
Reactivation and Foetal Infection in Pregnant Heifers Infected with Neospora caninum Live Tachyzoites at Prepubertal Age

 , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Samplings

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group (n) | Treatment BP (1) | Challenged at 210 DG (2) with NC-1 | Evaluation of |
|---|---|---|---|
| A (4) | 1 × 106 live tachyzoites of NC-Argentina LP1 strain | Yes | Protection |
| B (4) | PBS | Yes | Challenge control |
| C (4) | 1 × 106 live tachyzoites of NC-Argentina LP1 strain | No | Reactivation |
| D (3) | PBS | No | Negative control |
2.2. Neospora Caninum Inoculum and Native Antigen Extract
2.3. Specific IgG and Its Subisotypes in Serum and Colostrum Samples
2.4. Indirect Immunofluorescence Antibody Test (IFAT)
2.5. Immunoblot (IB) Analysis
2.6. Nested PCR (nPCR) for Neospora Caninum
2.7. Histopathological Studies on Placentas
2.8. Cytokine mRNA Expression in Dam PBMCs
2.9. Statistical Analysis
3. Results
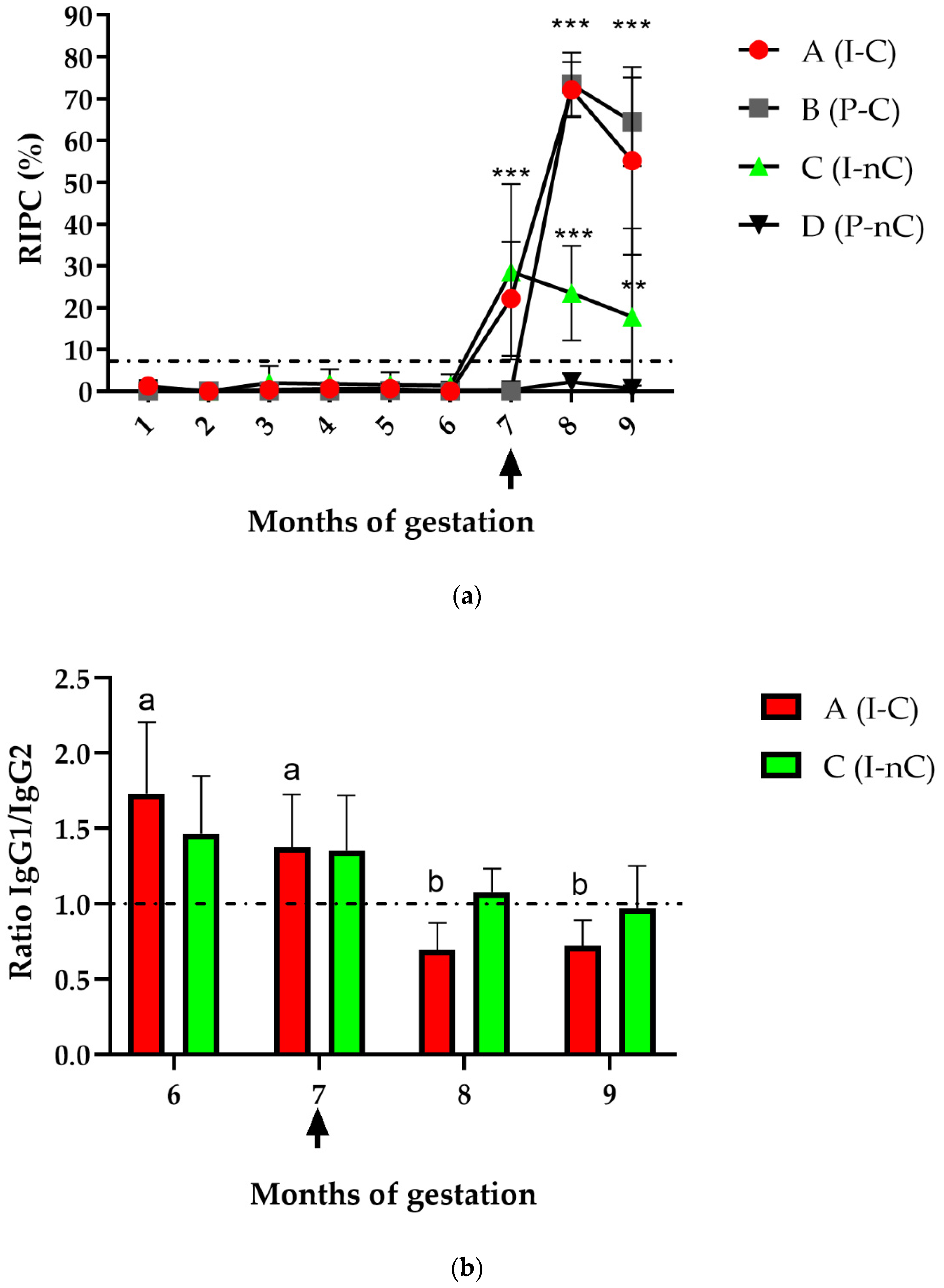
3.1. Serum Antibody Response in Dams
3.1.1. Specific Total IgG
3.1.2. IgG Subisotypes
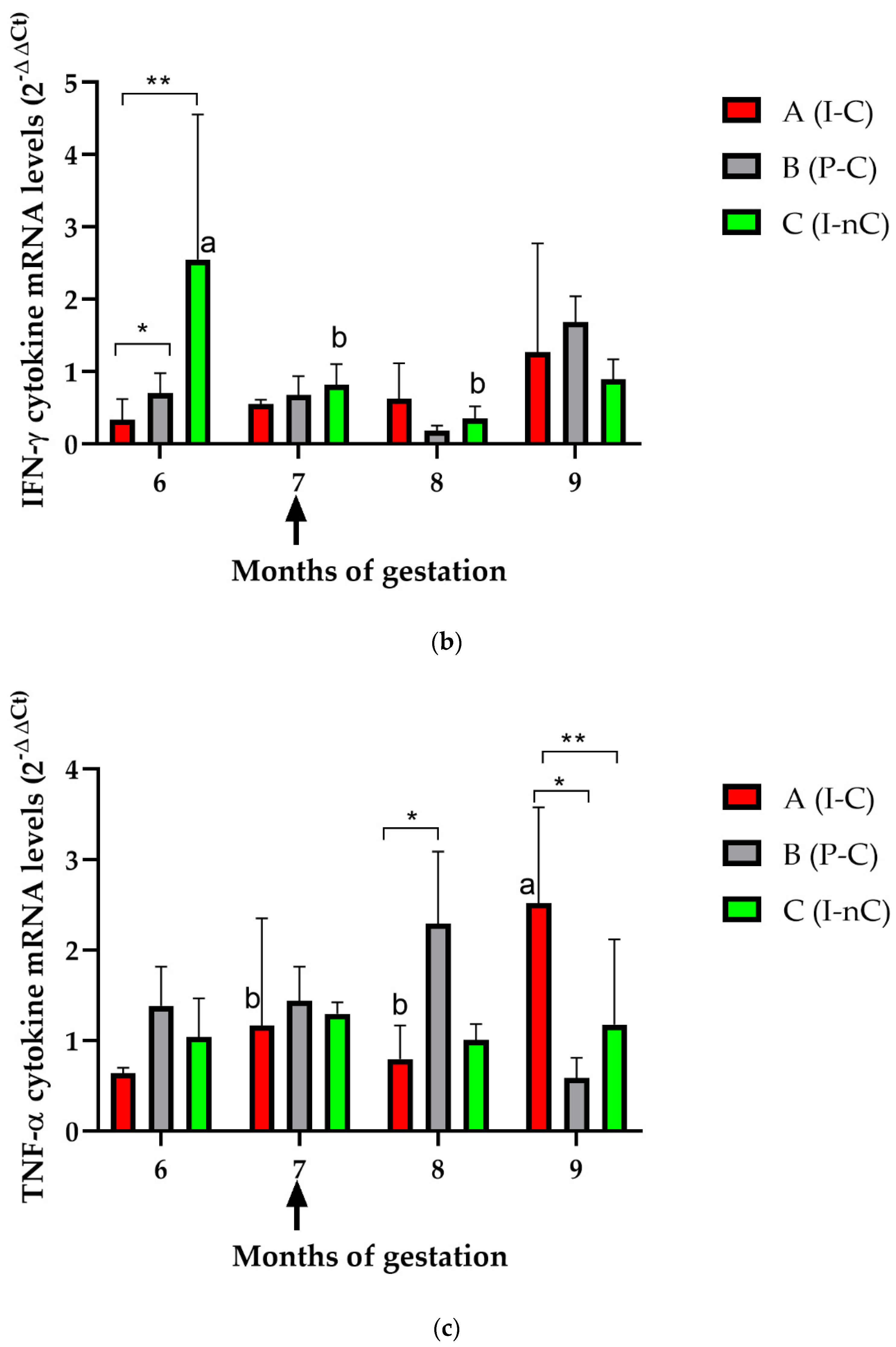
3.2. Cellular Immune Response in Dams
3.3. Neospora DNA Detection in Dams during Gestation and at Delivery
3.4. Histopathological Studies on Placentas and Aborted Foetuses
3.5. Detection of N. Caninum Infection in Calves
| Groups | Months of Gestation | 6 | 7 | 8 | 9 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Heifers | ELISA 1 | IFAT 2 | IB 3 | PCR 4 | ELISA 1 | IFAT 2 | IB 3 | PCR 4 | ELISA 1 | IFAT 2 | IB 3 | PCR 4 | ELISA 1 | IFAT 2 | IB 3 | PCR 4 | |
| A | #4 | − | 25 a | + | − | + | 100 a | + | + | + | 12,800 a | + | − | + | 6400 b | + | + |
| (I-C) | #5 | − | 100 a | + | − | + | 800 a | + | + | + | 25,600 a | + | − | + | 6400 b | + | + |
| #9 | − | 25 a | − | − | − | 100 a | − | + | + | 25,600 a | + | + | + | 12,800 b | + | − | |
| #12 | − | 100 a | + | + | + | 1600 a | + | + | + | 12,800 a | + | + | + | 800 b | + | + | |
| B | #32 | − | − b | − | − | − | − b | − | − | + | 12,800 a | + | − | + | 51,200 a | + | − |
| (P-C) | #34 | − | − b | − | − | − | − b | − | − | + | 12,800 a | + | − | + | 25,600 a | + | − |
| #38 | − | − b | − | − | − | − b | − | − | + | 25,600 a | + | − | + | 6400 a | + | − | |
| #47 | − | − b | − | − | − | − b | − | − | + | 25,600 a | + | − | + | 6400 a | + | − | |
| C | #2 | − | 100 a | + | − | + | 400 a | + | + | + | 800 b | + | + | + | 1600 b | − | − |
| (I-nC) | #14 | − | 25 a | − | − | + | 400 a | + | + | + | 200 b | + | − | + | 200 b | + | − |
| #18 | − | 25 a | + | − | − | 100 a | + | + | + | 3200 b | + | − | + | 3200 b | − | − | |
| #19 | − | 50 a | + | − | + | 400 a | + | + | + | 800 b | + | + | + | 3200 b | + | − | |
| D | #30 | − | − b | − | − | − | − b | − | − | − | − c | − | − | − | − c | − | − |
| (P-nC) | #47a | − | − b | − | − | − | − b | − | − | − | − c | − | − | − | − c | − | − |
| #48 | − | − b | − | − | − | − b | − | − | − | − c | − | − | − | − c | − | − | |
| Experimental Groups | Dams | Calves | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| ID | Specific IgG 1 | N. caninum DNA | ID | Specific IgG 1 | N. caninum DNA | |||||
| Serum 2 | Colostrum 1 | Placenta | PBMC | Colostrum | Serum 3 | Umbilical Cord | PBMC | |||
| A (I-C) | #4 | + | + | − | + | − | #1 | + | − | − |
| #5 | + | + | + | + | − | #6 | + | + | − | |
| #9 | + | + | − | − | − | #9 | + | − | − | |
| #12 | + | − | − | + | − | Aborted foetus | − | − | − | |
| B (P-C) | #32 | + | + | − | − | − | #3 | + | − | − |
| #34 | + | − | − | − | − | #4 | + | − | − | |
| #38 | + | − | − | − | − | #14 | + | − | − | |
| #47 | + | − | − | − | − | #13 | + | − | − | |
| C (I-nC) | #2 | + | − | − | − | − | #11 | − | − | − |
| #14 | + | − | − | − | − | #12 | + | − | − | |
| #18 | + | + | − | − | − | #5 | + | − | − | |
| #19 | + | + | − | − | − | #10 | − | − | − | |
| D (P-nC) | #30 | − | − | − | − | − | #8 | − | − | − |
| #47a | − | − | − | − | − | #15 | − | − | − | |
| #48 | − | − | − | − | − | #2 | − | − | − | |
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lindsay, D.S.; Dubey, J.P. Neosporosis, toxoplasmosis, and sarcocystosis in ruminants: An update. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 205–222. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.; Hemphill, A.; Calero-Bernal, R.; Schares, G. Neosporosis in Animals, 1st ed.; CRC Press: Boca Raton, FL, USA, 2017. [Google Scholar] [CrossRef]
- Reichel, M.P.; Ayanegui-Alcérreca, M.A.; Gondim, L.F.P.; Ellis, J.T. What is the global economic impact of Neospora caninum in cattle—The billion dollar question. Int. J. Parasitol. 2013, 43, 133–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horcajo, P.; Regidor-Cerrillo, J.; Aguado-Martínez, A.; Hemphill, A.; Ortega-Mora, L.M. Vaccines for bovine neosporosis: Current status and key aspects for development. Parasite Immunol. 2016, 38, 709–723. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Staska, L.M.; McGuire, T.C.; Davies, C.J.; Lewin, H.A.; Baszler, T.V. Neospora caninum-infected cattle develop parasite-specific CD4+ cytotoxic T lymphocytes. Infect. Immun. 2003, 71, 3272–3279. [Google Scholar] [CrossRef] [Green Version]
- Almería, S.; Nogareda, C.; Santolaria, P.; Garcia-Ispierto, I.; Yániz, J.L.; López-Gatius, F. Specific anti-Neospora caninum IgG1 and IgG2 antibody responses during gestation in naturally infected cattle and their relationship with gamma interferon production. Vet. Immunol. Immunopathol. 2009, 130, 35–42. [Google Scholar] [CrossRef]
- Almería, S.; Serrano-Pérez, B.; López-Gatius, F. Immune response in bovine neosporosis: Protection or contribution to the pathogenesis of abortion. Microb. Pathog. 2017, 109, 177–182. [Google Scholar] [CrossRef]
- McAllister, M.M.; Björkman, C.; Anderson-Sprecher, R.; Rogers, D.G. Evidence of point-source exposure to Neospora caninum and protective immunity in a herd of beef cows. J. Am. Vet. Med. Assoc. 2000, 217, 881–887. [Google Scholar] [CrossRef]
- Innes, E.A.; Wright, S.E.; Maley, S.; Rae, A.; Schock, A.; Kirvar, E.; Bartley, P.; Hamilton, C.; Carey, I.M.; Buxton, D. Protection against vertical transmission in bovine neosporosis. Int. J. Parasitol. 2001, 31, 1523–1534. [Google Scholar] [CrossRef]
- Campero, L.M.; Venturini, M.C.; Moore, D.P.; Massola, L.; Lagomarsino, H.; García, B.; Bacigalupe, D.; Rambeaud, M.; Pardini, L.; Leunda, M.R.; et al. Isolation and molecular characterization of a new Neospora caninum isolate from cattle in Argentina. Exp. Parasitol. 2015, 155, 8–12. [Google Scholar] [CrossRef]
- Hecker, Y.P.; Regidor-Cerrillo, J.; Fiorani, F.; Horcajo, P.; Soria, I.; Gual, I.; Torioni, S.; Campero, L.M.; Echaide, I.E.; Álvarez-García, G.; et al. Immune response to Neospora caninum live tachyzoites in prepubertal female calves. Parasitol. Res. 2019, 118, 2945–2955. [Google Scholar] [CrossRef]
- Paré, J.; Thurmond, M.C.; Hietala, S.K. Congenital Neospora caninum infection in dairy cattle and associated calfhood mortality. Can. J. Vet. Res. 1996, 60, 133–139. [Google Scholar] [PubMed]
- Dubey, J.P.; Hattel, A.L.; Lindsay, D.S.; Topper, M.J. Neonatal Neospora caninum infection in dogs: Isolation of the causative agent and experimental transmission. J. Am. Vet. Med. Assoc. 1988, 193, 1259–1263. [Google Scholar]
- Hecker, Y.P.; Moore, D.P.; Quattrocchi, V.; Regidor-Cerrillo, J.; Verna, A.; Leunda, M.R.; Morrell, E.; Ortega-Mora, L.M.; Zamorano, P.; Venturini, M.C.; et al. Immune response and protection provided by live tachyzoites and native antigens from the NC-6 Argentina strain of Neospora caninum in pregnant heifers. Vet. Parasitol. 2013, 197, 436–446. [Google Scholar] [CrossRef] [PubMed]
- Rivera, J.E.M.; Hecker, Y.P.; Burucúa, M.M.; Cirone, K.M.; Cheuquepán, F.A.; Fiorani, F.; Dorsch, M.A.; Colque, L.A.; Cantón, G.J.; Marin, M.S.; et al. Innate and humoral immune parameters at delivery in colostrum and calves from heifers experimentally infected with Neospora caninum. Mol. Immunol. 2021, 132, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Caspe, S.G.; Moore, D.P.; Leunda, M.R.; Cano, D.B.; Lischinsky, L.; Regidor-Cerrillo, J.; Álvarez-García, G.; Echaide, I.G.; Bacigalupe, D.; Ortega-Mora, L.M.; et al. The Neospora caninum-Spain 7 isolate induces placental damage, fetal death and abortion in cattle when inoculated in early gestation. Vet. Parasitol. 2012, 189, 171–181. [Google Scholar] [CrossRef]
- Campero, L.M.; Minkec, L.; Moré, G.; Rambeaud, M.; Bacigalupe, D.; Moore, D.P.; Hecker, Y.; Campero, C.M.; Schares, G.; Venturini, M.C. Evaluation and comparison of serological methods for the detection of bovine neosporosis in Argentina. Rev. Argent. Microbiol. 2015, 47, 295–301. [Google Scholar] [CrossRef] [Green Version]
- Buxton, D.; Maley, S.W.; Wright, S.; Thomson, K.M.; Rae, A.G.; Innes, E.A. The pathogenesis of experimental neosporosis in pregnant sheep. J. Comp. Pathol. 1998, 118, 267–279. [Google Scholar] [CrossRef]
- Campero, C.M.; Moore, D.P.; Odeón, A.C.; Cipolla, A.L.; Odriozola, E. Aetiology of bovine abortion in Argentina. Vet. Res. Commun. 2003, 27, 359–369. [Google Scholar] [CrossRef]
- Regidor-Cerrillo, J.; Arranz-Solís, D.; Benavides, J.; Gómez-Bautista, M.; Castro-Hermida, J.A.; Mezo, M.; Pérez, V.; Ortega-Mora, L.M.; González-Warleta, M. Neospora caninum infection during early pregnancy in cattle: How the isolate influences infection dynamics, clinical outcome and peripheral and local immune responses. Vet. Res. 2014, 45, 10. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Benavides, J.; Collantes-Fernández, E.; Ferre, I.; Pérez, V.; Campero, C.; Mota, R.; Innes, E.; Ortega-Mora, L.M. Experimental ruminant models for bovine neosporosis: What is known and what is needed. Parasitology 2014, 141, 1471–1488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Williams, D.J.; Guy, C.S.; Smith, R.F.; Guy, F.; McGarry, J.W.; McKay, J.S.; Trees, A.J. First demonstration of protective immunity against foetopathy in cattle with latent Neospora caninum infection. Int. J. Parasitol. 2003, 33, 1059–1065. [Google Scholar] [CrossRef]
- De Marez, T.; Liddell, S.; Dubey, J.P.; Jenkins, M.C.; Gasbarre, L. Oral infection of calves with Neospora caninum oocysts from dogs: Humoral and cellular immune responses. Int. J. Parasitol. 1999, 29, 1647–1657. [Google Scholar] [CrossRef]
- Maley, S.W.; Buxton, D.; Thomson, K.M.; Schriefer, C.E.S.; Innes, E.A. Serological analysis of calves experimentally infected with Neospora caninum: A 1-year study. Vet. Parasitol. 2001, 96, 1–9. [Google Scholar] [CrossRef]
- Klevar, S.; Kulberg, S.; Boysen, P.; Storset, A.K.; Moldal, T.; Björkman, C.; Olsen, I. Natural killer cells act as early responders in an experimental infection with Neospora caninum in calves. Int. J. Parasitol. 2007, 37, 329–339. [Google Scholar] [CrossRef]
- Quintanilla-Gozalo, A.; Pereira-Bueno, J.; Tabarés, E.; Innes, E.A.; González-Paniello, R.; Ortega-Mora, L.M. Seroprevalence of Neospora caninum infection in dairy and beef cattle in Spain. Int. J. Parasitol. 1999, 29, 1201–1208. [Google Scholar] [CrossRef]
- Maley, S.W.; Buxton, D.; Rae, A.G.; Wright, S.E.; Schock, A.; Bartley, P.M.; Esteban-Redondo, I.; Swales, C.; Hamilton, C.M.; Sales, J.; et al. The pathogenesis of neosporosis in pregnant cattle: Inoculation at mid-gestation. J. Comp. Pathol. 2003, 129, 186–195. [Google Scholar] [CrossRef]
- Williams, D.J.; Guy, C.S.; McGarry, J.W.; Guy, F.; Tasker, L.; Smith, R.F.; MacEachern, K.; Cripps, P.J.; Kelly, D.F.; Trees, A.J. Neospora caninum-associated abortion in cattle: The time of experimentally-induced parasitaemia during gestation determines foetal survival. Parasitology 2000, 121 Pt 4, 347–358. [Google Scholar] [CrossRef]
- Guy, C.S.; Williams, D.J.L.; Kelly, D.F.; McGarry, J.W.; Guy, F.; Björkman, C.; Smith, R.F.; Trees, A.J. Neospora caninum in persistently infected, pregnant cows: Spontaneous transplacental infection is associated with an acute increase in maternal antibody. Vet. Rec. 2001, 149, 443–449. [Google Scholar] [CrossRef]
- Nogareda, C.; López-Gatius, F.; Santolaria, P.; García-Ispierto, I.; Bech-Sàbat, G.; Pabón, M.; Mezo, M.; Gonzalez-Warleta, M.; Castro-Hermida, J.A.; Yániz, J.; et al. Dynamics of anti-Neospora caninum antibodies during gestation in chronically infected dairy cows. Vet. Parasitol. 2007, 148, 193–199. [Google Scholar] [CrossRef]
- Williams, D.J.; Guy, C.S.; Smith, R.F.; Ellis, J.; Björkman, C.; Reichel, M.P.; Trees, A.J. Immunization of cattle with live tachyzoites of Neospora caninum confers protection against fetal death. Infect. Immun. 2007, 75, 1343–1348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojo-Montejo, S.; Collantes-Fernández, E.; Blanco-Murcia, J.; Rodríguez-Bertos, A.; Risco-Castillo, V.; Ortega-Mora, L. Experimental infection with a low virulence isolate of Neospora caninum at 70 days gestation in cattle did not result in foetopathy. Vet. Res. 2009, 40, 49. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rojo-Montejo, S.; Collantes-Fernandez, E.; Perez-Zaballos, F.; Rodriguez-Marcos, S.; Blanco-Murcia, J.; Rodriguez-Bertos, A.; Prenafeta, A.; Ortega-Mora, L.M. Effect of vaccination of cattle with the low virulence Nc-Spain 1H isolate of Neospora caninum against a heterologous challenge in early and mid-gestation. Vet. Res. 2013, 44, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, F.H.; Jackson, J.A.; Sobecki, B.; Choromansky, L.; Olsen, M.; Meinert, T.; Frank, R.; Reichel, M.P.; Ellis, J.T. On the efficacy and safety of vaccination with live tachyzoites of N. caninum for prevention of Neospora-associated fetal loss in cattle. Clin. Vaccine Immunol. 2013, 20, 99–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mazuz, M.L.; Fisha, L.; Wolkomirsky, R.; Leibovich, B.; Reznikov, D.; Savitsky, I.; Golenser, J.; Shkap, V. The effect of a live Neospora caninum tachyzoite vaccine in naturally infected pregnant dairy cows. Prev. Med. 2015, 120, 232–235. [Google Scholar] [CrossRef]
- Mazuz, M.L.; Leibovich, B.; Savitsky, I.; Blinder, E.; Yasur-Landau, D.; Lavon, Y.; Sharir, B.; Tirosh-Levy, S. The effect of vaccination with Neospora caninum live-frozen tachyzoites on abortion rates of naturally infected pregnant cows. Vaccines 2021, 9, 401. [Google Scholar] [CrossRef]
- Regidor-Cerrillo, J.; Gómez-Bautista, M.; Pereira-Bueno, J.; Aduriz, G.; Navarro-Lozano, V.; Risco-Castillo, V.; Férnandez-García, A.; Pedraza-Díaz, S.; Ortega-Mora, L.M. Isolation and genetic characterization of Neospora caninum from asymptomatic calves in Spain. Parasitology 2008, 135, 1651–1659. [Google Scholar] [CrossRef]
- Benavides, J.; Katzer, F.; Maley, S.W.; Bartley, P.M.; Canton, G.; Palarea-Albaladejo, J.; Purslow, C.A.; Pang, Y.; Rocchi, M.S.; Chianini, F.; et al. High rate of transplacental infection and transmission of Neospora caninum following experimental challenge of cattle at day 210 of gestation. Vet. Res. 2012, 43, 83. [Google Scholar] [CrossRef] [Green Version]
- Rosbottom, A.; Guy, C.S.; Gibney, E.H.; Smith, R.F.; Valarcher, J.F.; Taylor, G.; Williams, D.J.L. Peripheral immune responses in pregnant cattle following Neospora caninum infection. Parasite Immunol. 2007, 29, 219–228. [Google Scholar] [CrossRef]
- Rosbottom, A.; Gibney, H.; Kaiser, P.; Hartley, C.; Smith, R.F.; Robinson, R.; Kipar, A.; Williams, D.J. Up regulation of the maternal immune response in the placenta of cattle naturally infected with Neospora caninum. PLoS ONE 2011, 6, e15799. [Google Scholar] [CrossRef] [Green Version]
- Gibney, E.H.; Kipar, A.; Rosbottom, A.; Guy, C.S.; Smith, R.F.; Hetzel, U.; Trees, A.J.; Williams, D.J. The extent of parasite-associated necrosis in the placenta and foetal tissues of cattle following Neospora caninum infection in early and late gestation correlates with foetal death. Int. J. Parasitol. 2008, 38, 579–588. [Google Scholar] [CrossRef] [PubMed]
- Bartley, P.M.; Katzer, F.; Rocchi, M.S.; Maley, S.W.; Benavides, J.; Nath, M.; Pang, Y.; Canton, G.; Thomson, J.; Chianini, F.; et al. Development of maternal and foetal immune responses in cattle following experimental challenge with Neospora caninum at day 210 of gestation. Vet. Res. 2013, 44, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartley, P.M.; Wright, S.; Chianini, F.; Buxton, D.; Innes, E.A. Inoculation of Balb/c mice with live attenuated tachyzoites protects against a lethal challenge of Neospora caninum. Parasitology 2008, 135, 13–21. [Google Scholar] [CrossRef] [PubMed]
- Aminia, L.; Namavarib, M.; Khodakaram-Taftia, A.; Divarc, M.R.; Hosseinib, S.M.H. The evaluation of attenuated Neospora caninum by long-term passages on murine macrophage cell line in prevention of vertical transmission in mice. Vet. Parasitol. 2020, 283, 109171. [Google Scholar] [CrossRef]
- Bancherau, J.; Pascual, V.; O’Garra, A. From IL-2 to IL-37: The expanding spectrum of anti-inflammatory cytokines. Nat. Immunol. 2012, 13, 925–931. [Google Scholar] [CrossRef] [PubMed]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hecker, Y.P.; Burucúa, M.M.; Fiorani, F.; Maldonado Rivera, J.E.; Cirone, K.M.; Dorsch, M.A.; Cheuquepán, F.A.; Campero, L.M.; Cantón, G.J.; Marín, M.S.; et al. Reactivation and Foetal Infection in Pregnant Heifers Infected with Neospora caninum Live Tachyzoites at Prepubertal Age. Vaccines 2022, 10, 1175. https://doi.org/10.3390/vaccines10081175
Hecker YP, Burucúa MM, Fiorani F, Maldonado Rivera JE, Cirone KM, Dorsch MA, Cheuquepán FA, Campero LM, Cantón GJ, Marín MS, et al. Reactivation and Foetal Infection in Pregnant Heifers Infected with Neospora caninum Live Tachyzoites at Prepubertal Age. Vaccines. 2022; 10(8):1175. https://doi.org/10.3390/vaccines10081175
Chicago/Turabian StyleHecker, Yanina P., Mercedes M. Burucúa, Franco Fiorani, Jaime E. Maldonado Rivera, Karina M. Cirone, Matías A. Dorsch, Felipe A. Cheuquepán, Lucía M. Campero, Germán J. Cantón, Maia S. Marín, and et al. 2022. "Reactivation and Foetal Infection in Pregnant Heifers Infected with Neospora caninum Live Tachyzoites at Prepubertal Age" Vaccines 10, no. 8: 1175. https://doi.org/10.3390/vaccines10081175