
Adults with Trisomy 21 Have Differential Antibody Responses to Influenza A


Abstract
:1. Introduction
2. Materials and Methods
2.1. Antibody Concentration Measurements
2.2. Hemagglutination Inhibition Assays (HIA)
2.3. Antibody Isotyping
2.4. Data Analysis and Statistics
3. Results
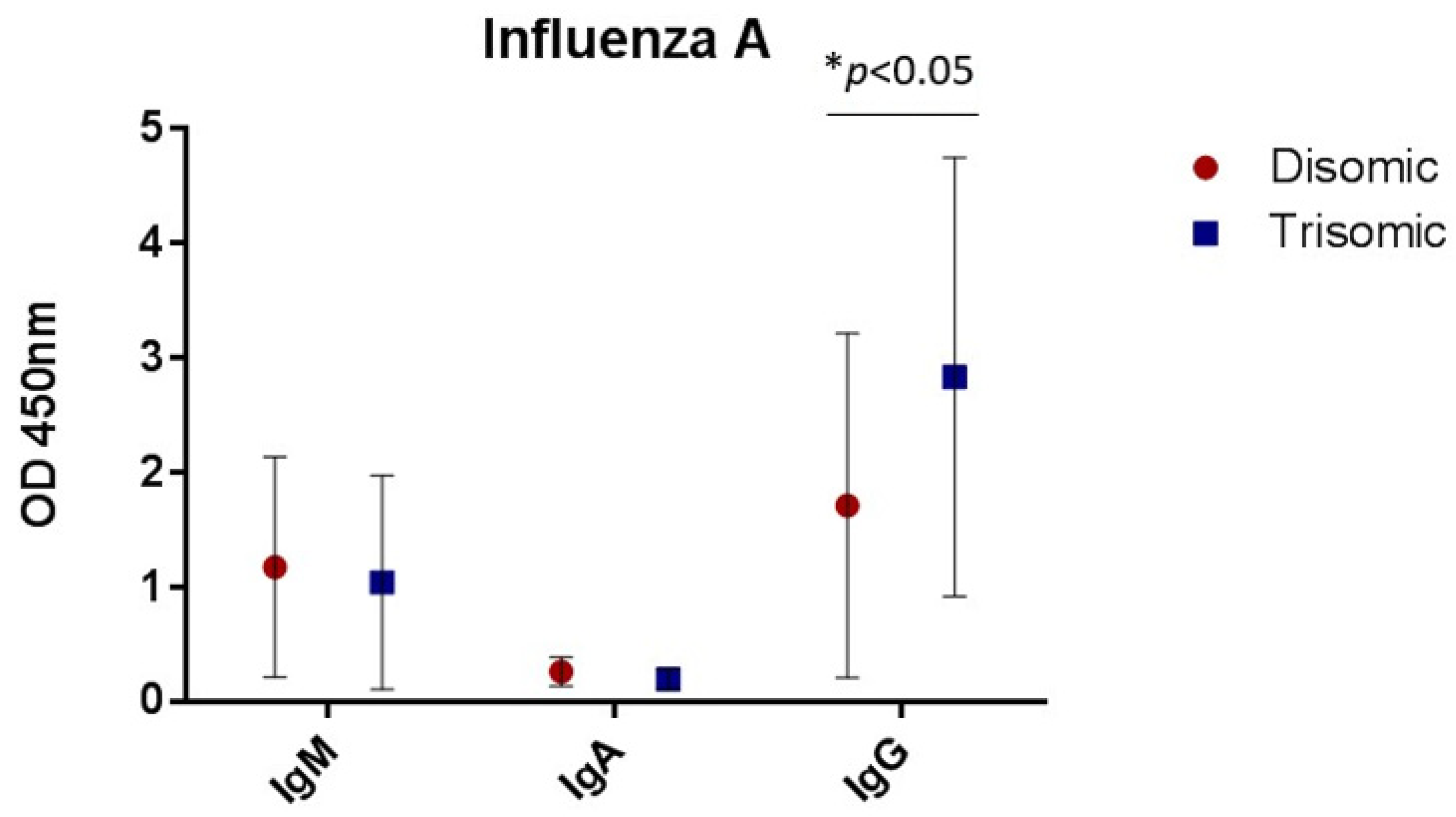
3.1. Antibody Concentrations
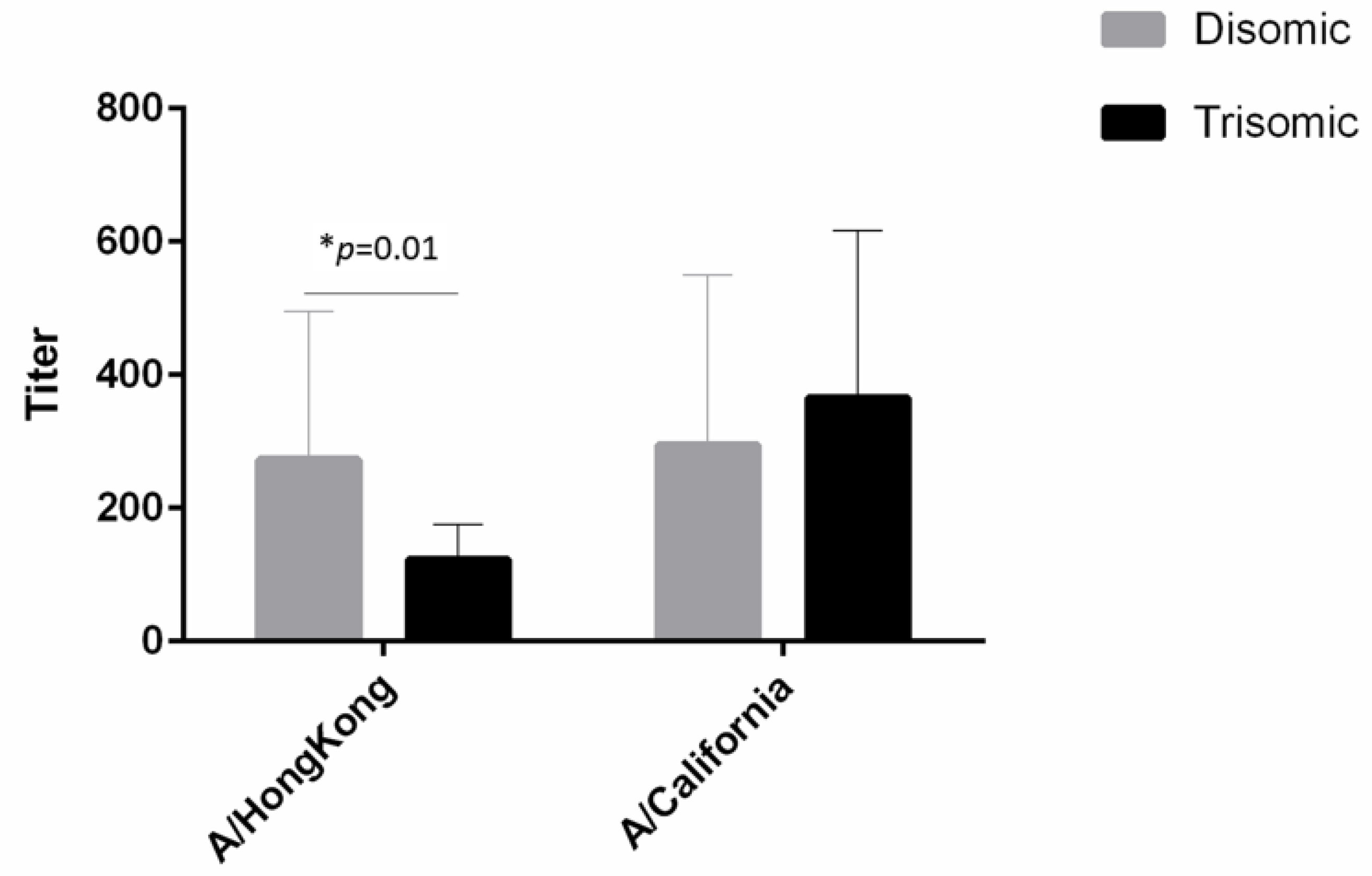
3.2. Hemagglutination Inhibition Assays
3.3. Antibody Isotyping
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Forrester, M.B.; Merz, R.D. Epidemiology of Down syndrome (Trisomy 21), Hawaii, 1986–1997. Teratology 2002, 65, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Presson, A.P.; Partyka, G.; Jensen, K.M.; Devine, O.J.; Rasmussen, S.A.; McCabe, L.L.; McCabe, E.R. Current Estimate of Down Syndrome Population Prevalence in the United States. J. Pediatr. 2013, 163, 1163–1168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, J.L.; Hasle, H.; Correa, A.; Schendel, D.; Friedman, J.; Olsen, J.; Rasmussen, S.A. Survival among people with Down syndrome: A nationwide population-based study in Denmark. Genet. Med. 2013, 15, 64–69. [Google Scholar] [CrossRef] [Green Version]
- Tenenbaum, A.; Chavkin, M.; Wexler, I.D.; Korem, M.; Merrick, J. Morbidity and hospitalizations of adults with Down syndrome. Res. Dev. Disabil. 2012, 33, 435–441. [Google Scholar] [CrossRef]
- Pérez-Padilla, R.; Fernández, R.; García-Sancho, C.; Franco-Marina, F.; Aburto, O.; Lopez-Gatell, H.; Bojorquez, I. Pandemic (H1N1) 2009 Virus and Down Syndrome Patients. Emerg. Infect. Dis. 2010, 16, 1312–1314. [Google Scholar] [CrossRef]
- Philip, R. Abnormalities of the in vitro cellular and humoreal responses to tetanus and influenza antigens with concomitant numberical alterations in lymphocyte subsets in Down syndrome (trisomy 21). J. Immunol. 1986, 136, 1661–1667. [Google Scholar] [PubMed]
- Epstein, L.B.; Philip, R. Abnormalities of the immune response to influenza antigen in Down syndrome (trisomy 21). Prog. Clin. Biol. Res. 1987, 246, 163–182. [Google Scholar]
- Nurmi, T.; Leinonen, M.; Haiva, V.M.; Tiilikainen, A.; Kouvalainen, K. Antibody response to pneumococcal vaccine in patients with trisomy-21 (Down’s syndrome). Clin. Exp. Immunol. 1982, 48, 485–490. [Google Scholar]
- Valentini, D.; Marcellini, V.; Bianchi, S.; Villani, A.; Facchini, M.; Donatelli, I.; Castrucci, M.R.; Marasco, E.; Farroni, C.; Carsetti, R. Generation of switched memory B cells in response to vaccination in Down syndrome children and their siblings. Vaccine 2015, 33, 6689–6696. [Google Scholar] [CrossRef] [Green Version]
- Eijsvoogel, N.B.; Hollegien, M.I.; Bok, V.L.A.; Derksen Lubsen, A.G.; Dikken, F.P.J.; Leenders, S.; Pijning, A.; Post, E.; Wojciechowski, M.; Hilbink, M.; et al. Declining antibody levels after hepatitis B vaccination in Down syndrome: A need for booster vaccination? J. Med. Virol. 2017, 89, 1682–1689. [Google Scholar] [CrossRef]
- A Baird, P.; Sadovnick, A.D. Causes of death to age 30 in Down syndrome. Am. J. Hum. Genet. 1988, 43, 239–248. [Google Scholar] [PubMed]
- Yang, Q.; Rasmussen, S.A.; Friedman, J.M. Mortality associated with Down’s syndrome in the USA from 1983 to 1997: A population-based study. Lancet 2002, 359, 1019–1025. [Google Scholar] [CrossRef]
- Scholl, T.; Stein, Z.; Hansen, H. Leukemia and other cancers, anomalies and infections as causes of death in Down’s syndrome in the United States during 1976. Dev. Med. Child Neurol. 1982, 24, 817–829. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.A.; Gridley, G.; Cnattingius, S.; Mellemkjaer, L.; Linet, M.; Adami, H.-O.; Olsen, J.H.; Nyren, O.; Fraumeni, J.F. Mortality and Cancer Incidence Among Individuals With Down Syndrome. Arch. Intern. Med. 2003, 163, 705–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iuliano, A.D.; Roguski, K.M.; Chang, H.H.; Muscatello, D.J.; Palekar, R.; Tempia, S.; Cohen, C.; Gran, J.M.; Schanzer, D.; Cowling, B.J.; et al. Estimates of global seasonal influenza-associated respiratory mortality: A modelling study. Lancet 2018, 391, 1285–1300. [Google Scholar] [CrossRef]
- Martin-Loeches, I.; van Someren Greve, F.; Schultz, M.J. Bacterial pneumonia as an influenza complication. Curr. Opin. Infect. Dis. 2017, 30, 201–207. [Google Scholar] [CrossRef]
- Ballinger, M.N.; Standiford, T.J. Postinfluenza Bacterial Pneumonia: Host Defenses Gone Awry. J. Interf. Cytokine Res. 2010, 30, 643–652. [Google Scholar] [CrossRef] [Green Version]
- Robinson, K.M.; Kolls, J.K.; Alcorn, J.F. The immunology of influenza virus-associated bacterial pneumonia. Curr. Opin. Immunol. 2015, 34, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Blanton, L.; Alabi, N.; Mustaquim, D.; Taylor, C.; Kniss, K.; Kramer, N.; Budd, A.; Garg, S.; Cummings, C.N.; Chung, J.; et al. Update: Influenza Activity in the United States During the 2016–17 Season and Composition of the 2017–18 Influenza Vaccine. Morb. Mortal. Wkly. Rep. 2017, 66, 668–676. [Google Scholar] [CrossRef]
- Verstegen, R.H.; Driessen, G.J.; Bartol, S.J.; van Noesel, C.J.; Boon, L.; van der Burg, M.; van Dongen, J.J.; de Vries, E.; van Zelm, M.C. Defective B-cell memory in patients with Down syndrome. J. Allergy Clin. Immunol. 2014, 134, 1346–1353. [Google Scholar] [CrossRef]
- Carsetti, R.; Valentini, D.; Marcellini, V.; Scarsella, M.; Marasco, E.; Giustini, F.; Bartuli, A.; Villani, A.; Ugazio, A.G. Reduced numbers of switched memory B cells with high terminal differentiation potential in Down syndrome. Eur. J. Immunol. 2014, 45, 903–914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nespoli, L. Immunological features of Down’s syndrome: A review. J. Intellect. Disabil. Res. 1993, 37, 543–551. [Google Scholar] [CrossRef] [PubMed]
- Barrena, M.J.; Echaniz, P.; Garcia-Serrano, C.; Cuadrado, E. Imbalance of the CD4+ Subpopulations Expressing CD45RA and CD29 Antigens in the Peripheral Blood of Adults and Children with Down Syndrome. Scand. J. Immunol. 1993, 38, 323–326. [Google Scholar] [CrossRef] [PubMed]
- Murphy, M.; Epstein, L.B. Down syndrome (DS) peripheral blood contains phenotypically mature CD3+TCR alpha, beta+ cells but abnormal proportions of TCR alpha, beta+, TCR gamma, delta+, and CD4+ CD45RA+ cells: Evidence for an inefficient release of mature T cells by the DS thymus. Clin. Immunol. Immunopathol. 1992, 62, 245–251. [Google Scholar] [CrossRef]
- Ram, G.; Chinen, J. Infections and immunodeficiency in Down syndrome. Clin. Exp. Immunology. 2011, 164, 9–16. [Google Scholar] [CrossRef]
- Rigas, D.A.; Elsasser, P.; Hecht, F. Impaired in vitro Response of Circulating Lymphocytes to Phytohemagglutinin in Down’s Syndrome: Dose- and Time-Response Curves and Relation to Cellular Immunity. Int. Arch. Allergy Immunol. 1970, 39, 587–608. [Google Scholar] [CrossRef]
- Sullivan, K.D.; Lewis, H.C.; A Hill, A.; Pandey, A.; Jackson, L.P.; Cabral, J.M.; Smith, K.P.; Liggett, L.A.; Gomez, E.B.; Galbraith, M.D.; et al. Author response: Trisomy 21 consistently activates the interferon response. eLife 2016, 5, e16220. [Google Scholar] [CrossRef]
- Huggard, D.; Kelly, L.; Ryan, E.; McGrane, F.; Lagan, N.; Roche, E.; Balfe, J.; Leahy, T.; Franklin, O.; Doherty, D. Increased systemic inflammation in children with Down syndrome. Cytokine 2020, 127, 154938. [Google Scholar] [CrossRef]
- Peters, M.; Ambrus, J.L.; Zheleznyak, A.; Walling, D.; Hoofnagle, J.H. Effect of interferon-alpha on immunoglobulin synthesis by human B cells. J. Immunol. 1986, 137, 3153–3157. [Google Scholar]



{kind=link}
{kind=link}
| Disomic Individuals | Trisomic Individuals | |
|---|---|---|
| N | 17 | 18 |
| % Female | 76.47 | 61.11 |
| % Male | 23.53 | 38.89 |
| Age [mean(sd)] | 37.32 (13.23) | 30.91 (10.36) |
| IgM (1) | IgA (2) | IgG (3) | |
|---|---|---|---|
| Trisomy 21 | −0.20 (0.31) | −017 (0.14) | 1.21 * (0.58) |
| Female | −0.29 (0.34) | −0.14 (0.15) | −0.43 (0.63) |
| Constant | 1.42 *** (0.34) | 0.55 *** (0.15) | 1.89 ** (0.64) |
| Observations | 36 | 36 | 37 |
| Adjusted R-squared | −0.03 | 0.00 | 0.09 |
| A/Hong Kong (4) | A/California (5) | |
|---|---|---|
| 20th Quartile | ||
| Trisomy 21 | −40.00 | 120.00 |
| Female | −80.00 | 20.00 |
| Constant | 160.00 ** | 20.00 |
| 40th Quartile | ||
| Trisomy 21 | 0.00 | 80.00 |
| Female | 0.00 | −80.00 |
| Constant | 160.00 | 160.00 |
| 60th Quartile | ||
| Trisomy 21 | 0.00 | 320.00 |
| Female | 0.00 | 160.00 |
| Constant | 160.00 | 160.00 |
| 80th Quartile | ||
| Trisomy 21 | −480.00 * | 0.00 |
| Female | −0.00 | 0.00 |
| Constant | 640.00 *** | 640.00 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
James, S.; Haight, R.C.; Hanna, C.; Furton, L. Adults with Trisomy 21 Have Differential Antibody Responses to Influenza A. Vaccines 2022, 10, 1145. https://doi.org/10.3390/vaccines10071145
James S, Haight RC, Hanna C, Furton L. Adults with Trisomy 21 Have Differential Antibody Responses to Influenza A. Vaccines. 2022; 10(7):1145. https://doi.org/10.3390/vaccines10071145
Chicago/Turabian StyleJames, Stephanie, Robert C. Haight, Cassandra Hanna, and Lindsey Furton. 2022. "Adults with Trisomy 21 Have Differential Antibody Responses to Influenza A" Vaccines 10, no. 7: 1145. https://doi.org/10.3390/vaccines10071145