
Lactobacillus acidophilus Expressing Murine Rotavirus VP8 and Mucosal Adjuvants Induce Virus-Specific Immune Responses
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. rLA Vaccine Strain Construction
2.2. Bacterial Growth Conditions
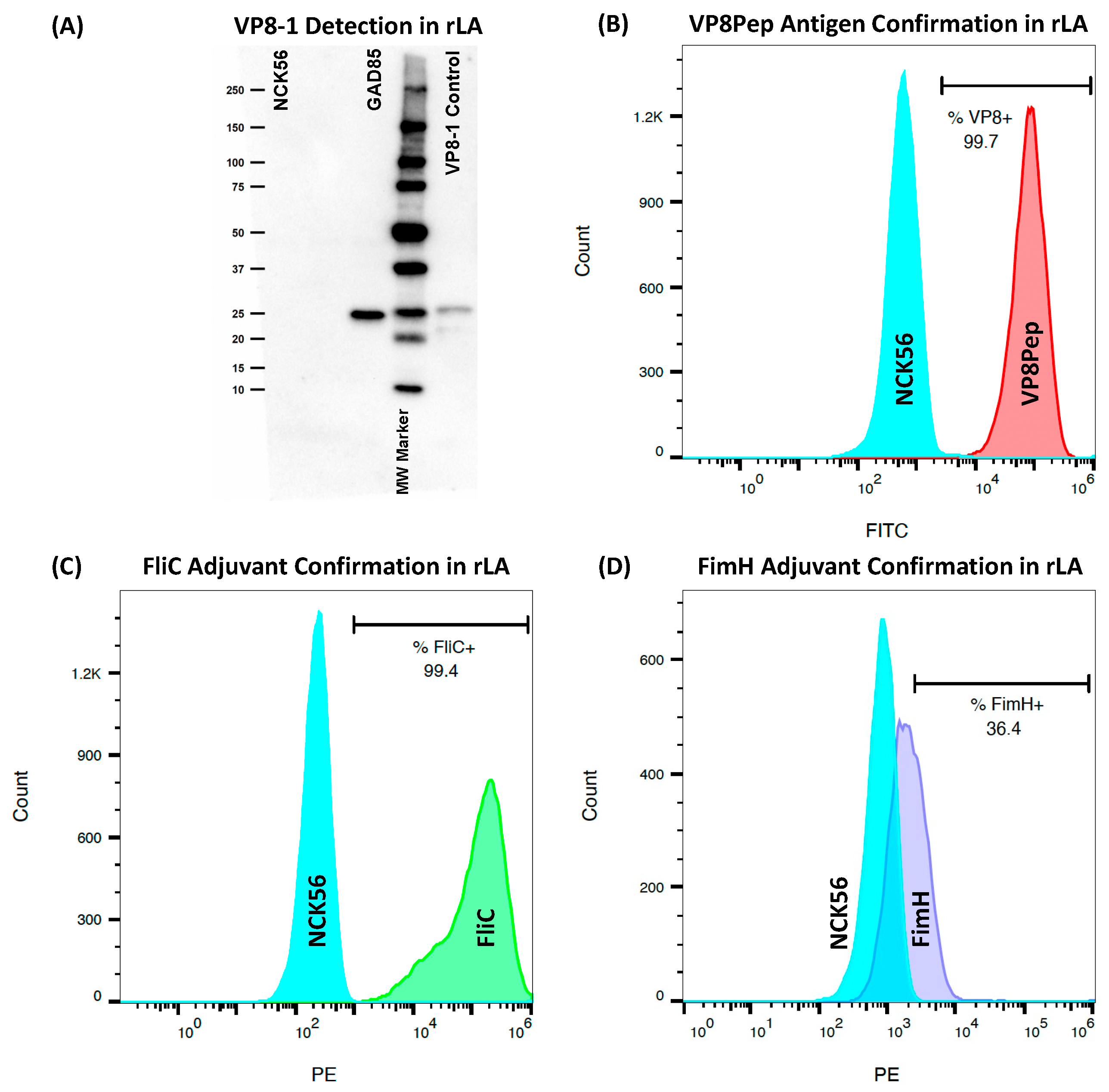
2.3. Confirmation of rLA Antigen and Adjuvant Surface Expression by Flow Cytometry
2.4. Confirmation of Cytosolic Accumulation of VP8-1 Antigen by SDS-PAGE Western Blot
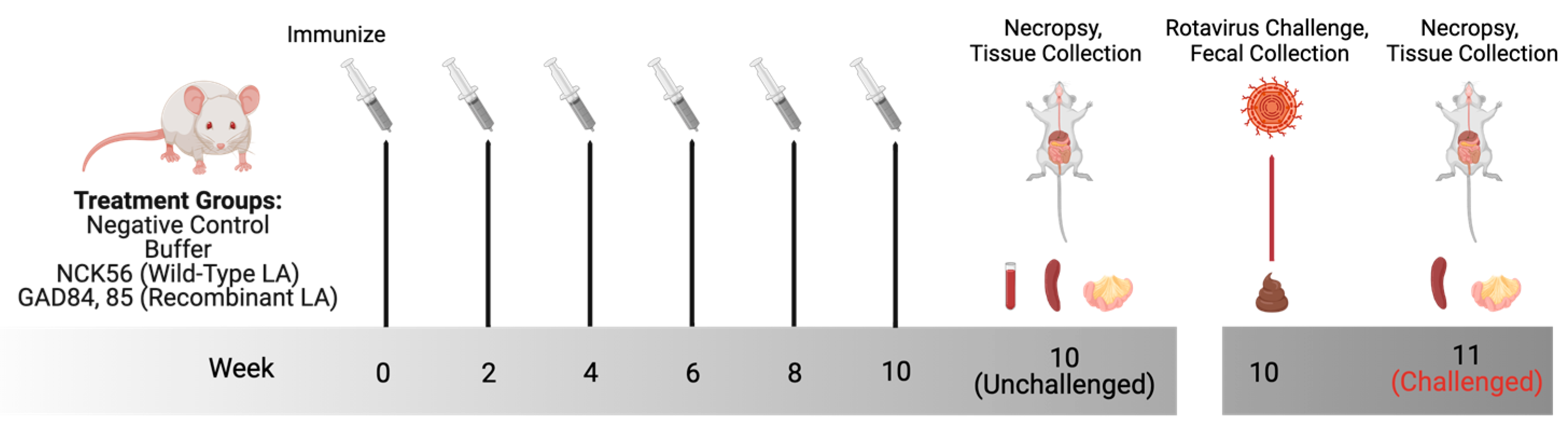
2.5. Animal Ethics and Murine Oral Immunization
2.6. Murine Rotavirus Challenge
2.7. Tissue Collection, Cell Isolation, and Antibody-Secreting Cell (ASC) FluoroSpot Assay
2.8. Fecal Antigen Shedding ELISA
2.9. Tissue-Culture-Adapted Murine RV (ETD) Propagation
2.10. ETD Immunoperoxidase Focus Reduction Neutralization Assay
2.11. ETD-Infected MA104 Cell-Based Antibody ELISA
2.12. Statistical Analyses
3. Results
3.1. rLA Expresses Rotavirus Antigens and Mucosal Adjuvants
3.2. rLA Vaccination Induces Antirotavirus Serum IgG
3.3. Rotavirus Challenge after Vaccination Boosted Antigen-Specific Immunity
3.4. rLA Vaccination Delays Rotavirus Shedding
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Troeger, C.; Khalil, I.A.; Rao, P.C.; Cao, S.; Blacker, B.F.; Ahmed, T.; Armah, G.; Bines, J.E.; Brewer, T.G.; Colombara, D.V.; et al. Rotavirus Vaccination and the Global Burden of Rotavirus Diarrhea Among Children Younger Than 5 Years. JAMA Pediatr. 2018, 172, 958–965. [Google Scholar] [CrossRef]
- Du, Y.; Chen, C.; Zhang, X.; Yan, D.; Jiang, D.; Liu, X.; Yang, M.; Ding, C.; Lan, L.; Hecht, R.; et al. Global burden and trends of rotavirus infection-associated deaths from 1990 to 2019: An observational trend study. Virol. J. 2022, 19, 166. [Google Scholar] [CrossRef] [PubMed]
- Burke, R.M.; Tate, J.E.; Parashar, U.D. Global Experience with Rotavirus Vaccines. J. Infect. Dis. 2021, 224, S792–S800. [Google Scholar] [CrossRef] [PubMed]
- International Vaccine Access Center (IVAC), Johns Hopkins Bloomberg School of Public Health. VIEW-Hub. Available online: www.view-hub.org (accessed on 30 October 2023).
- WHO. Rotavirus Vaccines: WHO Position Paper—July 2021. Wkly. Epidemiol. Rec. 2021, 96, 301–320. [Google Scholar]
- Soares-Weiser, K.; Bergman, H.; Henschke, N.; Pitan, F.; Cunliffe, N. Vaccines for preventing rotavirus diarrhoea: Vaccines in use. Cochrane Database Syst. Rev. 2019, 3, Cd008521. [Google Scholar] [CrossRef] [PubMed]
- Otero, C.E.; Langel, S.N.; Blasi, M.; Permar, S.R. Maternal antibody interference contributes to reduced rotavirus vaccine efficacy in developing countries. PLoS Pathog. 2020, 16, e1009010. [Google Scholar] [CrossRef]
- O’Ryan, M. Rotavirus Vaccines: A story of success with challenges ahead. F1000Research 2017, 6, 1517. [Google Scholar] [CrossRef]
- Burnett, E.; Parashar, U.; Tate, J. Rotavirus Vaccines: Effectiveness, Safety, and Future Directions. Paediatr. Drugs 2018, 20, 223–233. [Google Scholar] [CrossRef]
- Steele, A.D.; Victor, J.C.; Carey, M.E.; Tate, J.E.; Atherly, D.E.; Pecenka, C.; Diaz, Z.; Parashar, U.D.; Kirkwood, C.D. Experiences with rotavirus vaccines: Can we improve rotavirus vaccine impact in developing countries? Hum. Vaccin. Immunother. 2019, 15, 1215–1227. [Google Scholar] [CrossRef]
- Yadav, D.K.; Yadav, N.; Khurana, S.M.P. Chapter 26—Vaccines: Present status and applications. In Animal Biotechnology; Verma, A.S., Singh, A., Eds.; Academic Press: San Diego, CA, USA, 2014; pp. 491–508. [Google Scholar]
- Mak, T.W.; Saunders, M.E. 23—Vaccines and clinical immunization. In The Immune Response; Mak, T.W., Saunders, M.E., Eds.; Academic Press: Burlington, VA, USA, 2006; pp. 695–749. [Google Scholar]
- Blutt, S.E.; Warfield, K.L.; Lewis, D.E.; Conner, M.E. Early response to rotavirus infection involves massive B cell activation. J. Immunol. 2002, 168, 5716–5721. [Google Scholar] [CrossRef]
- Moser, C.A.; Offit, P.A. Distribution of rotavirus-specific memory B cells in gut-associated lymphoid tissue after primary immunization. J. Gen. Virol. 2001, 82, 2271–2274. [Google Scholar] [CrossRef]
- Saif, L.; Yuan, L.; Ward, L.; To, T. Comparative studies of the pathogenesis, antibody immune responses, and homologous protection to porcine and human rotaviruses in gnotobiotic piglets. Adv. Exp. Med. Biol. 1997, 412, 397–403. [Google Scholar] [CrossRef]
- Offit, P.A.; Clark, H.F. Protection against rotavirus-induced gastroenteritis in a murine model by passively acquired gastrointestinal but not circulating antibodies. J. Virol. 1985, 54, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Blutt, S.E.; Miller, A.D.; Salmon, S.L.; Metzger, D.W.; Conner, M.E. IgA is important for clearance and critical for protection from rotavirus infection. Mucosal. Immunol. 2012, 5, 712–719. [Google Scholar] [CrossRef] [PubMed]
- Ismail, B. The use of probiotics as vaccine vectors to prevent viral infections. In New Insights on Antiviral Probiotics: From Research to Applications; Al Kassaa, I., Ed.; Springer International Publishing: Cham, Switzerland, 2017; pp. 47–60. [Google Scholar]
- Vlasova, A.N.; Kandasamy, S.; Chattha, K.S.; Rajashekara, G.; Saif, L.J. Comparison of probiotic lactobacilli and bifidobacteria effects, immune responses and rotavirus vaccines and infection in different host species. Vet. Immunol. Immunopathol. 2016, 172, 72–84. [Google Scholar] [CrossRef] [PubMed]
- Lycke, N. Recent progress in mucosal vaccine development: Potential and limitations. Nat. Rev. Immunol. 2012, 12, 592–605. [Google Scholar] [CrossRef] [PubMed]
- Isolauri, E.; Joensuu, J.; Suomalainen, H.; Luomala, M.; Vesikari, T. Improved immunogenicity of oral D x RRV reassortant rotavirus vaccine by Lactobacillus casei GG. Vaccine 1995, 13, 310–312. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Wen, K.; Li, G.; Yang, X.; Kocher, J.; Bui, T.; Jones, D.; Pelzer, K.; Clark-Deener, S.; Yuan, L. Dual functions of Lactobacillus acidophilus NCFM as protection against rotavirus diarrhea. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 169–176. [Google Scholar] [CrossRef]
- Zhang, W.; Azevedo, M.S.; Wen, K.; Gonzalez, A.; Saif, L.J.; Li, G.; Yousef, A.E.; Yuan, L. Probiotic Lactobacillus acidophilus enhances the immunogenicity of an oral rotavirus vaccine in gnotobiotic pigs. Vaccine 2008, 26, 3655–3661. [Google Scholar] [CrossRef]
- Parreno, V.; Bai, M.; Liu, F.; Jing, J.; Olney, E.; Li, G.; Wen, K.; Yang, X.; Castellucc, T.B.; Kocher, J.F.; et al. Probiotic as Adjuvant Significantly Improves Protection of the Lanzhou Trivalent Rotavirus Vaccine against Heterologous Challenge in a Gnotobiotic Pig Model of Human Rotavirus Infection and Disease. Vaccines 2022, 10, 1529. [Google Scholar] [CrossRef]
- Yuan, L.; Saif, L.J. Induction of mucosal immune responses and protection against enteric viruses: Rotavirus infection of gnotobiotic pigs as a model. Vet. Immunol. Immunopathol. 2002, 87, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Bron, P.A.; van Baarlen, P.; Kleerebezem, M. Emerging molecular insights into the interaction between probiotics and the host intestinal mucosa. Nat. Rev. Microbiol. 2012, 10, 66–78. [Google Scholar] [CrossRef]
- Wells, J.M. Immunomodulatory mechanisms of lactobacilli. Microb. Cell Factories 2011, 10, S17. [Google Scholar] [CrossRef] [PubMed]
- Kajikawa, A.; Zhang, L.; LaVoy, A.; Bumgardner, S.; Klaenhammer, T.R.; Dean, G.A. Mucosal Immunogenicity of Genetically Modified Lactobacillus acidophilus Expressing an HIV-1 Epitope within the Surface Layer Protein. PLoS ONE 2015, 10, e0141713. [Google Scholar] [CrossRef] [PubMed]
- Qiao, X.; Li, G.; Wang, X.; Li, X.; Liu, M.; Li, Y. Recombinant porcine rotavirus VP4 and VP4-LTB expressed in Lactobacillus casei induced mucosal and systemic antibody responses in mice. BMC Microbiol. 2009, 9, 249. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, H.; Zhao, Y.; Sui, L.; Li, F.; Zhang, H.; Li, J.; Jiang, Y.; Cui, W.; Ding, G.; et al. Auxotrophic Lactobacillus Expressing Porcine Rotavirus VP4 Constructed Using CRISPR-Cas9D10A System Induces Effective Immunity in Mice. Vaccines 2022, 10, 1510. [Google Scholar] [CrossRef] [PubMed]
- Fox, B.E.; Vilander, A.C.; Gilfillan, D.; Dean, G.A.; Abdo, Z. Oral Vaccination Using a Probiotic Vaccine Platform Combined with Prebiotics Impacts Immune Response and the Microbiome. Vaccines 2022, 10, 1465. [Google Scholar] [CrossRef]
- Kajikawa, A.; Zhang, L.; Long, J.; Nordone, S.; Stoeker, L.; LaVoy, A.; Bumgardner, S.; Klaenhammer, T.; Dean, G. Construction and immunological evaluation of dual cell surface display of HIV-1 gag and Salmonella enterica serovar Typhimurium FliC in Lactobacillus acidophilus for vaccine delivery. Clin. Vaccine Immunol. 2012, 19, 1374–1381. [Google Scholar] [CrossRef]
- Xue, M.; Yu, L.; Che, Y.; Lin, H.; Zeng, Y.; Fang, M.; Li, T.; Ge, S.; Xia, N. Characterization and protective efficacy in an animal model of a novel truncated rotavirus VP8 subunit parenteral vaccine candidate. Vaccine 2015, 33, 2606–2613. [Google Scholar] [CrossRef]
- Kovacs-Nolan, J.; Yoo, D.; Mine, Y. Fine mapping of sequential neutralization epitopes on the subunit protein VP8 of human rotavirus. Biochem. J. 2003, 376, 269–275. [Google Scholar] [CrossRef]
- Clarke, E.; Desselberger, U. Correlates of protection against human rotavirus disease and the factors influencing protection in low-income settings. Mucosal. Immunol. 2015, 8, 1–17. [Google Scholar] [CrossRef]
- Vilander, A.C.; Shelton, K.; LaVoy, A.; Dean, G.A. Expression of E. coli FimH Enhances Trafficking of an Orally Delivered Lactobacillus acidophilus Vaccine to Immune Inductive Sites via Antigen-Presenting Cells. Vaccines 2023, 11, 1162. [Google Scholar] [CrossRef] [PubMed]
- Rhee, J.H.; Lee, S.E.; Kim, S.Y. Mucosal vaccine adjuvants update. Clin. Exp. Vaccine Res. 2012, 1, 50–63. [Google Scholar] [CrossRef] [PubMed]
- Barefoot, S.F.; Klaenhammer, T.R. Detection and activity of lactacin B, a bacteriocin produced by Lactobacillus acidophilus. Appl. Environ. Microbiol. 1983, 45, 1808–1815. [Google Scholar] [CrossRef] [PubMed]
- Sanders, M.E.; Klaenhammer, T.R. Invited Review: The Scientific Basis of Lactobacillus acidophilus NCFM Functionality as a Probiotic. J. Dairy Sci. 2001, 84, 319–331. [Google Scholar] [CrossRef]
- Douglas, G.L.; Goh, Y.J.; Klaenhammer, T.R. Integrative food grade expression system for lactic acid bacteria. In Strain Engineering: Methods and Protocols; Williams, J.A., Ed.; Humana Press: Totowa, NJ, USA, 2011; pp. 373–387. [Google Scholar]
- Goh, Y.J.; Azcárate-Peril, M.A.; O’Flaherty, S.; Durmaz, E.; Valence, F.; Jardin, J.; Lortal, S.; Klaenhammer, T.R. Development and application of a upp-based counterselective gene replacement system for the study of the S-layer protein SlpX of Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2009, 75, 3093–3105. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.C.; Aoyama, K.; Klaenhammer, T.R. Electrotransformation of Lactobacillus acidophilus group A1. FEMS Microbiol. Lett. 1996, 138, 233–237. [Google Scholar] [CrossRef]
- Kajikawa, A.; Nordone, S.K.; Zhang, L.; Stoeker, L.L.; LaVoy, A.S.; Klaenhammer, T.R.; Dean, G.A. Dissimilar properties of two recombinant Lactobacillus acidophilus strains displaying Salmonella FliC with different anchoring motifs. Appl. Environ. Microbiol. 2011, 77, 6587–6596. [Google Scholar] [CrossRef]
- Law, J.; Buist, G.; Haandrikman, A.; Kok, J.; Venema, G.; Leenhouts, K. A system to generate chromosomal mutations in Lactococcus lactis which allows fast analysis of targeted genes. J. Bacteriol. 1995, 177, 7011–7018. [Google Scholar] [CrossRef]
- Lindenbach, B.D. Measuring HCV infectivity produced in cell culture and in vivo. Methods Mol Biol 2009, 510, 329–336. [Google Scholar] [CrossRef]
- Feng, N.; Yasukawa, L.L.; Sen, A.; Greenberg, H.B. Permissive replication of homologous murine rotavirus in the mouse intestine is primarily regulated by VP4 and NSP1. J. Virol. 2013, 87, 8307–8316. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 8 November 2023).
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/ (accessed on 8 November 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer-Verlag: New York, NY, USA, 2016. [Google Scholar]
- Douglas, G.L.; Klaenhammer, T.R. Directed chromosomal integration and expression of the reporter gene gusA3 in Lactobacillus acidophilus NCFM. Appl. Environ. Microbiol. 2011, 77, 7365–7371. [Google Scholar] [CrossRef] [PubMed]
- O’Flaherty, S.; Klaenhammer, T.R. Multivalent Chromosomal Expression of the Clostridium botulinum Serotype A Neurotoxin Heavy-Chain Antigen and the Bacillus anthracis Protective Antigen in Lactobacillus acidophilus. Appl. Environ. Microbiol. 2016, 82, 6091–6101. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.J.; Ma, G.P.; Li, G.W.; Qiao, X.Y.; Ge, J.W.; Tang, L.J.; Liu, M.; Liu, L.W. Oral vaccination with the porcine rotavirus VP4 outer capsid protein expressed by Lactococcus lactis induces specific antibody production. J. Biomed. Biotechnol. 2010, 2010, 708460. [Google Scholar] [CrossRef]
- Stoeker, L.; Nordone, S.; Gunderson, S.; Zhang, L.; Kajikawa, A.; LaVoy, A.; Miller, M.; Klaenhammer, T.R.; Dean, G.A. Assessment of Lactobacillus gasseri as a Candidate Oral Vaccine Vector. Clin. Vaccine Immunol. 2011, 18, 1834–1844. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Iosef, C.; Azevedo, M.S.; Kim, Y.; Qian, Y.; Geyer, A.; Nguyen, T.V.; Chang, K.O.; Saif, L.J. Protective immunity and antibody-secreting cell responses elicited by combined oral attenuated Wa human rotavirus and intranasal Wa 2/6-VLPs with mutant Escherichia coli heat-labile toxin in gnotobiotic pigs. J. Virol. 2001, 75, 9229–9238. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Azevedo, M.S.P.; Gonzalez, A.M.; Jeong, K.-i.; Van Nguyen, T.; Lewis, P.; Iosef, C.; Herrmann, J.E.; Saif, L.J. Mucosal and systemic antibody responses and protection induced by a prime/boost rotavirus-DNA vaccine in a gnotobiotic pig model. Vaccine 2005, 23, 3925–3936. [Google Scholar] [CrossRef]
- Fransen, F.; Zagato, E.; Mazzini, E.; Fosso, B.; Manzari, C.; El Aidy, S.; Chiavelli, A.; D’Erchia, A.M.; Sethi, M.K.; Pabst, O.; et al. BALB/c and C57BL/6 Mice Differ in Polyreactive IgA Abundance, which Impacts the Generation of Antigen-Specific IgA and Microbiota Diversity. Immunity 2015, 43, 527–540. [Google Scholar] [CrossRef]
- McNeal, M.M.; Rae, M.N.; Bean, J.A.; Ward, R.L. Antibody-Dependent and -Independent Protection following Intranasal Immunization of Mice with Rotavirus Particles. J. Virol. 1999, 73, 7565–7573. [Google Scholar] [CrossRef]
- O’Neal, C.M.; Crawford, S.E.; Estes, M.K.; Conner, M.E. Rotavirus virus-like particles administered mucosally induce protective immunity. J. Virol. 1997, 71, 8707–8717. [Google Scholar] [CrossRef]
- Leshem, E.; Lopman, B.; Glass, R.; Gentsch, J.; Bányai, K.; Parashar, U.; Patel, M. Distribution of rotavirus strains and strain-specific effectiveness of the rotavirus vaccine after its introduction: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 847–856. [Google Scholar] [CrossRef] [PubMed]
- Amin, A.B.; Tate, J.E.; Waller, L.A.; Lash, T.L.; Lopman, B.A. Monovalent Rotavirus Vaccine Efficacy Against Different Rotavirus Genotypes: A Pooled Analysis of Phase II and III Trial Data. Clin. Infect. Dis. 2022, 76, e1150–e1156. [Google Scholar] [CrossRef] [PubMed]
- PATH. PATH Announces Early Closure of Pivotal Phase 3 Study of an Injectable Rotavirus Vaccine Candidate. 2022. Available online: https://www.path.org/media-center/path-announces-early-closure-of-pivotal-phase-3-study-of-an-injectable-rotavirus-vaccine-candidate/#:~:text=At%20this%20time%20there%20is,enrolled%20approximately%203%2C730%20healthy%20infants (accessed on 8 November 2023).
- Groome, M.J.; Koen, A.; Fix, A.; Page, N.; Jose, L.; Madhi, S.A.; McNeal, M.; Dally, L.; Cho, I.; Power, M.; et al. Safety and immunogenicity of a parenteral P2-VP8-P [8] subunit rotavirus vaccine in toddlers and infants in South Africa: A randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2017, 17, 843–853. [Google Scholar] [CrossRef] [PubMed]
- Groome, M.J.; Fairlie, L.; Morrison, J.; Fix, A.; Koen, A.; Masenya, M.; Jose, L.; Madhi, S.A.; Page, N.; McNeal, M.; et al. Safety and immunogenicity of a parenteral trivalent P2-VP8 subunit rotavirus vaccine: A multisite, randomised, double-blind, placebo-controlled trial. Lancet Infect. Dis. 2020, 20, 851–863. [Google Scholar] [CrossRef] [PubMed]
- Azagra-Boronat, I.; Massot-Cladera, M.; Knipping, K.; Garssen, J.; Ben Amor, K.; Knol, J.; Franch, À.; Castell, M.; Rodríguez-Lagunas, M.J.; Pérez-Cano, F.J. Strain-Specific Probiotic Properties of Bifidobacteria and Lactobacilli for the Prevention of Diarrhea Caused by Rotavirus in a Preclinical Model. Nutrients 2020, 12, 498. [Google Scholar] [CrossRef]
- Park, M.S.; Kwon, B.; Ku, S.; Ji, G.E. The Efficacy of Bifidobacterium longum BORI and Lactobacillus acidophilus AD031 Probiotic Treatment in Infants with Rotavirus Infection. Nutrients 2017, 9, 887. [Google Scholar] [CrossRef]
- Ward, R.L.; McNeal, M.M.; Sheridan, J.F. Development of an adult mouse model for studies on protection against rotavirus. J. Virol. 1990, 64, 5070–5075. [Google Scholar] [CrossRef]
- Cheever, F.S.; Mueller, J.H. Epidemic diarrheal disease of suckling mice: I. Manifestations, epidemiology, and attempts to transmit the disease. J. Exp. Med. 1947, 85, 405–416. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Components | Resistance | Reference |
|---|---|---|---|
| NCK56 | Wild-type lab LA NCFM strain | Erm | [38] |
| GAD84 | LA-expressing VP8Pep within SlpA and chromosomal VP8-1 downstream of enolase | None | This study |
| GAD85 | LA-expressing VP8Pep within SlpA, pFliC, and chromosomal VP8-1 and FimH downstream of enolase | Erm | This study |
| Group | N per Treatment | Treatment | Challenged with Rotavirus |
|---|---|---|---|
| Negative Control | 7 | No Intervention | No |
| Buffer | 8 | Bacterial Resuspension Media | Yes |
| NCK56 | 8 | Wild-type LA Control | Yes |
| GAD84 | 8 | Dual-Antigen rLA | No |
| GAD85 | 8 | Dual-Antigen and Dual-Adjuvant rLA | Yes |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilfillan, D.; Vilander, A.C.; Pan, M.; Goh, Y.J.; O’Flaherty, S.; Feng, N.; Fox, B.E.; Lang, C.; Greenberg, H.B.; Abdo, Z.; et al. Lactobacillus acidophilus Expressing Murine Rotavirus VP8 and Mucosal Adjuvants Induce Virus-Specific Immune Responses. Vaccines 2023, 11, 1774. https://doi.org/10.3390/vaccines11121774
Gilfillan D, Vilander AC, Pan M, Goh YJ, O’Flaherty S, Feng N, Fox BE, Lang C, Greenberg HB, Abdo Z, et al. Lactobacillus acidophilus Expressing Murine Rotavirus VP8 and Mucosal Adjuvants Induce Virus-Specific Immune Responses. Vaccines. 2023; 11(12):1774. https://doi.org/10.3390/vaccines11121774
Chicago/Turabian StyleGilfillan, Darby, Allison C. Vilander, Meichen Pan, Yong Jun Goh, Sarah O’Flaherty, Ningguo Feng, Bridget E. Fox, Callie Lang, Harry B. Greenberg, Zaid Abdo, and et al. 2023. "Lactobacillus acidophilus Expressing Murine Rotavirus VP8 and Mucosal Adjuvants Induce Virus-Specific Immune Responses" Vaccines 11, no. 12: 1774. https://doi.org/10.3390/vaccines11121774