

Protection of SPF Chickens by H9N2 Y439 and G1 Lineage Vaccine against Homologous and Heterologous Viruses
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Viruses and Vaccines
2.2. Statistical Analysis
2.3. Study 1: Protection of vac564 Vaccine against Challenge by Homologous and Heterologous Viruses in SPF Chickens
2.3.1. Vaccine Candidate Development by Multiple Passage
2.3.2. Vaccination and Challenge of SPF Chickens
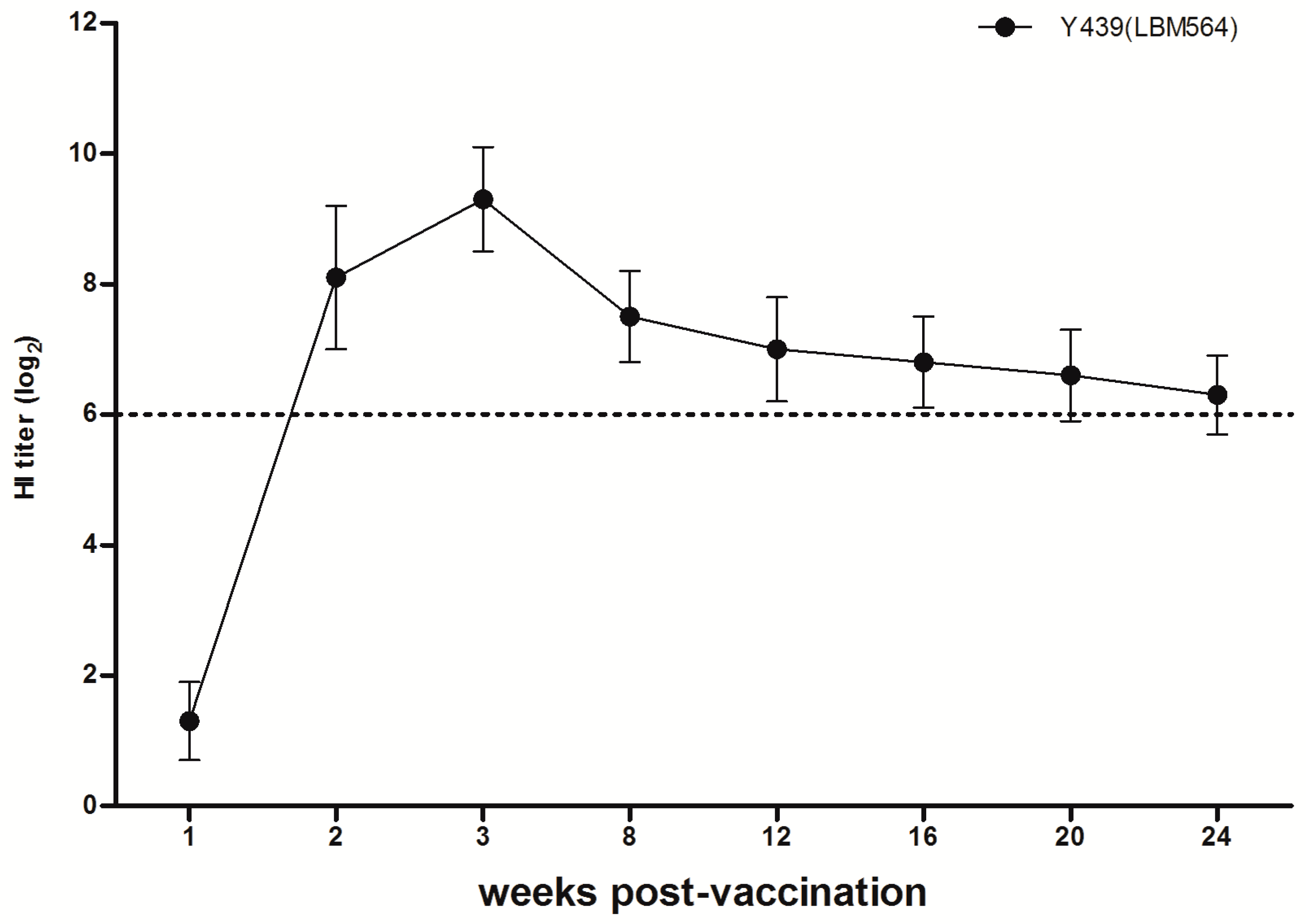
2.3.3. Serology and Antibody Persistence
2.3.4. Virus Shedding and Replication
2.4. Study 2: Protection of Commercial G1 Lineage Vaccine against Challenge by Heterologous Viruses in SPF Chickens
2.4.1. Vaccination and Challenge of SPF Chickens
2.4.2. Serology and Antibody Assays
2.4.3. Virus Shedding and Replication
3. Results
3.1. Study 1: Protection of vac564 Vaccine against Challenge by Homologous and Heterologous Viruses in SPF Chickens
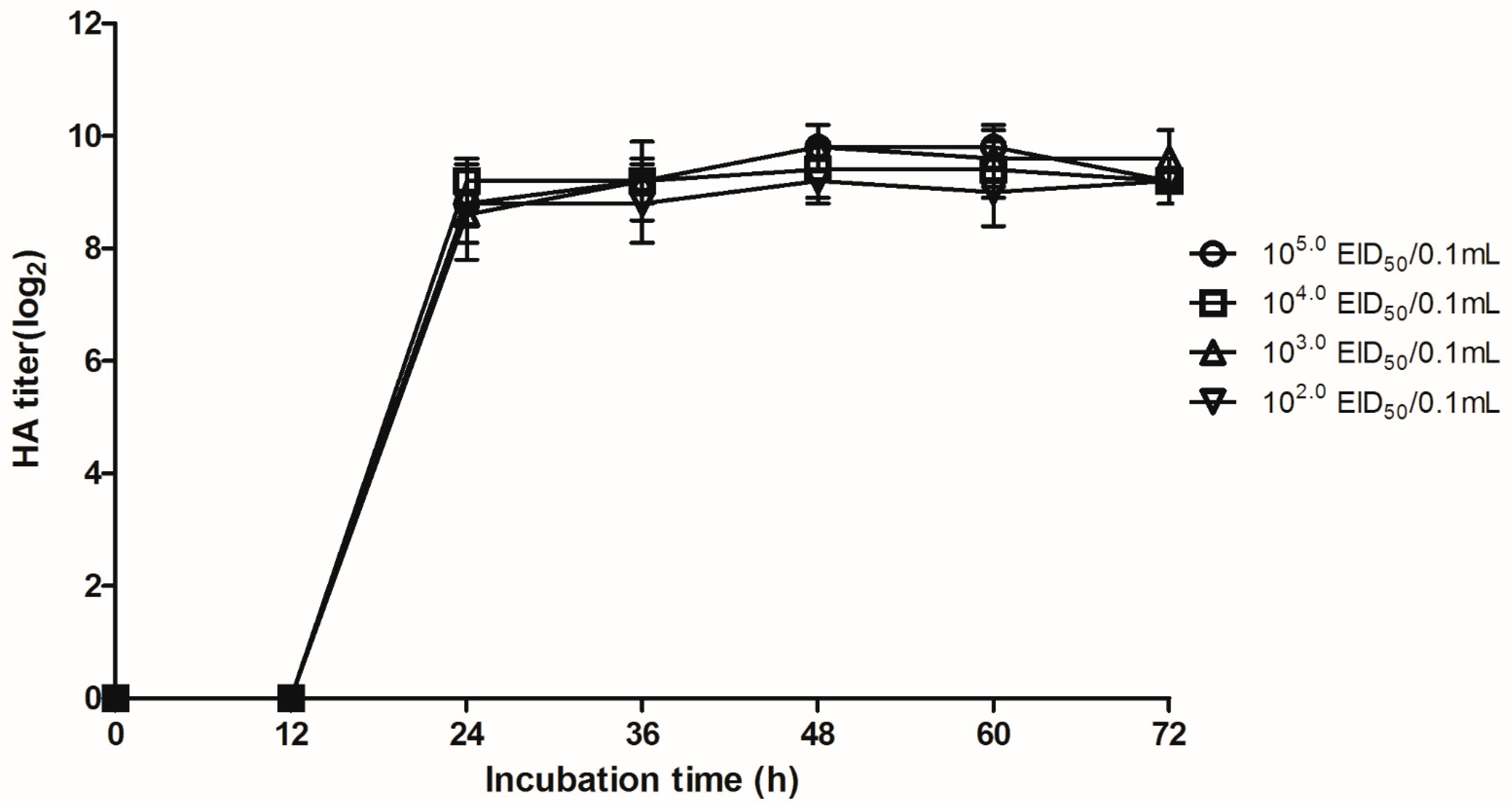
3.1.1. Development and Characteristics of Vaccine Candidate
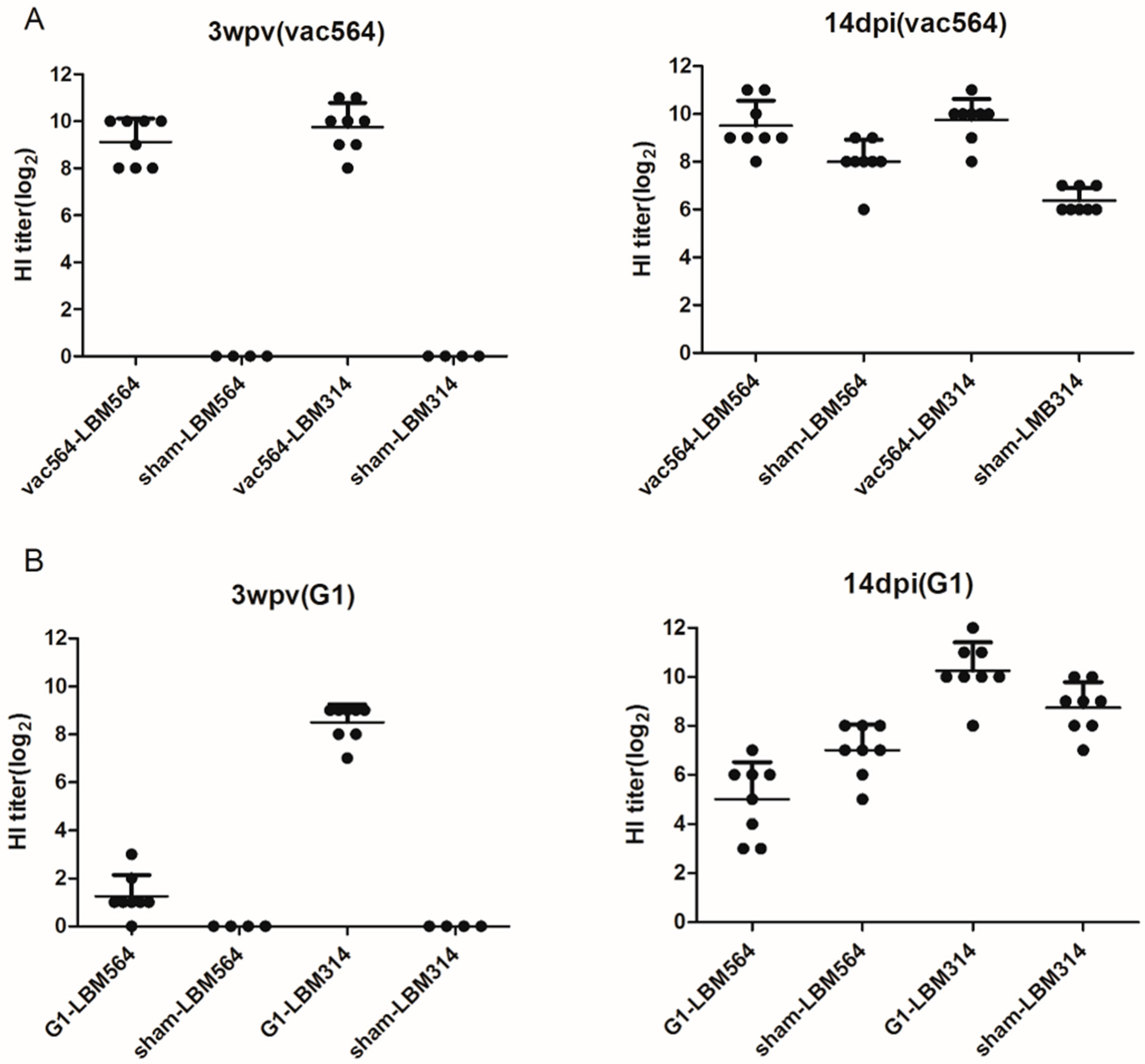
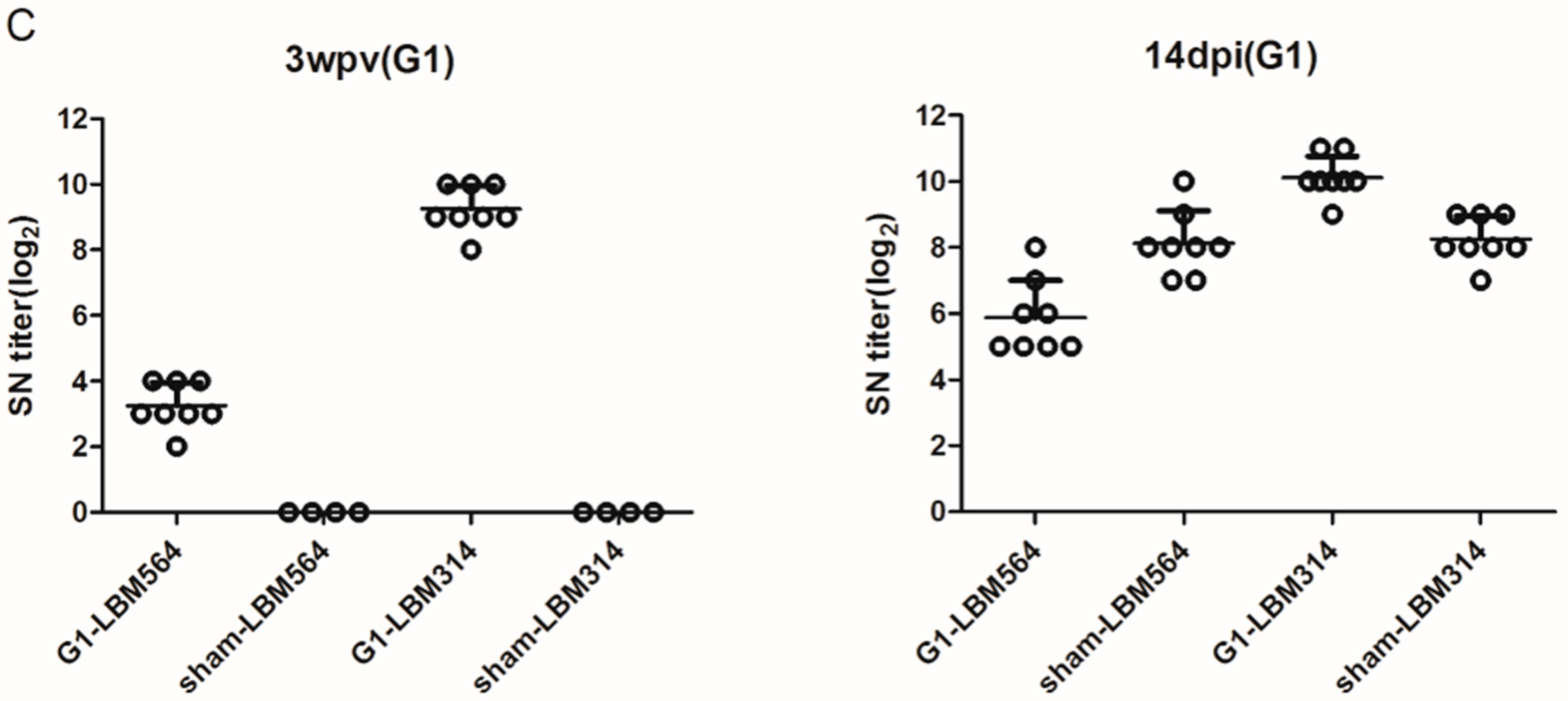
3.1.2. Serological Responses and Antibody Persistence
3.1.3. Virus Shedding and Replication Post-Challenge
3.2. Study 2: Protection of Commercial G1 Lineage Vaccine against Challenge by Heterologous Viruses in SPF Chickens
3.2.1. Serological Responses
3.2.2. Virus Shedding and Replication Post-Challenge
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Chin, P.S.; Dyrting, K.C.; Ellis, T.M.; Webster, R.G.; Peiris, M. H9N2 influenza viruses possessing H5N1-like internal genomes continue to circulate in poultry in Southeastern China. J. Virol. 2000, 74, 9372–9380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pusch, E.A.; Suarez, D.L. The multifaceted zoonotic risk of H9N2 avian influenza. Vet. Sci. 2018, 5, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ducatez, M.F.; Webster, R.G.; Webby, R.J. Animal influenza epidemiology. Vaccine 2008, 26, 67–69. [Google Scholar] [CrossRef] [Green Version]
- Lee, D.H.; Song, C.S. H9N2 avian influenza virus in Korea: Evolution and vaccination. Clin. Exp. Vaccine Res. 2013, 2, 26–33. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.G.; Lee, Y.J.; Kim, Y.J.; Lee, E.K.; Jeong, O.M.; Sung, H.W.; Kim, J.H.; Kwon, J.H. An inactivated vaccine to control the current H9N2 low pathogenic avian influenza in Korea. J. Vet. Sci. 2008, 9, 67–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gharaibeh, S.; Amareen, S. Vaccine efficacy against a new avian influenza (H9N2) field isolate from the Middle East (serology and challenge studies). Avian Dis. 2015, 59, 508–511. [Google Scholar] [CrossRef]
- Jin, F.; Dong, X.; Wan, Z.; Ren, D.; Liu, M.; Geng, T.; Zhang, J.; Gao, W.; Shao, H.; Qin, A.; et al. A single mutation N166D in hemagglutinin affects antigenicity and pathogenesis of H9N2 avian influenza virus. Viruses 2019, 11, 709. [Google Scholar] [CrossRef] [Green Version]
- Guan, Y.; Shortridge, K.F.; Krauss, S.; Webster, R.G. Molecular characterization of H9N2 influenza viruses: Were they the donors of the “internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA 1999, 96, 9363–9367. [Google Scholar] [CrossRef] [Green Version]
- Bonfante, F.; Mazzetto, E.; Zanardello, C.; Fortin, A.; Gobbo, F.; Maniero, S.; Bigolaro, M.; Davidson, I.; Haddas, R.; Cattoli, G.; et al. A G1-lineage H9N2 virus with oviduct tropism causes chronic pathological changes in the infundibulum and a long-lasting drop in egg production. Vet. Res. 2018, 49, 83. [Google Scholar] [CrossRef] [Green Version]
- Peacock, T.P.; James, J.; Sealy, J.E.; Iqbal, M. A global perspective on H9N2 avian influenza virus. Viruses 2019, 11, 620. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Pu, J.; Jiang, Z.; Guan, T.; Xia, Y.; Xu, Q.; Liu, L.; Ma, B.; Tian, F.; Brown, E.G.; et al. Genotypic evolution and antigenic drift of H9N2 influenza viruses in China from 1994 to 2008. Vet. Microbiol. 2010, 146, 215–225. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, S.; Bing, G.; Carter, R.A.; Wang, Z.; Wang, J.; Wang, C.; Wang, L.; Wu, G.; Webster, R.G.; et al. Genetic evolution of influenza H9N2 viruses isolated from various hosts in China from 1994 to 2013. Emerg. Microbes Infect. 2017, 6, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.K.; Ozaki, H.; Webby, R.J.; Webster, R.G.; Peiris, J.S.; Poon, L.; Butt, C.; Leung, Y.H.C.; Guan, Y. Continuing evolution of H9N2 influenza viruses in Southeastern China. J. Virol. 2004, 78, 8609–8614. [Google Scholar] [CrossRef] [Green Version]
- Kandeil, A.; El-Shesheny, R.; Maatouq, A.; Moatasim, Y.; Cai, Z.; Mckenzie, P.; Webby, R.; Kayali, G.; Ali, M.A. Novel reassortant H9N2 viruses in pigeons and evidence for antigenic diversity of H9N2 viruses isolated from quails in Egypt. J. Gen. Virol. 2017, 98, 548–562. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.H.; Swayne, D.E.; Sharma, P.; Rehmani, S.F.; Wajid, A.; Suarez, D.L.; Afonso, C.L. H9N2 low pathogenic avian influenza in Pakistan (2012–2015). Vet. Rec. 2016, 3, e000171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nili, H.; Asasi, K. Avian influenza (H9N2) outbreak in Iran. Avian Dis. 2003, 47, 828–831. [Google Scholar] [CrossRef]
- Zou, S.; Zhang, Y.; Li, X.; Bo, H.; Wei, H.; Dong, L.; Yang, L.; Dong, J.; Liu, J.; Shu, Y.; et al. Molecular characterization and receptor binding specificity of H9N2 avian influenza viruses based on poultry-related environmental surveillance in China between 2013 and 2016. Virology 2019, 529, 135–143. [Google Scholar] [CrossRef]
- Al-Garib, S.; Agha, A.; Al-Mesilaty, L. Low pathogenic avian influenza H9N2: World-wide distribution. Worlds Poult. Sci. 2016, 72, 125–136. [Google Scholar] [CrossRef]
- Mo, I.P.; Song, C.S.; Kim, K.S.; Rhee, J.C. An occurrence of non-highly pathogenic avian influenza in Korea. Avian Dis. 2003, 47, 379–383. [Google Scholar]
- Lee, C.W.; Song, C.S.; Lee, Y.J.; Mo, I.P.; Garcia, M.; Suarez, D.L.; Kim, S.J. Sequence analysis of the hemagglutinin gene of H9N2 Korean avian influenza viruses and assessment of the pathogenic potential of isolate MS96. Avian Dis. 2000, 44, 527–535. [Google Scholar] [CrossRef]
- Lee, Y.J.; Shin, J.Y.; Song, M.S.; Lee, Y.M.; Choi, J.G.; Lee, E.K.; Jeong, O.M.; Sung, H.W.; Kim, J.H.; Kwon, Y.K.; et al. Continuing evolution of H9 influenza viruses in Korean poultry. Virology 2007, 359, 313–323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, D.H.; Fusaro, A.; Song, C.S.; Suarez, D.L.; Swayne, D.E. Poultry vaccination directed evolution of H9N2 low pathogenic avian influenza viruses in Korea. Virology 2016, 488, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, G.B.; Kye, S.J.; Sagong, M.; Lee, E.K.; Lee, K.N.; Lee, Y.N.; Choi, K.S.; Lee, M.H.; Lee, Y.J. Genetic characterization of H9N2 avian influenza virus previously unrecognized in Korea. J. Vet. Sci. 2021, 22, e21. [Google Scholar] [CrossRef] [PubMed]
- Lai, V.D.; Kim, J.W.; Choi, Y.Y.; Kim, J.J.; So, H.H.; Mo, J. First report of field cases of Y280-like LPAI H9N2 strains in South Korean poultry farms: Pathological findings and genetic characterization. Avian Pathol. 2021, 50, 327–338. [Google Scholar] [CrossRef]
- Kye, S.J.; Park, M.J.; Kim, N.Y.; Lee, Y.N.; Heo, G.B.; Baek, Y.K.; Shin, J.I.; Lee, M.H.; Lee, Y.J. Pathogenicity of H9N2 low pathogenic avian influenza viruses of different lineages isolated from live bird markets tested in three animal models: SPF chickens, Korean native chickens, and ducks. Poult. Sci. 2021, 100, 101318. [Google Scholar] [CrossRef]
- Kim, D.Y.; Kang, Y.M.; Cho, H.K.; Park, S.J.; Lee, M.H.; Lee, Y.J.; Kang, H.M. Development of a recombinant H9N2 influenza vaccine candidate against the Y280 lineage field virus and its protective efficacy. Vaccine 2021, 39, 6201–6205. [Google Scholar] [CrossRef]
- Park, S.J.; Kang, Y.M.; Cho, H.K.; Kim, D.Y.; Kim, S.; Bae, Y.; Kim, J.; Kim, G.; Lee, Y.J.; Kang, H.M. Cross-protective efficacy of inactivated whole influenza vaccines against Korean Y280 and Y439 lineage H9N2 viruses in mice. Vaccine 2021, 39, 6213–6220. [Google Scholar] [CrossRef]
- Song, J.M.; Lee, Y.J.; Jeong, O.M.; Kang, H.M.; Kim, H.R.; Kwon, J.H.; Kim, J.H.; Seong, B.L.; Kim, Y.J. Generation and evaluation of reassortant influenza vaccines made by reverse genetics for H9N2 avian influenza in Korea. Vet. Microbiol. 2008, 130, 268–276. [Google Scholar] [CrossRef]
- Nagy, A.; Lee, J.; Mena, I.; Henningson, J.; Li, Y.; Ma, J.; Duff, M.; Li, Y.; Lang, Y.; Yang, J.; et al. Recombinant Newcastle disease virus expressing H9 HA protects chickens against heterologous avian influenza H9N2 virus challenge. Vaccine 2016, 34, 2537–2545. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.Y.; Shang, Y.H.; Yao, H.X.; Cui, B.A.; Zhang, H.Y.; Wang, Z.X.; Wang, Y.D.; Chao, A.J.; Duan, T.Y. Immune responses of chickens inoculated with a recombinant fowlpox vaccine coexpressing HA of H9N2 avian influenza virus and chicken IL-18. Antivir. Res. 2011, 91, 50–56. [Google Scholar] [CrossRef]
- Liu, L.; Wang, T.; Wang, M.; Tong, Q.; Sun, Y.; Pu, J.; Sun, H.; Liu, J. Recombinant turkey herpesvirus expressing H9 hemagglutinin providing protection against H9N2 avian influenza. Virology 2019, 529, 7–15. [Google Scholar] [CrossRef]
- Lee, D.H.; Park, J.K.; Lee, Y.N.; Song, J.M.; Kang, S.M.; Lee, J.B.; Park, S.Y.; Choi, I.S.; Song, C.S. H9N2 avian influenza virus-like particle vaccine provides protective immunity and a strategy for the differentiation of infected from vaccinated animals. Vaccine 2011, 29, 4003–4007. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Genzel, Y.; Reichl, U. Continuous cell lines as a production system for influenza vaccines. Expert Rev. Vaccines 2009, 8, 1681–1692. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Jia, H.; Lai, H.; Liu, X.; Tan, W.S. Highly efficient production of an influenza H9N2 vaccine using MDCK suspension cells. Bioresour. Bioprocess 2020, 7, 63. [Google Scholar] [CrossRef]
- Wong, S.S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramakrishnan, M.A. Determination of 50% endpoint titer using a simple formula. World J. Virol. 2016, 5, 85–86. [Google Scholar] [CrossRef] [PubMed]
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Ping, J.; Lopes, T.J.S.; Nidom, C.A.; Ghedin, E.; Macken, C.A.; Fitch, A.; Imai, M.; Maher, E.A.; Neumann, G.; Kawaoka, Y. Development of high-yield influenza A virus vaccine viruses. Nat. Commun. 2015, 6, 8148. [Google Scholar] [CrossRef] [Green Version]
- Webby, R.J.; Perez, D.R.; Coleman, J.S.; Guan, Y.; Knight, J.H.; Govorkova, E.A.; McClain-Moss, L.R.; Peiris, J.S.; Rehg, J.E.; Tuomanen, E.I.; et al. Responsiveness to a pandemic alert: Use of reverse genetics for rapid development of influenza vaccines. Lancet 2004, 363, 1099–1103. [Google Scholar] [CrossRef]
- Swayne, D.E. Principles for vaccine protection in chickens and domestic waterfowl against avian influenza. Ann. N. Y. Acad. Sci. 2006, 1081, 174–181. [Google Scholar] [CrossRef]
- Zhang, A.; Lai, H.; Xu, J.; Huang, W.; Liu, Y.; Zhao, D.; Chen, R. Evaluation of the protective efficacy of Poly I:C as an adjuvant for H9N2 subtype avian influenza inactivated vaccine and its mechanism of action in ducks. PLoS ONE 2017, 12, e0170681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tosh, C.; Nagarajan, S.; Behera, P.; Rajukumar, K.; Purohit, K.; Kamal, R.P.; Murugkar, H.V.; Gounalan, S.; Pattnaik, B.; Vanamayya, P.R.; et al. Genetic analysis of H9N2 avian influenza viruses isolated from India. Arch. Virol. 2008, 153, 1433–1439. [Google Scholar] [CrossRef] [PubMed]
- Swayne, D.E.; Perdue, M.; Beck, J.R.; Garcia, M.; Suarez, D.L. Vaccine protect chickens against H5 highly pathogenic avian influenza in the face of genetic changes in field viruses over multiple years. Vet. Microbiol. 2000, 74, 165–172. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Passage Number | LBM564 | |
|---|---|---|
| HA Unit | EID50/0.1 mL | |
| CE2 | 16 | 106.5 |
| CE5 | 512 | 107.2 |
| CE10 | 512 | 108.2 |
| CE15 | 1024 | 107.7 |
| CE20 | 1024 | 107.5 |
| CE30 | 1024 | 108.4 |
| CE40 | 1024 | 108.5 |
| Vaccine | Vaccination | HI Titer (log2) | |||
|---|---|---|---|---|---|
| No. of Chickens | Vaccine Dose (EID50/0.1 mL) | Vaccine Route | 2wpv | 3wpv | |
| vac564 | 8 | 108.0 | IM | 7.0 ± 1.2 * | 8.0 ± 1.2 * |
| 107.5 | 1.4 ± 1.3 ** | 2.9 ± 2.1 ** | |||
| Sham | PBS | - 1 | - | ||
| Group | Vaccine | No. of Chickens 1 (HI Titer at 3 wpv, log2) | Challenge Virus (Lineage) | Virus Shedding (log10EID50/0.1 mL) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1dpi | 3dpi | 5dpi | 7dpi | 10dpi | 14dpi | ||||||||||
| OP | CL | OP | CL | OP | CL | OP | CL | OP | CL | OP | CL | ||||
| Vaccinated | vac564 | 8/8 (9.1 ± 1.0) | LBM564 (Y439) | 0/8 3,** (-) 4 | 0/8 (-) | 0/8 ** (-) | 0/8 (-) | 0/8 ** (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) |
| Sham | 0/8 (-) 2 | 7/8 (2.8) | 0/8 (-) | 8/8 (3.5) | 1/8 (1.4) | 8/8 (2.9) | 3/8 (3.5) | 0/8 (-) | 2/8 (3.2) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | ||
| Vaccinated | 8/8 (9.8 ± 1.0) | LBM314 (Y280) | 8/8 (3.6) | 0/8 (-) | 8/8 (4.6) | 0/8 * (-) | 8/8 (2.4) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | |
| Sham | 0/8 (-) | 7/8 (3.8) | 0/8 (-) | 8/8 (5.2) | 4/8 (2.4) | 8/8 (3.2) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | ||
| Group | Vaccine | Challenge Virus (Lineage) | Virus Replication in Tissues (log10EID50/0.1 mL) | |||||
|---|---|---|---|---|---|---|---|---|
| Trachea | Lung | Kidney | Spleen | Liver | Cecal Tonsil | |||
| Vaccinated | vac564 | LBM564 (Y439) | 0/8 1 (-) 2 | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 * (-) |
| Sham | 0/8 (-) | 0/8 (-) | 0/8 (-) | 1/8 (1.4) | 0/8 (-) | 4/8 (2.6) | ||
| Vaccinated | LBM314 (Y280) | 4/8 (2.5) | 1/8 (1.2) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 1/8 (1.0) | |
| Sham | 6/8 (2.2) | 1/8 (1.4) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 3/8 (1.5) | ||
| Group | Vaccine | No. of Chickens 1 (HI Titer at 3 wpv, log2) | Challenge Virus (Lineage) | Virus Shedding (log10EID50/0.1 mL) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 1dpi | 3dpi | 5dpi | 7dpi | 10dpi | 14dpi | ||||||||||
| OP | CL | OP | CL | OP | CL | OP | CL | OP | CL | OP | CL | ||||
| Vaccinated | G1 | 8/8 (1.3 ± 0.8) | LBM564 (Y439) | 8/8 3 (2.6) | 0/8 (-) 4 | 8/8 (2.2) | 0/8 ** (-) | 6/8 (3.5) | 1/8 ** (1.5) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) |
| Sham | 0/8 (-) 2 | 8/8 (2.8) | 0/8 (-) | 8/8 (2.8) | 7/8 (4.1) | 7/8 (3.5) | 7/8 (4.5) | 0/8 (-) | 2/8 (2.8) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | ||
| Vaccinated | 8/8 (8.5 ± 0.7) | LBM314 (Y280) | 0/8 ** (-) | 0/8 (-) | 3/8 (4.8) | 0/8 ** (-) | 7/8 (1.5) | 1/8 (1.2) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | |
| Sham | 0/8 (-) | 7/8 (3.8) | 0/8 (-) | 6/8 (4.6) | 7/8 (2.6) | 8/8 (3.6) | 4/8 (2.4) | 0/8 (-) | 1/8 (1.7) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | ||
| Group | Vaccine | Challenge Virus (Lineage) | Virus Replication in Tissues (log10EID50/0.1 mL) | |||||
|---|---|---|---|---|---|---|---|---|
| Trachea | Lung | Kidney | Spleen | Liver | Cecal Tonsil | |||
| Vaccinated | G1 | LBM564 (Y439) | 0/8 1 (-) 2 | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) |
| Sham | 1/8 (1.4) | 0/8 (-) | 1/8 (-) | 1/8 (1.0) | 0/8 (-) | 2/8 (1.2) | ||
| Vaccinated | LBM314 (Y280) | 0/8 ** (-) | 0/8 ** (-) | 0/8 * (-) | 0/8 (-) | 0/8 (-) | 0/8 (-) | |
| Sham | 7/8 (2.6) | 6/8 (1.2) | 5/8 (-) | 0/8 (-) | 0/8 (-) | 1/8 (1.0) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cho, H.-K.; Kang, Y.-M.; Sagong, M.; Kim, J.; Kim, H.; An, S.; Lee, Y.-J.; Kang, H.-M. Protection of SPF Chickens by H9N2 Y439 and G1 Lineage Vaccine against Homologous and Heterologous Viruses. Vaccines 2023, 11, 538. https://doi.org/10.3390/vaccines11030538
Cho H-K, Kang Y-M, Sagong M, Kim J, Kim H, An S, Lee Y-J, Kang H-M. Protection of SPF Chickens by H9N2 Y439 and G1 Lineage Vaccine against Homologous and Heterologous Viruses. Vaccines. 2023; 11(3):538. https://doi.org/10.3390/vaccines11030538
Chicago/Turabian StyleCho, Hyun-Kyu, Yong-Myung Kang, Mingeun Sagong, Juhun Kim, Hyunjun Kim, Sungjun An, Youn-Jeong Lee, and Hyun-Mi Kang. 2023. "Protection of SPF Chickens by H9N2 Y439 and G1 Lineage Vaccine against Homologous and Heterologous Viruses" Vaccines 11, no. 3: 538. https://doi.org/10.3390/vaccines11030538