
Efficacy of a Modified Live Porcine Reproductive and Respiratory Syndrome Virus 1 (PRRSV-1) Vaccine against Experimental Infection with PRRSV AUT15-33 in Weaned Piglets

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Challenge Virus
2.3. Sample Collection and Necropsy
2.4. Histologic Lung Assessment
2.5. Serum Antibody Detection
2.6. Virological Analysis—PRRSV qRT-PCR
2.7. Statistical Analysis
3. Results
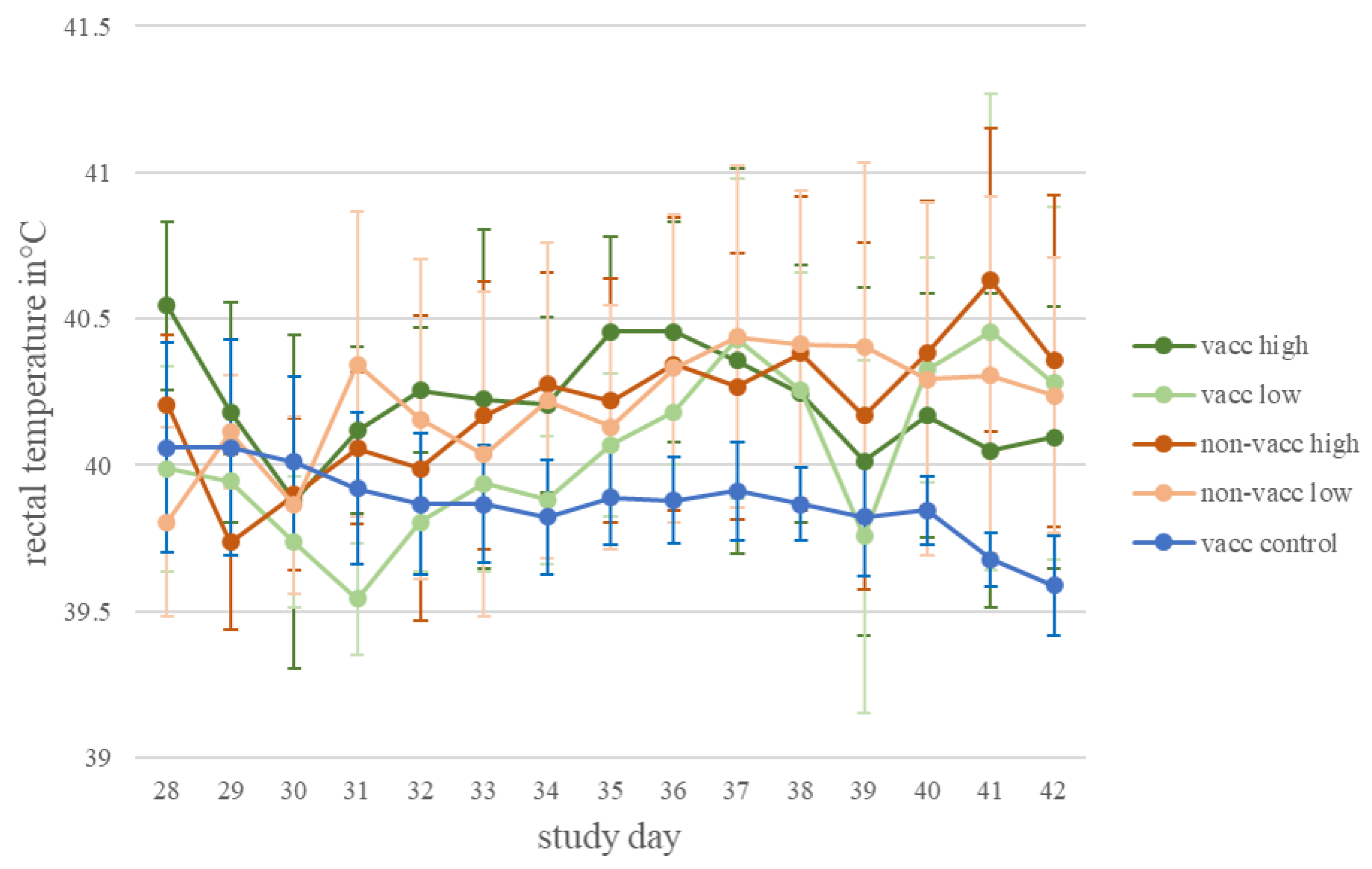
3.1. Clinical Observation
3.2. Cough Monitor
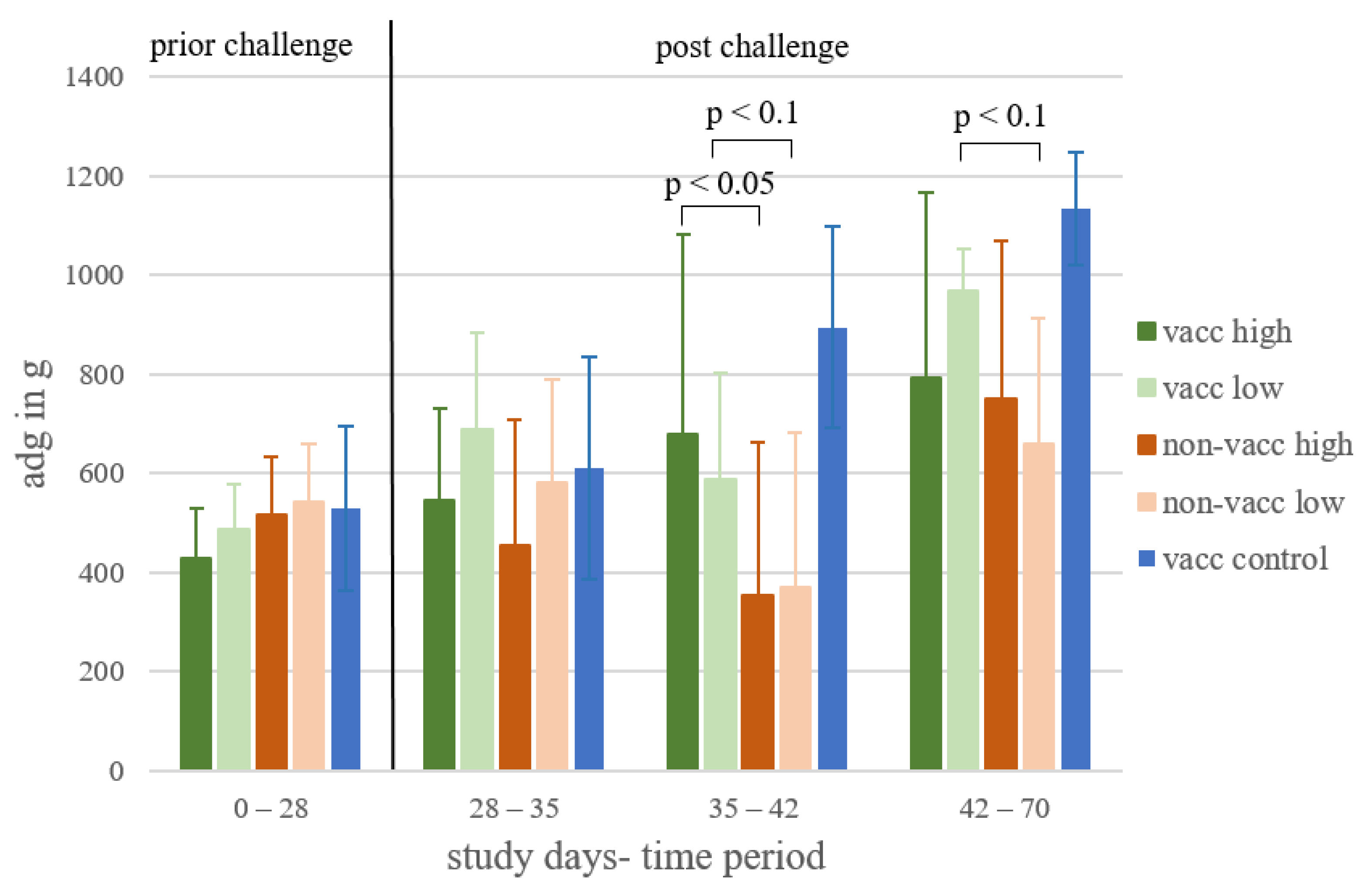
3.3. Average Daily Weight Gain
3.4. Serology
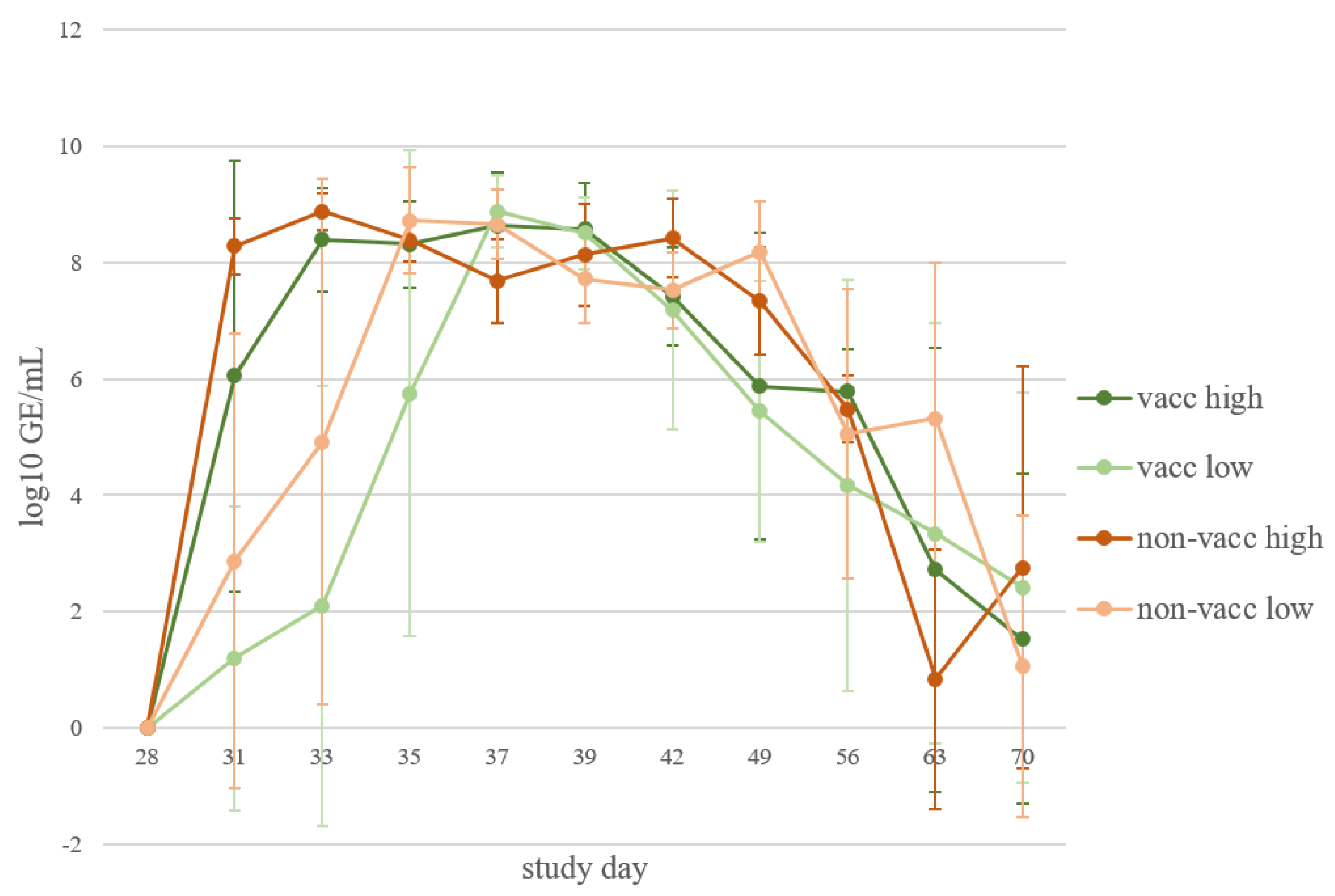
3.5. Viral Load in Serum
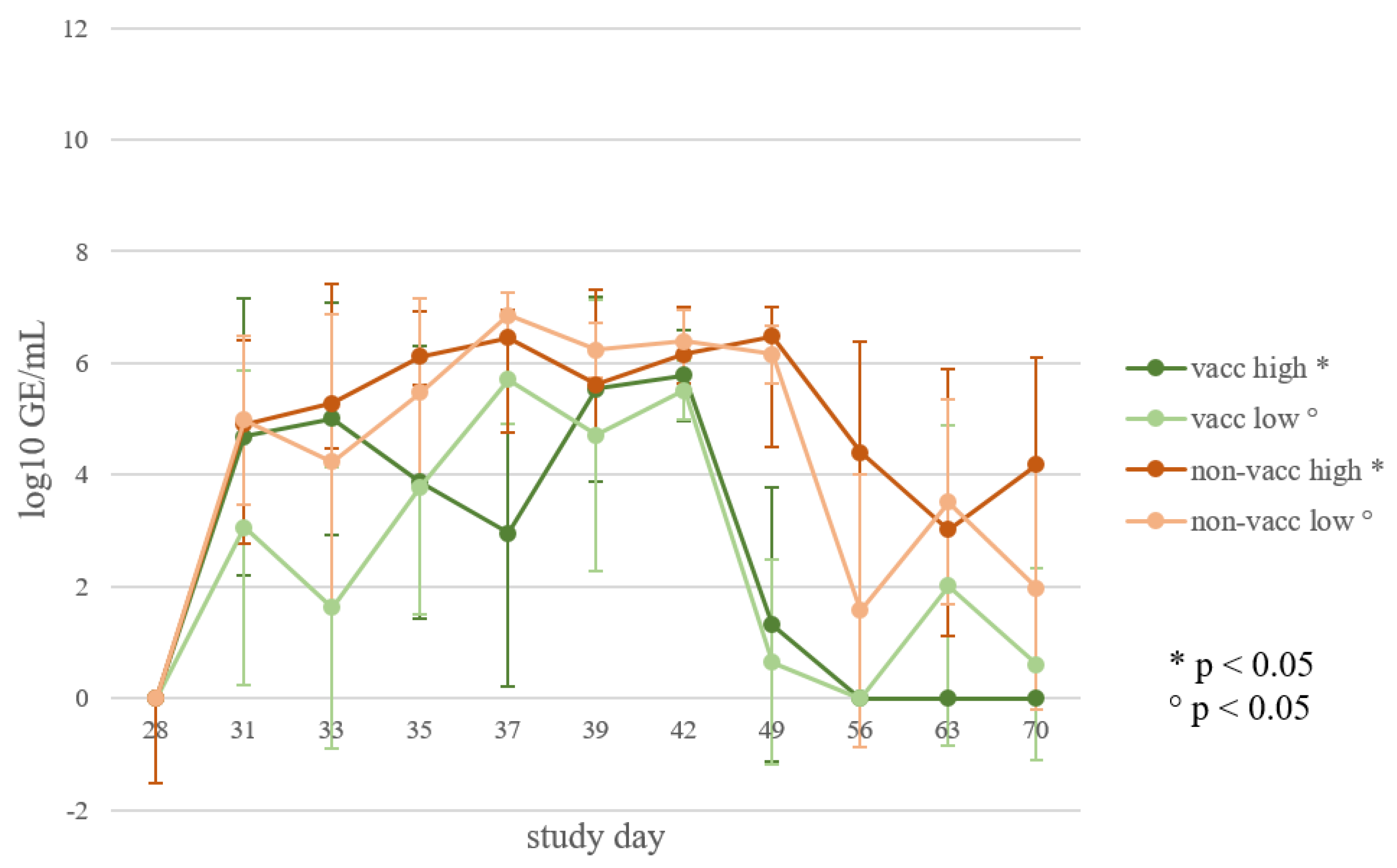
3.6. Viral Load in Oral Swabs
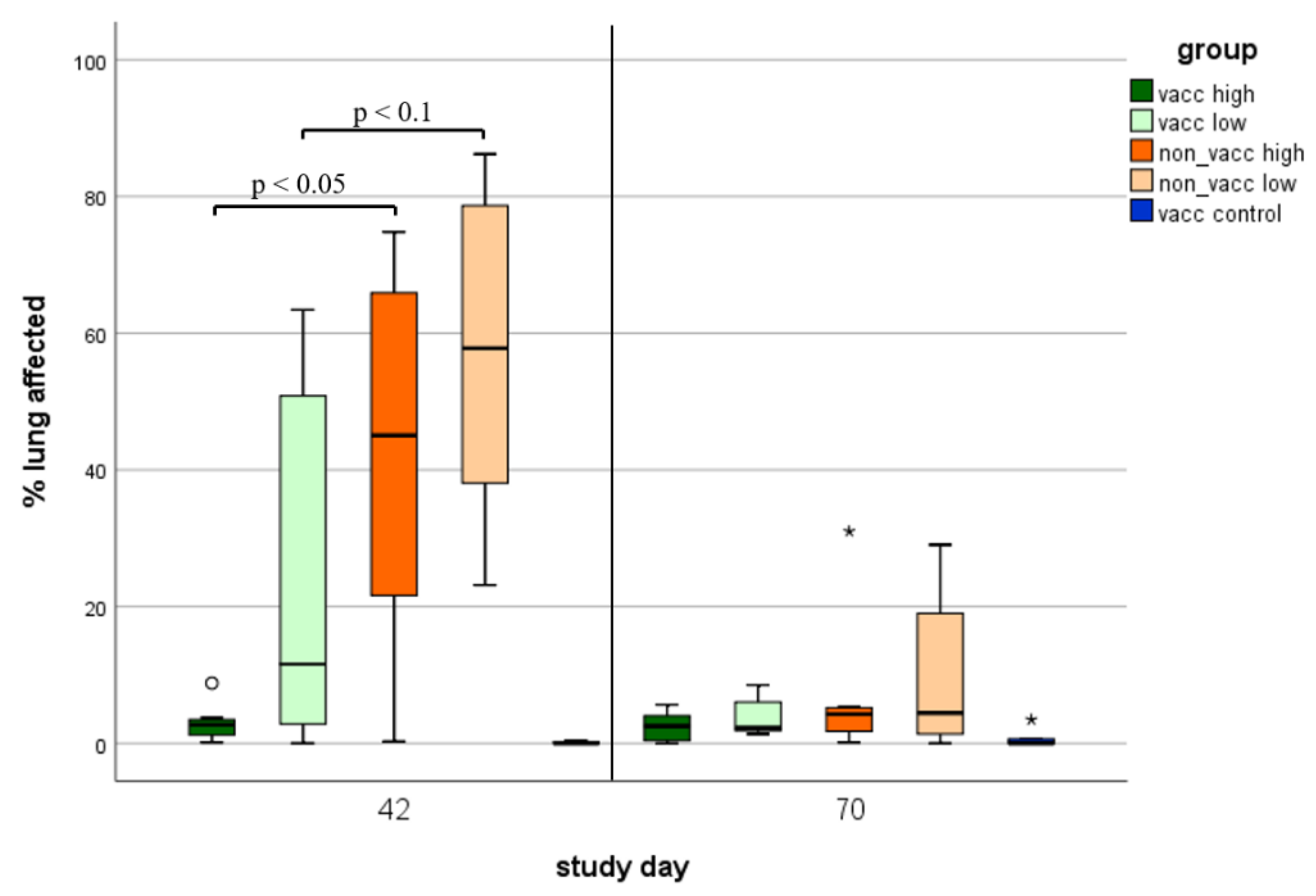
3.7. Macroscopic Lung Lesions
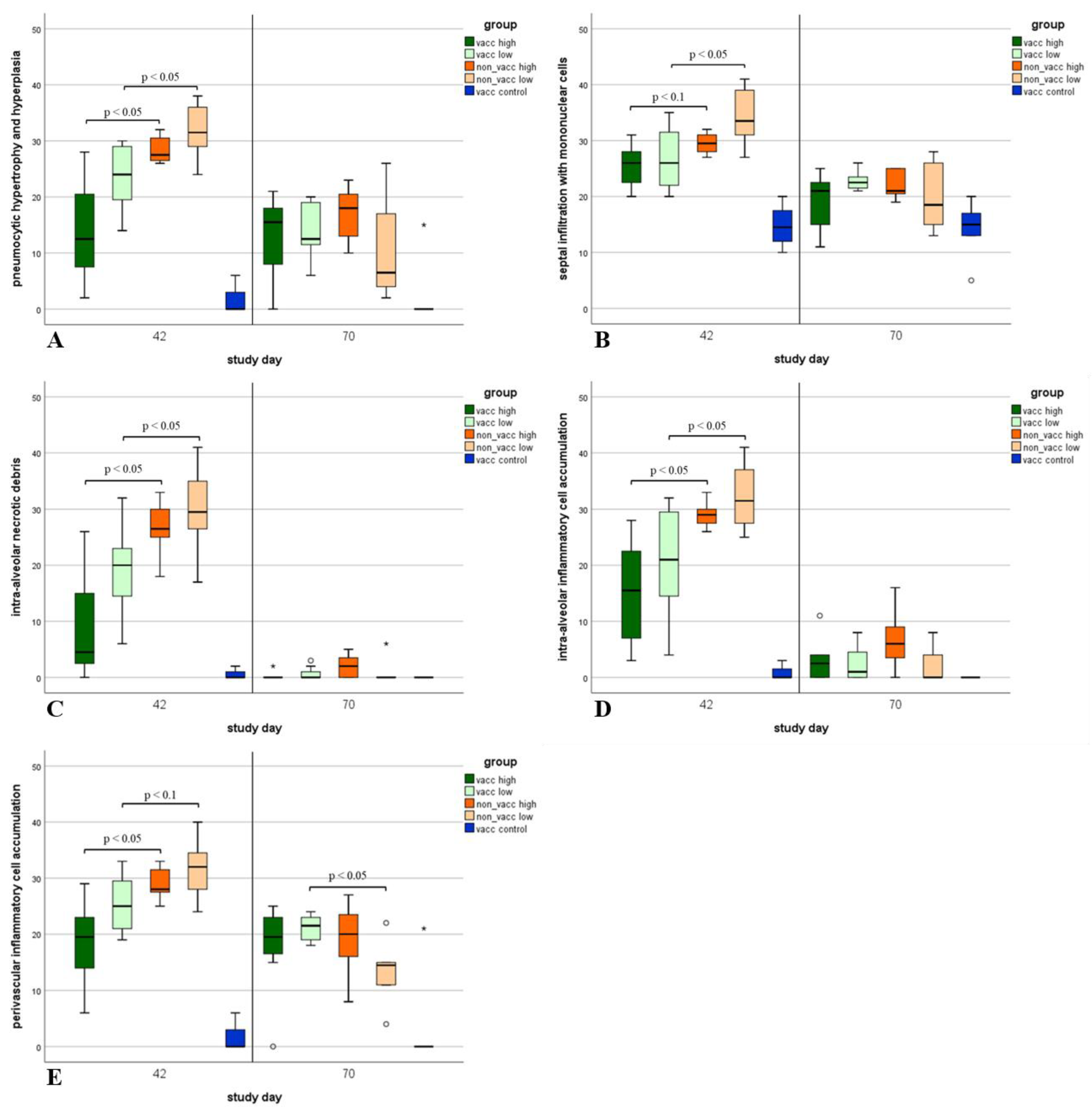
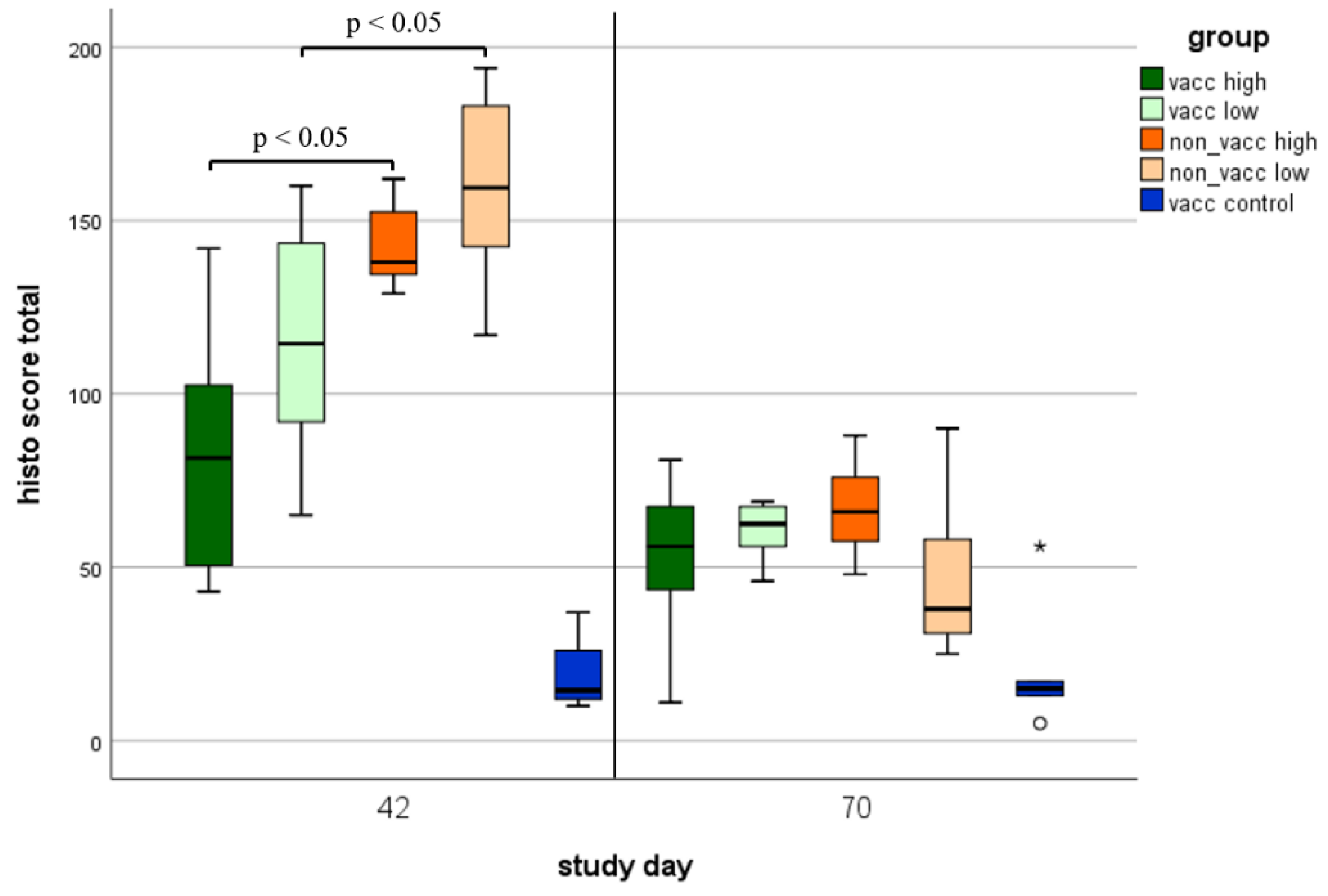
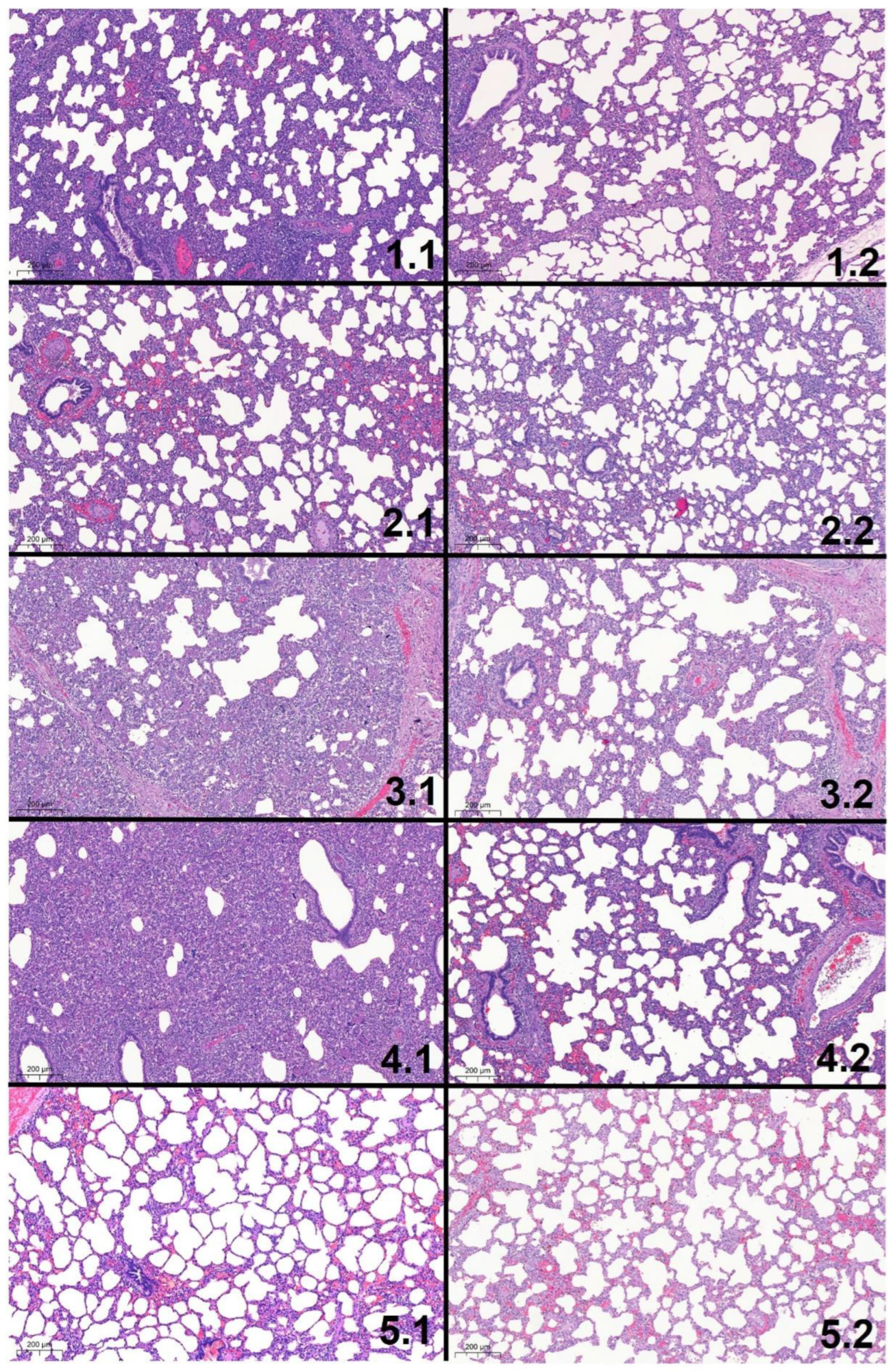
3.8. Histologic Lung Lesions
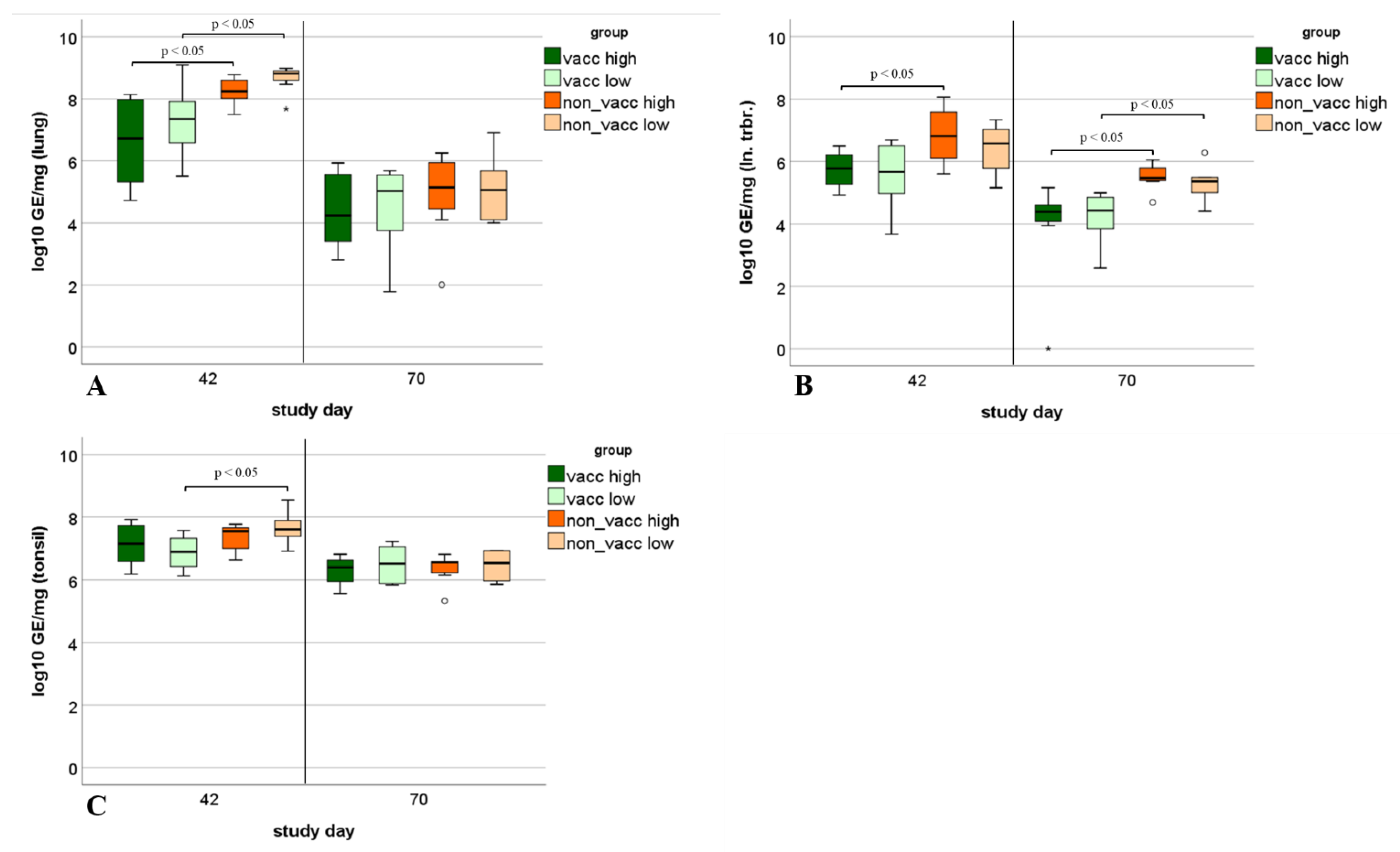
3.9. Viral Load in Tissue Samples
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Holtkamp, D.J.; Kliebenstein, J.B.; Neumann, E.J.; Zimmerman, J.; Rotto, H.F.; Yoder, T.K.; Wang, C.; Yeske, P.E.; Mowrer, C.L.; Haley, C.A. Assessment of the economic impact of porcine reproductive and respiratory syndrome virus on United States pork producers. J. Swine Health Prod. 2013, 21, 72–84. [Google Scholar]
- Benfield, D.A.; Nelson, E.; Collins, J.E.; Harris, L.; Goyal, S.M.; Robison, D.; Christianson, W.T.; Morrison, R.B.; Gorcyca, D.; Chladek, D. Characterization of swine infertility and respiratory syndrome (SIRS) virus (isolate ATCC VR-2332). J. Vet. Diagn. Investig. 1992, 4, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Cavanagh, D. Nidovirales: A new order comprising Coronaviridae and Arteriviridae. Arch. Virol. 1997, 142, 629–633. [Google Scholar] [PubMed]
- Adams, M.J.; Lefkowitz, E.J.; King, A.M.; Bamford, D.H.; Breitbart, M.; Davison, A.J.; Ghabrial, S.A.; Gorbalenya, A.E.; Knowles, N.J.; Krell, P.; et al. Ratification vote on taxonomic proposals to the International Committee on Taxonomy of Viruses (2015). Arch. Virol 2015, 160, 1837–1850. [Google Scholar] [CrossRef]
- Kuhn, J.H.; Lauck, M.; Bailey, A.L.; Shchetinin, A.M.; Vishnevskaya, T.V.; Bao, Y.; Ng, T.F.; LeBreton, M.; Schneider, B.S.; Gillis, A.; et al. Reorganization and expansion of the nidoviral family Arteriviridae. Arch. Virol. 2016, 161, 755–768. [Google Scholar] [CrossRef]
- Stadejek, T.; Oleksiewicz, M.B.; Scherbakov, A.V.; Timina, A.M.; Krabbe, J.S.; Chabros, K.; Potapchuk, D. Definition of subtypes in the European genotype of porcine reproductive and respiratory syndrome virus: Nucleocapsid characteristics and geographical distribution in Europe. Arch. Virol. 2008, 153, 1479–1488. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.; Hon, C.C.; Hui, R.K.; Faaberg, K.S.; Wennblom, T.; Murtaugh, M.P.; Stadejek, T.; Leung, F.C. Molecular epidemiology of PRRSV: A phylogenetic perspective. Virus Res. 2010, 154, 7–17. [Google Scholar] [CrossRef]
- Shi, M.; Lam, T.T.; Hon, C.C.; Murtaugh, M.P.; Davies, P.R.; Hui, R.K.; Li, J.; Wong, L.T.; Yip, C.W.; Jiang, J.W.; et al. Phylogeny-based evolutionary, demographical, and geographical dissection of North American type 2 porcine reproductive and respiratory syndrome viruses. J. Virol. 2010, 84, 8700–8711. [Google Scholar] [CrossRef]
- Magar, R.; Robinson, Y.; Dubuc, C.; Larochelle, R. Isolation and experimental oral transmission in pigs of a porcine reproductive and respiratory syndrome virus isolate. Adv. Exp. Med. Biol. 1995, 380, 139–144. [Google Scholar] [CrossRef]
- Van der Linden, I.F.; van der Linde-Bril, E.M.; Voermans, J.J.; van Rijn, P.A.; Pol, J.M.; Martin, R.; Steverink, P.J. Oral transmission of porcine reproductive and respiratory syndrome virus by muscle of experimentally infected pigs. Vet. Microbiol. 2003, 97, 45–54. [Google Scholar] [CrossRef]
- Christianson, W.T.; Choi, C.S.; Collins, J.E.; Molitor, T.W.; Morrison, R.B.; Joo, H.S. Pathogenesis of porcine reproductive and respiratory syndrome virus infection in mid-gestation sows and fetuses. Can. J. Vet. Res. 1993, 57, 262–268. [Google Scholar] [PubMed]
- Gradil, C.; Dubuc, C.; Eaglesome, M.D. Porcine reproductive and respiratory syndrome virus: Seminal transmission. Vet. Rec. 1996, 138, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Yaeger, M.J.; Prieve, T.; Collins, J.; Christopher-Hennings, J.; Nelson, E.; Benfield, D. Evidence for the transmission of porcine reproductive and respiratory syndrome (PRRS) virus in boar semen. Swine Health Prod. 1993, 1, 7–9. [Google Scholar]
- Keffaber, K.K. Reproductive failure of unknown etiology. Am. Assoc. Swine Pract. Newsl. 1989, 1, 1–10. [Google Scholar]
- Loula, T. Mystery pig disease. Agri-Practice 1991, 12, 23–34. [Google Scholar]
- Zimmerman, J.; Benfield, D.; Dee, S.A.; Murtaugh, M.P.; Stadejek, T.; Stevenson, G.W.; Torremorell, M. Porcine Reproductive and Respiratory Syndrome Virus (Porcine Arterivirus). In Diseases of Swine, 10th ed.; Zimmerman, J., Karriker, L., Ramirez, A., Schwartz, K., Stevenson, G.W., Eds.; John Wiley & Sons, Inc.: Chichester, UK, 2012; pp. 461–486. [Google Scholar]
- Zimmerman, J.J.; Yoon, K.J.; Wills, R.W.; Swenson, S.L. General overview of PRRSV: A perspective from the United States. Vet. Microbiol. 1997, 55, 187–196. [Google Scholar] [CrossRef]
- Corzo, C.A.; Mondaca, E.; Wayne, S.; Torremorell, M.; Dee, S.; Davies, P.; Morrison, R.B. Control and elimination of porcine reproductive and respiratory syndrome virus. Virus Res. 2010, 154, 185–192. [Google Scholar] [CrossRef]
- Charerntantanakul, W.; Platt, R.; Johnson, W.; Roof, M.; Vaughn, E.; Roth, J.A. Immune responses and protection by vaccine and various vaccine adjuvant candidates to virulent porcine reproductive and respiratory syndrome virus. Vet. Immunol. Immunopathol. 2006, 109, 99–115. [Google Scholar] [CrossRef]
- Linhares, D.C.; Cano, J.P.; Wetzell, T.; Nerem, J.; Torremorell, M.; Dee, S.A. Effect of modified-live porcine reproductive and respiratory syndrome virus (PRRSv) vaccine on the shedding of wild-type virus from an infected population of growing pigs. Vaccine 2012, 30, 407–413. [Google Scholar] [CrossRef]
- Sinn, L.J.; Klingler, E.; Lamp, B.; Brunthaler, R.; Weissenboeck, H.; Ruemenapf, T.; Ladinig, A. Emergence of a virulent porcine reproductive and respiratory syndrome virus (PRRSV) 1 strain in Lower Austria. Porc. Health Manag. 2016, 2, 28. [Google Scholar] [CrossRef]
- Kreutzmann, H.; Stadler, J.; Knecht, C.; Sassu, E.L.; Ruczizka, U.; Zablotski, Y.; Vatzia, E.; Balka, G.; Zaruba, M.; Chen, H.-W.; et al. Phenotypic characterization of a virulent PRRSV-1 isolate in a reproductive model with and without prior heterologous modified live PRRSV-1 vaccination. Front. Vet. Sci. 2022, 9, 820233. [Google Scholar] [CrossRef] [PubMed]
- Weiland, E.; Wieczorek-Krohmer, M.; Kohl, D.; Conzelmann, K.K.; Weiland, F. Monoclonal antibodies to the GP5 of porcine reproductive and respiratory syndrome virus are more effective in virus neutralization than monoclonal antibodies to the GP4. Vet. Microbiol. 1999, 66, 171–186. [Google Scholar] [CrossRef]
- Christensen, G.; Sørensen, V.; Mousing, J. Diseases of the respiratory system. In Diseases of Swine, 8th ed.; Straw, B.E., D’Allaire, S., Mengeling, W.L., Taylor, D.J., Eds.; Iowa State University Press: Ames, IA, USA, 1999; pp. 927–928. [Google Scholar]
- Balka, G.; Ladinig, A.; Ritzmann, M.; Saalmueller, A.; Gerner, W.; Kaeser, T.; Jakab, C.; Rusvai, M.; Weissenboeck, H. Immunohistochemical characterization of type II pneumocyte proliferation after challenge with type I porcine reproductive and respiratory syndrome virus. J. Comp. Pathol. 2013, 149, 322–330. [Google Scholar] [CrossRef] [PubMed]
- Egli, C.; Thur, B.; Liu, L.; Hofmann, M.A. Quantitative TaqMan RT-PCR for the detection and differentiation of European and North American strains of porcine reproductive and respiratory syndrome virus. J. Virol. Methods 2001, 98, 63–75. [Google Scholar] [CrossRef]
- Van Reeth, K.; Nauwynck, H.; Pensaert, M. Dual infections of feeder pigs with porcine reproductive and respiratory syndrome virus followed by porcine respiratory coronavirus or swine influenza virus: A clinical and virological study. Vet. Microbiol. 1996, 48, 325–335. [Google Scholar] [CrossRef]
- Done, S.H.; Paton, D.J. Porcine reproductive and respiratory syndrome: Clinical disease, pathology and immunosuppression. Vet. Rec. 1995, 136, 32–35. [Google Scholar] [CrossRef]
- Labarque, G.; van Reeth, K.; van Gucht, S.; Nauwynck, H.; Pensaert, M. Porcine reproductive-respiratory syndrome virus infection predisposes pigs for respiratory signs upon exposure to bacterial lipopolysaccharide. Vet. Microbiol. 2002, 88, 1–12. [Google Scholar] [CrossRef]
- Nodelijk, G.; de Jong, M.C.; van Leengoed, L.A.; Wensvoort, G.; Pol, J.M.; Steverink, P.J.; Verheijden, J.H. A quantitative assessment of the effectiveness of PRRSV vaccination in pigs under experimental conditions. Vaccine 2001, 19, 3636–3644. [Google Scholar] [CrossRef]
- Labarque, G.; van Gucht, S.; van Reeth, K.; Nauwynck, H.; Pensaert, M. Respiratory tract protection upon challenge of pigs vaccinated with attenuated porcine reproductive and respiratory syndrome virus vaccines. Vet. Microbiol. 2003, 95, 187–197. [Google Scholar] [CrossRef]
- Lyoo, K.S.; Choi, J.Y.; Hahn, T.W.; Park, K.T.; Kim, H.K. Effect of vaccination with a modified live porcine reproductive and respiratory syndrome virus vaccine on growth performance in fattening pigs under field conditions. J. Vet. Med. Sci. 2016, 78, 1533–1536. [Google Scholar] [CrossRef]
- Kritas, S.K.; Alexopoulos, C.; Kyriakis, C.S.; Tzika, E.; Kyriakis, S.C. Performance of fattening pigs in a farm infected with both porcine reproductive and respiratory syndrome (PRRS) virus and porcine circovirus type 2 following sow and piglet vaccination with an attenuated PRRS vaccine. J. Vet. Med. A Physiol. Pathol. Clin. Med. 2007, 54, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Jeong, J.; Kim, S.; Park, K.H.; Kang, I.; Park, S.J.; Yang, S.; Oh, T.; Chae, C. Vaccination with a porcine reproductive and respiratory syndrome virus vaccine at 1-day-old improved growth performance of piglets under field conditions. Vet. Microbiol. 2018, 214, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Charerntantanakul, W. Porcine reproductive and respiratory syndrome virus vaccines: Immunogenicity, efficacy and safety aspects. World J. Virol. 2012, 1, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Cano, J.P.; Dee, S.A.; Murtaugh, M.P.; Pijoan, C. Impact of a modified-live porcine reproductive and respiratory syndrome virus vaccine intervention on a population of pigs infected with a heterologous isolate. Vaccine 2007, 25, 4382–4391. [Google Scholar] [CrossRef]
- Lunney, J.K.; Fang, Y.; Ladinig, A.; Chen, N.; Li, Y.; Rowland, B.; Renukaradhya, G.J. Porcine Reproductive and Respiratory Syndrome Virus (PRRSV): Pathogenesis and Interaction with the Immune System. Annu. Rev. Anim. Biosci. 2016, 4, 129–154. [Google Scholar] [CrossRef]
- Guo, B.; Lager, K.M.; Henningson, J.N.; Miller, L.C.; Schlink, S.N.; Kappes, M.A.; Kehrli, M.E., Jr.; Brockmeier, S.L.; Nicholson, T.L.; Yang, H.C.; et al. Experimental infection of United States swine with a Chinese highly pathogenic strain of porcine reproductive and respiratory syndrome virus. Virology 2013, 435, 372–384. [Google Scholar] [CrossRef]
- Van der Linden, I.F.; Voermans, J.J.; van der Linde-Bril, E.M.; Bianchi, A.T.; Steverink, P.J. Virological kinetics and immunological responses to a porcine reproductive and respiratory syndrome virus infection of pigs at different ages. Vaccine 2003, 21, 1952–1957. [Google Scholar] [CrossRef]
- Cho, J.G.; Dee, S.A.; Deen, J.; Trincado, C.; Fano, E.; Jiang, Y.; Faaberg, K.; Murtaugh, M.P.; Guedes, A.; Collins, J.E.; et al. The impact of animal age, bacterial coinfection, and isolate pathogenicity on the shedding of porcine reproductive and respiratory syndrome virus in aerosols from experimentally infected pigs. Can. J. Vet. Res. 2006, 70, 297–301. [Google Scholar]
- Klinge, K.L.; Vaughn, E.M.; Roof, M.B.; Bautista, E.M.; Murtaugh, M.P. Age-dependent resistance to Porcine reproductive and respiratory syndrome virus replication in swine. Virol. J. 2009, 6, 177. [Google Scholar] [CrossRef]
- Balka, G.; Dreckmann, K.; Papp, G.; Kraft, C. Vaccination of piglets at 2 and 3 weeks of age with Ingelvac PRRSFLEX(R) EU provides protection against heterologous field challenge in the face of homologous maternally derived antibodies. Porc. Health Manag. 2016, 2, 24. [Google Scholar] [CrossRef]
- Cho, J.G.; Dee, S.A.; Deen, J.; Guedes, A.; Trincado, C.; Fano, E.; Jiang, Y.; Faaberg, K.; Collins, J.E.; Murtaugh, M.P.; et al. Evaluation of the effects of animal age, concurrent bacterial infection, and pathogenicity of porcine reproductive and respiratory syndrome virus on virus concentration in pigs. Am. J. Vet. Res. 2006, 67, 489–493. [Google Scholar] [CrossRef] [PubMed]
- Hermann, J.R.; Munoz-Zanzi, C.A.; Roof, M.B.; Burkhart, K.; Zimmerman, J.J. Probability of porcine reproductive and respiratory syndrome (PRRS) virus infection as a function of exposure route and dose. Vet. Microbiol. 2005, 110, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Pileri, E.; Gibert, E.; Soldevila, F.; Garcia-Saenz, A.; Pujols, J.; Diaz, I.; Darwich, L.; Casal, J.; Martin, M.; Mateu, E. Vaccination with a genotype 1 modified live vaccine against porcine reproductive and respiratory syndrome virus significantly reduces viremia, viral shedding and transmission of the virus in a quasi-natural experimental model. Vet. Microbiol. 2015, 175, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Mengeling, W.L.; Lager, K.M. A brief review of procedures and potential problems associated with the diagnosis of porcine reproductive and respiratory syndrome. Vet. Res. 2000, 31, 61–69. [Google Scholar] [CrossRef]
- Duan, X.; Nauwynck, H.J.; Pensaert, M.B. Effects of origin and state of differentiation and activation of monocytes/macrophages on their susceptibility to porcine reproductive and respiratory syndrome virus (PRRSV). Arch. Virol. 1997, 142, 2483–2497. [Google Scholar] [CrossRef]
- Allende, R.; Laegreid, W.W.; Kutish, G.F.; Galeota, J.A.; Wills, R.W.; Osorio, F.A. Porcine reproductive and respiratory syndrome virus: Description of persistence in individual pigs upon experimental infection. J. Virol. 2000, 74, 10834–10837. [Google Scholar] [CrossRef]
- Wills, R.W.; Doster, A.R.; Galeota, J.A.; Sur, J.H.; Osorio, F.A. Duration of infection and proportion of pigs persistently infected with porcine reproductive and respiratory syndrome virus. J. Clin. Microbiol. 2003, 41, 58–62. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. of Animals | Room | Treatment (Study Day 0) | PRRSV Challenge (Study Day 28) |
|---|---|---|---|---|
| 1 (vacc 1 high) | 16 | A | Vaccination | 105 TCID50 2/dose |
| 2 (vacc low) | 16 | A | Vaccination | 103 TCID50/dose |
| 3 (non-vacc high) | 16 | B | Sham treatment | 105 TCID50/dose |
| 4 (non-vacc low) | 16 | B | Sham treatment | 103 TCID50/dose |
| 5 (vacc control) | 10 | C | Vaccination | Sham inoculation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duerlinger, S.; Knecht, C.; Sawyer, S.; Balka, G.; Zaruba, M.; Ruemenapf, T.; Kraft, C.; Rathkjen, P.H.; Ladinig, A. Efficacy of a Modified Live Porcine Reproductive and Respiratory Syndrome Virus 1 (PRRSV-1) Vaccine against Experimental Infection with PRRSV AUT15-33 in Weaned Piglets. Vaccines 2022, 10, 934. https://doi.org/10.3390/vaccines10060934
Duerlinger S, Knecht C, Sawyer S, Balka G, Zaruba M, Ruemenapf T, Kraft C, Rathkjen PH, Ladinig A. Efficacy of a Modified Live Porcine Reproductive and Respiratory Syndrome Virus 1 (PRRSV-1) Vaccine against Experimental Infection with PRRSV AUT15-33 in Weaned Piglets. Vaccines. 2022; 10(6):934. https://doi.org/10.3390/vaccines10060934
Chicago/Turabian StyleDuerlinger, Sophie, Christian Knecht, Spencer Sawyer, Gyula Balka, Marianne Zaruba, Till Ruemenapf, Christian Kraft, Poul Henning Rathkjen, and Andrea Ladinig. 2022. "Efficacy of a Modified Live Porcine Reproductive and Respiratory Syndrome Virus 1 (PRRSV-1) Vaccine against Experimental Infection with PRRSV AUT15-33 in Weaned Piglets" Vaccines 10, no. 6: 934. https://doi.org/10.3390/vaccines10060934