M, Adesola RO, Adamu AM, Adegboye OA. Changing Ecotypes of Dengue Virus 2 Serotype in Nigeria and the Emergence of Cosmopolitan and Asian I Lineages, 1966–2019. Vaccines. 2023; 11(3):547. https://doi.org/10.3390/vaccines11030547
Chicago/Turabian StyleOnoja, Bernard A., Mamoudou Maiga, Ridwan O. Adesola, Andrew M. Adamu, and Oyelola A. Adegboye. 2023. "Changing Ecotypes of Dengue Virus 2 Serotype in Nigeria and the Emergence of Cosmopolitan and Asian I Lineages, 1966–2019" Vaccines 11, no. 3: 547. https://doi.org/10.3390/vaccines11030547