
Impact of DNA Prime/Protein Boost Vaccination against Campylobacter jejuni on Immune Responses and Gut Microbiota in Chickens

Abstract
:1. Introduction
2. Materials and Methods
2.1. Production of DNA Vaccine
2.2. Production of the Recombinant Flagellin (recFlaA) Protein Vaccine
2.3. Campylobacter Strain and Growth
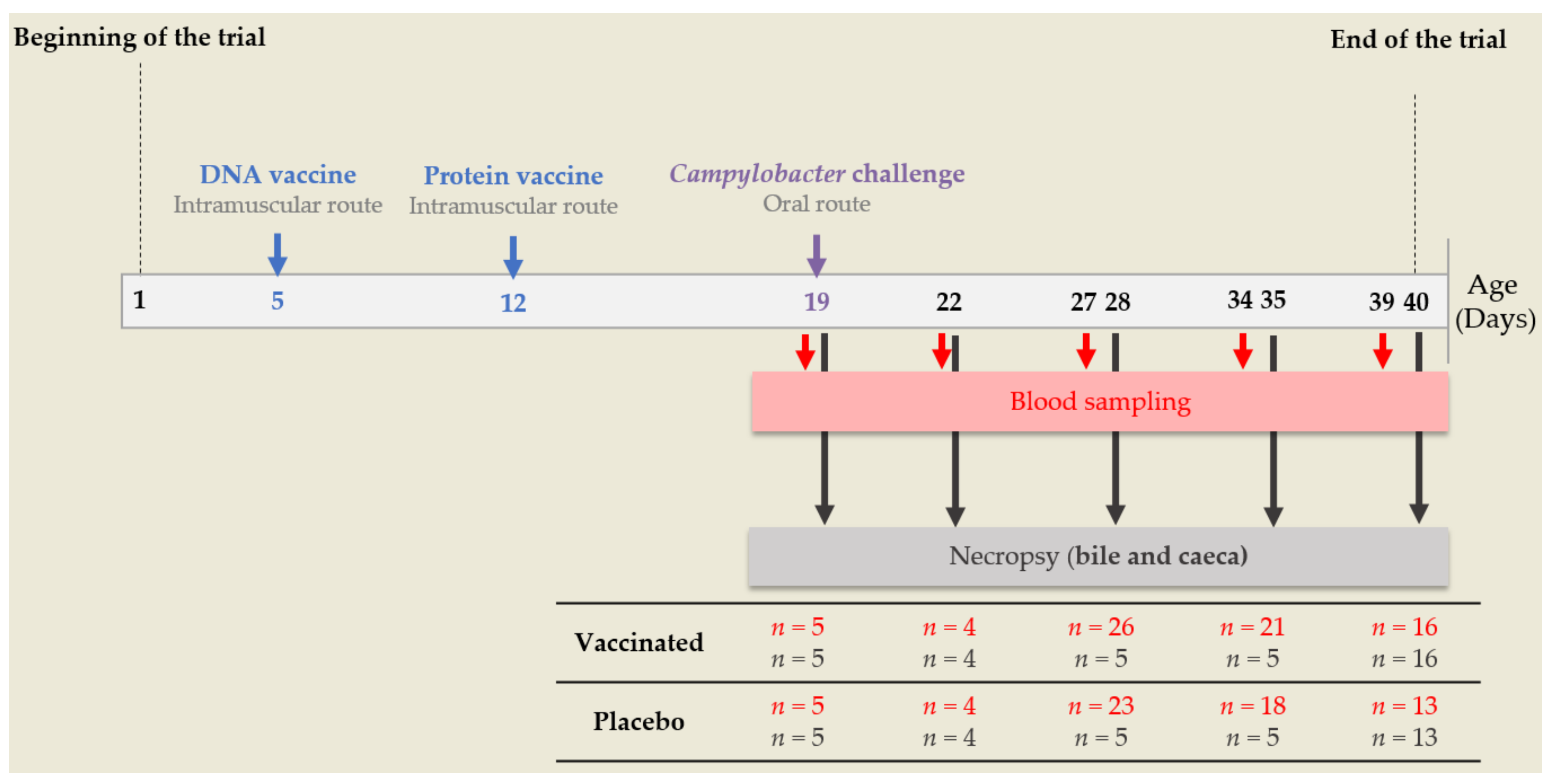
2.4. Avian Vaccine Experiment
2.5. Campylobacter Caecal Enumeration
2.6. Serum (IgY) and Bile (IgA) Anti-Flagellin Antibodies by Specific ELISAs
2.7. Relative Cytokine and Chemokine Expressions Determined by RT-qPCR
2.7.1. RNA Extraction
2.7.2. Reverse Transcription of Total RNA
2.7.3. qPCR
2.8. Statistical Analyses
2.9. DNA Extraction and PCR Amplification of 16S rRNA Gene Sequences and Microbiota Diversity Analysis
2.9.1. DNA Extraction
2.9.2. Sequencing of the V3/V4 Variable Region of the 16S Ribosomal Genes
2.9.3. Sequence Analyses
2.9.4. Statistical Analyses
3. Results
3.1. Body Weight
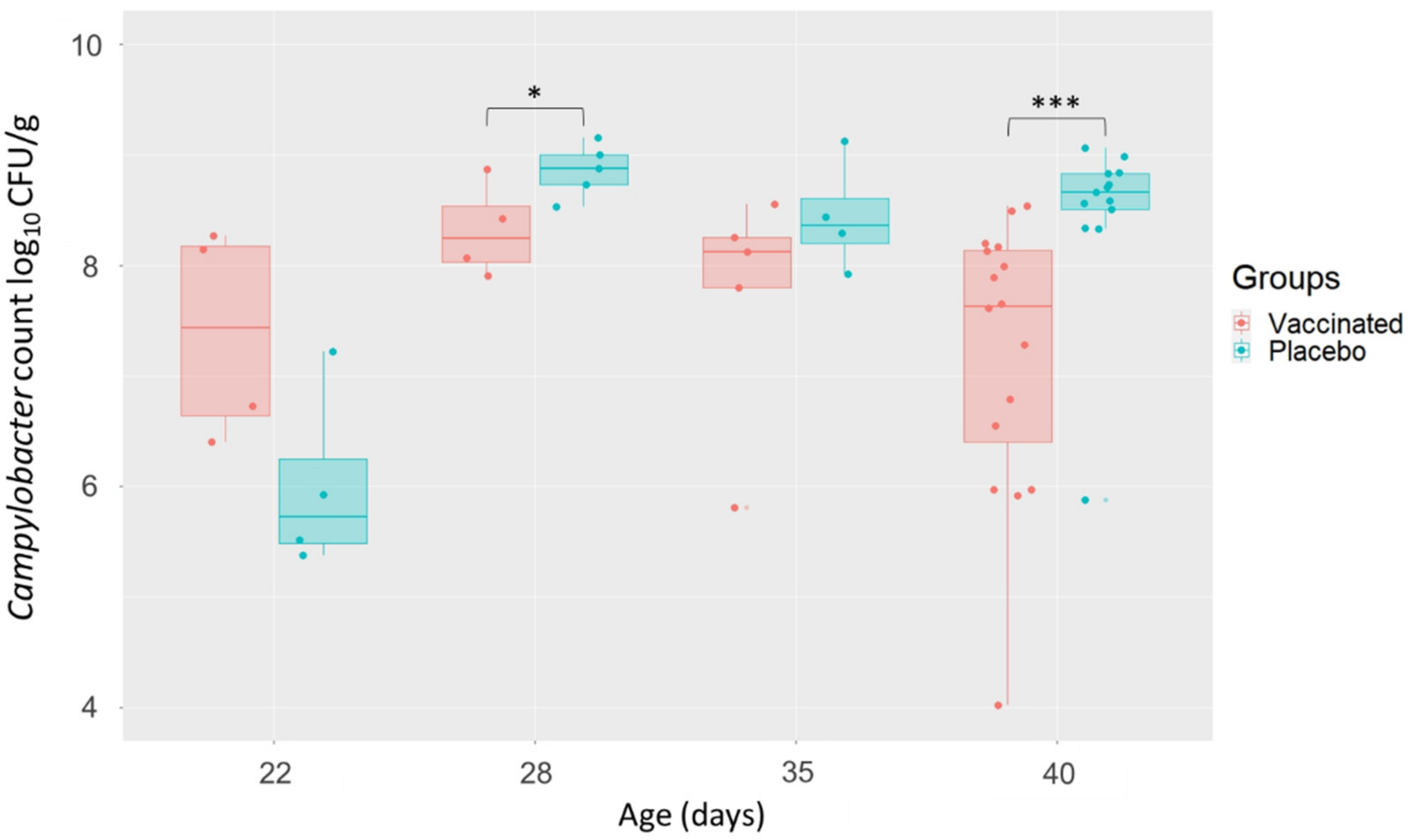
3.2. Campylobacter Caecal Enumeration
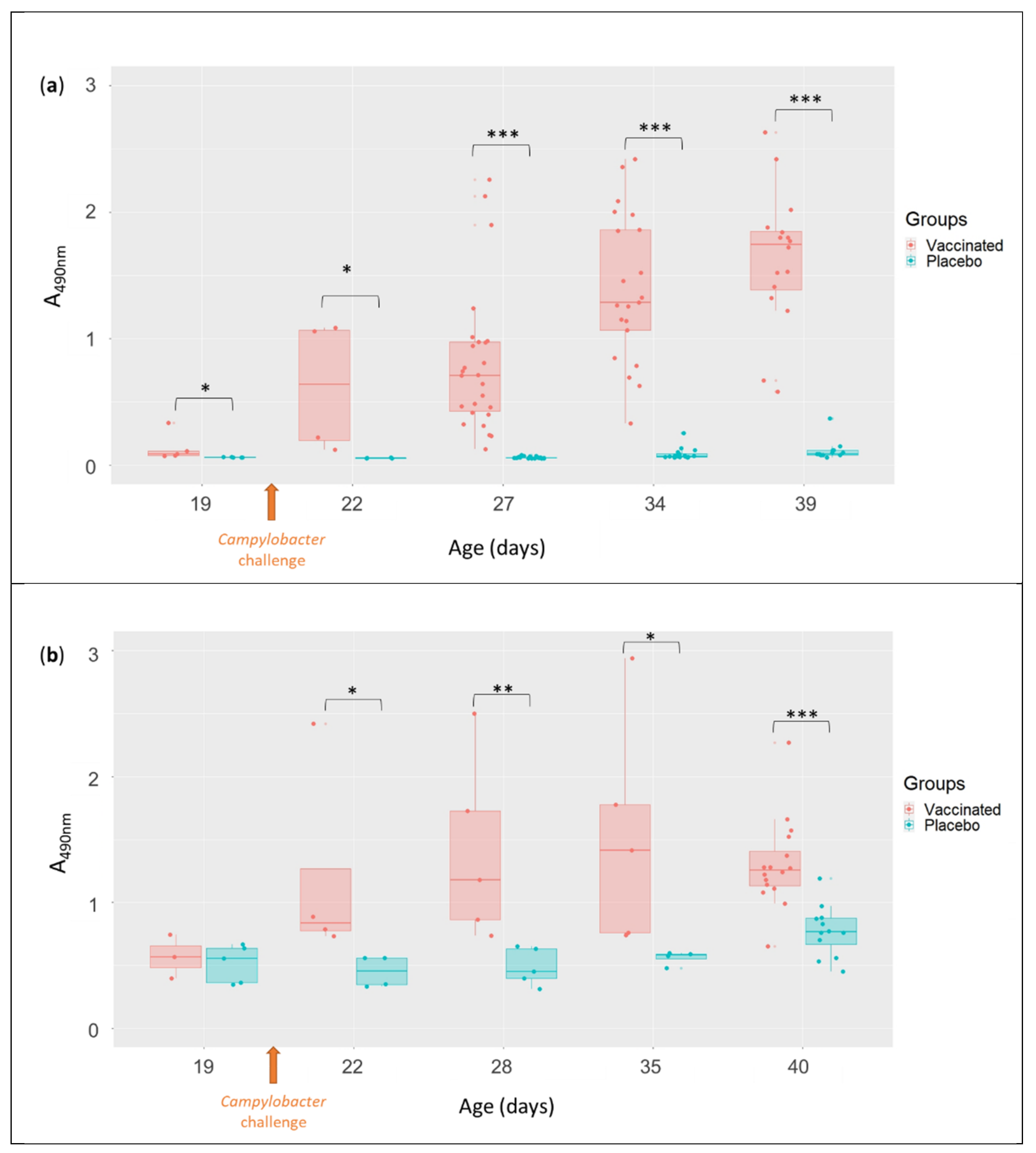
3.3. Serum (IgY) and Mucosal (IgA) Anti-Flagellin Antibody Levels
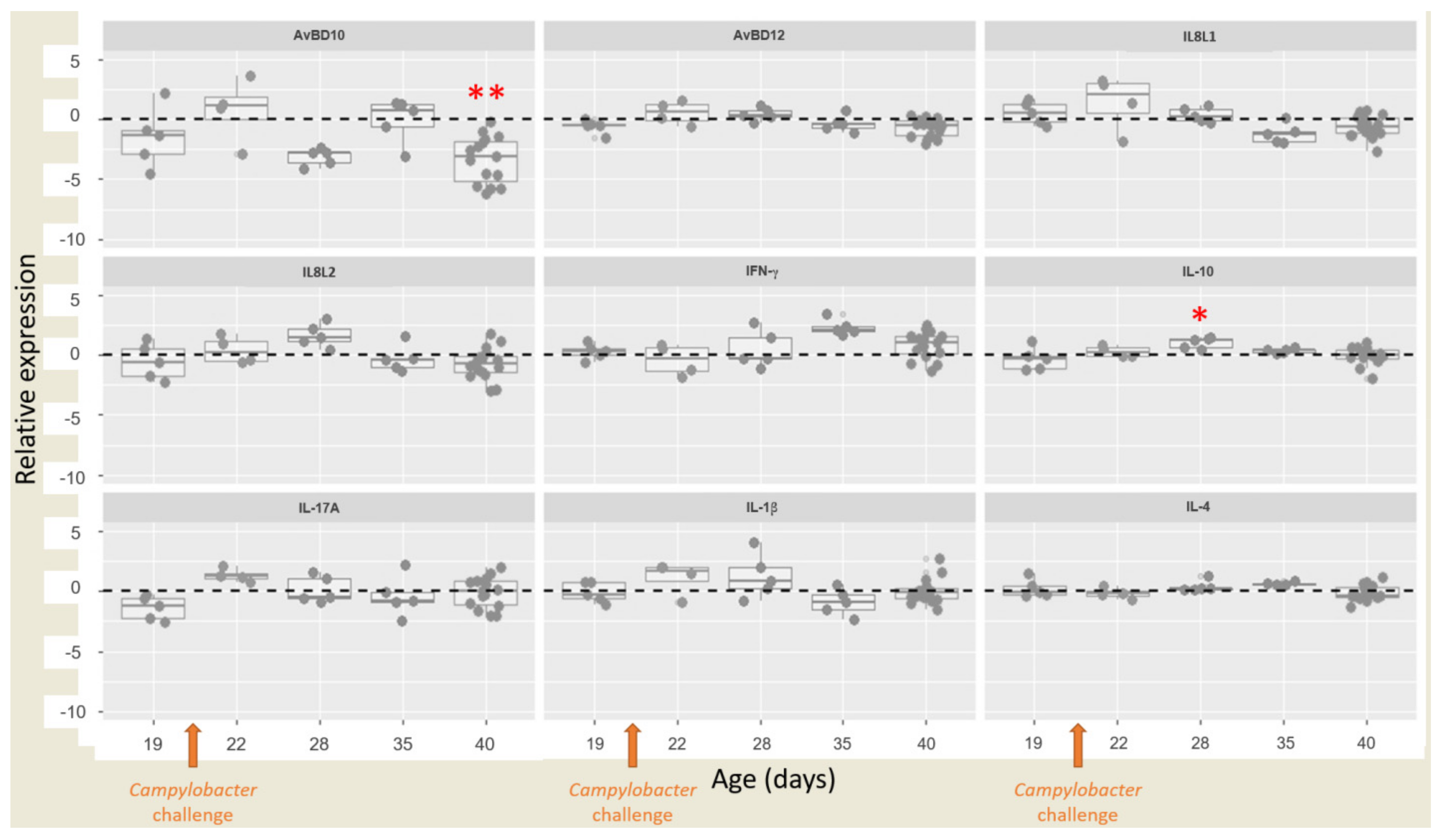
3.4. Relative Expressions of Cytokines/Chemokines and Antimicrobial Peptides
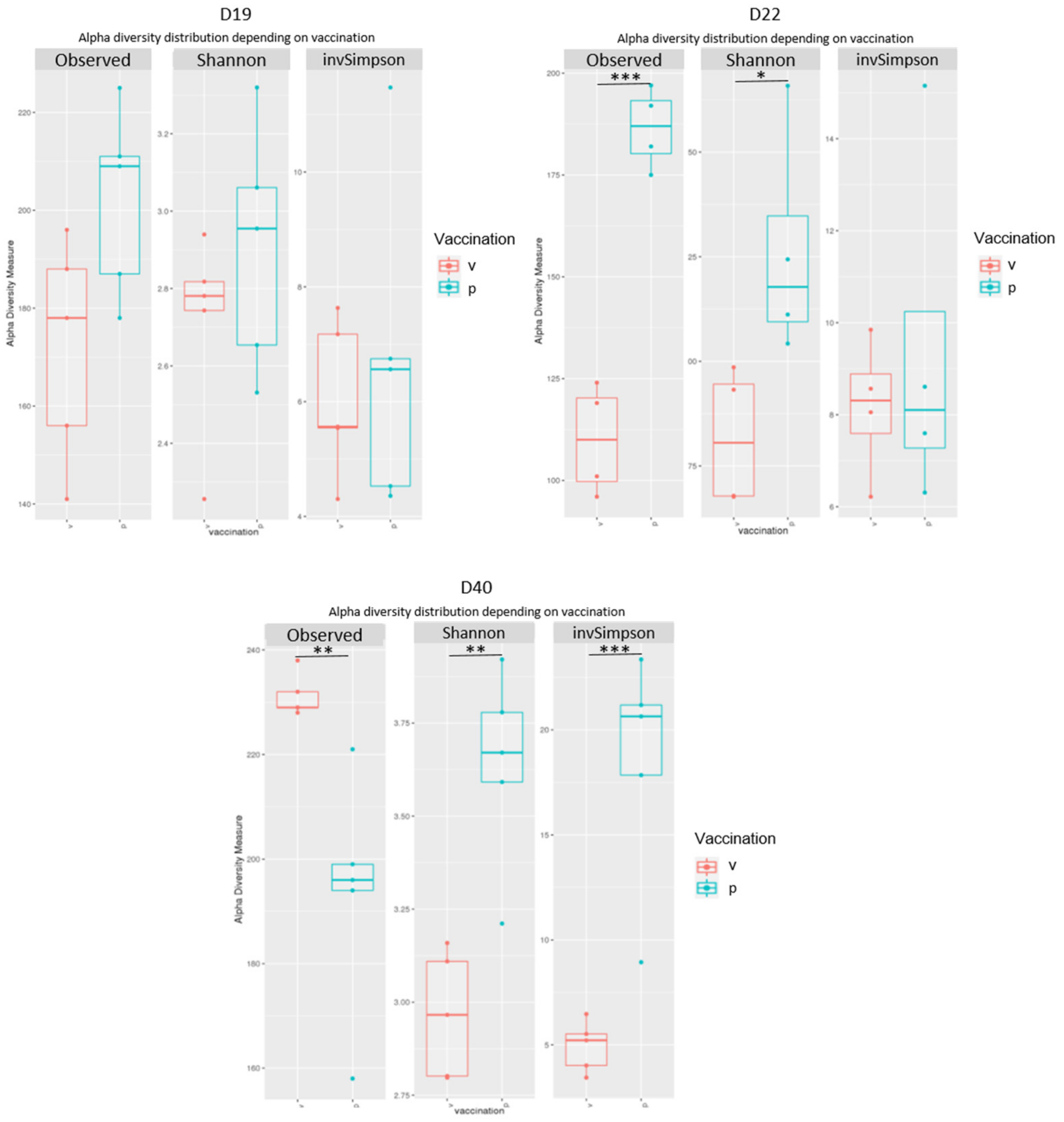
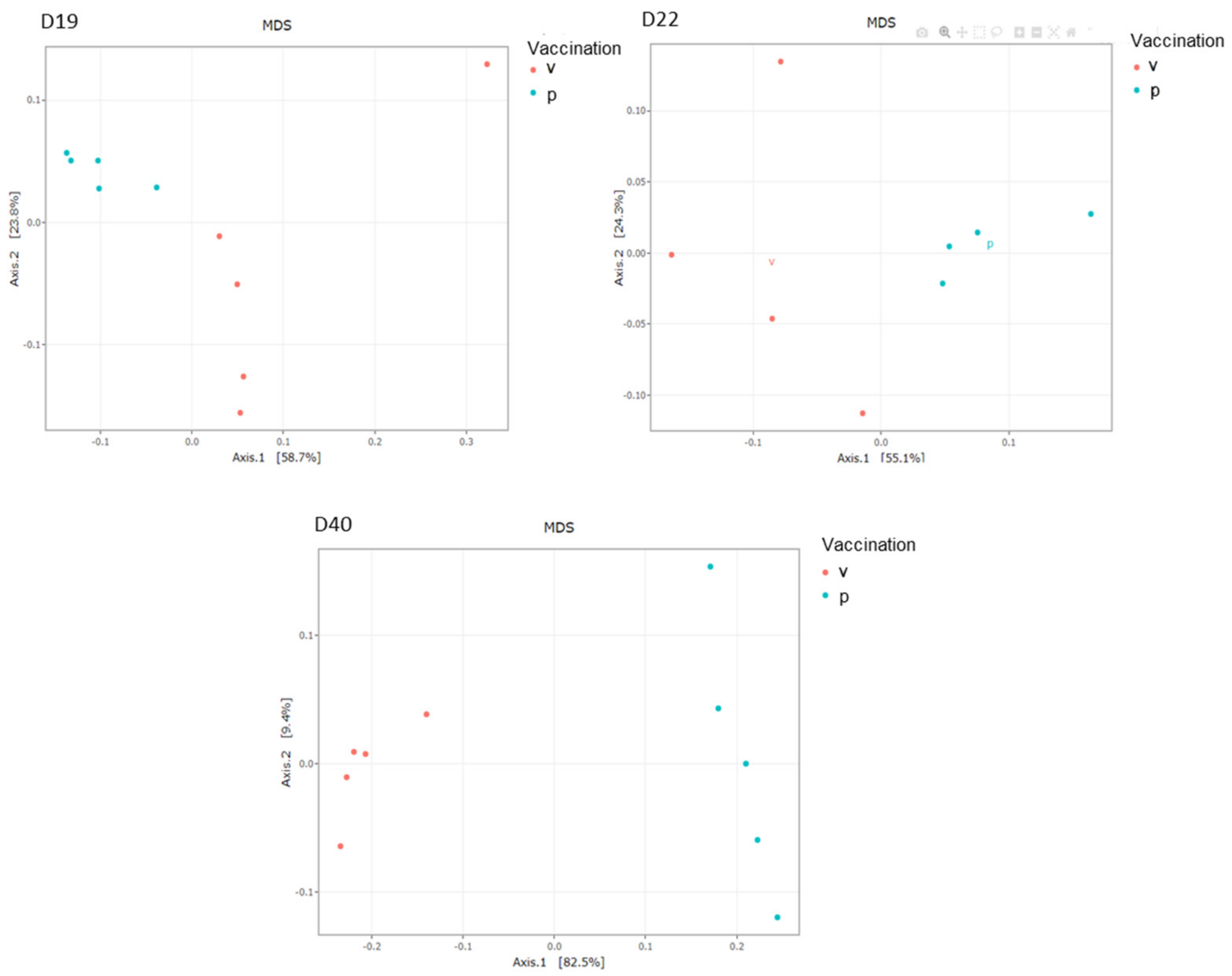
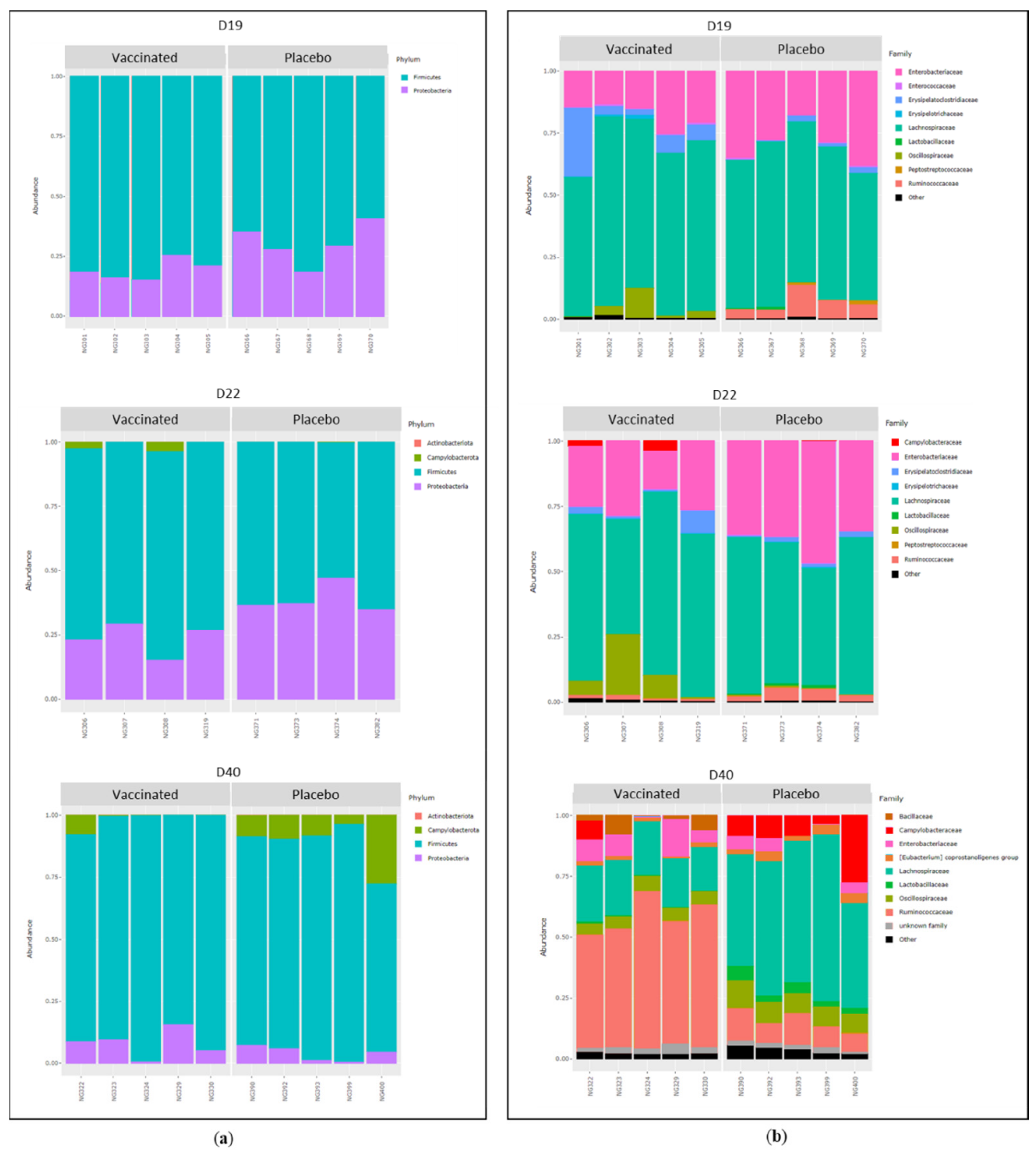
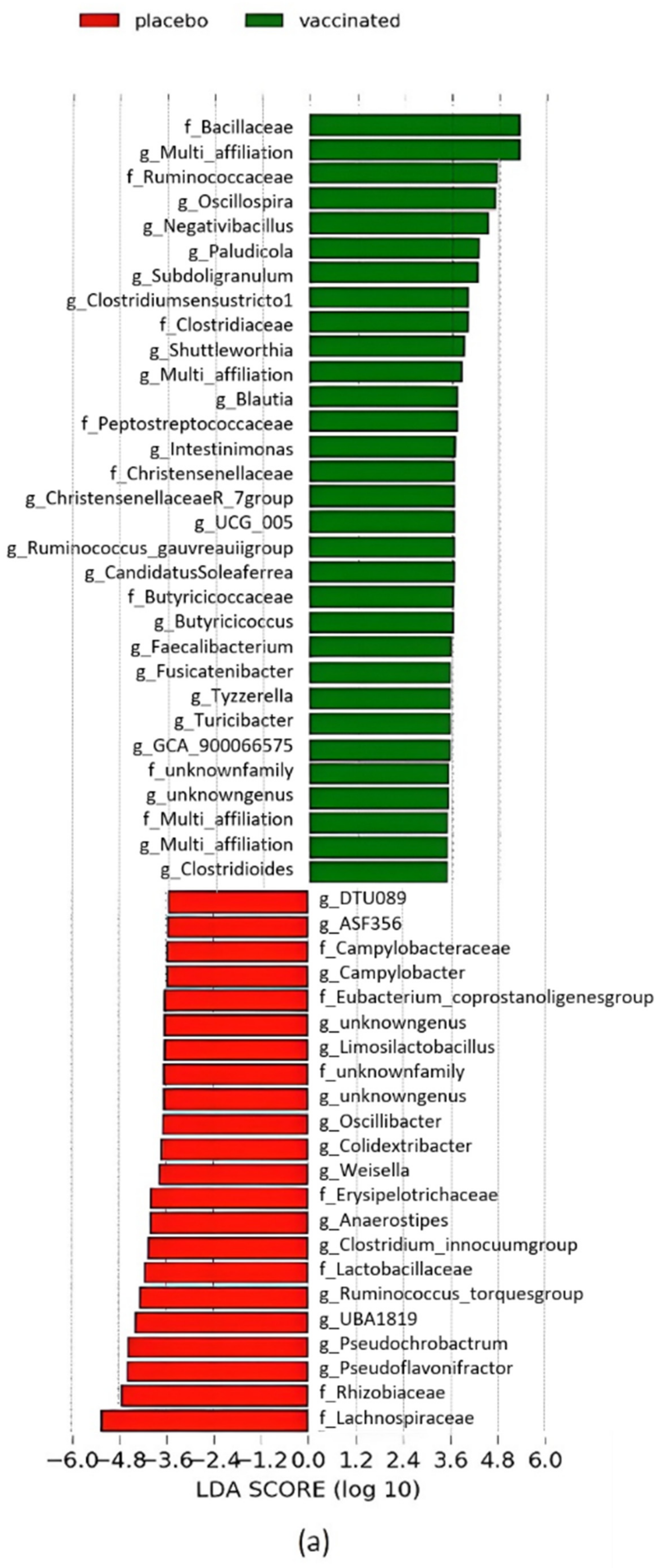
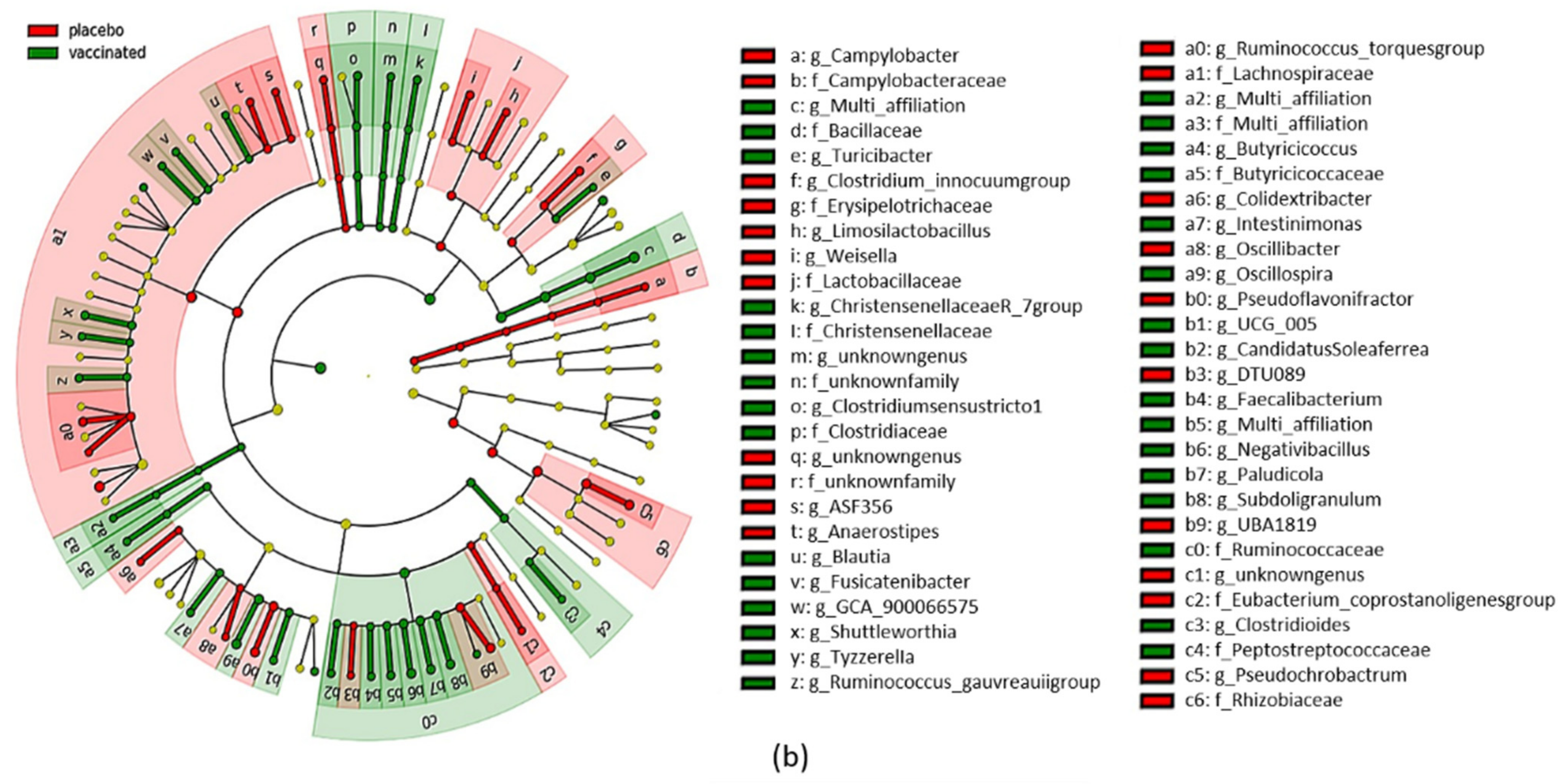
3.5. Caecal Microbiota Analyses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- EFSA and ECDC. The European Union One Health 2019 Zoonoses Report. EFSA J. 2021, 19, e06406. [Google Scholar] [CrossRef]
- Wassenaar, T.M.; Blaser, M.J. Pathophysiology of Campylobacter Jejuni Infections of Humans. Microbes Infect. 1999, 1, 1023–1033. [Google Scholar] [CrossRef]
- Nyati, K.K.; Nyati, R. Role of Campylobacter Jejuni Infection in the Pathogenesis of Guillain-Barré Syndrome: An Update. BioMed Res. Int. 2013, 2013, 852195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansson, I.; Pudas, N.; Harbom, B.; Engvall, E.O. Within-Flock Variations of Campylobacter Loads in Caeca and on Carcasses from Broilers. Int. J. Food Microbiol. 2010, 141, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Hue, O.; Allain, V.; Laisney, M.-J.; Le Bouquin, S.; Lalande, F.; Petetin, I.; Rouxel, S.; Quesne, S.; Gloaguen, P.-Y.; Picherot, M.; et al. Campylobacter Contamination of Broiler Caeca and Carcasses at the Slaughterhouse and Correlation with Salmonella Contamination. Food Microbiol. 2011, 28, 862–868. [Google Scholar] [CrossRef]
- EFSA Update and Review of Control Options for Campylobacter in Broilers at Primary Production. EFSA J. 2020, 18, e06090. [CrossRef]
- The European Commission. Commission Regulation (EU) 2017/1495 of 23 August 2017 Amending Regulation (EC) No 2073/2005 as Regards Campylobacter in Broiler Carcases. Off. J. Eur. Union 2017, L218, 1–6. Available online: https://eur-lex.europa.eu/legal-content/EN/TXT/?uri=CELEX:32017R1495 (accessed on 2 May 2022).
- Nothaft, H.; Davis, B.; Lock, Y.Y.; Perez-Munoz, M.E.; Vinogradov, E.; Walter, J.; Coros, C.; Szymanski, C.M. Engineering the Campylobacter Jejuni N-Glycan to Create an Effective Chicken Vaccine. Sci. Rep. 2016, 6, 26511. [Google Scholar] [CrossRef] [Green Version]
- Meunier, M.; Guyard-Nicodème, M.; Vigouroux, E.; Poezevara, T.; Beven, V.; Quesne, S.; Bigault, L.; Amelot, M.; Dory, D.; Chemaly, M. Promising New Vaccine Candidates against Campylobacter in Broilers. PLoS ONE 2017, 12, e0188472. [Google Scholar] [CrossRef] [Green Version]
- Meunier, M.; Guyard-Nicodème, M.; Vigouroux, E.; Poezevara, T.; Béven, V.; Quesne, S.; Amelot, M.; Parra, A.; Chemaly, M.; Dory, D. A DNA Prime/Protein Boost Vaccine Protocol Developed against Campylobacter Jejuni for Poultry. Vaccine 2018, 36, 2119–2125. [Google Scholar] [CrossRef]
- Cui, Y.; Guo, F.; Guo, J.; Cao, X.; Wang, H.; Yang, B.; Zhou, H.; Su, X.; Zeng, X.; Lin, J.; et al. Immunization of Chickens with the Enterobactin Conjugate Vaccine Reduced Campylobacter Jejuni Colonization in the Intestine. Vaccines 2020, 8, 747. [Google Scholar] [CrossRef]
- Vohra, P.; Chintoan-Uta, C.; Terra, V.S.; Bremner, A.; Cuccui, J.; Wren, B.W.; Vervelde, L.; Stevens, M.P. Evaluation of Glycosylated FlpA and SodB as Subunit Vaccines Against Campylobacter Jejuni Colonisation in Chickens. Vaccines 2020, 8, 520. [Google Scholar] [CrossRef]
- Sahin, O.; Luo, N.; Huang, S.; Zhang, Q. Effect of Campylobacter-Specific Maternal Antibodies on Campylobacter Jejuni Colonization in Young Chickens. Appl. Environ. Microbiol. 2003, 69, 5372–5379. [Google Scholar] [CrossRef] [Green Version]
- Shoaf-Sweeney, K.D.; Larson, C.L.; Tang, X.; Konkel, M.E. Identification of Campylobacter Jejuni Proteins Recognized by Maternal Antibodies of Chickens. Appl. Environ. Microbiol. 2008, 74, 6867–6875. [Google Scholar] [CrossRef] [Green Version]
- De Zoete, M.R.; Keestra, A.M.; Roszczenko, P.; van Putten, J.P.M. Activation of Human and Chicken Toll-Like Receptors by Campylobacter spp. Infect. Immun. 2010, 78, 1229–1238. [Google Scholar] [CrossRef] [Green Version]
- Reid, W.D.K.; Close, A.J.; Humphrey, S.; Chaloner, G.; Lacharme-Lora, L.; Rothwell, L.; Kaiser, P.; Williams, N.J.; Humphrey, T.J.; Wigley, P.; et al. Cytokine Responses in Birds Challenged with the Human Food-Borne Pathogen Campylobacter Jejuni Implies a Th17 Response. R. Soc. Open Sci. 2016, 3, 150541. [Google Scholar] [CrossRef] [Green Version]
- Han, Z.; Willer, T.; Li, L.; Pielsticker, C.; Rychlik, I.; Velge, P.; Kaspers, B.; Rautenschlein, S. Influence of the Gut Microbiota Composition on Campylobacter Jejuni Colonization in Chickens. Infect. Immun. 2017, 85, e00380-17. [Google Scholar] [CrossRef] [Green Version]
- Lacharme-Lora, L.; Chaloner, G.; Gilroy, R.; Humphrey, S.; Gibbs, K.; Jopson, S.; Wright, E.; Reid, W.; Ketley, J.; Humphrey, T.; et al. B Lymphocytes Play a Limited Role in Clearance of Campylobacter Jejuni from the Chicken Intestinal Tract. Sci. Rep. 2017, 7, 45090. [Google Scholar] [CrossRef]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The Effect of the Timing of Exposure to Campylobacter Jejuni on the Gut Microbiome and Inflammatory Responses of Broiler Chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef]
- Garcia, J.S.; Byrd, J.A.; Wong, E.A. Expression of Nutrient Transporters and Host Defense Peptides in Campylobacter Challenged Broilers. Poult. Sci. 2018, 97, 3671–3680. [Google Scholar] [CrossRef]
- Skånseng, B.; Kaldhusdal, M.; Rudi, K. Comparison of Chicken Gut Colonisation by the Pathogens Campylobacter Jejuni and Clostridium Perfringens by Real-Time Quantitative PCR. Mol. Cell. Probes 2006, 20, 269–279. [Google Scholar] [CrossRef]
- Sergeant, M.J.; Constantinidou, C.; Cogan, T.A.; Bedford, M.R.; Penn, C.W.; Pallen, M.J. Extensive Microbial and Functional Diversity within the Chicken Cecal Microbiome. PLoS ONE 2014, 9, e91941. [Google Scholar] [CrossRef]
- Wei, S.; Morrison, M.; Yu, Z. Bacterial Census of Poultry Intestinal Microbiome. Poult. Sci. 2013, 92, 671–683. [Google Scholar] [CrossRef]
- Qu, A.; Brulc, J.M.; Wilson, M.K.; Law, B.F.; Theoret, J.R.; Joens, L.A.; Konkel, M.E.; Angly, F.; Dinsdale, E.A.; Edwards, R.A.; et al. Comparative Metagenomics Reveals Host Specific Metavirulomes and Horizontal Gene Transfer Elements in the Chicken Cecum Microbiome. PLoS ONE 2008, 3, e2945. [Google Scholar] [CrossRef] [Green Version]
- Thibodeau, A.; Fravalo, P.; Yergeau, É.; Arsenault, J.; Lahaye, L.; Letellier, A. Chicken Caecal Microbiome Modifications Induced by Campylobacter Jejuni Colonization and by a Non-Antibiotic Feed Additive. PLoS ONE 2015, 10, e0131978. [Google Scholar] [CrossRef] [Green Version]
- Taha-Abdelaziz, K.; Yitbarek, A.; Alkie, T.N.; Hodgins, D.C.; Read, L.R.; Weese, J.S.; Sharif, S. PLGA-Encapsulated CpG ODN and Campylobacter Jejuni Lysate Modulate Cecal Microbiota Composition in Broiler Chickens Experimentally Challenged with C. Jejuni. Sci. Rep. 2018, 8, 12076. [Google Scholar] [CrossRef] [Green Version]
- Awad, W.A.; Mann, E.; Dzieciol, M.; Hess, C.; Schmitz-Esser, S.; Wagner, M.; Hess, M. Age-Related Differences in the Luminal and Mucosa-Associated Gut Microbiome of Broiler Chickens and Shifts Associated with Campylobacter Jejuni Infection. Front. Cell. Infect. Microbiol. 2016, 6, 154. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.K.; AbuOun, M.; Cawthraw, S.A.; Humphrey, T.J.; Rothwell, L.; Kaiser, P.; Barrow, P.A.; Jones, M.A. Campylobacter Colonization of the Chicken Induces a Proinflammatory Response in Mucosal Tissues. FEMS Immunol. Med. Microbiol. 2008, 54, 114–121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thépault, A.; Roulleau, X.; Loiseau, P.; Cauquil, L.; Poezevara, T.; Hyronimus, B.; Quesne, S.; Souchaud, F.; Keita, A.; Chemaly, M.; et al. Effect of Litter Treatment on Campylobacter Jejuni in Broilers and on Cecal Microbiota. Pathogens 2020, 9, 333. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M.; Wang, D.; Fitzgerald, S.D.; Sun, S. Severe Necrotic Dermatitis in the Combs of Line 63 Chickens Infected with Marek’s Disease Virus. Avian Pathol. 2016, 45, 582–592. [Google Scholar] [CrossRef] [Green Version]
- Nang, N.T.; Lee, J.S.; Song, B.M.; Kang, Y.M.; Kim, H.S.; Seo, S.H. Induction of Inflammatory Cytokines and Toll-like Receptors in Chickens Infected with Avian H9N2 Influenza Virus. Vet. Res. 2011, 42, 64. [Google Scholar] [CrossRef] [Green Version]
- Cox, C.M.; Sumners, L.H.; Kim, S.; McElroy, A.P.; Bedford, M.R.; Dalloul, R.A. Immune Responses to Dietary β-Glucan in Broiler Chicks during an Eimeria Challenge. Poult. Sci. 2010, 89, 2597–2607. [Google Scholar] [CrossRef]
- Rasoli, M.; Yeap, S.K.; Tan, S.W.; Roohani, K.; Kristeen-Teo, Y.W.; Alitheen, N.B.; Rahaman, Y.A.; Aini, I.; Bejo, M.H.; Kaiser, P.; et al. Differential Modulation of Immune Response and Cytokine Profiles in the Bursae and Spleen of Chickens Infected with Very Virulent Infectious Bursal Disease Virus. BMC Vet. Res. 2015, 11, 75. [Google Scholar] [CrossRef]
- Su, S.; Dwyer, D.M.; Miska, K.B.; Fetterer, R.H.; Jenkins, M.C.; Wong, E.A. Expression of Host Defense Peptides in the Intestine of Eimeria-Challenged Chickens. Poult. Sci. 2017, 96, 2421–2427. [Google Scholar] [CrossRef]
- Escudié, F.; Auer, L.; Bernard, M.; Mariadassou, M.; Cauquil, L.; Vidal, K.; Maman, S.; Hernandez-Raquet, G.; Combes, S.; Pascal, G. FROGS: Find, Rapidly, OTUs with Galaxy Solution. Bioinformatics 2018, 34, 1287–1294. [Google Scholar] [CrossRef]
- Lynn, D.J.; Higgs, R.; Lloyd, A.T.; O’Farrelly, C.; Hervé-Grépinet, V.; Nys, Y.; Brinkman, F.S.L.; Yu, P.-L.; Soulier, A.; Kaiser, P.; et al. Avian Beta-Defensin Nomenclature: A Community Proposed Update. Immunol. Lett. 2007, 110, 86–89. [Google Scholar] [CrossRef]
- Min, W.; Kim, W.H.; Lillehoj, E.P.; Lillehoj, H.S. Recent Progress in Host Immunity to Avian Coccidiosis: IL-17 Family Cytokines as Sentinels of the Intestinal Mucosa. Dev. Comp. Immunol. 2013, 41, 418–428. [Google Scholar] [CrossRef]
- Chaloner, G.; Wigley, P.; Humphrey, S.; Kemmett, K.; Lacharme-Lora, L.; Humphrey, T.; Williams, N. Dynamics of Dual Infection with Campylobacter Jejuni Strains in Chickens Reveals Distinct Strain-to-Strain Variation in Infection Ecology. Appl. Environ. Microbiol. 2014, 80, 6366–6372. [Google Scholar] [CrossRef] [Green Version]
- Widders, P.R.; Thomas, L.M.; Long, K.A.; Tokhi, M.A.; Panaccio, M.; Apos, E. The Specificity of Antibody in Chickens Immunised to Reduce Intestinal Colonisation with Campylobacter Jejuni. Vet. Microbiol. 1998, 64, 39–50. [Google Scholar] [CrossRef]
- Radomska, K.A.; Vaezirad, M.M.; Verstappen, K.M.; Wösten, M.M.S.M.; Wagenaar, J.A.; Putten, J.P.M. van Chicken Immune Response after In Ovo Immunization with Chimeric TLR5 Activating Flagellin of Campylobacter Jejuni. PLoS ONE 2016, 11, e0164837. [Google Scholar] [CrossRef] [Green Version]
- Huang, J.; Yin, Y.-X.; Pan, Z.; Zhang, G.; Zhu, A.; Liu, X.; Jiao, X. Intranasal Immunization with Chitosan/PCAGGS-FlaA Nanoparticles Inhibits Campylobacter Jejuni in a White Leghorn Model. J. Biomed. Biotechnol. 2010, 2010, 589476. [Google Scholar] [CrossRef] [Green Version]
- Neal-McKinney, J.M.; Samuelson, D.R.; Eucker, T.P.; Nissen, M.S.; Crespo, R.; Konkel, M.E. Reducing Campylobacter Jejuni Colonization of Poultry via Vaccination. PLoS ONE 2014, 9, e114254. [Google Scholar] [CrossRef] [Green Version]
- Chintoan-Uta, C.; Cassady-Cain, R.L.; Al-Haideri, H.; Watson, E.; Kelly, D.J.; Smith, D.G.E.; Sparks, N.H.C.; Kaiser, P.; Stevens, M.P. Superoxide Dismutase SodB Is a Protective Antigen against Campylobacter Jejuni Colonisation in Chickens. Vaccine 2015, 33, 6206–6211. [Google Scholar] [CrossRef] [Green Version]
- Davis, S.K.; Selva, K.J.; Kent, S.J.; Chung, A.W. Serum IgA Fc Effector Functions in Infectious Disease and Cancer. Immunol. Cell Biol. 2020, 98, 276–286. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Swaggerty, C.L.; Kogut, M.H.; Chiang, H.-I.; Wang, Y.; Genovese, K.J.; He, H.; Zhou, H. Gene Expression Profiling of the Local Cecal Response of Genetic Chicken Lines That Differ in Their Susceptibility to Campylobacter Jejuni Colonization. PLoS ONE 2010, 5, e11827. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, S.; Chaloner, G.; Kemmett, K.; Davidson, N.; Williams, N.; Kipar, A.; Humphrey, T.; Wigley, P. Campylobacter Jejuni Is Not Merely a Commensal in Commercial Broiler Chickens and Affects Bird Welfare. mBio 2014, 5, e01364-14. [Google Scholar] [CrossRef] [Green Version]
- Šefcová, M.; Larrea-Álvarez, M.; Larrea-Álvarez, C.; Revajová, V.; Karaffová, V.; Koščová, J.; Nemcová, R.; Ortega-Paredes, D.; Vinueza-Burgos, C.; Levkut, M.; et al. Effects of Lactobacillus Fermentum Supplementation on Body Weight and Pro-Inflammatory Cytokine Expression in Campylobacter Jejuni-Challenged Chickens. Vet. Sci. 2020, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Cerqueira, C.; Manfroi, B.; Fillatreau, S. IL-10-Producing Regulatory B Cells and Plasmocytes: Molecular Mechanisms and Disease Relevance. Semin. Immunol. 2019, 44, 101323. [Google Scholar] [CrossRef] [PubMed]
- Faber, E.; Gripp, E.; Maurischat, S.; Kaspers, B.; Tedin, K.; Menz, S.; Zuraw, A.; Kershaw, O.; Yang, I.; Rautenschlein, S.; et al. Novel Immunomodulatory Flagellin-Like Protein FlaC in Campylobacter Jejuni and Other Campylobacterales. mSphere 2015, 1, e00028-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thibodeau, A.; Letellier, A.; Yergeau, É.; Larrivière-Gauthier, G.; Fravalo, P. Lack of Evidence That Selenium-Yeast Improves Chicken Health and Modulates the Caecal Microbiota in the Context of Colonization by Campylobacter Jejuni. Front. Microbiol. 2017, 8, 451. [Google Scholar] [CrossRef] [Green Version]
- Kogut, M.H.; Lee, A.; Santin, E. Microbiome and Pathogen Interaction with the Immune System. Poult. Sci. 2020, 99, 1906–1913. [Google Scholar] [CrossRef]
- Hermans, D.; Pasmans, F.; Heyndrickx, M.; Immerseel, F.V.; Martel, A.; Deun, K.V.; Haesebrouck, F. A Tolerogenic Mucosal Immune Response Leads to Persistent Campylobacter Jejuni Colonization in the Chicken Gut. Crit. Rev. Microbiol. 2012, 38, 17–29. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Primer Sequence (5′-3′) | Product Size (bp) | NCBI Accession Number | Reference |
|---|---|---|---|---|
| B-actin | F: CCCACCTGAGCGCAAGTACT R: AAGCATTTGCGGTGGACAAT | 132 | NM_205518.1 | [31] |
| IFN-γ | F: TGAGCCAGATTGTTTCGATG R: CTTGGCCAGGTCCATGATA | 152 | NM_205149.1 | [32] |
| IL-1β | F: GTGAGGCTCAACATTGCGCTGTA R: TGTCCAGGCGGTAGAAGATGAAG | 214 | NM_204524.1 | [32] |
| IL-4 | F: GCTCTCAGTGCCGCTGATG R: GAAACCTCTCCCTGGATGTCAT | 60 | NM_204524.1 | [33] |
| IL-10 | F: CGCTGTCACCGCTTCTTCA R: CGAACGTCTCCTTGATCTGCTT | 67 | NM_001004414.2 | Primer Express® |
| IL-17A | R: CATGGGATTACAGGATCGATGA F: GCGGCACTGGGCATCA | 68 | NM_204460.1 | [16] |
| IL8L1 | F: CCGATGCCAGTGCATAGAG R: CCTTGTCCAGAATTGCCTTG | 191 | NM_205018.1 | [34] |
| IL8L2 | F: CCTGGTTTCAGCTGCTCTGT R: GCGTCAGCTTCACATCTTGA | 128 | NM_205498.1 | [34] |
| AvBD10 | F: CAGACCCACTTTTCCCTGACA R: CCCAGCACGGCAGAAATT | 64 | NM_001001609.2 | [35] |
| AvBD12 | F: TGTAACCACGACAGGGGATTG R: GGGAGTTGGTGACAGAGGTTT | 114 | NM_001001607.2 | [35] |
| Groups | Day 19 | Day 22 | Day 28 | Day 35 | Day 40 |
|---|---|---|---|---|---|
| Placebo | 181 ± 27 | 218 ± 37 | 312 ± 40 | 432 ± 53 | 524 ± 74 |
| Vaccinated | 183 ± 22 | 223 ± 30 | 319 ± 31 | 443 ± 48 | 530 ± 58 |
| D19 | D22 | D40 | |
|---|---|---|---|
| Number of sequences after read demultiplexing and pre-process (merging, denoising and dereplication) | 1,133,635 (min: 74,225; max: 153,026) | 735,454 (min: 61, 456; max: 125,181) | 1,032,938 (min: 45,196; max: 163,639) |
| Number of OTU | 332 Placebo: (min: 219; max:263) Vaccinated: (min: 163, max:234) | 348 Placebo: (min: 256; max: 299) Vaccinated: (min: 115, max: 182) | 412 Placebo: (min: 203; max: 296) Vaccinated: (min: 271, max: 314) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gloanec, N.; Dory, D.; Quesne, S.; Béven, V.; Poezevara, T.; Keita, A.; Chemaly, M.; Guyard-Nicodème, M. Impact of DNA Prime/Protein Boost Vaccination against Campylobacter jejuni on Immune Responses and Gut Microbiota in Chickens. Vaccines 2022, 10, 981. https://doi.org/10.3390/vaccines10060981
Gloanec N, Dory D, Quesne S, Béven V, Poezevara T, Keita A, Chemaly M, Guyard-Nicodème M. Impact of DNA Prime/Protein Boost Vaccination against Campylobacter jejuni on Immune Responses and Gut Microbiota in Chickens. Vaccines. 2022; 10(6):981. https://doi.org/10.3390/vaccines10060981
Chicago/Turabian StyleGloanec, Noémie, Daniel Dory, Ségolène Quesne, Véronique Béven, Typhaine Poezevara, Alassane Keita, Marianne Chemaly, and Muriel Guyard-Nicodème. 2022. "Impact of DNA Prime/Protein Boost Vaccination against Campylobacter jejuni on Immune Responses and Gut Microbiota in Chickens" Vaccines 10, no. 6: 981. https://doi.org/10.3390/vaccines10060981