
Ageing Curtails the Diversity and Functionality of Nascent CD8+ T Cell Responses against SARS-CoV-2
 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Donors
2.2. Peptides and Tetramers
2.3. In Vitro Priming of Human Antigen-Specific CD8+ T Cells
2.4. ELISpot Assay
2.5. Flow Cytometry
2.6. HLA–Peptide Binding Assay
2.7. Statistics
3. Results
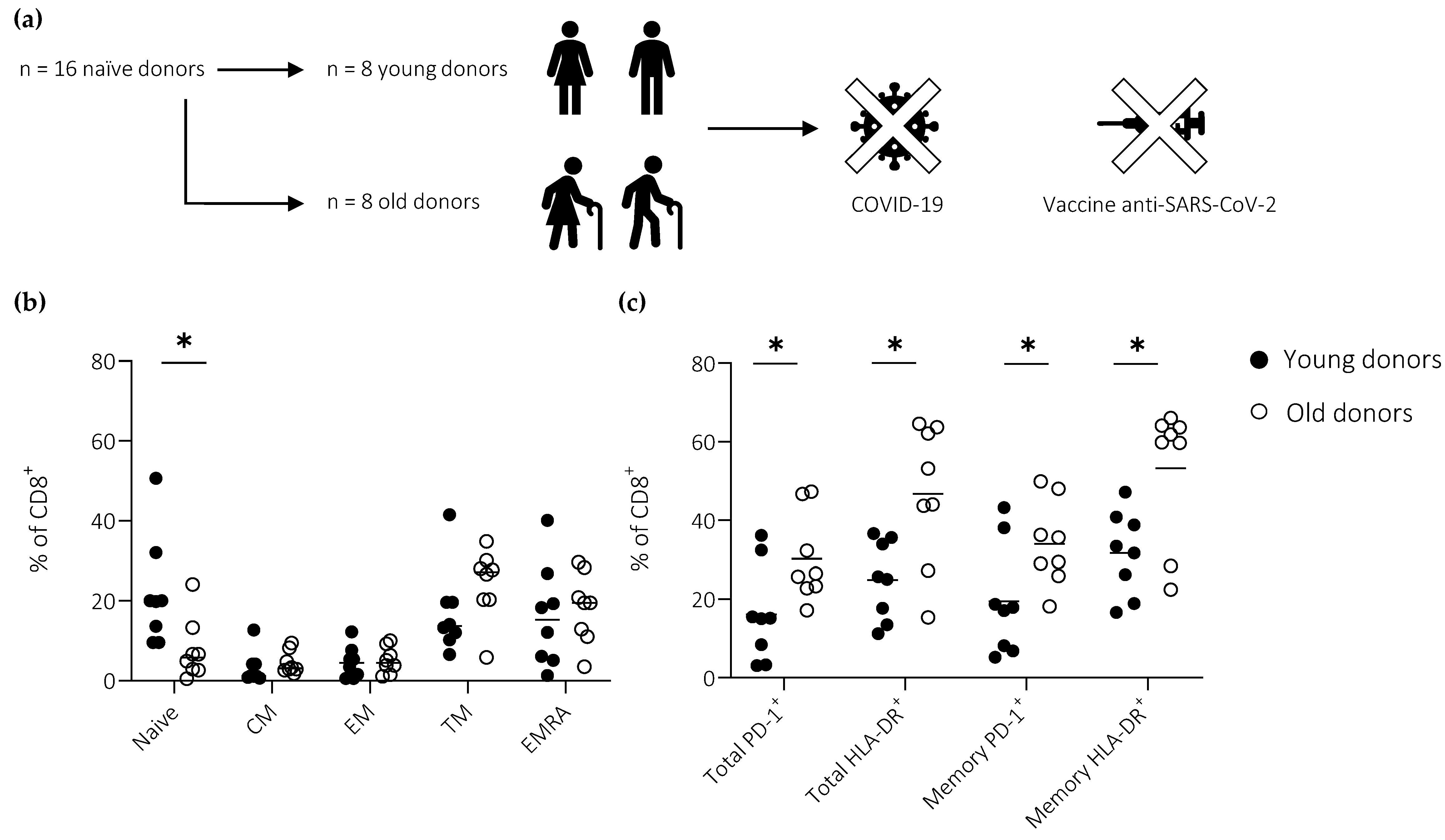
3.1. Cohort Description
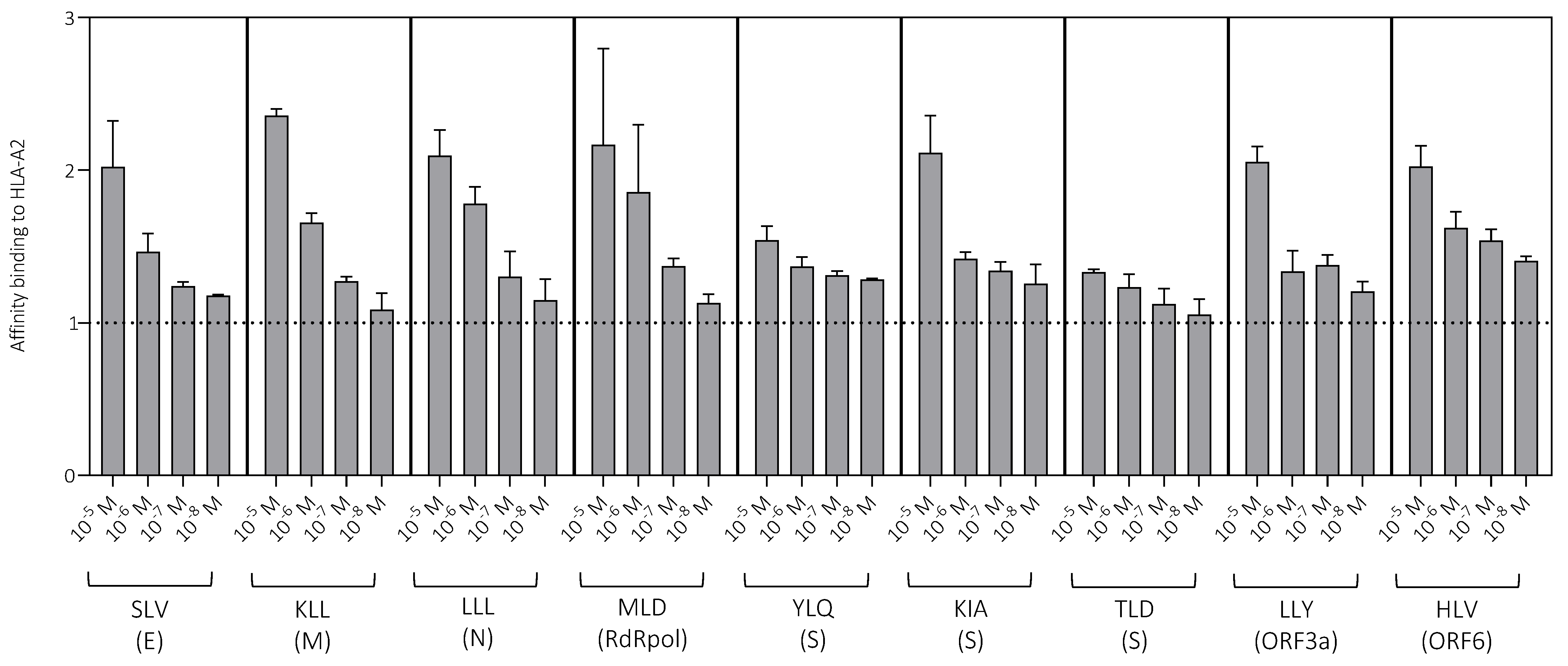
3.2. Peptide Characterization
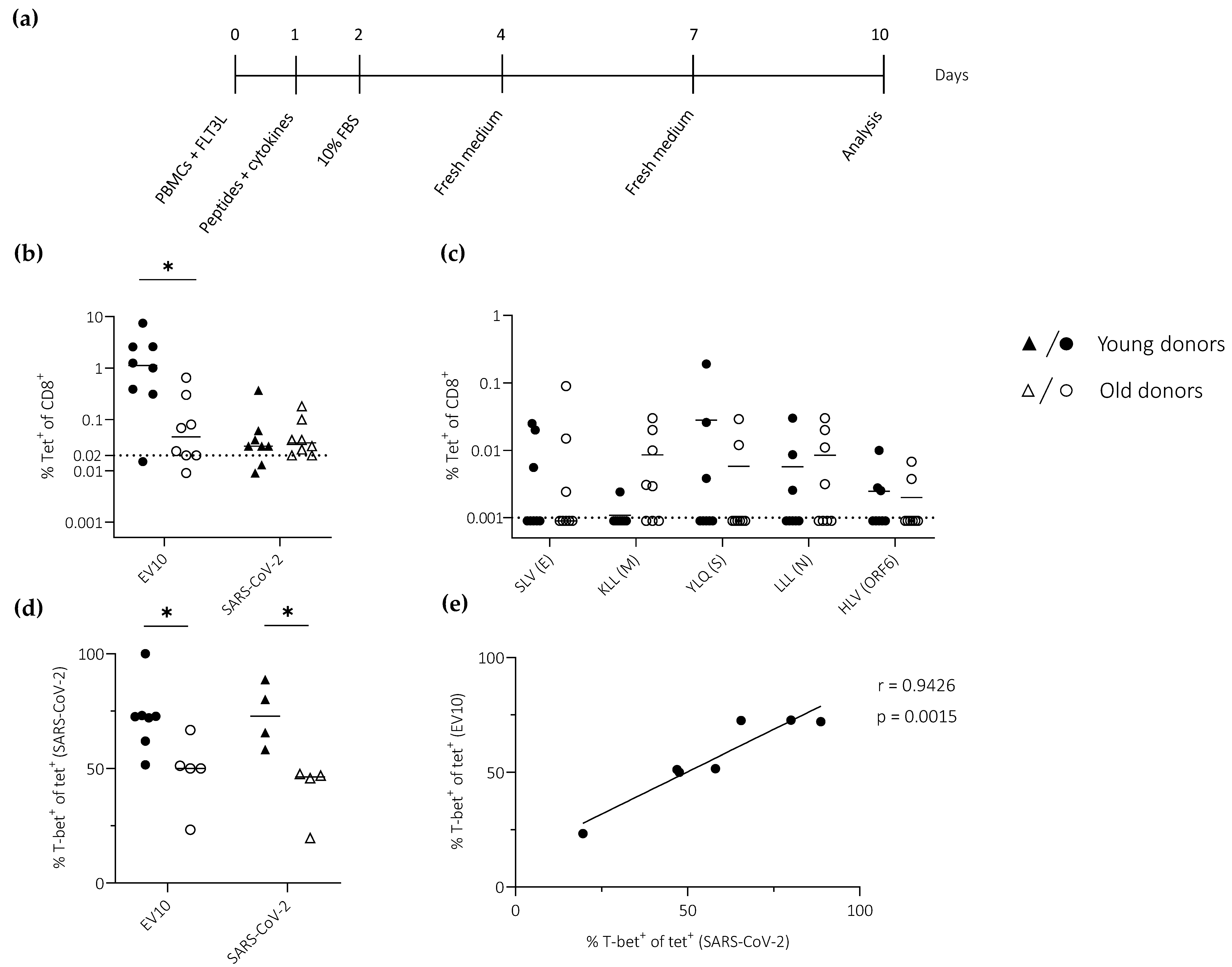
3.3. Ageing Impairs the Induction of Effector CD8+ T Cells Defined by the Expression of T-bet
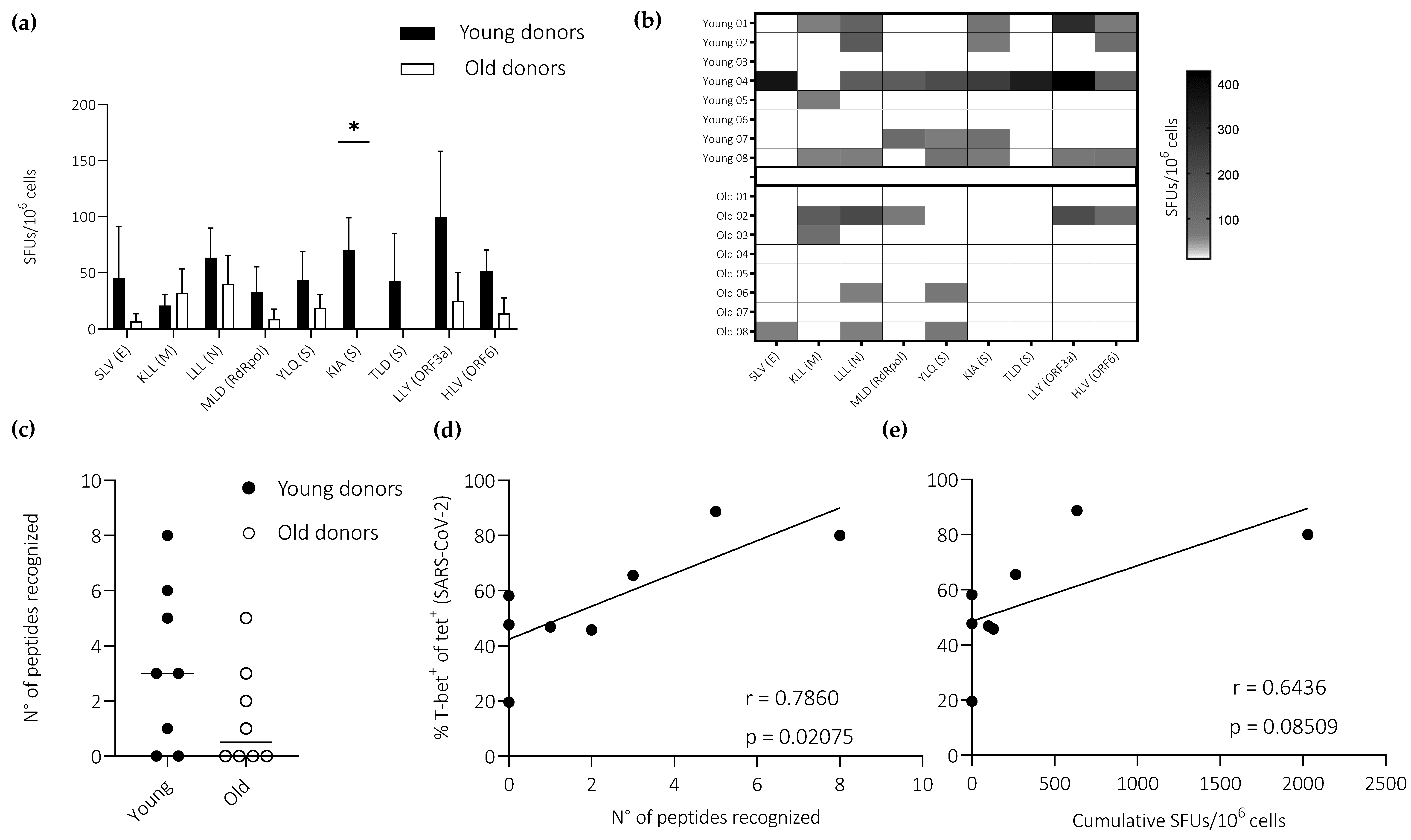
3.4. Ageing Impairs the Induction of Effector CD8+ T Cells Defined by the Expression of IFN-γ
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nicoli, F.; Paudel, D.; Solis-Soto, M.T. Old and new coronaviruses in the elderly. Aging 2021, 13, 12295–12296. [Google Scholar] [CrossRef] [PubMed]
- Nicoli, F.; Solis-Soto, M.T.; Paudel, D.; Marconi, P.; Gavioli, R.; Appay, V.; Caputo, A. Age-related decline of de novo T cell responsiveness as a cause of COVID-19 severity. Geroscience 2020, 42, 1015–1019. [Google Scholar] [CrossRef] [PubMed]
- Arregoces-Castillo, L.; Fernandez-Nino, J.; Rojas-Botero, M.; Palacios-Clavijo, A.; Galvis-Pedraza, M.; Rincon-Medrano, L.; Pinto-Alvarez, M.; Ruiz-Gomez, F.; Trejo-Valdivia, B. Effectiveness of COVID-19 vaccines in older adults in Colombia: A retrospective, population-based study of the ESPERANZA cohort. Lancet Healthy Longev. 2022, 3, e242–e252. [Google Scholar] [CrossRef] [PubMed]
- Verity, R.; Okell, L.C.; Dorigatti, I.; Winskill, P.; Whittaker, C.; Imai, N.; Cuomo-Dannenburg, G.; Thompson, H.; Walker, P.G.T.; Fu, H.; et al. Estimates of the severity of coronavirus disease 2019: A model-based analysis. Lancet Infect. Dis. 2020, 20, 669–677. [Google Scholar] [CrossRef]
- Hazeldine, J.; Lord, J.M. Innate immunesenescence: Underlying mechanisms and clinical relevance. Biogerontology 2015, 16, 187–201. [Google Scholar] [CrossRef]
- Cunha, L.L.; Perazzio, S.F.; Azzi, J.; Cravedi, P.; Riella, L.V. Remodeling of the immune response with aging: Immunosenescence and its potential impact on COVID-19 immune response. Front. Immunol. 2020, 11, 1748. [Google Scholar] [CrossRef]
- Rydyznski Moderbacher, C.; Ramirez, S.I.; Dan, J.M.; Grifoni, A.; Hastie, K.M.; Weiskopf, D.; Belanger, S.; Abbott, R.K.; Kim, C.; Choi, J.; et al. Antigen-specific adaptive immunity to SARS-CoV-2 in acute COVID-19 and associations with age and disease severity. Cell 2020, 183, 996–1012.e19. [Google Scholar] [CrossRef]
- Nicoli, F.; Cabral-Piccin, M.P.; Papagno, L.; Gallerani, E.; Fusaro, M.; Folcher, V.; Dubois, M.; Clave, E.; Vallet, H.; Frere, J.J.; et al. Altered basal lipid metabolism underlies the functional impairment of naive CD8+ T cells in elderly humans. J. Immunol. 2022, 208, 562–570. [Google Scholar] [CrossRef]
- Nicoli, F.; Clave, E.; Wanke, K.; von Braun, A.; Bondet, V.; Alanio, C.; Douay, C.; Baque, M.; Lependu, C.; Marconi, P.; et al. Primary immune responses are negatively impacted by persistent herpesvirus infections in older people: Results from an observational study on healthy subjects and a vaccination trial on subjects aged more than 70 years old. EBioMedicine 2022, 76, 103852. [Google Scholar] [CrossRef]
- Briceno, O.; Lissina, A.; Wanke, K.; Afonso, G.; von Braun, A.; Ragon, K.; Miquel, T.; Gostick, E.; Papagno, L.; Stiasny, K.; et al. Reduced naive CD8+ T-cell priming efficacy in elderly adults. Aging Cell 2016, 15, 14–21. [Google Scholar] [CrossRef]
- Westmeier, J.; Paniskaki, K.; Karakose, Z.; Werner, T.; Sutter, K.; Dolff, S.; Overbeck, M.; Limmer, A.; Liu, J.; Zheng, X.; et al. Impaired cytotoxic CD8+ T cell response in elderly COVID-19 patients. mBio 2020, 11, e02243-20. [Google Scholar] [CrossRef]
- Pittet, M.J.; Valmori, D.; Dunbar, P.R.; Speiser, D.E.; Lienard, D.; Lejeune, F.; Fleischhauer, K.; Cerundolo, V.; Cerottini, J.C.; Romero, P. High frequencies of naive Melan-A/MART-1-specific CD8+ T cells in a large proportion of human histocompatibility leukocyte antigen (HLA)-A2 individuals. J. Exp. Med. 1999, 190, 705–715. [Google Scholar] [CrossRef] [Green Version]
- Gallerani, E.; Proietto, D.; Dallan, B.; Campagnaro, M.; Pacifico, S.; Albanese, V.; Marzola, E.; Marconi, P.; Caputo, A.; Appay, V.; et al. Impaired priming of SARS-CoV-2-specific naive CD8+ T cells in older subjects. Front. Immunol. 2021, 12, 693054. [Google Scholar] [CrossRef]
- Price, D.A.; Brenchley, J.M.; Ruff, L.E.; Betts, M.R.; Hill, B.J.; Roederer, M.; Koup, R.A.; Migueles, S.A.; Gostick, E.; Wooldridge, L.; et al. Avidity for antigen shapes clonal dominance in CD8+ T cell populations specific for persistent DNA viruses. J. Exp. Med. 2005, 202, 1349–1361. [Google Scholar] [CrossRef]
- Prakash, S.; Srivastava, R.; Coulon, P.G.; Dhanushkodi, N.R.; Chentoufi, A.A.; Tifrea, D.F.; Edwards, R.A.; Figueroa, C.J.; Schubl, S.D.; Hsieh, L.; et al. Genome-wide B cell, CD4+, and CD8+ T cell epitopes that are highly conserved between human and animal coronaviruses, identified from SARS-CoV-2 as targets for preemptive pan-coronavirus vaccines. J. Immunol. 2021, 206, 2566–2582. [Google Scholar] [CrossRef]
- Lee, E.; Sandgren, K.; Duette, G.; Stylianou, V.V.; Khanna, R.; Eden, J.S.; Blyth, E.; Gottlieb, D.; Cunningham, A.L.; Palmer, S. Identification of SARS-CoV-2 nucleocapsid and spike T-cell epitopes for assessing T-cell immunity. J. Virol. 2021, 95, e02002-20. [Google Scholar] [CrossRef]
- Gutjahr, A.; Papagno, L.; Nicoli, F.; Lamoureux, A.; Vernejoul, F.; Lioux, T.; Gostick, E.; Price, D.A.; Tiraby, G.; Perouzel, E.; et al. Cutting Edge: A dual TLR2 and TLR7 ligand induces highly potent humoral and cell-mediated immune responses. J. Immunol. 2017, 198, 4205–4209. [Google Scholar] [CrossRef] [Green Version]
- Fali, T.; Fabre-Mersseman, V.; Yamamoto, T.; Bayard, C.; Papagno, L.; Fastenackels, S.; Zoorab, R.; Koup, R.A.; Boddaert, J.; Sauce, D.; et al. Elderly human hematopoietic progenitor cells express cellular senescence markers and are more susceptible to pyroptosis. JCI Insight 2018, 3, e95319. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Weyand, C.M.; Goronzy, J.J.; Gustafson, C.E. Understanding T cell aging to improve anti-viral immunity. Curr. Opin. Virol. 2021, 51, 127–133. [Google Scholar] [CrossRef]
- Mathew, D.; Giles, J.R.; Baxter, A.E.; Oldridge, D.A.; Greenplate, A.R.; Wu, J.E.; Alanio, C.; Kuri-Cervantes, L.; Pampena, M.B.; D’Andrea, K.; et al. Deep immune profiling of COVID-19 patients reveals distinct immunotypes with therapeutic implications. Science 2020, 369, eabc8511. [Google Scholar] [CrossRef]
- Witkowski, J.M. Immune system aging and the aging-related diseases in the COVID-19 era. Immunol. Lett. 2022, 243, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Szeto, C.; Nguyen, A.T.; Lobos, C.A.; Chatzileontiadou, D.S.M.; Jayasinghe, D.; Grant, E.J.; Riboldi-Tunnicliffe, A.; Smith, C.; Gras, S. Molecular basis of a dominant SARS-CoV-2 spike-derived epitope presented by HLA-A*02:01 recognised by a public TCR. Cells 2021, 10, 2646. [Google Scholar] [CrossRef] [PubMed]
- Chaurasia, P.; Nguyen, T.H.O.; Rowntree, L.C.; Juno, J.A.; Wheatley, A.K.; Kent, S.J.; Kedzierska, K.; Rossjohn, J.; Petersen, J. Structural basis of biased T cell receptor recognition of an immunodominant HLA-A2 epitope of the SARS-CoV-2 spike protein. J. Biol. Chem. 2021, 297, 101065. [Google Scholar] [CrossRef] [PubMed]
- Qiu, C.; Xiao, C.; Wang, Z.; Zhu, G.; Mao, L.; Chen, X.; Gao, L.; Deng, J.; Su, J.; Su, H.; et al. CD8+ T-cell epitope variations suggest a potential antigen HLA-A2 binding deficiency for spike protein of SARS-CoV-2. Front. Immunol. 2022, 12, 764949. [Google Scholar] [CrossRef]
- Rowntree, L.C.; Petersen, J.; Juno, J.A.; Chaurasia, P.; Wragg, K.; Koutsakos, M.; Hensen, L.; Wheatley, A.K.; Kent, S.J.; Rossjohn, J.; et al. SARS-CoV-2-specific CD8+ T-cell responses and TCR signatures in the context of a prominent HLA-A*24:02 allomorph. Immunol. Cell Biol. 2021, 99, 990–1000. [Google Scholar] [CrossRef]
- Habel, J.R.; Nguyen, T.H.O.; van de Sandt, C.E.; Juno, J.A.; Chaurasia, P.; Wragg, K.; Koutsakos, M.; Hensen, L.; Jia, X.; Chua, B.; et al. Suboptimal SARS-CoV-2-specific CD8+ T cell response associated with the prominent HLA-A*02:01 phenotype. Proc. Natl. Acad. Sci. USA 2020, 117, 24384–24391. [Google Scholar] [CrossRef]
- Ranieri, E.; Popescu, I.; Gigante, M. CTL ELISPOT assay. Methods Mol. Biol. 2014, 1186, 75–86. [Google Scholar] [CrossRef]
- Chen, Y.; Klein, S.L.; Garibaldi, B.T.; Li, H.; Wu, C.; Osevala, N.M.; Li, T.; Margolick, J.B.; Pawelec, G.; Leng, S.X. Aging in COVID-19: Vulnerability, immunity and intervention. Ageing Res. Rev. 2021, 65, 101205. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Peptide ID | Sequence | Residues | Length | Tetramer |
|---|---|---|---|---|---|
| E | SLV | SLVKPSFYV | 50–58 | 9 | A*02:01 SV9 |
| M | KLL | KLLEQWNLV | 15–23 | 9 | A*02:01 KV9 |
| N | LLL | LLLDRLNQL | 222–230 | 9 | A*02:01 LL9 |
| RdRpol | MLD | MLDMYSVML | 5291–5299 | 9 | - |
| S | YLQ | YLQPRTFLL | 269–277 | 9 | A*02:01 YL9 |
| S | KIA | KIADYNYKL | 417–425 | 9 | - |
| S | TLD | TLDSKTQSL | 109–117 | 9 | - |
| ORF3a | LLY | LLYDANYFL | 139–147 | 9 | - |
| ORF6 | HLV | HLVDFQVTI | 3–11 | 9 | A*02:01 HI9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Proietto, D.; Dallan, B.; Gallerani, E.; Albanese, V.; Llewellyn-Lacey, S.; Price, D.A.; Appay, V.; Pacifico, S.; Caputo, A.; Nicoli, F.; et al. Ageing Curtails the Diversity and Functionality of Nascent CD8+ T Cell Responses against SARS-CoV-2. Vaccines 2023, 11, 154. https://doi.org/10.3390/vaccines11010154
Proietto D, Dallan B, Gallerani E, Albanese V, Llewellyn-Lacey S, Price DA, Appay V, Pacifico S, Caputo A, Nicoli F, et al. Ageing Curtails the Diversity and Functionality of Nascent CD8+ T Cell Responses against SARS-CoV-2. Vaccines. 2023; 11(1):154. https://doi.org/10.3390/vaccines11010154
Chicago/Turabian StyleProietto, Davide, Beatrice Dallan, Eleonora Gallerani, Valentina Albanese, Sian Llewellyn-Lacey, David A. Price, Victor Appay, Salvatore Pacifico, Antonella Caputo, Francesco Nicoli, and et al. 2023. "Ageing Curtails the Diversity and Functionality of Nascent CD8+ T Cell Responses against SARS-CoV-2" Vaccines 11, no. 1: 154. https://doi.org/10.3390/vaccines11010154