
Biological Properties of SARS-CoV-2 Variants: Epidemiological Impact and Clinical Consequences



Abstract
:1. Introduction
Virus Transmission between Sick and Asymptomatic Individuals
2. Variants
2.1. D614G Variant
2.1.1. Transmissibility
2.1.2. Infectivity Rate
2.1.3. Disease Severity
2.1.4. Affinity to Angiotensin-Converting Enzyme 2 (ACE2) Receptors
2.1.5. Viral Load
2.1.6. Reproduction Number (R0/Rt)
2.1.7. Vaccine Effectiveness and Vaccine Breakthrough
2.2. Alpha Variant
2.2.1. Transmissibility
2.2.2. Infectivity Rate
2.2.3. Disease Severity
2.2.4. Affinity to Angiotensin-Converting Enzyme 2 (ACE2) Receptors
2.2.5. Viral Load
2.2.6. Reproduction Number (R0/Rt)
2.2.7. Vaccine Effectiveness and Vaccine Breakthrough
2.3. Beta Variant
2.3.1. Transmissibility
2.3.2. Infectivity Rate
2.3.3. Disease Severity
2.3.4. Affinity to Angiotensin-Converting Enzyme 2 (ACE2) Receptors
2.3.5. Viral Load
2.3.6. Reproduction Number (R0/Rt)
2.3.7. Vaccine Effectiveness and Vaccine Breakthrough
2.4. Gamma
2.4.1. Transmissibility
2.4.2. Infectivity Rate
2.4.3. Disease Severity
2.4.4. Affinity to Angiotensin-Converting Enzyme 2 (ACE2) Receptors
2.4.5. Viral Load
2.4.6. Reproduction Number (R0/Rt)
2.4.7. Vaccine Effectiveness and Vaccine Breakthrough
2.5. Delta
2.5.1. Transmissibility
2.5.2. Infectivity Rate
2.5.3. Disease Severity
2.5.4. Affinity to Angiotensin-Converting Enzyme 2 (ACE2) Receptors
2.5.5. Viral Load
2.5.6. Reproduction Number (R0/Rt)
2.5.7. Vaccine Effectiveness and Vaccine Breakthrough
2.6. Omicron
2.6.1. Transmissibility
2.6.2. Infectivity Rate
2.6.3. Disease Severity
2.6.4. Affinity to Angiotensin-Converting Enzyme 2 (ACE2) Receptors
2.6.5. Viral Load
2.6.6. Reproduction Number (R0/Rt)
2.6.7. Vaccine Effectiveness and Vaccine Breakthrough
3. Epidemiological Factors
3.1. Weather Conditions and Environmental Factors
3.1.1. Air Pollution
3.1.2. Temperature, Humidity, and Wind Speed
3.2. Indoor Settings
3.3. Population Density, Race and Life Condition
3.4. Age
3.4.1. Children
3.4.2. Adults and Older Age
3.5. Gender
4. COVID-19 Disease Clinical Treatment Methods
4.1. Oxygenation and Ventilation
4.2. Antiviral Therapies
4.3. Anti-SARS-CoV-2 Neutralizing Antibodies
4.4. Steroid Treatment
4.5. Immunomodulatory Agents
5. Transmission Rate of Other Coronaviruses
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

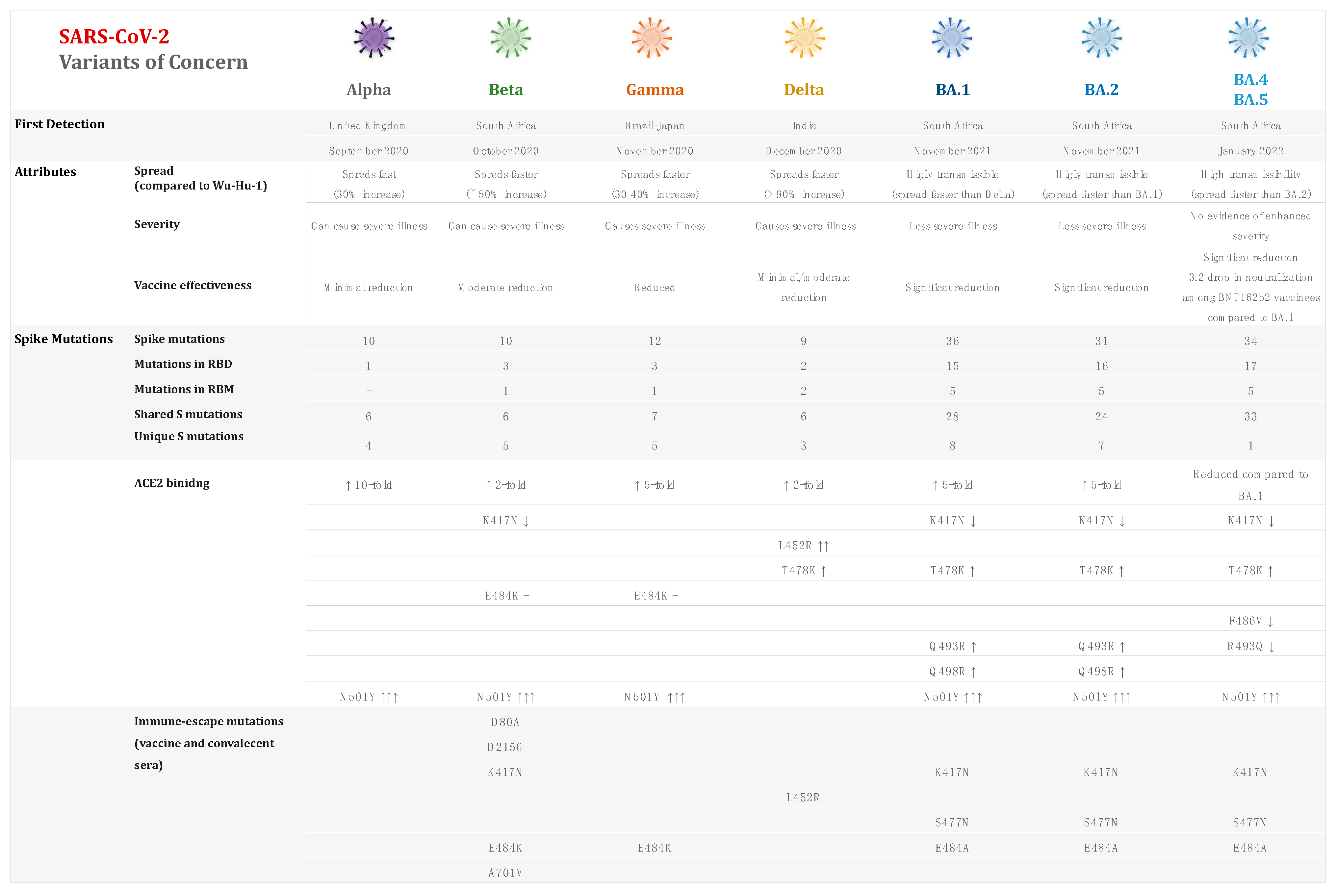{kind=link}
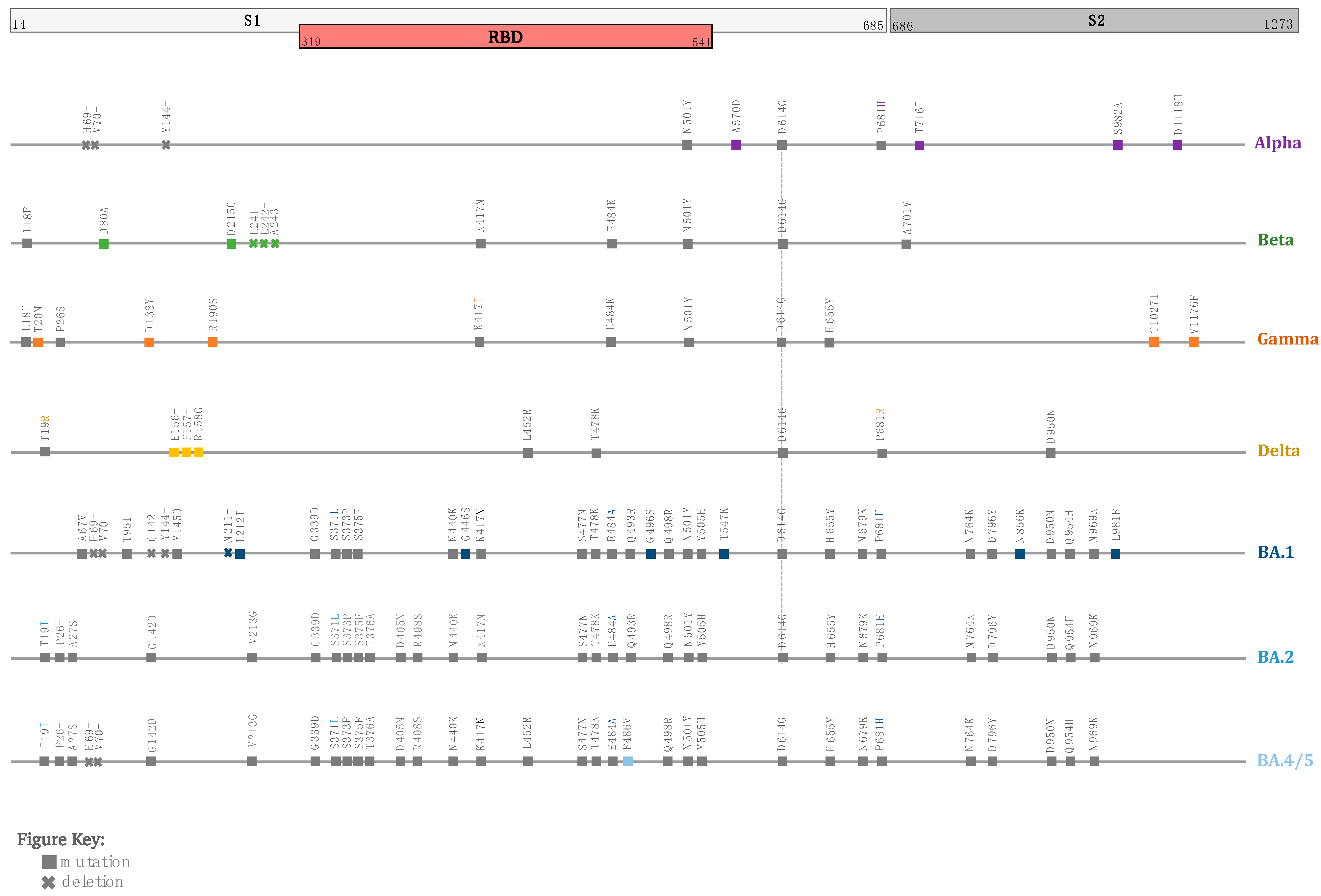{kind=link}
| SARS-CoV-2 Variant | Alpha | Beta | Gamma | Delta | Omicron |
|---|---|---|---|---|---|
| Scientific Name | B.1.1.7 | B.1.351 | P.1 | B.1.617.2 | B.1.1.529 |
| First reported (geographical location/date) | United Kingdom September 2020 | South Africa May 2020 | Brazil November 2020 | India October 2020 | Botswana, South Africa November 2021 |
| Number of mutations/spike mutation of interest | 23 mutations N501Y, D614G, P681H | 21 mutations K417N, E484K, N501Y, D614G, A701V | 17 mutations K417T, E484K, N501Y, D614G, H655Y | ∼23 mutations L452R, T478K, D614G, P681R | ∼50 mutations G339D, S371L S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, D614G, T547K, H655Y, N679K, P681R |
| Transmissibility | ∼50–70% [100,105,106] compared to the wild type ∼30–40% [100] Compared to other circulating lineages ∼43–90% [104,108] compared to proceeding variants. | ∼23–50% [157,158,159] compared to ancestral lineages | 1.4–2.2 [193,197] times compared to the wild type and 46% [197] compared to previous variants. | 1.4–2.0 [229,237] compared to other lineages. 60–70% [232,233] more transmissible than B.1.1.7. | 2.7–3.7 [306,307] times compared to the Delta variant. |
| Infectivity | 0.1% in early October to 49.7% in late November 2020 [63]. 3.7-fold rise in December 2020 [109]. | Reinfection cases were reported [163]. | 5.3% in the biweekly period of 11–24 April 2021 to 11.1% in the period of 23 May–5 June 2021 [202]. 0% in the period from November 2020 to 73% in January 2021 [203]. | 60% more infectious than the wild type [242]. 0.6% during April 2021 to 11.1% in May–June 2021, then surging to 83.2% in July 2021 [202]. | 13% to 73% in the period of 13–17 December 2021 [98]. >50% infections in mid-November 2021 [315]. 5.41-10-fold higher risk of reinfection than Delta [249,319]. Omicron is estimated to infect three to six times as many people as Delta over the same time [320]. |
| Disease severity | 1.5 to 1.7 [114,499] increased risk of hospitalization 62% higher risk of hospital admission compared to wild type [120]. 30% to 50% greater mortality rate [119,122] 1.64 mortality hazard ratio compared to preceding lineages [121]. | 2.16–3.6-fold higher risk of hospitalization and a 2.23–3.3-fold elevated risk of ICU admission [166,168]. 1.24-fold higher risk of progressing to severe disease than B.1.1.7 and 1.57-fold increased risk of mortality [170]. | 1.7–2.6-fold higher risk of hospitalization and a 2.06–2.2-fold higher risk of ICU admission [166,208,209]. | 2.26–2.83-fold higher risk of hospitalization compared to the B.1.1.7 variant [115,248,251]. 108–120% higher risk of hospitalization, a 235–287% higher risk of ICU admission, and a 133–137% higher risk of mortality [243,251]. | 50–70% less likely to be admitted to hospital than those infected with the Delta variant [323,360]. Two-thirds reduction in the risk of COVID-19 hospitalization when compared to Delta [321]. |
| Viral load/Ct values | The median Ct values for the ORF gene target (22.30 vs. 18.16; p < 0.0001) and N-gene target (23.16 vs. 19.39; p < 0.0001) were considerably lower in SGTF samples compared to non-SGTF samples [129]. Virus loads in SGTF samples can be 104 times higher than those in non-SGTF samples [129]. Viral load was higher in B.1.1.7 samples than in non-B.1.1.7 samples, as determined by cycle threshold value (mean 28.8, SD 4.7 vs. 32.0, 4.8) [123]. | Viral load at symptom onset was higher in B.1.351 variants than in historical variants, with a Ct value of −1.15 (−1.57, −0.697) lower than preceding strains [171]. | Viral load was ten times (Ct = 19.8 vs. 23.0; p < 0.0001) higher than in non-Gamma patients [197]. Decrease in the median Ct values (25 to 22) from nasopharyngeal swab samples examined by RT-qPCR (p < 0.0001) between June 2020 and February 2021 [211]. | Larger viral loads and a longer Ct ≤ 30 [246]. Much larger viral load than wild-type infections, with median Ct values for the N gene of the Delta variant of 23.0, which is significantly lower than the wild-type N gene’s values (median: 36.5) [9]. Viral levels up to 1260 times higher than those infected with the original strain [260]. | Viral RNA content in the lungs was 3 log10 lower than in animals infected with D614G [338]. Mean peak Ct value for Omicron was 23.3 vs. 20.5 for Delta, underlining that the lower the Ct value, the higher the peak viral load [339]. Lower infectious virus loads than Delta-infected patients (5.2-fold, 0.715 log10) [341]. |
| Estimated reproduction numbers (R0/Rt) | Rt = 1.25 during lockdown (UK) [132]. Rt increased 1.35 between September and December 2020, when compared to pre-existing variants (UK) [110]. R0 = 1–2 in November 2020 (UK) [104]. Rt = 1.4 in February 2021, then decreased to reach 1 in March, then 0.8 in April 2021 (Italy) [133]. R0 = 0.8–1.5 between January and March 2021 (Czech Republic) [134]. Rt > 2 between January and April 2021 (India) [136]. Rt = 1.44 between December 2020 and 3 February 2021 (Canada) [137]. Rt = 0.97 between August 2020 and January 2021, then increased > 1 from January 2021 to March 2021 [45], reaching 1.6 in January 2021 (Qatar) [45]. Rt varied from 1.1 to 2.8 (systematic review of ∼15 studies) [106]. | Rt = 1.55 (95% CI: 1.43–1.69) (based on data from multiple countries. including England, Wales, Scotland, Denmark, USA, and South Africa) until 12 February 2021 [138]. | Rt = 2.6 based on data collected before January 2021 (Brazil) [197]. Rt = 38% higher than non-VOCs, 10% higher than Alpha, and 17% higher than Beta (based on global data) until June 2021 [159]. | R0 = 3.2–5.0 between May and June 2021 (China) [239,265]. R0 = 5.08–6.0 in July 2021 (China) [266]. R0 = 5.2 during May 2021 (UK) [267]. | Rt = 0.8–2.5 in November 2021 (South Africa) [315,342]. Rt > 3 between November and December 2021 (UK) [319]. Rt was 4.2 times higher than the Delta variant (95% CI: 2.1, 9.1) during November 2021 (South Africa) [306]. R0 was equal to 1.90 (95% CI: 1.50–2.43) during November and December 2021 (South Korea) [343]. Rt = 3.19 (95% CI: 2.82–3.61) times more than that of Delta during December 2021 (Denmark) [344]. Rt = 2.5 during December 2021 (US) [345]. Rt = 1.8–3.1 in December 2021 (Italy) [346]. Rt = 1.34–3.57 depending on the location and vaccination rate, according to data obtained in many areas as of January 2022 (India) [377]. |
| Vaccine Effectiveness Pfizer (BNT162b2) AstraZeneca (ChAdOx1) Moderna (mRNA-1273) Novavax (NVX-CoV2373) Johnson & Johnson (Ad26.COV2.S) | Estimated reduction in neutralization by less than 2-fold for both Pfizer and Moderna vaccines [145,146,148], by 5- to <10-folds for AstraZeneca and between 2 and <5-fold for Johnson & Johnson [147]. Novavax was 85.6% effective in reducing symptomatic COVID-19 infection (UK) [139,141]. Moderna was 88% effective 14 days after the first dose and 100% ≥ 14 days after the second dose [140]. Pfizer was 83% effective in the overall population and 93% in SARS-CoV-2-experienced subjects (100-day cumulative incidence: 5.78%) (Italy) [150]. | Estimated reduction in neutralization by 5 to <10-fold for Pfizer, Moderna, and AstraZeneca vaccines and by more than 10-fold for Ad26.COV2.S [147]. Novavax revealed 89% efficacy (UK) and 60% efficacy (South Africa) [181,182]. Moderna and Pfizer exhibited 1.2-fold reduced nAb titers 3–4 weeks post-second dose (US) [183]. Moderna was 72% (US), 66% (Latin America), and 57% (South Africa) effective 28 days after immunization [181,182]. Moderna was 61% effective after the first dose and 96% after the second dose [140]. | Estimated reduction in neutralization by 2 to <5-fold for Pfizer, Moderna, AstraZeneca, and Johnson & Johnson vaccines [147]. The median pseudovirus neutralizing antibody titers generated by Ad26.COV2.S were 3.3-fold lower against the P.1 variant [185]. Neutralizing ability of plasma from people who had previously been infected was 8.6 times lower against the P.1 isolates [218]. AstraZeneca vaccination was 77.9% effective against infection, 87.6% effective against hospitalization, and 93.6% effective against death following the two-dose regimen (Brazil) [214]. | Estimated reduction in neutralization by 5 to <10-fold for Pfizer, by 2 to <5-folds for Moderna and AstraZeneca vaccines, and by less than 2-fold for Johnson & Johnson [149]. Delta was found to be more resistant to neutralization of convalescent serum (by a factor of 2) and vaccine serum (by a factor of 2.5–3.33) than the wild-type virus in his investigation [277]. In 95% of people, two doses produced a neutralizing response, with titers against Delta being 3-to-5 times lower than those against Alpha [276]. Both the Pfizer and AstraZeneca vaccines reduced the neutralizing titer for B.1.351 by eight to ninefold [178]. Pfizer and AstraZeneca vaccine have similar efficacies against the Delta variant, with 30.7% following a single dose (UK) [269]. Two AstraZeneca doses were less effective (67%) against Delta variant infection (UK) [269]. Pfizer was 45.3% effective ≥14 days after the first vaccine dose and 51.9% effective ≥14 days after the second dose (Qatar) [270]. Pfizer provided 95% protection against infection in people aged 16 and older following the two-dose regimen (data obtained from US, Argentina, Brazil, South Africa, Germany, and Turkey) [271]. Moderna was 73.7% and 73.1% effective ≥14 days after the first and second doses, respectively (Qatar) [270]. Moderna was found to be 94.1-96.8% effective in avoiding COVID-19 sickness, including severe disease (US) [273]. Gam-COVID-Vac (Sputnik V) was 91.6% effective (Moscow, Russia) [274]. | Estimated 20- to 40-fold reduction in neutralizing activity with two doses of Pfizer vaccine [362] and by more than 10-fold for both AstraZeneca and Moderna vaccines [361,362]. 22.9-fold higher neutralization resistance than the ancestral D614G 3–4 weeks after receiving a second dose of either mRNA-1273 or BNT162b2 (US) [183]. Three to eightfold reduction in neutralization titers for Omicron compared to Delta in a study conducted in South Africa [359]. Pfizer was 70% effective during the proxy Omicron period (South Africa) [356]. Pfizer and Moderna booster effectiveness was ∼50% (Qatar) [304]. Reduced vaccine effectiveness against infection to 33%, down from 80% against Delta (South Africa) [295]. Pfizer and Moderna third/booster vaccine doses were associated with a 57% reduction in the risk of symptomatic infection when compared to 25 weeks after the second dose [321]. Booster doses (3X) were found to provide a high level or protection, exceeding 80% under certain circumstances [317]. Omicron may be twice as likely to evade existing vaccinations compared to Delta [299]. |
References
- Chan, J.F.; Li, K.S.; To, K.K.; Cheng, V.C.; Chen, H.; Yuen, K.-Y. Is the discovery of the novel human betacoronavirus 2c EMC/2012 (HCoV-EMC) the beginning of another SARS-like pandemic? J. Infect. 2012, 65, 477–489. [Google Scholar] [CrossRef] [Green Version]
- Gallagher, T.M.; Buchmeier, M.J. Coronavirus spike proteins in viral entry and pathogenesis. Virology 2001, 279, 371–374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seah, I.; Agrawal, R. Can the coronavirus disease 2019 (COVID-19) affect the eyes? A review of coronaviruses and ocular implications in humans and animals. Ocul. Immunol. Inflamm. 2020, 28, 391–395. [Google Scholar] [CrossRef]
- Liu, J.; Xie, W.; Wang, Y.; Xiong, Y.; Chen, S.; Han, J.; Wu, Q. A comparative overview of COVID-19, MERS and SARS. Int. J. Surg. 2020, 81, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Singhal, T. A review of coronavirus disease-2019 (COVID-19). Indian J. Pediatr. 2020, 87, 281–286. [Google Scholar] [CrossRef] [Green Version]
- Sanches, P.R.; Charlie-Silva, I.; Braz, H.L.; Bittar, C.; Calmon, M.F.; Rahal, P.; Cilli, E.M. Recent advances in SARS-CoV-2 Spike protein and RBD mutations comparison between new variants Alpha (B. 1.1. 7, United Kingdom), Beta (B. 1.351, South Africa), Gamma (P. 1, Brazil) and Delta (B. 1.617. 2, India). J. Virus Erad. 2021, 7, 100054. [Google Scholar] [CrossRef]
- Ziegler, C.G.; Allon, S.J.; Nyquist, S.K.; Mbano, I.M.; Miao, V.N.; Tzouanas, C.N.; Cao, Y.; Yousif, A.S.; Bals, J.; Hauser, B.M. SARS-CoV-2 receptor ACE2 is an interferon-stimulated gene in human airway epithelial cells and is detected in specific cell subsets across tissues. Cell 2020, 181, 1016–1035.e19. [Google Scholar] [CrossRef]
- van Dorp, L.; Acman, M.; Richard, D.; Shaw, L.P.; Ford, C.E.; Ormond, L.; Owen, C.J.; Pang, J.; Tan, C.C.; Boshier, F.A. Emergence of genomic diversity and recurrent mutations in SARS-CoV-2. Infect. Genet. Evol. 2020, 83, 104351. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Peng, W.; Zhu, Y.; Lu, S.; Zhou, M.; Lin, W.; Wu, W.; Huang, S.; Jiang, L.; Luo, X. Recent progress in understanding 2019 novel coronavirus (SARS-CoV-2) associated with human respiratory disease: Detection, mechanisms and treatment. Int. J. Antimicrob. Agents 2020, 55, 105950. [Google Scholar] [CrossRef]
- Cevik, M.; Tate, M.; Lloyd, O.; Maraolo, A.E.; Schafers, J.; Ho, A. SARS-CoV-2, SARS-CoV, and MERS-CoV viral load dynamics, duration of viral shedding, and infectiousness: A systematic review and meta-analysis. Lancet Microb. 2021, 2, e13–e22. [Google Scholar] [CrossRef]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R. A novel coronavirus from patients with pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Hui, D.S.; Azhar, E.I.; Madani, T.A.; Ntoumi, F.; Kock, R.; Dar, O.; Ippolito, G.; Mchugh, T.D.; Memish, Z.A.; Drosten, C. The continuing 2019-nCoV epidemic threat of novel coronaviruses to global health—The latest 2019 novel coronavirus outbreak in Wuhan, China. Int. J. Infect. Dis. 2020, 91, 264–266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Lin, Q.; Jin, S.; You, L. Coronavirus 2019-nCoV: A Brief Perspective from the Front Line. J. Infect. 2020, 80, 373–377. [Google Scholar] [CrossRef] [Green Version]
- WHO. WHO Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 4 February 2022).
- Bamgboye, E.L.; Omiye, J.A.; Afolaranmi, O.J.; Davids, M.R.; Tannor, E.K.; Wadee, S.; Niang, A.; Were, A.; Naicker, S. COVID-19 pandemic: Is Africa different? J. Natl. Med. Assoc. 2021, 113, 324–335. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Cong, F.; Zeng, F.; Lian, Y.; Liu, X.; Luo, M.; Guo, P.; Ma, J. Development of a real time reverse transcription loop-mediated isothermal amplification method (RT-LAMP) for detection of a novel swine acute diarrhea syndrome coronavirus (SADS-CoV). J. Virol. Methods 2018, 260, 45–48. [Google Scholar] [CrossRef]
- Benzarti, M. Coronaviruses in farm animals: Epidemiology and public health implications. Vet. Med. Sci. 2020, 7, 322–347. [Google Scholar]
- Jia, Y.; Shen, G.; Nguyen, S.; Zhang, Y.; Huang, K.-S.; Ho, H.-Y.; Hor, W.-S.; Yang, C.-H.; Bruning, J.B.; Li, C. Analysis of the mutation dynamics of SARS-CoV-2 reveals the spread history and emergence of RBD mutant with lower ACE2 binding affinity. bioRxiv 2021. [Google Scholar] [CrossRef] [Green Version]
- Boni, M.F.; Lemey, P.; Jiang, X.; Lam, T.T.-Y.; Perry, B.W.; Castoe, T.A.; Rambaut, A.; Robertson, D.L. Evolutionary origins of the SARS-CoV-2 sarbecovirus lineage responsible for the COVID-19 pandemic. Nat. Microbiol. 2020, 5, 1408–1417. [Google Scholar] [CrossRef]
- Tonkin-Hill, G.; Martincorena, I.; Amato, R.; Lawson, A.R.; Gerstung, M.; Johnston, I.; Jackson, D.K.; Park, N.; Lensing, S.V.; Quail, M.A. Patterns of within-host genetic diversity in SARS-CoV-2. eLife 2021, 10, e66857. [Google Scholar] [CrossRef]
- Bar-On, Y.M.; Flamholz, A.; Phillips, R.; Milo, R. Science Forum: SARS-CoV-2 (COVID-19) by the numbers. eLife 2020, 9, e57309. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Li, H.; Wu, X.; Zhong, Y.; Zhang, K.; Zhang, Y.-P.; Boerwinkle, E.; Fu, Y.-X. Moderate mutation rate in the SARS coronavirus genome and its implications. BMC Evol. Biol. 2004, 4, 21. [Google Scholar] [CrossRef] [Green Version]
- Koyama, T.; Platt, D.; Parida, L. Variant analysis of SARS-CoV-2 genomes. Bull. World Health Organ. 2020, 98, 495. [Google Scholar] [CrossRef] [PubMed]
- Cotten, M.; Watson, S.J.; Zumla, A.I.; Makhdoom, H.Q.; Palser, A.L.; Ong, S.H.; Al Rabeeah, A.A.; Alhakeem, R.F.; Assiri, A.; Al-Tawfiq, J.A. Spread, circulation, and evolution of the Middle East respiratory syndrome coronavirus. MBio 2014, 5, e01062-13. [Google Scholar] [CrossRef] [Green Version]
- Ren, L.; Zhang, Y.; Li, J.; Xiao, Y.; Zhang, J.; Wang, Y.; Chen, L.; Paranhos-Baccalà, G.; Wang, J. Genetic drift of human coronavirus OC43 spike gene during adaptive evolution. Sci. Rep. 2015, 5, 11451. [Google Scholar] [CrossRef]
- Pyrc, K.; Dijkman, R.; Deng, L.; Jebbink, M.F.; Ross, H.A.; Berkhout, B.; Van der Hoek, L. Mosaic structure of human coronavirus NL63, one thousand years of evolution. J. Mol. Biol. 2006, 364, 964–973. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, S.; Govindarajulu, M.; Parise, R.S.; Neel, L.; Shankar, T.; Patel, S.; Lowery, P.; Smith, F.; Dhanasekaran, M.; Moore, T. Emerging SARS-CoV-2 variants: A review of its mutations, its implications and vaccine efficacy. Vaccines 2021, 9, 1195. [Google Scholar] [CrossRef] [PubMed]
- Olsen, R.J.; Christensen, P.A.; Long, S.W.; Subedi, S.; Hodjat, P.; Olson, R.; Nguyen, M.; Davis, J.J.; Yerramilli, P.; Saavedra, M.O. Trajectory of growth of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) variants in Houston, Texas, January through May 2021, based on 12,476 genome sequences. Am. J. Pathol. 2021, 191, 1754–1773. [Google Scholar] [CrossRef] [PubMed]
- Oran, D.P.; Topol, E.J. The proportion of SARS-CoV-2 infections that are asymptomatic: A systematic review. Ann. Intern. Med. 2021, 174, 655–662. [Google Scholar] [CrossRef]
- Team, E. The epidemiological characteristics of an outbreak of 2019 novel coronavirus diseases (COVID-19)—China, 2020. China CDC Wkly. 2020, 2, 113. [Google Scholar]
- WHO. Tracking SARS-CoV-2 Variants. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 31 January 2022).
- Parums, D.V. Revised World Health Organization (WHO) terminology for variants of concern and variants of interest of SARS-CoV-2. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2021, 27, e933622-21. [Google Scholar] [CrossRef] [PubMed]
- Rambaut, A.; Holmes, E.C.; O’Toole, Á.; Hill, V.; McCrone, J.T.; Ruis, C.; du Plessis, L.; Pybus, O.G. A dynamic nomenclature proposal for SARS-CoV-2 lineages to assist genomic epidemiology. Nat. Microbiol. 2020, 5, 1403–1407. [Google Scholar] [CrossRef]
- Wallinga, J.; Lipsitch, M. How generation intervals shape the relationship between growth rates and reproductive numbers. Proc. R. Soc. B Biol. Sci. 2007, 274, 599–604. [Google Scholar] [CrossRef] [Green Version]
- Ridenhour, B.; Kowalik, J.M.; Shay, D.K. Unraveling r 0: Considerations for public health applications. Am. J. Public Health 2018, 108, S445–S454. [Google Scholar] [CrossRef]
- Forsberg White, L.; Pagano, M. A likelihood-based method for real-time estimation of the serial interval and reproductive number of an epidemic. Stat. Med. 2008, 27, 2999–3016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Healthknowledge. Epidemic Theory (Effective & Basic Reproduction Numbers, Epidemic Thresholds) & Techniques for Analysis of Infectious Disease Data (Construction & Use of Epidemic Curves, Generation Numbers, Exceptional Reporting & Identification of Significant Clusters). Available online: https://www.healthknowledge.org.uk/public-health-textbook/research-methods/1a-epidemiology/epidemic-theory (accessed on 20 February 2022).
- Yuan, J.; Li, M.; Lv, G.; Lu, Z.K. Monitoring transmissibility and mortality of COVID-19 in Europe. Int. J. Infect. Dis. 2020, 95, 311–315. [Google Scholar] [CrossRef]
- Bacaër, N.; Gomes, M.G.M. On the final size of epidemics with seasonality. Bull. Math. Biol. 2009, 71, 1954. [Google Scholar] [CrossRef] [Green Version]
- Chen, J. Pathogenicity and transmissibility of 2019-nCoV—A quick overview and comparison with other emerging viruses. Microb. Infect. 2020, 22, 69–71. [Google Scholar] [CrossRef] [PubMed]
- Gostic, K.M.; McGough, L.; Baskerville, E.B.; Abbott, S.; Joshi, K.; Tedijanto, C.; Kahn, R.; Niehus, R.; Hay, J.A.; De Salazar, P.M. Practical considerations for measuring the effective reproductive number, Rt. PLoS Comput. Biol. 2020, 16, e1008409. [Google Scholar] [CrossRef] [PubMed]
- Ayoub, H.H.; Chemaitelly, H.; Seedat, S.; Makhoul, M.; Al Kanaani, Z.; Al Khal, A.; Al Kuwari, E.; Butt, A.A.; Coyle, P.; Jeremijenko, A. Mathematical modeling of the SARS-CoV-2 epidemic in Qatar and its impact on the national response to COVID-19. J. Glob. Health 2021, 11, 05005. [Google Scholar] [CrossRef]
- Khailaie, S.; Mitra, T.; Bandyopadhyay, A.; Schips, M.; Mascheroni, P.; Vanella, P.; Lange, B.; Binder, S.C.; Meyer-Hermann, M. Development of the reproduction number from coronavirus SARS-CoV-2 case data in Germany and implications for political measures. BMC Med. 2021, 19, 32. [Google Scholar] [CrossRef]
- Annunziato, A.; Asikainen, T. Effective reproduction number estimation from data series. JRC121343 2020, pra (VA), Italy. Available online: https://data.europa.eu/doi/10.2760/036156 (accessed on 31 March 2022).
- Abu-Raddad, L.J.; Chemaitelly, H.; Ayoub, H.H.; Coyle, P.; Malek, J.A.; Ahmed, A.A.; Mohamoud, Y.A.; Younuskunju, S.; Tang, P.; Al Kanaani, Z. Introduction and expansion of the SARS-CoV-2 B. 1.1. 7 variant and reinfections in Qatar: A nationally representative cohort study. PLoS Med. 2021, 18, e1003879. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.-Y.; Zhang, Y.; Peng, L.-Q.; Gao, R.-R.; Jing, J.-R.; Wang, J.-L.; Ren, B.-Z.; Xu, J.-G.; Wang, T. Demand for longer quarantine period among common and uncommon COVID-19 infections: A scoping review. Infect. Dis. Poverty 2021, 10, 5–13. [Google Scholar] [CrossRef] [PubMed]
- Chun, J.Y.; Baek, G.; Kim, Y. Transmission onset distribution of COVID-19. Int. J. Infect. Dis. 2020, 99, 403–407. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Zhang, G.; Yu, H.; Wang, J.; Wang, S.; Tang, G.; Guo, C.; Li, J.; Wei, S.; Wang, C. The epidemiologic and clinical features of suspected and confirmed cases of imported 2019 novel coronavirus pneumonia in north Shanghai, China. Ann. Transl. Med. 2020, 8, 637. [Google Scholar] [CrossRef]
- CDC. Post-COVID Conditions. Available online: https://www.cdc.gov/coronavirus/2019-ncov/long-term-effects/ (accessed on 21 February 2022).
- Healthline. Omicron and Long COVID: What We Know So Far. Available online: https://www.healthline.com/health-news/omicron-and-long-covid-what-we-know-so-far (accessed on 21 February 2022).
- McCall, B. Different Variants May Cause Different Long COVID Symptoms: Study. Available online: https://www.medscape.com/viewarticle/970982?reg=1 (accessed on 10 May 2022).
- Taquet, M. Over a Third of COVID-19 Patients Diagnosed with at Least One Long-COVID Symptom. Available online: https://www.ox.ac.uk/news/2021-09-29-over-third-covid-19-patients-diagnosed-least-one-long-covid-symptom (accessed on 21 February 2022).
- Group, S.C.-G. Genomewide association study of severe COVID-19 with respiratory failure. N. Engl. J. Med. 2020, 383, 1522–1534. [Google Scholar]
- Pairo-Castineira, E.; Clohisey, S.; Klaric, L.; Bretherick, A.D.; Rawlik, K.; Pasko, D.; Walker, S.; Parkinson, N.; Fourman, M.H.; Russell, C.D. Genetic mechanisms of critical illness in COVID-19. Nature 2021, 591, 92–98. [Google Scholar] [CrossRef]
- Zhang, Q.; Bastard, P.; Liu, Z.; Le Pen, J.; Moncada-Velez, M.; Chen, J.; Ogishi, M.; Sabli, I.K.; Hodeib, S.; Korol, C. Inborn errors of type I IFN immunity in patients with life-threatening COVID-19. Science 2020, 370, eabd4570. [Google Scholar] [CrossRef] [PubMed]
- Asgari, S.; Pousaz, L.A. Human Genetic Variants Identified That Affect COVID Susceptibility and Severity; Nature Publishing Group: New York, NY, USA, 2021. [Google Scholar]
- Zhang, M.; Liang, Y.; Yu, D.; Du, B.; Cheng, W.; Li, L.; Yu, Z.; Luo, S.; Zhang, Y.; Wang, H. A systematic review of Vaccine Breakthrough Infections by SARS-CoV-2 Delta Variant. Int. J. Biol. Sci. 2022, 18, 889. [Google Scholar] [CrossRef] [PubMed]
- Noori, M.; Nejadghaderi, S.A.; Rezaei, N. “Original antigenic sin”: A potential threat beyond the development of booster vaccination against novel SARS-CoV-2 variants. Infect. Control Hosp. Epidemiol. 2021, 1–2. [Google Scholar] [CrossRef]
- Forchette, L.; Sebastian, W.; Liu, T. A comprehensive review of COVID-19 virology, vaccines, variants, and therapeutics. Curr. Med. Sci. 2021, 41, 1037–1051. [Google Scholar] [CrossRef]
- Noor, R. Developmental Status of the Potential Vaccines for the Mitigation of the COVID-19 Pandemic and a Focus on the Effectiveness of the Pfizer-BioNTech and Moderna mRNA Vaccines. Curr. Clin. Microbiol. Rep. 2021, 8, 178–185. [Google Scholar] [CrossRef]
- Fan, Y.-J.; Chan, K.-H.; Hung, I.F.-N. Safety and Efficacy of COVID-19 Vaccines: A Systematic Review and Meta-Analysis of Different Vaccines at Phase 3. Vaccines 2021, 9, 989. [Google Scholar] [CrossRef]
- Abebe, E.C.; Dejenie, T.A.; Shiferaw, M.Y.; Malik, T. The newly emerged COVID-19 disease: A systemic review. Virol. J. 2020, 17, 96. [Google Scholar] [CrossRef]
- Leung, K.; Shum, M.H.; Leung, G.M.; Lam, T.T.; Wu, J.T. Early transmissibility assessment of the N501Y mutant strains of SARS-CoV-2 in the United Kingdom, October to November 2020. Eurosurveillance 2021, 26, 2002106. [Google Scholar] [CrossRef]
- Vardoulakis, S.; Oyarce, D.A.E.; Donner, E. Transmission of COVID-19 and other infectious diseases in public washrooms: A systematic review. Sci. Total Environ. 2022, 803, 149932. [Google Scholar] [CrossRef] [PubMed]
- Triggle, C.R.; Bansal, D.; Ding, H.; Islam, M.M.; Farag, E.A.B.A.; Hadi, H.A.; Sultan, A.A. A comprehensive review of viral characteristics, transmission, pathophysiology, immune response, and management of SARS-CoV-2 and COVID-19 as a basis for controlling the pandemic. Front. Immunol. 2021, 12, 338. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Thambiraja, T.S.; Karuppanan, K.; Subramaniam, G. Omicron and Delta Variant of SARS-CoV-2: A Comparative Computational Study of Spike Protein. J. Med. Virol. 2021, 94, 1641–1649. [Google Scholar] [CrossRef] [PubMed]
- Thakur, V.; Thakur, P.; Ratho, R.K. Evolutionary Dynamics of the Emergent SARS-Cov-2 Variants: Just Within a Year of Circulation. Immunome Res. 2021, 17, 1–4. [Google Scholar]
- Dudas, G.; Hong, S.L.; Potter, B.I.; Calvignac-Spencer, S.; Niatou-Singa, F.S.; Tombolomako, T.B.; Fuh-Neba, T.; Vickos, U.; Ulrich, M.; Leendertz, F.H. Emergence and spread of SARS-CoV-2 lineage B. 1.620 with variant of concern-like mutations and deletions. Nat. Commun. 2021, 12, 5769. [Google Scholar] [CrossRef]
- Korber, B.; Fischer, W.M.; Gnanakaran, S.; Yoon, H.; Theiler, J.; Abfalterer, W.; Hengartner, N.; Giorgi, E.E.; Bhattacharya, T.; Foley, B. Tracking changes in SARS-CoV-2 spike: Evidence that D614G increases infectivity of the COVID-19 virus. Cell 2020, 182, 812–827.e19. [Google Scholar] [CrossRef]
- Hu, J.; He, C.L.; Gao, Q.; Zhang, G.J.; Cao, X.X.; Long, Q.X.; Deng, H.J.; Huang, L.Y.; Chen, J.; Wang, K. The D614G mutation of SARS-CoV-2 spike protein enhances viral infectivity. bioRxiv 2020. [Google Scholar] [CrossRef]
- Volz, E.; Hill, V.; McCrone, J.T.; Price, A.; Jorgensen, D.; O’Toole, Á.; Southgate, J.; Johnson, R.; Jackson, B.; Nascimento, F.F. Evaluating the effects of SARS-CoV-2 spike mutation D614G on transmissibility and pathogenicity. Cell 2021, 184, 64–75.e11. [Google Scholar] [CrossRef]
- Hou, Y.J.; Chiba, S.; Halfmann, P.; Ehre, C.; Kuroda, M.; Dinnon, K.H.; Leist, S.R.; Schäfer, A.; Nakajima, N.; Takahashi, K. SARS-CoV-2 D614G variant exhibits efficient replication ex vivo and transmission in vivo. Science 2020, 370, 1464–1468. [Google Scholar] [CrossRef]
- Plante, J.A.; Liu, Y.; Liu, J.; Xia, H.; Johnson, B.A.; Lokugamage, K.G.; Zhang, X.; Muruato, A.E.; Zou, J.; Fontes-Garfias, C.R. Spike mutation D614G alters SARS-CoV-2 fitness. Nature 2021, 592, 116–121. [Google Scholar] [CrossRef]
- Daniloski, Z.; Jordan, T.X.; Ilmain, J.K.; Guo, X.; Bhabha, G.; Sanjana, N.E. The Spike D614G mutation increases SARS-CoV-2 infection of multiple human cell types. eLife 2021, 10, e65365. [Google Scholar] [CrossRef]
- Ogawa, J.; Zhu, W.; Tonnu, N.; Singer, O.; Hunter, T.; Ryan, A.L.; Pao, G.M. The D614G mutation in the SARS-CoV2 Spike protein increases infectivity in an ACE2 receptor dependent manner. bioRxiv 2020. [Google Scholar] [CrossRef]
- Pandey, U.; Yee, R.; Shen, L.; Judkins, A.R.; Bootwalla, M.; Ryutov, A.; Maglinte, D.T.; Ostrow, D.; Precit, M.; Biegel, J.A. High prevalence of SARS-CoV-2 genetic variation and D614G mutation in pediatric patients with COVID-19. In Open Forum Infectious Diseases; Oxford University Press: Oxford, UK, 2020; Volume 8, p. ofaa551. [Google Scholar]
- Yurkovetskiy, L.; Wang, X.; Pascal, K.E.; Tomkins-Tinch, C.; Nyalile, T.P.; Wang, Y.; Baum, A.; Diehl, W.E.; Dauphin, A.; Carbone, C. Structural and functional analysis of the D614G SARS-CoV-2 spike protein variant. Cell 2020, 183, 739–751.e8. [Google Scholar] [CrossRef]
- Zhang, L.; Jackson, C.B.; Mou, H.; Ojha, A.; Peng, H.; Quinlan, B.D.; Rangarajan, E.S.; Pan, A.; Vanderheiden, A.; Suthar, M.S. SARS-CoV-2 spike-protein D614G mutation increases virion spike density and infectivity. Nat. Commun. 2020, 11, 6013. [Google Scholar] [CrossRef]
- Ozono, S.; Zhang, Y.; Ode, H.; Sano, K.; Tan, T.S.; Imai, K.; Miyoshi, K.; Kishigami, S.; Ueno, T.; Iwatani, Y. SARS-CoV-2 D614G spike mutation increases entry efficiency with enhanced ACE2-binding affinity. Nat. Commun. 2021, 12, 848. [Google Scholar] [CrossRef]
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Eurosurveillance 2020, 25, 2000045. [Google Scholar] [CrossRef] [Green Version]
- Raghav, S.; Ghosh, A.; Turuk, J.; Kumar, S.; Jha, A.; Madhulika, S.; Priyadarshini, M.; Biswas, V.K.; Shyamli, P.S.; Singh, B. Analysis of Indian SARS-CoV-2 genomes reveals prevalence of D614G mutation in spike protein predicting an increase in interaction with TMPRSS2 and virus infectivity. Front. Microbiol. 2020, 11, 594928. [Google Scholar] [CrossRef] [PubMed]
- Groves, D.C.; Rowland-Jones, S.L.; Angyal, A. The D614G mutations in the SARS-CoV-2 spike protein: Implications for viral infectivity, disease severity and vaccine design. Biochem. Biophys. Res. Commun. 2021, 538, 104–107. [Google Scholar] [CrossRef]
- Mansbach, R.A.; Chakraborty, S.; Nguyen, K.; Montefiori, D.C.; Korber, B.; Gnanakaran, S. The SARS-CoV-2 spike variant D614G favors an open conformational state. Sci. Adv. 2021, 7, eabf3671. [Google Scholar] [CrossRef]
- Lorenzo-Redondo, R.; Nam, H.H.; Roberts, S.C.; Simons, L.M.; Jennings, L.J.; Qi, C.; Achenbach, C.J.; Hauser, A.R.; Ison, M.G.; Hultquist, J.F. A unique clade of SARS-CoV-2 viruses is associated with lower viral loads in patient upper airways. medRxiv 2020. [Google Scholar] [CrossRef]
- WHO. Statement on the Second Meeting of the International Health Regulations. Emergency Committee Regarding the Outbreak of Novel Coronavirus (2019-nCoV). 2005. Available online: https://www.who.int/news/item/30-01-2020-statement-on-the-second-meeting-of-the-international-health-regulations-(2005)-emergency-committee-regarding-the-outbreak-of-novel-coronavirus-(2019-ncov) (accessed on 27 February 2022).
- Liu, Y.; Gayle, A.A.; Wilder-Smith, A.; Rocklöv, J. The reproductive number of COVID-19 is higher compared to SARS coronavirus. J. Travel Med. 2020, 13, taaa021. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Bi, Y.; Gao, G.F. Epidemiological model suggests D614G spike protein mutation accelerates transmission of COVID-19—worldwide, 2020. China CDC Wkly. 2020, 2, 94. [Google Scholar] [CrossRef]
- Khan, A.; Zia, T.; Suleman, M.; Khan, T.; Ali, S.S.; Abbasi, A.A.; Mohammad, A.; Wei, D.Q. Higher infectivity of the SARS-CoV-2 new variants is associated with K417N/T, E484K, and N501Y mutants: An insight from structural data. J. Cell. Physiol. 2021, 236, 7045–7057. [Google Scholar] [CrossRef]
- Islam, S.R.; Prusty, D.; Manna, S.K. Structural basis of fitness of emerging SARS-COV-2 variants and considerations for screening, testing and surveillance strategy to contain their threat. medRxiv 2021. [Google Scholar] [CrossRef]
- Bartsch, S.M.; O’Shea, K.J.; Ferguson, M.C.; Bottazzi, M.E.; Wedlock, P.T.; Strych, U.; McKinnell, J.A.; Siegmund, S.S.; Cox, S.N.; Hotez, P.J. Vaccine efficacy needed for a COVID-19 coronavirus vaccine to prevent or stop an epidemic as the sole intervention. Am. J. Prev. Med. 2020, 59, 493–503. [Google Scholar] [CrossRef]
- Krause, P.; Fleming, T.R.; Longini, I.; Henao-Restrepo, A.M.; Peto, R.; Dean, N.; Halloran, M.; Huang, Y.; Fleming, T.; Gilbert, P. COVID-19 vaccine trials should seek worthwhile efficacy. Lancet 2020, 396, 741–743. [Google Scholar] [CrossRef]
- Leung, K.; Pei, Y.; Leung, G.M.; Lam, T.T.; Wu, J.T. Estimating the transmission advantage of the D614G mutant strain of SARS-CoV-2, December 2019 to June 2020. Eurosurveillance 2021, 26, 2002005. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. COVID-19: Death rate is 0.66% and increases with age, study estimates. BMJ 2020, 369, m1327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakraborty, C.; Bhattacharya, M.; Sharma, A.R. Present variants of concern and variants of interest of severe acute respiratory syndrome coronavirus 2: Their significant mutations in S-glycoprotein, infectivity, re-infectivity, immune escape and vaccines activity. Rev. Med. Virol. 2021, 32, e2270. [Google Scholar] [CrossRef]
- Akkiz, H. Implications of the Novel Mutations in the SARS-CoV-2 Genome for Transmission, Disease Severity, and the Vaccine Development. Front. Med. 2021, 8, 636532. [Google Scholar] [CrossRef]
- Thorne, L.G.; Bouhaddou, M.; Reuschl, A.-K.; Zuliani-Alvarez, L.; Polacco, B.; Pelin, A.; Batra, J.; Whelan, M.V.; Ummadi, M.; Rojc, A. Evolution of enhanced innate immune evasion by the SARS-CoV-2 B. 1.1. 7 UK variant. bioRxiv 2021. [Google Scholar] [CrossRef]
- Volz, E.; Mishra, S.; Chand, M.; Barrett, J.C.; Johnson, R.; Geidelberg, L.; Hinsley, W.R.; Laydon, D.J.; Dabrera, G.; O’Toole, Á. Assessing transmissibility of SARS-CoV-2 lineage B. 1.1. 7 in England. Nature 2021, 593, 266–269. [Google Scholar] [CrossRef]
- CDC. US COVID-19 Cases Caused by Variants. Available online: https://stacks.cdc.gov/view/cdc/104580 (accessed on 12 January 2022).
- GVN. Alpha (B.1.1.7). Available online: https://gvn.org/covid-19/alpha-b-1-1-7/ (accessed on 22 February 2022).
- Mahase, E. COVID-19: How many variants are there, and what do we know about them? BMJ 2021, 374, n1971. [Google Scholar] [CrossRef]
- Yang, W.; Shaman, J. Development of a model-inference system for estimating epidemiological characteristics of SARS-CoV-2 variants of concern. Nat. Commun. 2021, 12, 5573. [Google Scholar] [CrossRef]
- Lyngse, F.P.; Mølbak, K.; Skov, R.L.; Christiansen, L.E.; Mortensen, L.H.; Albertsen, M.; Møller, C.H.; Krause, T.G.; Rasmussen, M.; Michaelsen, T.Y. Increased transmissibility of SARS-CoV-2 lineage B. 1.1. 7 by age and viral load. Nat. Commun. 2021, 12, 7251. [Google Scholar] [CrossRef] [PubMed]
- Althaus, C.L.; Reichmuth, M.; Hodcroft, E.; Riou, J.; Schibler, M.; Eckerle, I.; Kaiser, L.; Suter, F.; Huber, M.; Trkola, A. Transmission of SARS-CoV-2 variants in Switzerland. Inst. Soc. Prev. Med. 2021. Available online: https://ispmbern.github.io/covid-19/variants/ (accessed on 27 February 2022).
- Davies, N.G.; Abbott, S.; Barnard, R.C.; Jarvis, C.I.; Kucharski, A.J.; Munday, J.D.; Pearson, C.A.; Russell, T.W.; Tully, D.C.; Washburne, A.D. Estimated transmissibility and impact of SARS-CoV-2 lineage B. 1.1. 7 in England. Science 2021, 372, eabg3055. [Google Scholar] [CrossRef] [PubMed]
- Zhao, S.; Lou, J.; Cao, L.; Zheng, H.; Chong, M.K.; Chen, Z.; Chan, R.W.; Zee, B.C.; Chan, P.K.; Wang, M.H. Quantifying the transmission advantage associated with N501Y substitution of SARS-CoV-2 in the UK: An early data-driven analysis. J. Travel Med. 2021, 28, taab011. [Google Scholar] [CrossRef]
- Curran, J.; Dol, J.; Boulos, L.; Somerville, M.; McCulloch, H.; MacDonald, M.; LeBlanc, J.; Barrett, L.; Hatchette, T.; Comeau, J. Transmission characteristics of SARS-CoV-2 variants of concern Rapid Scoping Review. medRxiv 2021. [Google Scholar] [CrossRef]
- Brown, K.A.; Tibebu, S.; Daneman, N.; Schwartz, K.L.; Whelan, M.; Buchan, S.A. Comparative Household Secondary Attack Rates associated with B. 1.1. 7, B. 1.351, and P. 1 SARS-CoV-2 Variants. medRxiv 2021. [Google Scholar] [CrossRef]
- Washington, N.L.; Gangavarapu, K.; Zeller, M.; Bolze, A.; Cirulli, E.T.; Barrett, K.M.S.; Larsen, B.B.; Anderson, C.; White, S.; Cassens, T. Emergence and rapid transmission of SARS-CoV-2 B. 1.1. 7 in the United States. Cell 2021, 184, 2587–2594.e7. [Google Scholar] [CrossRef]
- Kirby, T. New variant of SARS-CoV-2 in UK causes surge of COVID-19. Lancet Respir. Med. 2021, 9, e20–e21. [Google Scholar] [CrossRef]
- Graham, M.S.; Sudre, C.H.; May, A.; Antonelli, M.; Murray, B.; Varsavsky, T.; Kläser, K.; Canas, L.S.; Molteni, E.; Modat, M. Changes in symptomatology, reinfection, and transmissibility associated with the SARS-CoV-2 variant B. 1.1. 7: An ecological study. Lancet Public Health 2021, 6, e335–e345. [Google Scholar] [CrossRef]
- Kow, C.S.; Hasan, S.S. Could it be that the B. 1.1. 7 lineage is more deadly? Infect. Control. Hosp. Epidemiol. 2021, 43, 678–679. [Google Scholar] [CrossRef]
- Martínez-García, L.; Espinel, M.A.; Abreu, M.; González-Alba, J.M.; Gijón, D.; McGee, A.; Cantón, R.; Galán, J.C.; Aranaz, J. Emergence and Spread of B. 1.1. 7 Lineage in Primary Care and Clinical Impact in the Morbi-Mortality among Hospitalized Patients in Madrid, Spain. Microorganisms 2021, 9, 1517. [Google Scholar] [CrossRef]
- Tober-Lau, P.; Schwarz, T.; Hillus, D.; Spieckermann, J.; Helbig, E.T.; Lippert, L.J.; Thibeault, C.; Koch, W.; Bergfeld, L.; Niemeyer, D. Outbreak of SARS-CoV-2 B. 1.1. 7 lineage after vaccination in long-term care facility, Germany, February–March 2021. Emerg. Infect. Dis. 2021, 27, 2169. [Google Scholar] [CrossRef] [PubMed]
- Nyberg, T.; Twohig, K.A.; Harris, R.J.; Seaman, S.R.; Flannagan, J.; Allen, H.; Charlett, A.; De Angelis, D.; Dabrera, G.; Presanis, A.M. Risk of hospital admission for patients with SARS-CoV-2 variant B. 1.1. 7: Cohort analysis. BMJ 2021, 373. [Google Scholar]
- Bager, P.; Wohlfahrt, J.; Rasmussen, M.; Albertsen, M.; Krause, T. Hospitalisation associated with SARS-CoV-2 delta variant in Denmark. Lancet. Infect. Dis. 2021, 21, 1351. [Google Scholar] [CrossRef]
- Younes, M.; Hamze, K.; Nassar, H.; Makki, M.; Ghadar, M.; Nguewa, P.; Sater, F.A. Emergence and fast spread of B. 1.1. 7 lineage in Lebanon. medRxiv 2021. [Google Scholar] [CrossRef]
- Borges, V.; Sousa, C.; Menezes, L.; Gonçalves, A.M.; Picão, M.; Almeida, J.P.; Vieita, M.; Santos, R.; Silva, A.R.; Costa, M. Tracking SARS-CoV-2 lineage B. 1.1. 7 dissemination: Insights from nationwide spike gene target failure (SGTF) and spike gene late detection (SGTL) data, Portugal, week 49 2020 to week 3 2021. Eurosurveillance 2021, 26, 2100131. [Google Scholar] [CrossRef]
- Strålin, K.; Bruce, D.; Wahlström, E.; Walther, S.; Rehn, M.; Carnahan, A.; Andersson, E.; Bark, A.M.B.; Hanberger, H. Impact of the Alpha VOC on disease severity in SARS-CoV-2-positive adults in Sweden. J. Infect. 2021, 84, e3–e5. [Google Scholar] [CrossRef] [PubMed]
- Iacobucci, G. COVID-19: New UK variant may be linked to increased death rate, early data indicate. BMJ 2021, 372, n230. [Google Scholar] [CrossRef] [PubMed]
- Grint, D.J.; Wing, K.; Houlihan, C.; Gibbs, H.P.; Evans, S.J.; Williamson, E.; McDonald, H.I.; Bhaskaran, K.; Evans, D.; Walker, A.J. Severity of SARS-CoV-2 alpha variant (B. 1.1. 7) in England. Clin. Infect. Dis. 2021, ciab754. [Google Scholar] [CrossRef] [PubMed]
- Challen, R.; Brooks-Pollock, E.; Read, J.M.; Dyson, L.; Tsaneva-Atanasova, K.; Danon, L. Risk of mortality in patients infected with SARS-CoV-2 variant of concern 202012/1: Matched cohort study. BMJ 2021, 372, n579. [Google Scholar] [CrossRef]
- Davies, N.G.; Jarvis, C.I.; Edmunds, W.J.; Jewell, N.P.; Diaz-Ordaz, K.; Keogh, R.H. Increased mortality in community-tested cases of SARS-CoV-2 lineage B. 1.1. 7. Nature 2021, 593, 270–274. [Google Scholar] [CrossRef]
- Frampton, D.; Rampling, T.; Cross, A.; Bailey, H.; Heaney, J.; Byott, M.; Scott, R.; Sconza, R.; Price, J.; Margaritis, M. Genomic characteristics and clinical effect of the emergent SARS-CoV-2 B. 1.1. 7 lineage in London, UK: A whole-genome sequencing and hospital-based cohort study. Lancet Infect. Dis. 2021, 21, 1246–1256. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Q.; Wei, P.; Chen, Z.; Aviszus, K.; Yang, J.; Downing, W.; Jiang, C.; Liang, B.; Reynoso, L. The basis of a more contagious 501Y. V1 variant of SARS-COV-2. Cell Res. 2021, 31, 720–722. [Google Scholar] [CrossRef]
- Fratev, F. N501Y and K417N Mutations in the Spike Protein of SARS-CoV-2 Alter the Interactions with Both hACE2 and Human-Derived Antibody: A Free Energy of Perturbation Retrospective Study. J. Chem. Inf. Model. 2021, 61, 6079–6084. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, X.; Man, V.H.; Zhai, J.; Ji, B.; Wang, J. Binding Profile Assessment of N501Y: A More Infectious Mutation on the Receptor Binding Domain of SARS-CoV-2 Spike Protein. ChemRxiv 2021. [Google Scholar] [CrossRef]
- Ramanathan, M.; Ferguson, I.D.; Miao, W.; Khavari, P.A. SARS-CoV-2 B. 1.1. 7 and B. 1.351 Spike variants bind human ACE2 with increased affinity. Lancet Infect. Dis. 2021; in preprint. [Google Scholar] [CrossRef]
- Ontario, P.H. Focus on: An Overview of Cycle Threshold Values and Their Role in SARS-Cov-2 Real-Time PCR Test Interpretation. Available online: https://www.publichealthontario.ca/-/media/documents/ncov/main/2020/09/cycle-thresholdvalues-sars-cov2-pcr.pdf?la=en (accessed on 15 January 2022).
- Kidd, M.; Richter, A.; Best, A.; Cumley, N.; Mirza, J.; Percival, B.; Mayhew, M.; Megram, O.; Ashford, F.; White, T. S-Variant SARS-CoV-2 Lineage B1. 1.7 Is Associated With Significantly Higher Viral Load in Samples Tested by TaqPath Polymerase Chain Reaction. J. Infect. Dis. 2021, 223, 1666–1670. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.S.; Vihta, K.D.; Gethings, O.; Pritchard, E.; Jones, J.; House, T.; Bell, I.; Bell, J.; Newton, J.; Farrar, J. Increased infections, but not viral burden, with a new SARS-CoV-2 variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Dagpunar, J.S. Interim estimates of increased transmissibility, growth rate, and reproduction number of the COVID-19 B. 1.617. 2 variant of concern in the United Kingdom. medRxiv 2021. [Google Scholar] [CrossRef]
- Vöhringer, H.; Sinnott, M.; Amato, R.; Martincorena, I.; Kwiatkowski, D.; Barrett, J.C.; Gerstung, M. Lineage-specific growth of SARS-CoV-2 B. 1.1. 7 during the English national lockdown. Virological 2020. Available online: https://virological.org/t/lineage-specific-growth-of-sars-cov-2-b-1-1-7-during-the-english-national-lockdown/575 (accessed on 11 April 2022).
- Bonifazi, G.; Lista, L.; Menasce, D.; Mezzetto, M.; Pedrini, D.; Spighi, R.; Zoccoli, A. Study on the effects of the restrictive measures for containment of the COVID-19 pandemic on the reproduction number Rt in Italian regions. arXiv 2021, arXiv:2106.02603. [Google Scholar]
- Diabelko, D.; Dvorackova, M.; Heroldova, M.D.; Forte, G.; Cundrle, I.; Ruzicka, F.; Vrbsky, J. Monitoring of SARS-CoV-2 B. 1.1. 7 variant early-phase spreading in South-Moravian Region in the Czech Republic and evaluation of its pathogenicity. medRxiv 2021. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, E.-J.; Lee, S.-W.; Kwon, D. Review of the early reports of the epidemiological characteristics of the B. 1.1. 7 variant of SARS-CoV-2 and its spread worldwide. Osong Public Health Res. Perspect. 2021, 12, 139–148. [Google Scholar] [CrossRef]
- Singh, J.; Rahman, S.A.; Ehtesham, N.Z.; Hira, S.; Hasnain, S.E. SARS-CoV-2 variants of concern are emerging in India. Nat. Med. 2021, 27, 1131–1133. [Google Scholar] [CrossRef]
- Brown, K.A.; Gubbay, J.; Hopkins, J.; Patel, S.; Buchan, S.A.; Daneman, N.; Goneau, L. Rapid rise of S-gene target failure and the UK variant B. 1.1. 7 among COVID-19 isolates in the greater Toronto area, Canada. medRxiv 2021. [Google Scholar] [CrossRef]
- Grabowski, F.; Preibisch, G.; Giziński, S.; Kochańczyk, M.; Lipniacki, T. SARS-CoV-2 variant of concern 202012/01 has about twofold replicative advantage and acquires concerning mutations. Viruses 2021, 13, 392. [Google Scholar] [CrossRef] [PubMed]
- Novavax. Novavax COVID-19 Vaccine Demonstrates 89.3% Efficacy in UK Phase 3 Trial. Available online: https://ir.novavax.com/newsreleases/news-release-details/novavax-covid-19-vaccine-demonstrates-893-efficacy-uk-phase-3 (accessed on 18 January 2022).
- Chemaitelly, H.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; Tang, P.; Hasan, M.R.; Malek, J.A.; Coyle, P.; Ayoub, H.H.; Al Kanaani, Z. mRNA-1273 COVID-19 vaccine effectiveness against the B. 1.1. 7 and B. 1.351 variants and severe COVID-19 disease in Qatar. Nat. Med. 2021, 27, 1614–1621. [Google Scholar] [CrossRef]
- Callaway, E.; Mallapaty, S. Novavax offers first evidence that COVID vaccines protect people against variants. Nature 2021, 590, 17. [Google Scholar] [CrossRef]
- Shen, X.; Tang, H.; McDanal, C.; Wagh, K.; Fischer, W.; Theiler, J.; Yoon, H.; Li, D.; Haynes, B.F.; Sanders, K.O. SARS-CoV-2 variant B. 1.1. 7 is susceptible to neutralizing antibodies elicited by ancestral spike vaccines. Cell Host Microb. 2021, 29, 529–539.e3. [Google Scholar] [CrossRef]
- Wang, P.; Nair, M.S.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P.D. Antibody resistance of SARS-CoV-2 variants B. 1.351 and B. 1.1. 7. Nature 2021, 593, 130–135. [Google Scholar] [CrossRef]
- Wu, K.; Werner, A.P.; Moliva, J.I.; Koch, M.; Choi, A.; Stewart-Jones, G.B.; Bennett, H.; Boyoglu-Barnum, S.; Shi, W.; Graham, B.S. mRNA-1273 vaccine induces neutralizing antibodies against spike mutants from global SARS-CoV-2 variants. bioRxiv 2021. [Google Scholar] [CrossRef]
- Wang, Z.; Schmidt, F.; Weisblum, Y.; Muecksch, F.; Barnes, C.O.; Finkin, S.; Schaefer-Babajew, D.; Cipolla, M.; Gaebler, C.; Lieberman, J.A. mRNA vaccine-elicited antibodies to SARS-CoV-2 and circulating variants. Nature 2021, 592, 616–622. [Google Scholar] [CrossRef]
- Xie, X.; Liu, Y.; Liu, J.; Zhang, X.; Zou, J.; Fontes-Garfias, C.R.; Xia, H.; Swanson, K.A.; Cutler, M.; Cooper, D. Neutralization of SARS-CoV-2 spike 69/70 deletion, E484K and N501Y variants by BNT162b2 vaccine-elicited sera. Nat. Med. 2021, 27, 620–621. [Google Scholar] [CrossRef]
- Collier, D.A.; De Marco, A.; Ferreira, I.A.; Meng, B.; Datir, R.; Walls, A.C.; Bassi, J.; Pinto, D.; Fregni, C.S.; Bianchi, S. SARS-CoV-2 B. 1.1. 7 sensitivity to mRNA vaccine-elicited, convalescent and monoclonal antibodies. medRxiv 2021. [Google Scholar] [CrossRef]
- Muik, A.; Wallisch, A.-K.; Sänger, B.; Swanson, K.A.; Mühl, J.; Chen, W.; Cai, H.; Maurus, D.; Sarkar, R.; Türeci, Ö. Neutralization of SARS-CoV-2 lineage B. 1.1. 7 pseudovirus by BNT162b2 vaccine–elicited human sera. Science 2021, 371, 1152–1153. [Google Scholar] [CrossRef] [PubMed]
- WHO. Weekly Epidemiological Update on COVID-19. 6 July 2021. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---6-july-2021 (accessed on 20 February 2022).
- Rovida, F.; Cassaniti, I.; Paolucci, S.; Percivalle, E.; Sarasini, A.; Piralla, A.; Giardina, F.; Sammartino, J.C.; Ferrari, A.; Bergami, F. SARS-CoV-2 vaccine breakthrough infections with the alpha variant are asymptomatic or mildly symptomatic among health care workers. Nat. Commun. 2021, 12, 6032. [Google Scholar] [CrossRef] [PubMed]
- Duerr, R.; Dimartino, D.; Marier, C.; Zappile, P.; Wang, G.; Lighter, J.; Elbel, B.; Troxel, A.B.; Heguy, A. Dominance of Alpha and Iota variants in SARS-CoV-2 vaccine breakthrough infections in New York City. J. Clin. Investig. 2021, 131, e152702. [Google Scholar] [CrossRef] [PubMed]
- Tegally, H.; Wilkinson, E.; Althaus, C.L.; Giovanetti, M.; San, J.E.; Giandhari, J.; Pillay, S.; Naidoo, Y.; Ramphal, U.; Msomi, N. Rapid replacement of the Beta variant by the Delta variant in South Africa. medRxiv 2021. [Google Scholar] [CrossRef]
- Yadav, P.D.; Nyayanit, D.A.; Sahay, R.R.; Shete, A.M.; Majumdar, T.; Patil, S.; Patil, D.Y.; Gupta, N.; Kaur, H.; Aggarwal, N. Imported SARS-CoV-2 V501Y. V2 variant (B. 1.351) detected in travelers from South Africa and Tanzania to India. Travel Med. Infect. Dis. 2021, 41, 102023. [Google Scholar] [CrossRef]
- Jungnick, S.; Hobmaier, B.; Mautner, L.; Hoyos, M.; Haase, M.; Baiker, A.; Lahne, H.; Eberle, U.; Wimmer, C.; Hepner, S. Detection of the new SARS-CoV-2 variants of concern B. 1.1. 7 and B. 1.351 in five SARS-CoV-2 rapid antigen tests (RATs), Germany, March 2021. Eurosurveillance 2021, 26, 2100413. [Google Scholar] [CrossRef]
- Feder, K.A.; Pearlowitz, M.; Goode, A.; Duwell, M.; Williams, T.W.; Chen-Carrington, P.A.; Patel, A.; Dominguez, C.; Keller, E.N.; Klein, L. Linked Clusters of SARS-CoV-2 Variant B. 1.351—Maryland, January–February 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 627. [Google Scholar] [CrossRef]
- Boloko, L.; Lifson, A.; Little, F.; De Wet, T.; Papavarnavas, N.; Marais, G.; Hsiao, N.-y.; Roslee, M.-J.; Doolabh, D.; Iranzadeh, A. Severity and inpatient mortality of COVID-19 pneumonia from Beta variant infection: A clinical cohort study in Cape Town, South Africa. medRxiv 2021. [Google Scholar] [CrossRef]
- Centre for Mathematical Modelling of Infectious Diseases. Estimates of Severity and Transmissibility of Novel South Africa SARSCoV-2 Variant 501Y.V2. Available online: https://cmmid.github.io/topics/covid19/reports/sa-novelvariant/2021_01_11_Transmissibility_and_severity_of_501Y_V2_in_SA.pdf (accessed on 15 January 2022).
- Tegally, H.; Wilkinson, E.; Giovanetti, M.; Iranzadeh, A.; Fonseca, V.; Giandhari, J.; Doolabh, D.; Pillay, S.; San, E.J.; Msomi, N. Emergence and rapid spread of a new severe acute respiratory syndrome-related coronavirus 2 (SARS-CoV-2) lineage with multiple spike mutations in South Africa. medRxiv 2020. [Google Scholar] [CrossRef]
- Campbell, F.; Archer, B.; Laurenson-Schafer, H.; Jinnai, Y.; Konings, F.; Batra, N.; Pavlin, B.; Vandemaele, K.; Van Kerkhove, M.D.; Jombart, T. Increased transmissibility and global spread of SARS-CoV-2 variants of concern as at June 2021. Eurosurveillance 2021, 26, 2100509. [Google Scholar] [CrossRef]
- Vermeulen, M.; Mhlanga, L.; Sykes, W.; Coleman, C.; Pietersen, N.; Cable, R.; Swanevelder, R.; Glatt, T.N.; Grebe, E.; Welte, A. Prevalence of anti-SARS-CoV-2 antibodies among blood donors in South Africa during the period January–May 2021. Res. Sq. 2021; preprint. [Google Scholar] [CrossRef]
- Roquebert, B.; Trombert-Paolantoni, S.; Haim-Boukobza, S.; Lecorche, E.; Verdurme, L.; Foulongne, V.; Sofonea, M.T.; Alizon, S. The SARS-CoV-2 B. 1.351 lineage (VOC β) is outgrowing the B. 1.1. 7 lineage (VOC α) in some French regions in April 2021. Eurosurveillance 2021, 26, 2100447. [Google Scholar] [CrossRef] [PubMed]
- Motozono, C.; Toyoda, M.; Zahradnik, J.; Saito, A.; Nasser, H.; Tan, T.S.; Ngare, I.; Kimura, I.; Uriu, K.; Kosugi, Y. SARS-CoV-2 spike L452R variant evades cellular immunity and increases infectivity. Cell Host Microb. 2021, 29, 1124–1136.e11. [Google Scholar] [CrossRef] [PubMed]
- Staub, T.; Arendt, V.; de la Vega, E.C.L.; Braquet, P.; Michaux, C.; Kohnen, M.; Tsobo, C.; Abdelrahman, T.; Wienecke-Baldacchino, A.; Francois, J.-H. Case series of four re-infections with a SARS-CoV-2 B. 1.351 variant, Luxembourg, February 2021. Eurosurveillance 2021, 26, 2100423. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Chen, Q.; Yang, G.; He, L.; Fan, H.; Deng, Y.-Q.; Wang, Y.; Teng, Y.; Zhao, Z.; Cui, Y. Adaptation of SARS-CoV-2 in BALB/c mice for testing vaccine efficacy. Science 2020, 369, 1603–1607. [Google Scholar] [CrossRef] [PubMed]
- Abu-Raddad, L.J.; Chemaitelly, H.; Bertollini, R. Severity of SARS-CoV-2 reinfections as compared with primary infections. N. Engl. J. Med. 2021, 385, 2487–2489. [Google Scholar] [CrossRef]
- Cantón, R.; Ramos, P.D.L.; García-Botella, A.; García-Lledó, A.; Gómez-Pavón, J.; del Castillo, J.G.; Hernández-Sampelayo, T.; Martín-Delgado, M.C.; Sánchez, F.J.M.; Martínez-Sellés, M. New variants of SARS-CoV-2. Rev. Española Quimioter. 2021, 34, 419. [Google Scholar] [CrossRef] [PubMed]
- GVN. Beta (B.1.351). Available online: https://gvn.org/covid-19/beta-b-1-351/ (accessed on 11 April 2022).
- Li, Y.; Wang, X.; Campbell, H.; Nair, H.; for COVID, U.N. The association of community mobility with the time-varying reproduction number (R) of SARS-CoV-2: A modelling study across 330 local UK authorities. Lancet Digit. Health 2021, 3, e676–e683. [Google Scholar] [CrossRef]
- Kleynhans, J.; Tempia, S.; Wolter, N.; von Gottberg, A.; Bhiman, J.N.; Buys, A.; Moyes, J.; McMorrow, M.L.; Kahn, K.; Gómez-Olivé, F.X. SARS-CoV-2 Seroprevalence in a rural and urban household cohort during first and second waves of infections, South Africa, July 2020–March 2021. Emerg. Infect. Dis. 2021, 27, 3020. [Google Scholar] [CrossRef]
- Abu-Raddad, L.J.; Chemaitelly, H.; Ayoub, H.H.; Yassine, H.M.; Benslimane, F.; Al Khatib, H.A.; Tang, P.; Hasan, M.R.; Coyle, P.; AlMukdad, S. Severity, criticality, and fatality of the SARS-CoV-2 Beta variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Cosentino, G.; Bernard, M.; Giannoli, J.-M.; Florence, D. SARS-CoV-2 viral dynamics in infections with Alpha and Beta variants of concern in the French community. J. Infect. 2021, 84, 94–118. [Google Scholar] [CrossRef]
- Radvak, P.; Kwon, H.-J.; Kosikova, M.; Ortega-Rodriguez, U.; Xiang, R.; Phue, J.-N.; Shen, R.-F.; Rozzelle, J.; Kapoor, N.; Rabara, T. SARS-CoV-2 B. 1.1. 7 (alpha) and B. 1.351 (beta) variants induce pathogenic patterns in K18-hACE2 transgenic mice distinct from early strains. Nat. Commun. 2021, 12, 6559. [Google Scholar] [CrossRef]
- Golubchik, T.; Lythgoe, K.A.; Hall, M.D.; Ferretti, L.; Fryer, H.R.; MacInyre-Cockett, G.; de Cesare, M.; Trebes, A.; Piazza, P.; Buck, D. Early analysis of a potential link between viral load and the N501Y mutation in the SARS-COV-2 spike protein. medRxiv 2021. [Google Scholar] [CrossRef]
- Pan, T.; Chen, R.; He, X.; Yuan, Y.; Deng, X.; Li, R.; Yan, H.; Yan, S.; Liu, J.; Zhang, Y. Infection of wild-type mice by SARS-CoV-2 B. 1.351 variant indicates a possible novel cross-species transmission route. Signal Transduct. Target. Ther. 2021, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- Linka, K.; Peirlinck, M.; Schäfer, A.; Tikenogullari, O.Z.; Goriely, A.; Kuhl, E. Effects of B. 1.1. 7 and B. 1.351 on COVID-19 dynamics: A campus reopening study. Arch. Comput. Methods Eng. 2021, 28, 4225–4236. [Google Scholar] [CrossRef]
- Brum, E.; Saha, S.; Sania, A.; Tanmoy, A.M.; Hooda, Y.; Tanni, A.; Goswami, S.; Al Sium, S.M.; Sajib, M.S.I.; Malaker, R. Surging COVID-19 in Bangladesh driven by B. 1.351 variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Wibmer, C.; Ayres, F.; Hermanus, T.; Madzivhandila, M.; Kgagudi, P.; Lambson, B.; Vermeulen, M.; Berg, K.; van den Rossouw, T.; Boswell, M. SARS-CoV-2 501Y.V2 escapes neutralization by South African COVID-19 donor plasma. Nat. Med. 2021, 27, 622–625. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.; Dejnirattisai, W.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.; Tuekprakhon, A.; Nutalai, R. Evidence of escape of SARS-CoV-2 variant B. 1.351 from natural and vaccine-induced sera. Cell 2021, 184, 2348–2361.e6. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhang, Q.; Ge, J.; Ren, W.; Zhang, R.; Lan, J.; Ju, B.; Su, B.; Yu, F.; Chen, P. Analysis of SARS-CoV-2 variant mutations reveals neutralization escape mechanisms and the ability to use ACE2 receptors from additional species. Immunity 2021, 54, 1611–1621.e5. [Google Scholar] [CrossRef] [PubMed]
- Wu, K.; Werner, A.P.; Koch, M.; Choi, A.; Narayanan, E.; Stewart-Jones, G.B.; Colpitts, T.; Bennett, H.; Boyoglu-Barnum, S.; Shi, W. Serum neutralizing activity elicited by mRNA-1273 vaccine. N. Engl. J. Med. 2021, 384, 1468–1470. [Google Scholar] [CrossRef]
- Madhi, S.A.; Baillie, V.; Cutland, C.L.; Voysey, M.; Koen, A.L.; Fairlie, L.; Padayachee, S.D.; Dheda, K.; Barnabas, S.L.; Bhorat, Q.E. Efficacy of the ChAdOx1 nCoV-19 COVID-19 vaccine against the B. 1.351 variant. N. Engl. J. Med. 2021, 384, 1885–1898. [Google Scholar] [CrossRef]
- Ho, D.; Wang, P.; Liu, L.; Iketani, S.; Luo, Y.; Guo, Y.; Wang, M.; Yu, J.; Zhang, B.; Kwong, P. Increased resistance of SARS-CoV-2 variants B. 1.351 and B. 1.1. 7 to antibody neutralization. Res. Sq. 2021, rs. 3, rs-155394. [Google Scholar]
- Zeng, C.; Evans, J.P.; Qu, P.; Faraone, J.; Zheng, Y.-M.; Carlin, C.; Bednash, J.S.; Zhou, T.; Lozanski, G.; Mallampalli, R. Neutralization and stability of SARS-CoV-2 Omicron variant. bioRxiv 2021. [Google Scholar] [CrossRef]
- Tostanoski, L.H.; Yu, J.; Mercado, N.B.; McMahan, K.; Jacob-Dolan, C.; Martinot, A.J.; Piedra-Mora, C.; Anioke, T.; Chang, A.; Giffin, V.M. Immunity elicited by natural infection or Ad26. COV2. S vaccination protects hamsters against SARS-CoV-2 variants of concern. Sci. Transl. Med. 2021, 13, eabj3789. [Google Scholar] [CrossRef]
- Alter, G.; Yu, J.; Liu, J.; Chandrashekar, A.; Borducchi, E.N.; Tostanoski, L.H.; McMahan, K.; Jacob-Dolan, C.; Martinez, D.R.; Chang, A. Immunogenicity of Ad26. COV2. S vaccine against SARS-CoV-2 variants in humans. Nature 2021, 596, 268–272. [Google Scholar] [CrossRef]
- Wang, R.; Chen, J.; Hozumi, Y.; Yin, C.; Wei, G.-W. Emerging vaccine-breakthrough SARS-CoV-2 variants. ACS Infect. Dis. 2022, 8, 546–556. [Google Scholar] [CrossRef] [PubMed]
- Sabino, E.C.; Buss, L.F.; Carvalho, M.P.; Prete, C.A.; Crispim, M.A.; Fraiji, N.A.; Pereira, R.H.; Parag, K.V.; da Silva Peixoto, P.; Kraemer, M.U. Resurgence of COVID-19 in Manaus, Brazil, despite high seroprevalence. Lancet 2021, 397, 452–455. [Google Scholar] [CrossRef]
- Duong, D. Alpha, Beta, Delta, Gamma: What’s important to know about SARS-CoV-2 variants of concern? CMAJ 2021, 193, E1059–E1060. [Google Scholar] [CrossRef] [PubMed]
- Di Giallonardo, F.; Puglia, I.; Curini, V.; Cammà, C.; Mangone, I.; Calistri, P.; Cobbin, J.C.; Holmes, E.C.; Lorusso, A. Emergence and spread of SARS-CoV-2 lineages B. 1.1. 7 and P. 1 in Italy. Viruses 2021, 13, 794. [Google Scholar] [CrossRef] [PubMed]
- Panzera, Y.; Goñi, N.; Calleros, L.; Ramos, N.; Frabasile, S.; Marandino, A.; Tomás, G.; Techera, C.; Grecco, S.; Fuques, E. Genome Sequences of SARS-CoV-2 P. 1 (Variant of Concern) and P. 2 (Variant of Interest) Identified in Uruguay. Microbiol. Resour. Announc. 2021, 10, e00410–e00421. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Zhou, D.; Supasa, P.; Liu, C.; Mentzer, A.J.; Ginn, H.M.; Zhao, Y.; Duyvesteyn, H.M.; Tuekprakhon, A.; Nutalai, R. Antibody evasion by the P. 1 strain of SARS-CoV-2. Cell 2021, 184, 2939–2954.e9. [Google Scholar] [CrossRef]
- Padilha, D.A.; Benetti-Filho, V.; Moreira, R.S.; Soratto, T.S.T.; Maia, G.A.; Christoff, A.P.; Barazzetti, F.H.; Schorner, M.A.; Ferrari, F.L.; Martins, C.L. The emergence of two distinct SARS-CoV-2 Gamma related variants during the second wave of COVID-19 in Santa Catarina, Southern Brazil and the rapid spread of P. 1-like-II SARS-CoV-2 variant transmission and a regionalization in the Western region. medRxiv 2022. [Google Scholar] [CrossRef]
- Faria, N.R.; Mellan, T.A.; Whittaker, C.; Claro, I.M.; Candido, D.d.S.; Mishra, S.; Crispim, M.A.; Sales, F.C.; Hawryluk, I.; McCrone, J.T. Genomics and epidemiology of the P. 1 SARS-CoV-2 lineage in Manaus, Brazil. Science 2021, 372, 815–821. [Google Scholar] [CrossRef]
- Janik, E.; Niemcewicz, M.; Podogrocki, M.; Majsterek, I.; Bijak, M. The emerging concern and interest SARS-CoV-2 variants. Pathogens 2021, 10, 633. [Google Scholar] [CrossRef]
- Sanyaolu, A.; Okorie, C.; Marinkovic, A.; Haider, N.; Abbasi, A.F.; Jaferi, U.; Prakash, S.; Balendra, V. The emerging SARS-CoV-2 variants of concern. Ther. Adv. Infect. Dis. 2021, 8, 20499361211024372. [Google Scholar] [CrossRef]
- Coutinho, R.M.; Marquitti, F.M.D.; Ferreira, L.S.; Borges, M.E.; da Silva, R.L.P.; Canton, O.; Portella, T.P.; Lyra, S.P.; Franco, C.; Plucinski, M.M. Model-based estimation of transmissibility and reinfection of SARS-CoV-2 P. 1 variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Naveca, F.G.; Nascimento, V.; de Souza, V.C.; de Lima Corado, A.; Nascimento, F.; Silva, G.; Costa, Á.; Duarte, D.; Pessoa, K.; Mejía, M. COVID-19 in Amazonas, Brazil, was driven by the persistence of endemic lineages and P. 1 emergence. Nat. Med. 2021, 27, 1230–1238. [Google Scholar] [CrossRef]
- Hogan, C.A.; Jassem, A.N.; Sbihi, H.; Joffres, Y.; Tyson, J.R.; Noftall, K.; Taylor, M.; Lee, T.; Fjell, C.; Wilmer, A. Rapid Increase in SARS-CoV-2 P. 1 Lineage Leading to Codominance with B. 1.1. 7 Lineage, British Columbia, Canada, January–April 2021. Emerg. Infect. Dis. 2021, 27, 2802. [Google Scholar] [CrossRef]
- Stefanelli, P.; Trentini, F.; Guzzetta, G.; Marziano, V.; Mammone, A.; Poletti, P.; Grane, C.M.; Manica, M.; del Manso, M.; Andrianou, X. Co-circulation of SARS-CoV-2 variants B. 1.1. 7 and P. 1. medRxiv 2021. [Google Scholar] [CrossRef]
- UN. COVID Variants ‘Winning the Race against Vaccines’ Warns WHO Chief. Available online: https://news.un.org/en/story/2021/07/1095432 (accessed on 20 February 2022).
- GISAID. Outbreak.Info: P.1 Lineage Report. 2021. Available online: https://outbreak.info/ (accessed on 15 February 2022).
- CDC. COVID Data Tracker: Variant Proportions. Available online: https://covid.cdc.gov/covid-data-tracker/#variant-proportions (accessed on 4 February 2022).
- Naveca, F.; Nascimento, V.; Souza, V.; Corado, A.; Nascimento, F.; Silva, G.; Costa, Á.; Duarte, D.; Pessoa, K.; Mejía, M. COVID-19 epidemic in the Brazilian state of Amazonas was driven by long-term persistence of endemic SARS-CoV-2 lineages and the recent emergence of the new Variant of Concern P. 1. Res. Sq. 2021; in preprint. [Google Scholar] [CrossRef]
- L’Huillier, A.G.; Torriani, G.; Pigny, F.; Kaiser, L.; Eckerle, I. Culture-competent SARS-CoV-2 in nasopharynx of symptomatic neonates, children, and adolescents. Emerg. Infect. Dis. 2020, 26, 2494. [Google Scholar] [CrossRef] [PubMed]
- Waudby-West, R.; Parcell, B.J.; Palmer, C.N.; Bell, S.; Chalmers, J.D.; Siddiqui, M.K. The association between SARS-CoV-2 RT-PCR cycle threshold and mortality in a community cohort. Eur. Respir. J. 2021, 58, 2100360. [Google Scholar] [CrossRef]
- Taylor, L. COVID-19: Researchers Find Higher than Expected Reinfections with P. 1 Variant among the Brazilian Amazon. BMJ 2021, 373, n1353. [Google Scholar] [CrossRef]
- Romano, C.M.; Felix, A.C.; Paula, A.V.d.; Jesus, J.G.d.; Andrade, P.S.; Cândido, D.; Oliveira, F.M.d.; Ribeiro, A.C.; Silva, F.C.d.; Inemami, M. SARS-CoV-2 reinfection caused by the P. 1 lineage in Araraquara city, Sao Paulo State, Brazil. Rev. Inst. Med. Trop. São Paulo 2021, 63, e36. [Google Scholar] [CrossRef]
- Funk, T.; Pharris, A.; Spiteri, G.; Bundle, N.; Melidou, A.; Carr, M.; Gonzalez, G.; Garcia-Leon, A.; Crispie, F.; O’Connor, L. Characteristics of SARS-CoV-2 variants of concern B. 1.1. 7, B. 1.351 or P. 1: Data from seven EU/EEA countries, weeks 38/2020 to 10/2021. Eurosurveillance 2021, 26, 2100348. [Google Scholar] [CrossRef] [PubMed]
- Freitas, A.R.; Lemos, D.R.; Beckedorff, O.A.; Cavalcante, L.P.; Siqueira, A.M.; Mello, R.C.; Barros, E.N. The increase in the risk of severity and fatality rate of covid-19 in southern Brazil after the emergence of the Variant of Concern (VOC) SARS-CoV-2 P. 1 was greater among young adults without pre-existing risk conditions. medRxiv 2021. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Hilton, S.K.; Ellis, D.; Crawford, K.H.; Dingens, A.S.; Navarro, M.J.; Bowen, J.E.; Tortorici, M.A.; Walls, A.C. Deep mutational scanning of SARS-CoV-2 receptor binding domain reveals constraints on folding and ACE2 binding. Cell 2020, 182, 1295–1310.e20. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, C.K.V.; Gräf, T.; de Lorenzo Barcia, C.A.; Costa, V.F.; de Oliveira, J.L.; da Hora Passos, R.; Bastos, I.N.; de Santana, M.C.B.; Santos, I.M.; de Sousa, K.A.F. SARS-CoV-2 variant of concern P. 1 (Gamma) infection in young and middle-aged patients admitted to the intensive care units of a single hospital in Salvador, Northeast Brazil, February 2021. Int. J. Infect. Dis. 2021, 111, 47–54. [Google Scholar] [CrossRef]
- To, A.; Wong, T.A.S.; Lieberman, M.M.; Thompson, K.; Ball, A.H.; Pessaint, L.; Greenhouse, J.; Daham, N.; Cook, A.; Narvaez, B. A Recombinant Subunit Vaccine Induces a Potent, Broadly Neutralizing, and Durable Antibody Response in Macaques against the SARS-CoV-2 P. 1 (Gamma) Variant. ACS Infect. Dis. 2022, 8, 825–840. [Google Scholar] [CrossRef]
- Oróstica, K.Y.; Contreras, S.; Mohr, S.B.; Dehning, J.; Bauer, S.; Medina-Ortiz, D.; Iftekhar, E.N.; Mujica, K.; Covarrubias, P.C.; Ulloa, S. Mutational signatures and transmissibility of SARS-CoV-2 Gamma and Lambda variants. arXiv 2021, arXiv:2108.10018. [Google Scholar]
- Hitchings, M.D.; Ranzani, O.T.; Dorion, M.; D’Agostini, T.L.; de Paula, R.C.; de Paula, O.F.P.; de Moura Villela, E.F.; Torres, M.S.S.; de Oliveira, S.B.; Schulz, W. Effectiveness of ChAdOx1 vaccine in older adults during SARS-CoV-2 Gamma variant circulation in São Paulo. Nat. Commun. 2021, 12, 6220. [Google Scholar] [CrossRef]
- Wang, P.; Casner, R.G.; Nair, M.S.; Wang, M.; Yu, J.; Cerutti, G.; Liu, L.; Kwong, P.D.; Huang, Y.; Shapiro, L. Increased resistance of SARS-CoV-2 variant P. 1 to antibody neutralization. Cell Host Microb. 2021, 29, 747–751.e4. [Google Scholar] [CrossRef]
- Mallapaty, S. China COVID vaccine reports mixed results—What does that mean for the pandemic. Nature 2021, 15. [Google Scholar] [CrossRef]
- Shapiro, J.; Dean, N.E.; Madewell, Z.J.; Yang, Y.; Halloran, M.E.; Longini, I.M. Efficacy estimates for various COVID-19 vaccines: What we know from the literature and reports. medRxiv 2021. [Google Scholar] [CrossRef]
- Souza, W.M.; Amorim, M.R.; Sesti-Costa, R.; Coimbra, L.D.; Brunetti, N.S.; Toledo-Teixeira, D.A.; de Souza, G.F.; Muraro, S.P.; Parise, P.L.; Barbosa, P.P. Neutralisation of SARS-CoV-2 lineage P. 1 by antibodies elicited through natural SARS-CoV-2 infection or vaccination with an inactivated SARS-CoV-2 vaccine: An immunological study. Lancet Microb. 2021, 2, e527–e535. [Google Scholar] [CrossRef]
- Thye, A.Y.-K.; Law, J.W.-F.; Pusparajah, P.; Letchumanan, V.; Chan, K.-G.; Lee, L.-H. Emerging SARS-CoV-2 variants of concern (VOCs): An impending global crisis. Biomedicines 2021, 9, 1303. [Google Scholar] [CrossRef] [PubMed]
- Hodcroft, E.B. CoVariants: SARS-CoV-2 Mutations and Variants of Interest. 2021. Available online: https://covariants.org/ (accessed on 24 May 2022).
- Peacock, T.P.; Goldhill, D.H.; Zhou, J.; Baillon, L.; Frise, R.; Swann, O.C.; Kugathasan, R.; Penn, R.; Brown, J.C.; Sanchez-David, R.Y. The furin cleavage site in the SARS-CoV-2 spike protein is required for transmission in ferrets. Nat. Microbiol. 2021, 6, 899–909. [Google Scholar] [CrossRef]
- Hoffmann, M.; Kleine-Weber, H.; Pöhlmann, S. A multibasic cleavage site in the spike protein of SARS-CoV-2 is essential for infection of human lung cells. Mol. Cell 2020, 78, 779–784.e5. [Google Scholar] [CrossRef]
- Ingraham, N.E.; Ingbar, D.H. The omicron variant of SARS-CoV-2: Understanding the known and living with unknowns. Clin. Transl. Med. 2021, 11, e685. [Google Scholar] [CrossRef] [PubMed]
- Malabadi, R.B.; Kolkar, K.P.; Meti, N.T.; Chalannavar, R.K. Outbreak of Coronavirus (SARS-CoV-2) Delta variant (B. 1.617. 2) and Delta Plus (AY. 1) with fungal infections, Mucormycosis: Herbal medicine treatment. Int. J. Res. Sci. Innov. 2021, 8, 59–70. [Google Scholar]
- Liu, C.; Ginn, H.M.; Dejnirattisai, W.; Supasa, P.; Wang, B.; Tuekprakhon, A.; Nutalai, R.; Zhou, D.; Mentzer, A.J.; Zhao, Y. Reduced neutralization of SARS-CoV-2 B. 1.617 by vaccine and convalescent serum. Cell 2021, 184, 4220–4236.e13. [Google Scholar] [CrossRef] [PubMed]
- Rahman, F.I.; Ether, S.A.; Islam, M.R. The “Delta Plus” COVID-19 variant has evolved to become the next potential variant of concern: Mutation history and measures of prevention. J. Basic Clin. Physiol. Pharmacol. 2021, 33, 109–112. [Google Scholar] [CrossRef]
- Earnest, R.; Uddin, R.; Matluk, N.; Renzette, N.; Turbett, S.E.; Siddle, K.J.; Loreth, C.; Adams, G.; Tomkins-Tinch, C.H.; Petrone, M.E. Comparative transmissibility of SARS-CoV-2 variants delta and alpha in New England, USA. Cell Rep. Med. 2022, 3, 100583. [Google Scholar] [CrossRef]
- Adam, D. What scientists know about new, fast-spreading coronavirus variants. Nature 2021, 594, 19–20. [Google Scholar] [CrossRef]
- Hagen, A. How dangerous Is the Delta variant (B. 1.617. 2). Am. Soc. Microbiol. 2021. Available online: https://asm.org/Articles/2021/July/How-Dangerous-is-the-Delta-Variant-B-1-617-2 (accessed on 3 March 2022).
- Joseph, R.J.; Ser, H.-L. Stories from the East: COVID-19 Situation in India. Prog. Microb. Mol. Biol. 2021, 4. [Google Scholar] [CrossRef]
- England, P.H. Variants: Distribution of case data, 11 June 2021. 18 June2021. Available online: https://www.gov.uk/government/publications/covid-19-variants-genomically-confirmed-case-numbers/variants-distribution-of-case-data-18-june-2021 (accessed on 4 February 2022).
- SAGE. SPI-M-O: Summary of Further Modelling of Easing Restrictions—Roadmap Step 4 on 19 July 2021. Available online: https://www.gov.uk/government/publications/spi-m-o-summary-of-further-modelling-of-easing-restrictions-roadmap-step-4-on-19-july-2021-7-july-2021/spi-m-o-summary-of-further-modelling-of-easing-restrictions-roadmap-step-4-on-19-july-2021-7-july-2021 (accessed on 10 April 2022).
- Vaughan, A. Delta to Dominate World; Elsevier: Amsterdam, The Netherlands, 2021. [Google Scholar]
- Dyer, O. COVID-19: South Africa’s surge in cases deepens alarm over omicron variant. BMJ 2021, 375, n3013. [Google Scholar] [CrossRef] [PubMed]
- Mahase, E. Delta variant: What is happening with transmission, hospital admissions, and restrictions? BMJ 2021, 373, n1513. [Google Scholar] [CrossRef]
- Allen, H.; Vusirikala, A.; Flannagan, J.; Twohig, K.A.; Zaidi, A.; Chudasama, D.; Lamagni, T.; Groves, N.; Turner, C.; Rawlinson, C. Household transmission of COVID-19 cases associated with SARS-CoV-2 delta variant (B. 1.617. 2): National case-control study. Lancet Reg. Health-Eur. 2022, 12, 100252. [Google Scholar] [CrossRef]
- Research, J.U.P.A.E. Potential Community Transmission of B.1.617.2 Inferred by S-gene Positivity. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/988205/S1239_Joint_UNIversities_Pandemic_and_Epidemiological_Research.pdf (accessed on 31 January 2022).
- Kang, M.; Xin, H.; Yuan, J.; Ali, S.T.; Liang, Z.; Zhang, J.; Hu, T.; Lau, E.; Zhang, Y.; Zhang, M. Transmission dynamics and epidemiological characteristics of Delta variant infections in China. Medrxiv 2021. [Google Scholar] [CrossRef]
- Zhang, M.; Xiao, J.; Deng, A.; Zhang, Y.; Zhuang, Y.; Hu, T.; Li, J.; Tu, H.; Li, B.; Zhou, Y. Transmission dynamics of an outbreak of the COVID-19 Delta variant B. 1.617. 2—Guangdong Province, China, May–June 2021. China CDC Wkly. 2021, 3, 584. [Google Scholar] [CrossRef]
- Deng, X.; Garcia-Knight, M.A.; Khalid, M.M.; Servellita, V.; Wang, C.; Morris, M.K.; Sotomayor-González, A.; Glasner, D.R.; Reyes, K.R.; Gliwa, A.S. Transmission, infectivity, and neutralization of a spike L452R SARS-CoV-2 variant. Cell 2021, 184, 3426–3437.e8. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Wang, M.; Wei, G.-W. Mutations strengthened SARS-CoV-2 infectivity. J. Mol. Biol. 2020, 432, 5212–5226. [Google Scholar] [CrossRef] [PubMed]
- Yang, W.; Shaman, J. COVID-19 pandemic dynamics in India and impact of the SARS-CoV-2 Delta (B. 1.617. 2) variant. medrxiv 2021. [Google Scholar] [CrossRef]
- Shiehzadegan, S.; Alaghemand, N.; Fox, M.; Venketaraman, V. Analysis of the delta variant B. 1.617. 2 COVID-19. Clin. Pract. 2021, 11, 778–784. [Google Scholar] [CrossRef]
- Pastor, J.; Vega-Zelaya, L.; Martin Abad, E. Specific EEG encephalopathy pattern in SARS-CoV-2 patients. J. Clin. Med. 2020, 9, 1545. [Google Scholar] [CrossRef] [PubMed]
- Brucki, S.M.; Corazza, L.A.; de Queiroz, A.P.; Barros, M.P.; Tatsch, J.F.; Riso, I.L.; Batista, N.A.; Manfroi, G.; Sawada, L.A.; Batista, L.L. Neurological complications in COVID-19 patients from Latin America. Brain 2021, 144, e29. [Google Scholar] [CrossRef]
- Ong, S.W.X.; Chiew, C.J.; Ang, L.W.; Mak, T.-M.; Cui, L.; Toh, M.P.H.; Lim, Y.D.; Lee, P.H.; Lee, T.H.; Chia, P.Y. Clinical and Virological Features of SARS-CoV-2 Variants of Concern: A Retrospective Cohort Study Comparing B. 1.1. 7 (Alpha), B. 1.315 (Beta), and B. 1.617. 2 (Delta). Clin. Infect. Dis. 2021, 23, ciab721. [Google Scholar] [CrossRef]
- Rashedi, R.; Samieefar, N.; Akhlaghdoust, M.; Mashhadi, M.; Darzi, P.; Rezaei, N. Delta Variant: The New Challenge of COVID-19 Pandemic, an Overview of Epidemiological, Clinical, and Immune Characteristics. Acta Biomed. 2022, 93, 2. [Google Scholar]
- Twohig, K.A.; Nyberg, T.; Zaidi, A.; Thelwall, S.; Sinnathamby, M.A.; Aliabadi, S.; Seaman, S.R.; Harris, R.J.; Hope, R.; Lopez-Bernal, J. Hospital admission and emergency care attendance risk for SARS-CoV-2 delta (B. 1.617. 2) compared with alpha (B. 1.1. 7) variants of concern: A cohort study. Lancet Infect. Dis. 2021. [Google Scholar] [CrossRef]
- Sheikh, A.; McMenamin, J.; Taylor, B.; Robertson, C. SARS-CoV-2 Delta VOC in Scotland: Demographics, risk of hospital admission, and vaccine effectiveness. Lancet 2021, 397, 2461–2462. [Google Scholar] [CrossRef]
- Torjesen, I. COVID-19: Delta variant is now UK’s most dominant strain and spreading through schools. BMJ 2021, 373, n1445. [Google Scholar] [CrossRef]
- Fisman, D.; Tuite, A. Progressive Increase in Virulence of Novel SARS-CoV-2 Variants in Ontario, Canada, February to June, 2021. medRxiv 2021. [Google Scholar] [CrossRef]
- Khatri, R.; Siddqui, G.; Sadhu, S.; Maithil, V.; Vishwakarma, P.; Lohiya, B.; Goswami, A.; Ahmed, S.; Awasthi, A.; Samal, S. SARS-CoV-2 variants’-Alpha, Delta, and Omicron D614G and P681R/H mutations impact virus entry, fusion, and infectivity. Res. Sq. 2022; in preprint. [Google Scholar] [CrossRef]
- Zhang, J.; Xiao, T.; Cai, Y.; Lavine, C.L.; Peng, H.; Zhu, H.; Anand, K.; Tong, P.; Gautam, A.; Mayer, M.L. Membrane fusion and immune evasion by the spike protein of SARS-CoV-2 Delta variant. Science 2021, 374, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Garcia-Knight, M.A.; Khalid, M.M.; Servellita, V.; Wang, C.; Morris, M.K.; Sotomayor-González, A.; Glasner, D.R.; Reyes, K.R.; Gliwa, A.S. Transmission, infectivity, and antibody neutralization of an emerging SARS-CoV-2 variant in California carrying a L452R spike protein mutation. medRxiv 2021. [Google Scholar] [CrossRef]
- Mohammadi, M.; Shayestehpour, M.; Mirzaei, H. The impact of spike mutated variants of SARS-CoV2 [Alpha, Beta, Gamma, Delta, and Lambda] on the efficacy of subunit recombinant vaccines. Braz. J. Infect. Dis. 2021, 25, 101606. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Hofmann-Winkler, H.; Krüger, N.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Arora, P.; Sidarovich, A.; Moldenhauer, A.-S.; Winkler, M.S. SARS-CoV-2 variant B. 1.617 is resistant to Bamlanivimab and evades antibodies induced by infection and vaccination. Cell Rep. 2021, 36, 109415. [Google Scholar] [CrossRef]
- Motozono, C.; Toyoda, M.; Zahradnik, J.; Ikeda, T.; Saito, A.; Tan, T.S.; Ngare, I.; Nasser, H.; Kimura, I.; Uriu, K. An Emerging SARS-CoV-2 Mutant Evades Cellular Immunity and Increases Infectivity. Cell Host Microbe 2021. [Google Scholar] [CrossRef]
- Blanquart, F.; Abad, C.; Ambroise, J.; Bernard, M.; Cosentino, G.; Giannoli, J.-M.; Débarre, F. Characterisation of vaccine breakthrough infections of SARS-CoV-2 Delta and Alpha variants and within-host viral load dynamics in the community, France, June to July 2021. Eurosurveillance 2021, 26, 2100824. [Google Scholar] [CrossRef]
- Teyssou, E.; Delagrèverie, H.; Visseaux, B.; Lambert-Niclot, S.; Brichler, S.; Ferre, V.; Marot, S.; Jary, A.; Todesco, E.; Schnuriger, A. The Delta SARS-CoV-2 variant has a higher viral load than the Beta and the historical variants in nasopharyngeal samples from newly diagnosed COVID-19 patients. J. Infect. 2021, 83, e1–e3. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Deng, A.; Li, K.; Hu, Y.; Li, Z.; Xiong, Q.; Liu, Z.; Guo, Q.; Zou, L.; Zhang, H. Viral infection and transmission in a large well-traced outbreak caused by the Delta SARS-CoV-2 variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Chia, P.Y.; Ong, S.W.X.; Chiew, C.J.; Ang, L.W.; Chavatte, J.-M.; Mak, T.-M.; Cui, L.; Kalimuddin, S.; Chia, W.N.; Tan, C.W. Virological and serological kinetics of SARS-CoV-2 Delta variant vaccine-breakthrough infections: A multi-center cohort study. Clin. Microbiol. Infect. 2021, 28, 612.e1–612.e7. [Google Scholar]
- Despres, H.W.; Mills, M.G.; Shirley, D.J.; Schmidt, M.M.; Huang, M.-L.; Roychoudhury, P.; Jerome, K.R.; Greninger, A.L.; Bruce, E.A. Measuring infectious SARS-CoV-2 in clinical samples reveals a higher viral titer: RNA ratio for Delta and Epsilon vs. Alpha variants. Proc. Natl. Acad. Sci. USA 2022, 119, e2116518119. [Google Scholar] [CrossRef]
- Kissler, S.M.; Fauver, J.R.; Mack, C.; Tai, C.G.; Breban, M.I.; Watkins, A.E.; Samant, R.M.; Anderson, D.J.; Metti, J.; Khullar, G. Viral dynamics of SARS-CoV-2 variants in vaccinated and unvaccinated persons. N. Engl. J. Med. 2021, 385, 2489–2491. [Google Scholar] [CrossRef]
- Puhach, O.; Adea, K.; Hulo, N.; Sattonnet, P.; Genecand, C.; Iten, A.; Bausch, F.J.; Kaiser, L.; Vetter, P.; Eckerle, I. Infectious viral load in unvaccinated and vaccinated individuals infected with ancestral, Delta or Omicron SARS-CoV-2. Nat. Med. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Shi, Q.; Gao, X.; Hu, B. Research progress on characteristics, epidemiology and control measure of SARS-CoV-2 Delta VOC. Chin. J. Nosocomiol 2021, 31, 3703–3707. [Google Scholar]
- Liu, Y.; Rocklöv, J. The reproductive number of the Delta variant of SARS-CoV-2 is far higher compared to the ancestral SARS-CoV-2 virus. J. Travel Med. 2021, 28, taab124. [Google Scholar] [CrossRef] [PubMed]
- Mackie, D. Global Vulnerabilities to the COVID-19 Variant B.1.617.2. Available online: https://www.suerf.org/suer-policy-brief/26931/global-vulnerabilities-to-the-covid-19-variant-b-1-617-2 (accessed on 29 January 2022).
- Burki, T.K. Lifting of COVID-19 restrictions in the UK and the Delta variant. Lancet Respir. Med. 2021, 9, e85. [Google Scholar] [CrossRef]
- Bernal, J.L.; Andrews, N.; Gower, C.; Gallagher, E.; Simmons, R.; Thelwall, S.; Tessier, E.; Groves, N.; Dabrera, G.; Myers, R. Effectiveness of COVID-19 vaccines against the B. 1.617. 2 variant. medRxiv 2021. [Google Scholar] [CrossRef]
- Tang, P.; Hasan, M.R.; Chemaitelly, H.; Yassine, H.M.; Benslimane, F.M.; Al Khatib, H.A.; AlMukdad, S.; Coyle, P.; Ayoub, H.H.; Al Kanaani, Z. BNT162b2 and mRNA-1273 COVID-19 vaccine effectiveness against the SARS-CoV-2 Delta variant in Qatar. Nat. Med. 2021, 27, 2136–2143. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Marc, G.P.; Moreira, E.D.; Zerbini, C. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 31, 2603–2615. [Google Scholar] [CrossRef]
- Williams, S.V.; Vusirikala, A.; Ladhani, S.N.; De Olano, E.F.R.; Iyanger, N.; Aiano, F.; Stoker, K.; Rao, G.G.; John, L.; Patel, B. An outbreak caused by the SARS-CoV-2 Delta (B. 1.617. 2) variant in a care home after partial vaccination with a single dose of the COVID-19 vaccine Vaxzevria, London, England, April 2021. Eurosurveillance 2021, 26, 2100626. [Google Scholar] [CrossRef] [PubMed]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B. Efficacy and safety of the mRNA-1273 SARS-CoV-2 vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Logunov, D.Y.; Dolzhikova, I.V.; Shcheblyakov, D.V.; Tukhvatulin, A.I.; Zubkova, O.V.; Dzharullaeva, A.S.; Kovyrshina, A.V.; Lubenets, N.L.; Grousova, D.M.; Erokhova, A.S. Safety and efficacy of an rAd26 and rAd5 vector-based heterologous prime-boost COVID-19 vaccine: An interim analysis of a randomised controlled phase 3 trial in Russia. Lancet 2021, 397, 671–681. [Google Scholar] [CrossRef]
- Mlcochova, P.; Kemp, S.A.; Dhar, M.S.; Papa, G.; Meng, B.; Ferreira, I.A.; Datir, R.; Collier, D.A.; Albecka, A.; Singh, S. SARS-CoV-2 B. 1.617. 2 Delta variant replication and immune evasion. Nature 2021, 599, 114–119. [Google Scholar] [CrossRef]
- Planas, D.; Veyer, D.; Baidaliuk, A.; Staropoli, I.; Guivel-Benhassine, F.; Rajah, M.M.; Planchais, C.; Porrot, F.; Robillard, N.; Puech, J. Reduced sensitivity of SARS-CoV-2 variant Delta to antibody neutralization. Nature 2021, 596, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.-L.; Wang, Z.-Y.; Duan, L.-J.; Meng, Q.-C.; Jiang, M.-D.; Cao, J.; Yao, L.; Zhu, K.-L.; Cao, W.-C.; Ma, M.-J. Susceptibility of circulating SARS-CoV-2 variants to neutralization. N. Engl. J. Med. 2021, 384, 2354–2356. [Google Scholar] [CrossRef]
- Salvatore, M.; Bhattacharyya, R.; Purkayastha, S.; Zimmermann, L.; Ray, D.; Hazra, A.; Kleinsasser, M.; Mellan, T.A.; Whittaker, C.; Flaxman, S. Resurgence of SARS-CoV-2 in India: Potential role of the B. 1.617. 2 (Delta) variant and delayed interventions. medRxiv 2021. [Google Scholar] [CrossRef]
- Mohammadpour, H.; Ziai, A.; Sadr, M.; Rezaei, M.; Marjani, M.; Tabarsi, P. A novel coronavirus disease (COVID-19): A review of host cell signaling pathways. Tanaffos 2020, 19, 108. [Google Scholar]
- Luo, C.H.; Morris, C.P.; Sachithanandham, J.; Amadi, A.; Gaston, D.; Li, M.; Swanson, N.J.; Schwartz, M.; Klein, E.Y.; Pekosz, A. Infection with the SARS-CoV-2 delta variant is associated with higher infectious virus loads compared to the alpha variant in both unvaccinated and vaccinated individuals. medRxiv 2021. [Google Scholar] [CrossRef]
- Singanayagam, A.; Hakki, S.; Dunning, J.; Madon, K.J.; Crone, M.A.; Koycheva, A.; Derqui-Fernandez, N.; Barnett, J.L.; Whitfield, M.G.; Varro, R. Community transmission and viral load kinetics of the SARS-CoV-2 delta (B. 1.617. 2) variant in vaccinated and unvaccinated individuals in the UK: A prospective, longitudinal, cohort study. Lancet Infect. Dis. 2021, 22, 183–195. [Google Scholar] [CrossRef]
- Mohapatra, R.K.; Tiwari, R.; Sarangi, A.K.; Sharma, S.K.; Khandia, R.; Saikumar, G.; Dhama, K. Twin combination of Omicron and Delta variant triggering a Tsunami wave of ever high surges in COVID-19 cases: A challenging global threat with a special focus on Indian sub-continent. J. Med. Virol. 2022, 94, 1761–1765. [Google Scholar] [CrossRef]
- WHO. Classification of Omicron (B.1.1.529): SARS-CoV-2 Variant of Concern. Available online: https://www.who.int/news/item/26-11-2021-classification-of-omicron-(b.1.1.529)-sars-cov-2-variant-of-concern (accessed on 10 January 2022).
- Kim, S.; Nguyen, T.T.; Taitt, A.S.; Jhun, H.; Park, H.-Y.; Kim, S.-H.; Kim, Y.-G.; Song, E.Y.; Lee, Y.; Yum, H. SARS-CoV-2 Omicron Mutation Is Faster than the Chase: Multiple Mutations on Spike/ACE2 Interaction Residues. Immune Netw. 2021, 21, e38. [Google Scholar] [CrossRef]
- ELPAIS. Omicron: ‘The Fastest-Spreading Virus in History’. Available online: https://english.elpais.com/usa/2022-01-03/omicron-the-fastest-spreading-virus-in-history.html (accessed on 11 January 2022).
- Quarleri, J.; Galvan, V.; Delpino, M. Omicron Variant of the SARS-CoV-2: A Quest to Define the Consequences of Its High Mutational Load; Springer: Berlin/Heidelberg, Germany, 2021; pp. 1–4. [Google Scholar]
- Desingu, P.A.; Nagarajan, K.; Dhama, K. Emergence of Omicron third lineage BA. 3 and its importance. J. Med. Virol. 2022, 94, 1808–1810. [Google Scholar] [CrossRef]
- Guo, Q.; Ruhan, A.; Liang, L.; Zhao, X.; Deng, A.; Hu, Y.; Zheng, Z.; Zou, L.; Chen, C.; Li, B. An Imported Case of BA. 2 Lineage of Omicron Variant COVID-19—Guangdong Province, China, December 28, 2021. China CDC Wkly. 2022, 4, 98–99. [Google Scholar] [CrossRef]
- Kumar, S.; Karuppanan, K.; Subramaniam, G. Omicron (BA. 1) and Sub-Variants (BA. 1, BA. 2 and BA. 3) of SARS-CoV-2 Spike Infectivity and Pathogenicity: A Comparative Sequence and Structural-based Computational Assessment. bioRxiv 2022. [Google Scholar] [CrossRef]
- WHO. Contact Tracing and Quarantine in the Context of the Omicron SARS-CoV-2 Variant. Available online: https://apps.who.int/iris/rest/bitstreams/1411031/retrieve (accessed on 21 February 2022).
- Nature. Why Does the Omicron Sub-Variant Spread Faster than the Original? Available online: https://www.nature.com/articles/d41586-022-00471-2 (accessed on 21 February 2022).
- Mukherjee, A.G.; Wanjari, U.R.; Murali, R.; Chaudhary, U.; Renu, K.; Madhyastha, H.; Iyer, M.; Vellingiri, B.; Gopalakrishnan, A.V. Omicron variant infection and the associated immunological scenario. Immunobiology 2022, 4, 152222. [Google Scholar] [CrossRef]
- Team, W.W. WHO tracks two new Omicron subvariants: What are BA.4 and BA.5 strains? CNBC, 15 April 2022. Available online: https://www.cnbc.com/2022/04/15/who-tracks-omicron-bapoint4-and-bapoint5-subvariants-as-they-spread-through-africa-and-europe.html (accessed on 15 April 2022).
- Kupferschmidt, K. Where Did ‘Weird’ Omicron Come from? American Association for the Advancement of Science: Washington, DC, USA, 2021. [Google Scholar]
- Ren, S.-Y.; Wang, W.-B.; Gao, R.-D.; Zhou, A.-M. Omicron variant (B. 1.1. 529) of SARS-CoV-2: Mutation, infectivity, transmission, and vaccine resistance. World J. Clin. Cases 2022, 10, 1. [Google Scholar] [CrossRef]
- WHO. Update on SARS-CoV-2 Variant of Concern Omicron. Available online: https://www.who.int/news-room/events/detail/2022/02/01/default-calendar/update-on-sars-cov-2-variant-of-concern-omicron (accessed on 2 February 2022).
- Gong, S.Y.; Chatterjee, D.; Richard, J.; Prévost, J.; Tauzin, A.; Gasser, R.; Bo, Y.; Vézina, D.; Goyette, G.; Gendron-Lepage, G. Contribution of single mutations to selected SARS-CoV-2 emerging variants spike antigenicity. Virology 2021, 563, 134–145. [Google Scholar] [CrossRef]
- Times, T.N.Y. Omicron’s spread drives the global daily average of known new cases past a staggering 2 million. The New York Times, 8 January 2022. [Google Scholar]
- Chowdhury, S.; Bappy, M.H.; Chowdhury, S.; Chowdhury, M.S.; Chowdhury, N.S. Omicron Variant (B. 1.1. 529) of SARS-CoV-2, A Worldwide Public Health Emergency! Eur. J. Clin. Med. 2022, 3, 5–9. [Google Scholar] [CrossRef]
- CDC. COVID Data Tracker. Available online: https://covid.cdc.gov/covid-data-tracker/ (accessed on 31 January 2022).
- Ranjan, R. Omicron Impact in India: An Early Analysis of the Ongoing COVID-19 Third Wave. medRxiv 2022. [Google Scholar] [CrossRef]
- Foxnews. Omicron Is Better at Evading Vaccines, New COVID-19 Transmissibility Data Confirms. Available online: https://www.foxnews.com/world/omicron-vaccines-new-covid-transmissibility-data (accessed on 15 May 2022).
- Matsumaya, K. A New Study Suggests That the Omicron Variant is over Four Times More Transmissible in Its Early Stage than Delta. Available online: https://fortune.com/2021/12/08/omicron-covid-variant-data-more-transmissible-than-delta-new-study/ (accessed on 25 January 2022).
- Abu-Raddad, L.J.; Chemaitelly, H.; Ayoub, H.H.; AlMukdad, S.; Tang, P.; Hasan, M.R.; Coyle, P.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K. Effectiveness of BNT162b2 and mRNA-1273 COVID-19 boosters against SARS-CoV-2 Omicron (B. 1.1. 529) infection in Qatar. medRxiv 2022. [Google Scholar] [CrossRef]
- Chemaitelly, H.; Ayoub, H.H.; AlMukdad, S.; Tang, P.; Hasan, M.R.; Yassine, H.M.; Al Khatib, H.A.; Smatti, M.K.; Coyle, P.; Al Kanaani, Z. Duration of protection of BNT162b2 and mRNA-1273 COVID-19 vaccines against symptomatic SARS-CoV-2 Omicron infection in Qatar. medRxiv 2022. [Google Scholar] [CrossRef]
- Nishiura, H.; Ito, K.; Anzai, A.; Kobayashi, T.; Piantham, C.; Rodríguez-Morales, A.J. Relative Reproduction Number of SARS-CoV-2 Omicron (B. 1.1. 529) Compared with Delta Variant in South Africa. J. Clin. Med. 2022, 11, 30. [Google Scholar] [CrossRef]
- Lyngse, F.P.; Mortensen, L.H.; Denwood, M.J.; Christiansen, L.E.; Møller, C.H.; Skov, R.L.; Spiess, K.; Fomsgaard, A.; Lassauniere, R.; Rasmussen, M. SARS-CoV-2 Omicron VOC Transmission in Danish Households. medRxiv 2021. [Google Scholar] [CrossRef]
- Statista. Number of SARS-CoV-2 Omicron Variant Cases Worldwide as of February 3, 2022, by Country or Territory. Available online: https://www.statista.com/statistics/1279100/number-omicron-variant-worldwide-by-country/ (accessed on 6 February 2022).
- Lyngse, F.P.; Kirkeby, C.T.; Denwood, M.; Christiansen, L.E.; Mølbak, K.; Møller, C.H.; Skov, R.L.; Krause, T.G.; Rasmussen, M.; Sieber, R.N. Transmission of SARS-CoV-2 Omicron VOC subvariants BA. 1 and BA. 2: Evidence from Danish Households. medRxiv 2022. [Google Scholar] [CrossRef]
- Bi, K.; Herrera-Diestra, J.L.; Bai, Y.; Du, Z.; Wang, L.; Gibson, G.; Johnson-Leon, M.; Fox, S.; Meyers, L.A. The risk of SARS-CoV-2 Omicron variant emergence in low and middle-income countries (LMICs). medRxiv 2022. [Google Scholar] [CrossRef]
- Benvenuto, D.; Angeletti, S.; Giovanetti, M.; Bianchi, M.; Pascarella, S.; Cauda, R.; Ciccozzi, M.; Cassone, A. Evolutionary analysis of SARS-CoV-2: How mutation of Non-Structural Protein 6 (NSP6) could affect viral autophagy. J. Infect. 2020, 81, e24–e27. [Google Scholar] [CrossRef]
- Pang, X.; Li, P.; Zhang, L.; Que, L.; Dong, M.; Xie, B.; Wang, Q.; Wei, Y.; Xie, X.; Li, L. Emerging SARS-CoV-2 mutation hotspots associated with clinical outcomes and transmission. Front. Microbiol. 2021, 12, 3047. [Google Scholar] [CrossRef]
- Li, W.; Shi, Z.; Yu, M.; Ren, W.; Smith, C.; Epstein, J.H.; Wang, H.; Crameri, G.; Hu, Z.; Zhang, H. Bats are natural reservoirs of SARS-like coronaviruses. Science 2005, 310, 676–679. [Google Scholar] [CrossRef]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Gozzi, N.; Chinazzi, M.; Davis, J.T.; Mu, K.; y Piontti, A.P.; Vespignani, A.; Perra, N. Preliminary modeling estimates of the relative transmissibility and immune escape of the Omicron SARS-CoV-2 variant of concern in South Africa. medRxiv 2022. [Google Scholar] [CrossRef]
- Pulliam, J.R.; van Schalkwyk, C.; Govender, N.; von Gottberg, A.; Cohen, C.; Groome, M.J.; Dushoff, J.; Mlisana, K.; Moultrie, H. Increased risk of SARS-CoV-2 reinfection associated with emergence of the Omicron variant in South Africa. medRxiv 2021. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: Omicron and the need for boosters. BMJ 2021, 375, n3079. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Gilby, N.B.; Wei, G.-W. Omicron (B. 1.1. 529): Infectivity, vaccine breakthrough, and antibody resistance. arXiv 2021, arXiv:2112.01318v1. [Google Scholar]
- London, I.C. Report 49—Growth, Population Distribution and Immune Escape of Omicron in England. Available online: https://www.imperial.ac.uk/mrc-global-infectious-disease-analysis/covid-19/report-49-Omicron/ (accessed on 26 January 2022).
- Callaway, E.; Ledford, H. How bad is Omicron? What scientists know so far. Nature 2021, 600, 197–199. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, A.; Kerr, S.; Woolhouse, M.; McMenamin, J.; Robertson, C. Severity of Omicron variant of concern and vaccine effectiveness against symptomatic disease: National cohort with nested test negative design study in Scotland. Lancet Infect. Dis. 2021; Preprint. [Google Scholar] [CrossRef]
- Chan, M.C.; Hui, K.P.; Ho, J.; Cheung, M.-C.; Ng, K.-C.; Ching, R.; Lai, K.-L.; Kam, T.; Gu, H.; Sit, K.-Y. SARS-CoV-2 Omicron variant replication in human respiratory tract ex vivo. Nature 2021, 603, 715–720. [Google Scholar]
- Ulloa, A.C.; Buchan, S.A.; Daneman, N.; Brown, K.A. Early estimates of SARS-CoV-2 Omicron variant severity based on a matched cohort study, Ontario, Canada. medRxiv 2021. [Google Scholar] [CrossRef]
- Menni, C.; Valdes, A.M.; Polidori, L.; Antonelli, M.; Penamakuri, S.; Nogal, A.; Louca, P.; May, A.; Figueiredo, J.C.; Hu, C. A comparison of SARS-CoV-2 symptom prevalence, duration, and risk of hospitalisation among vaccinated individuals from the ZOE COVID Study during omicron vs. delta variant dominant periods. Lancet 2022, 399, 453–456. [Google Scholar] [CrossRef]
- Houhamdi, L.; Gautret, P.; Hoang, V.T.; Fournier, P.E.; Colson, P.; Raoult, D. Characteristics of the first 1119 SARS-CoV-2 Omicron variant cases, in Marseille, France, November− December 2021. J. Med. Virol. 2022, 94, 2290–2295. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention (U.S.). SARS-CoV-2 Infection and Hospitalization Among Adults Aged ≥18 Years, by Vaccination Status, Before and During SARS-CoV-2 B. 1.1. 529 (Omicron) Variant Predominance—Los Angeles County, California, November 7, 2021–January 8, 2022. Available online: https://stacks.cdc.gov/view/cdc/113859 (accessed on 1 February 2022).
- GHN. Omicron in the US: An Avoidable Tragedy. Available online: https://www.globalhealthnow.org/2022-02/omicron-us-avoidable-tragedy (accessed on 28 February 2022).
- Kim, M.-K.; Lee, B.; Choi, Y.Y.; Um, J.; Lee, K.-S.; Sung, H.K.; Kim, Y.; Park, J.-S.; Lee, M.; Jang, H.-C. Clinical Characteristics of 40 Patients Infected With the SARS-CoV-2 Omicron Variant in Korea. J. Korean Med. Sci. 2022, 37, e31. [Google Scholar] [CrossRef]
- Christie, B. COVID-19: Early studies give hope omicron is milder than other variants. BMJ 2021, 375, n3144. [Google Scholar] [CrossRef]
- Mahase, E. COVID-19: Hospital admission 50–70% less likely with omicron than delta, but transmission a major concern. BMJ 2021, 375, n3151. [Google Scholar] [CrossRef]
- Karim, S.S.A.; Karim, Q.A. Omicron SARS-CoV-2 variant: A new chapter in the COVID-19 pandemic. Lancet 2021, 398, 2126–2128. [Google Scholar] [CrossRef]
- Kannan, S.; Sheeza, A. Omicron (B. 1.1. 529)-variant of concern-molecular profile and epidemiology: A mini review. Eur. Rev. Med. Pharmacol. Sci. 2021, 25, 8019–8022. [Google Scholar]
- Zahradník, J.; Marciano, S.; Shemesh, M.; Zoler, E.; Harari, D.; Chiaravalli, J.; Meyer, B.; Rudich, Y.; Li, C.; Marton, I. SARS-CoV-2 variant prediction and antiviral drug design are enabled by RBD in vitro evolution. Nat. Microbiol. 2021, 6, 1188–1198. [Google Scholar] [CrossRef]
- Khan, A.; Waris, H.; Rafique, M.; Suleman, M.; Mohammad, A.; Ali, S.S.; Khan, T.; Waheed, Y.; Liao, C.; Wei, D.-Q. The Omicron (B. 1.1. 529) variant of SARS-CoV-2 binds to the hACE2 receptor more strongly and escapes the antibody response: Insights from structural and simulation data. Int. J. Biol. Macromol. 2022, 200, 438–448. [Google Scholar] [CrossRef]
- Wang, R.; Chen, J.; Wei, G.-W. Mechanisms of SARS-CoV-2 evolution revealing vaccine-resistant mutations in Europe and America. J. Phys. Chem. Lett. 2021, 12, 11850–11857. [Google Scholar] [CrossRef]
- Leary, S.; Gaudieri, S.; Parker, M.D.; Chopra, A.; James, I.; Pakala, S.; Alves, E.; John, M.; Lindsey, B.B.; Keeley, A.J. Generation of a novel SARS-CoV-2 sub-genomic RNA due to the R203K/G204R variant in nucleocapsid. bioRxiv 2021. [Google Scholar] [CrossRef]
- Mourier, T.; Shuaib, M.; Hala, S.; Mfarrej, S.; Alofi, F.; Naeem, R.; Alsomali, A.; Jorgensen, D.; Subudhi, A.; Ben Rached, F. Saudi Arabian SARS-CoV-2 genomes implicate a mutant Nucleocapsid protein in modulating host interactions and increased viral load in COVID-19 patients. medRxiv 2021. [Google Scholar] [CrossRef]
- Abdelnabi, R.; Foo, C.S.; Zhang, X.; Lemmens, V.; Maes, P.; Slechten, B.; Raymenants, J.; André, E.; Weynand, B.; Dallemier, K. The omicron (B. 1.1. 529) SARS-CoV-2 variant of concern does not readily infect Syrian hamsters. Antivir. Res. 2022, 198, 105253. [Google Scholar] [CrossRef] [PubMed]
- Hay, J.A.; Kissler, S.M.; Fauver, J.R.; Mack, C.; Tai, C.G.; Samant, R.M.; Connelly, S.; Anderson, D.J.; Khullar, G.; MacKay, M. Viral dynamics and duration of PCR positivity of the SARS-CoV-2 Omicron variant. medRxiv 2022. [Google Scholar] [CrossRef]
- Sentis, C.; Billaud, G.; Bal, A.; Frobert, E.; Bouscambert, M.; Destras, G.; Josset, L.; Lina, B.; Morfin, F.; Gaymard, A. SARS-CoV-2 Omicron variant, lineage BA. 1, is associated with lower viral load in nasopharyngeal samples compared to Delta variant. Viruses 2022, 14, 919. [Google Scholar] [CrossRef] [PubMed]
- Meyer, B.; Puhach, O.; Adea, K.; Hulo, N.; Sattonnet, P.; Genecand, C.; Iten, A.; Bausch, F.J.; Kaiser, L.; Vetter, P. Infectious viral load in unvaccinated and vaccinated patients infected with SARS-CoV-2 WT, Delta and Omicron. Nat. Med. 2022; online ahead of print. [Google Scholar] [CrossRef]
- Rossouw, L. Estimating the Effective Reproduction Number of COVID-19 in South Africa. Unsupervized. 2021. Available online: https://unsupervised.online/static/covid-19/estimating_r_za.html (accessed on 20 April 2022).
- Kim, D.; Jo, J.; Lim, J.-S.; Ryu, S. Serial interval and basic reproduction number of SARS-CoV-2 Omicron variant in South Korea. medRxiv 2021. [Google Scholar] [CrossRef]
- Ito, K.; Piantham, C.; Nishiura, H. Relative instantaneous reproduction number of Omicron SARS-CoV-2 variant with respect to the Delta variant in Denmark. J. Med. Virol. 2021, 94, 2265–2268. [Google Scholar] [CrossRef]
- Hladish, T.J.; Pillai, A.N.; Longini, I.M. Updated projections for COVID-19 omicron wave in Florida. medRxiv 2022. [Google Scholar] [CrossRef]
- Stefanelli, P.; Trentini, F.; Petrone, D.; Mammone, A.; Ambrosio, L.; Manica, M.; Guzzetta, G.; Andrea, V.D.; Marziano, V.; Zardini, A. Tracking the progressive spread of the SARS-CoV-2 Omicron variant in Italy, December 2021-January 2022. medRxiv 2022. [Google Scholar] [CrossRef]
- Ranjan, R. Omicron Impact in India: Analysis of the Ongoing COVID-19 Third Wave Based on Global Data. medRxiv 2022. [Google Scholar] [CrossRef]
- Chen, J.; Wang, R.; Gilby, N.B.; Wei, G.-W. Omicron variant (B. 1.1. 529): Infectivity, vaccine breakthrough, and antibody resistance. J. Chem. Inf. Model. 2022, 62, 412–422. [Google Scholar] [CrossRef] [PubMed]
- Escalera, A.; Gonzalez-Reiche, A.S.; Aslam, S.; Mena, I.; Pearl, R.L.; Laporte, M.; Fossati, A.; Rathnasinghe, R.; Alshammary, H.; van de Guchte, A. SARS-CoV-2 variants of concern have acquired mutations associated with an increased spike cleavage. bioRxiv 2021, preprint. [Google Scholar] [CrossRef]
- Kannan, S.R.; Spratt, A.N.; Sharma, K.; Chand, H.S.; Byrareddy, S.N.; Singh, K. Omicron SARS-CoV-2 variant: Unique features and their impact on pre-existing antibodies. J. Autoimmun. 2022, 126, 102779. [Google Scholar] [CrossRef]
- Kozlov, M. Omicron overpowers key COVID antibody treatments in early tests. Nature, 2021; ahead of print. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Wu, B.; Yang, Q.; Chen, A.; Li, Y.; Zhang, Y.; Pan, T.; Zhang, H.; He, X. SARS-CoV-2 Omicron strain exhibits potent capabilities for immune evasion and viral entrance. Signal Transduct. Target. Ther. 2021, 6, 430. [Google Scholar] [CrossRef] [PubMed]
- Callaway, E. Omicron likely to weaken COVID vaccine protection. Nature 2021, 600, 367–368. [Google Scholar] [CrossRef]
- Cross, R. Omicron puts scientists on red alert. Chem. Eng. News 2021, 99, 6. [Google Scholar] [CrossRef]
- Holmes, L., Jr.; Dabney, K.W.; Ogundele, B.; Chavan, P.; Adhikari, R.; Halloran, D.; Ogungbade, G.; Mellekate, V.S.; Nelson, C.; Williams, J. SARS-CoV-2 Omicron Variant Exponential Transmission & Mitigation Dynamics: Translational Immuno-Epidemiologic Perspective. 2021. Available online: http://dx.doi.org/10.2139/ssrn.3993979 (accessed on 5 February 2022).
- Collie, S.; Champion, J.; Moultrie, H.; Bekker, L.-G.; Gray, G. Effectiveness of BNT162b2 vaccine against omicron variant in South Africa. N. Engl. J. Med. 2021, 385, 494–496. [Google Scholar] [CrossRef]
- Rosenberg, E.S.; Dorabawila, V.; Easton, D.; Bauer, U.E.; Kumar, J.; Hoen, R.; Hoefer, D.; Wu, M.; Lutterloh, E.; Conroy, M.B. COVID-19 vaccine effectiveness in New York state. N. Engl. J. Med. 2022, 386, 116–127. [Google Scholar] [CrossRef]
- Ai, J.; Zhang, H.; Zhang, Y.; Lin, K.; Zhang, Y.; Wu, J.; Wan, Y.; Huang, Y.; Song, J.; Fu, Z. Omicron variant showed lower neutralizing sensitivity than other SARS-CoV-2 variants to immune sera elicited by vaccines after boost. Emerg. Microb. Infect. 2021, 11, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Pearson, C.A.; Silal, S.P.; Li, M.W.; Dushoff, J.; Bolker, B.M.; Abbott, S.; van Schalkwyk, C.; Davies, N.G.; Barnard, R.C.; Edmunds, W.J. Bounding the levels of transmissibility & immune evasion of the Omicron variant in South Africa. medRxiv 2021. [Google Scholar] [CrossRef]
- Agency, U.H.S. SARS-CoV-2 Variants of Concern and Variants under Investigation in England: Technical Briefing 33. Available online: https://assets.publishing.service.gov.uk/government/uploads/system/uploads/attachment_data/file/1043680/technical-briefing-33.pdf (accessed on 31 January 2022).
- Veneti, L.; Bøås, H.; Kristoffersen, A.B.; Stålcrantz, J.; Bragstad, K.; Hungnes, O.; Storm, M.L.; Aasand, N.; Rø, G.; Starrfelt, J. Reduced risk of hospitalisation among reported COVID-19 cases infected with the SARS-CoV-2 Omicron BA. 1 variant compared with the Delta variant, Norway, December 2021 to January 2022. Eurosurveillance 2022, 27, 2200077. [Google Scholar] [CrossRef]
- WHO. Coronavirus disease (COVID-19) Weekly Epidemiological Update and Weekly Operational Update. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019/situation-reports (accessed on 20 February 2022).
- Lee, J.J.; Choe, Y.J.; Jeong, H.; Kim, M.; Kim, S.; Yoo, H.; Park, K.; Kim, C.; Choi, S.; Sim, J. Importation and transmission of SARS-CoV-2 B. 1.1. 529 (Omicron) variant of concern in Korea, November 2021. J. Korean Med. Sci. 2021, 36, e346. [Google Scholar] [CrossRef]
- Zhang, X.; Maggioni, V.; Houser, P.; Xue, Y.; Mei, Y. The impact of weather condition and social activity on COVID-19 transmission in the United States. J. Environ. Manag. 2022, 302, 114085. [Google Scholar] [CrossRef]
- Coccia, M. Effects of the spread of COVID-19 on public health of polluted cities: Results of the first wave for explaining the dejà vu in the second wave of COVID-19 pandemic and epidemics of future vital agents. Environ. Sci. Pollut. Res. 2021, 28, 19147–19154. [Google Scholar] [CrossRef]
- Coccia, M. The effects of atmospheric stability with low wind speed and of air pollution on the accelerated transmission dynamics of COVID-19. Int. J. Environ. Stud. 2021, 78, 1–27. [Google Scholar] [CrossRef]
- Domingo, J.L.; Marquès, M.; Rovira, J. Influence of airborne transmission of SARS-CoV-2 on COVID-19 pandemic. A review. Environ. Res. 2020, 188, 109861. [Google Scholar] [CrossRef] [PubMed]
- Bowe, B.; Xie, Y.; Gibson, A.K.; Cai, M.; van Donkelaar, A.; Martin, R.V.; Burnett, R.; Al-Aly, Z. Ambient fine particulate matter air pollution and the risk of hospitalization among COVID-19 positive individuals: Cohort study. Environ. Int. 2021, 154, 106564. [Google Scholar] [CrossRef]
- Raiteux, J.; Eschlimann, M.; Marangon, A.; Rogée, S.; Dadvisard, M.; Taysse, L.; Larigauderie, G. Inactivation of SARS-CoV-2 by Simulated Sunlight on Contaminated Surfaces. Microbiol. Spectr. 2021, 9, e0033321. [Google Scholar] [CrossRef]
- Shy, C.-G.; Lu, J.-H.; Lin, H.-C.; Hung, M.-N.; Chang, H.-C.; Lu, M.-L.; Chao, H.-R.; Chen, Y.-S.; Wang, P.-S. Rapid Control of a SARS-CoV-2 B. 1.617. 2 (Delta) Variant COVID-19 Community Outbreak: The Successful Experience in Pingtung County of Taiwan. Int. J. Environ. Res. Public Health 2022, 19, 1421. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, J.; Yao, J.; Zhang, X.; Li, L.; Xu, X.; He, X.; Wang, B.; Fu, S.; Niu, T. Impact of meteorological factors on the COVID-19 transmission: A multi-city study in China. Sci. Total Environ. 2020, 726, 138513. [Google Scholar] [CrossRef]
- Haque, S.E.; Rahman, M. Association between temperature, humidity, and COVID-19 outbreaks in Bangladesh. Environ. Sci. Policy 2020, 114, 253–255. [Google Scholar] [CrossRef]
- Wang, J.; Tang, K.; Feng, K.; Lin, X.; Lv, W.; Chen, K.; Wang, F. Impact of temperature and relative humidity on the transmission of COVID-19: A modelling study in China and the United States. BMJ Open 2021, 11, e043863. [Google Scholar] [CrossRef]
- Xie, J.; Zhu, Y. Association between ambient temperature and COVID-19 infection in 122 cities from China. Sci. Total Environ. 2020, 724, 138201. [Google Scholar] [CrossRef]
- Majumder, P.; Ray, P.P. A systematic review and meta-analysis on correlation of weather with COVID-19. Sci. Rep. 2021, 11, 10746. [Google Scholar] [CrossRef]
- Coccia, M. How do low wind speeds and high levels of air pollution support the spread of COVID-19? Atmos. Pollut. Res. 2021, 12, 437–445. [Google Scholar] [CrossRef]
- Tan, L.; Schultz, D.M. How Is COVID-19 Affected by Weather? Metaregression of 158 Studies and Recommendations for Best Practices in Future Research. Weather Clim. Soc. 2022, 14, 237–255. [Google Scholar] [CrossRef]
- Ansar, B.; Asmin, W.; Tosepu, R. Meteorological parameters and air pollutants affect the transmission of COVID-19: A review. IOP Conf. Ser. Mater. Sci. Eng. 2021, 1088, 012117. [Google Scholar]
- Leclerc, Q.J.; Fuller, N.M.; Knight, L.E.; Funk, S.; Knight, G.M.; Group, C.C.-W. What settings have been linked to SARS-CoV-2 transmission clusters? Wellcome Open Res. 2020, 5, 83. [Google Scholar] [CrossRef]
- Ladhani, S.N.; Ireland, G.; Baawuah, F.; Beckmann, J.; Okike, I.O.; Ahmad, S.; Garstang, J.; Brent, A.J.; Brent, B.; Aiano, F. Emergence of SARS-CoV-2 Alpha (B. 1.1. 7) variant, infection rates, antibody seroconversion and seroprevalence rates in secondary school students and staff: Active prospective surveillance, December 2020 to March 2021, England. J. Infect. 2021, 83, 573–580. [Google Scholar] [CrossRef]
- Mensah, A.A.; Sinnathamby, M.; Zaidi, A.; Coughlan, L.; Simmons, R.; Ismail, S.A.; Ramsay, M.E.; Saliba, V.; Ladhani, S.N. SARS-CoV-2 infections in children following the full re-opening of schools and the impact of national lockdown: Prospective, national observational cohort surveillance, July-December 2020, England. J. Infect. 2021, 82, 67–74. [Google Scholar] [CrossRef]
- Lam-Hine, T.; McCurdy, S.A.; Santora, L.; Duncan, L.; Corbett-Detig, R.; Kapusinszky, B.; Willis, M. Outbreak associated with SARS-CoV-2 B. 1.617. 2 (delta) variant in an elementary school—Marin County, California, May–June 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 1214. [Google Scholar] [CrossRef]
- Dougherty, K.; Mannell, M.; Naqvi, O.; Matson, D.; Stone, J. SARS-CoV-2 B. 1.617. 2 (Delta) variant COVID-19 outbreak associated with a gymnastics facility—Oklahoma, April–May 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 1004. [Google Scholar] [CrossRef]
- Ives, A.R.; Bozzuto, C. Estimating and explaining the spread of COVID-19 at the county level in the USA. Commun. Biol. 2021, 4, 60. [Google Scholar] [CrossRef]
- Ilardi, A.; Chieffi, S.; Iavarone, A.; Ilardi, C.R. SARS-CoV-2 in Italy: Population density correlates with morbidity and mortality. Jpn. J. Infect. Dis. 2020, 74, 61–64. [Google Scholar] [CrossRef]
- Jabłońska, K.; Aballéa, S.; Toumi, M. Factors influencing the COVID-19 daily deaths’ peak across European countries. Public Health 2021, 194, 135–142. [Google Scholar] [CrossRef]
- Aw, S.B.; Teh, B.T.; Ling, G.H.T.; Leng, P.C.; Chan, W.H.; Ahmad, M.H. The COVID-19 Pandemic Situation in Malaysia: Lessons Learned from the Perspective of Population Density. Int. J. Environ. Res. Public Health 2021, 18, 6566. [Google Scholar] [CrossRef]
- Chishinga, N.; Gandhi, N.R.; Onwubiko, U.N.; Telford, C.; Prieto, J.; Smith, S.; Chamberlain, A.T.; Khan, S.; Williams, S.; Khan, F. Characteristics and Risk Factors for Hospitalization and Mortality among Persons with COVID-19 in Atlanta Metropolitan Area. medRxiv 2020. [Google Scholar] [CrossRef]
- Hamidi, S.; Sabouri, S.; Ewing, R. Does density aggravate the COVID-19 pandemic? Early findings and lessons for planners. J. Am. Plan. Assoc. 2020, 86, 495–509. [Google Scholar] [CrossRef]
- Bhadra, A.; Mukherjee, A.; Sarkar, K. Impact of population density on COVID-19 infected and mortality rate in India. Model. Earth Syst. Environ. 2021, 7, 623–629. [Google Scholar] [CrossRef]
- WorldBank. Urban Density Is Not an Enemy in the Coronavirus Fight: Evidence from China. Available online: https://blogs.worldbank.org/sustainablecities/urban-density-not-enemy-coronavirus-fight-evidence-china (accessed on 3 February 2022).
- Modes, M.E.; Directo, M.P.; Melgar, M.; Johnson, L.R.; Yang, H.; Chaudhary, P.; Bartolini, S.; Kho, N.; Noble, P.W.; Isonaka, S. Clinical Characteristics and Outcomes Among Adults Hospitalized with Laboratory-Confirmed SARS-CoV-2 Infection During Periods of B. 1.617. 2 (Delta) and B. 1.1. 529 (Omicron) Variant Predominance—One Hospital, California, July 15–September 23, 2021, and December 21, 2021–January 27, 2022. Morb. Mortal. Wkly. Rep. 2022, 71, 217. [Google Scholar]
- Dowd, J.B.; Andriano, L.; Brazel, D.M.; Rotondi, V.; Block, P.; Ding, X.; Liu, Y.; Mills, M.C. Demographic science aids in understanding the spread and fatality rates of COVID-19. Proc. Natl. Acad. Sci. 2020, 117, 9696–9698. [Google Scholar] [CrossRef] [Green Version]
- Promislow, D.E. A geroscience perspective on COVID-19 mortality. J. Gerontol. Ser. A 2020, 75, e30–e33. [Google Scholar] [CrossRef] [Green Version]
- Conti, P.; Caraffa, A.; Gallenga, C.; Ross, R.; Kritas, S.; Frydas, I.; Younes, A.; Di Emidio, P.; Ronconi, G.; Toniato, E. IL-1 induces throboxane-A2 (TxA2) in COVID-19 causing inflammation and micro-thrombi: Inhibitory effect of the IL-1 receptor antagonist (IL-1Ra). J. Biol. Regul. Homeost. Agents 2020, 34, 1623–1627. [Google Scholar]
- CDC. Coronavirus disease 2019 in children—United States, february 12–april 2, 2020. Morb. Mortal. Wkly. Rep. 2020, 69, 422. [Google Scholar] [CrossRef]
- Parri, N.; Lenge, M.; Buonsenso, D. Children with COVID-19 in pediatric emergency departments in Italy. N. Engl. J. Med. 2020, 383, 187–190. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Zhang, L.; Du, H.; Zhang, J.; Li, Y.Y.; Qu, J.; Zhang, W.; Wang, Y.; Bao, S.; Li, Y. SARS-CoV-2 infection in children. N. Engl. J. Med. 2020, 382, 1663–1665. [Google Scholar] [CrossRef] [Green Version]
- Posfay-Barbe, K.M.; Wagner, N.; Gauthey, M.; Moussaoui, D.; Loevy, N.; Diana, A.; L’Huillier, A.G. COVID-19 in children and the dynamics of infection in families. Pediatrics 2020, 146, e20201576. [Google Scholar] [CrossRef] [PubMed]
- Heald-Sargent, T.; Muller, W.J.; Zheng, X.; Rippe, J.; Patel, A.B.; Kociolek, L.K. Age-related differences in nasopharyngeal severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) levels in patients with mild to moderate coronavirus disease 2019 (COVID-19). JAMA Pediatr. 2020, 174, 902–903. [Google Scholar] [CrossRef] [PubMed]
- Biswas, M.; Rahaman, S.; Biswas, T.K.; Haque, Z.; Ibrahim, B. Association of sex, age, and comorbidities with mortality in COVID-19 patients: A systematic review and meta-analysis. Intervirology 2021, 64, 36–47. [Google Scholar] [CrossRef]
- Bloomberg. The Delta Wave Is Tough on Kids, But Deadly for the Middle-Aged; Bloomberg Opinion: New York, NY, USA, 2021. [Google Scholar]
- Malden, D.E.; Bruxvoort, K.J.; Tseng, H.F.; Ackerson, B.; Choi, S.K.; Florea, A.; Tubert, J.; Takhar, H.; Aragones, M.; Hong, V. Distribution of SARS-CoV-2 Variants in a Large Integrated Health Care System—California, March–July 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 1415. [Google Scholar] [CrossRef] [PubMed]
- Hwang, H.; Lim, J.-S.; Song, S.-A.; Achangwa, C.; Sim, W.; Kim, G.; Ryu, S. Transmission dynamics of the Delta variant of SARS-CoV-2 infections in South Korea. J. Infect. Dis. 2021, 225, 793–799. [Google Scholar] [CrossRef]
- Kumar, A.; Asghar, A.; Raza, K.; Narayan, R.K.; Jha, R.K.; Satyam, A.; Kumar, G.; Dwivedi, P.; Sahni, C.; Kumari, C. Demographic characteristics of SARS-CoV-2 B. 1.617. 2 (Delta) variant infections in Indian population. medRxiv 2021. [Google Scholar] [CrossRef]
- Danza, P.; Koo, T.H.; Haddix, M.; Fisher, R.; Traub, E.; OYong, K.; Balter, S. SARS-CoV-2 infection and hospitalization among adults aged ≥18 Years, by vaccination status, before and during SARS-CoV-2 B. 1.1. 529 (omicron) variant predominance—Los angeles county, California, november 7, 2021–january 8, 2022. Morb. Mortal. Wkly. Rep. 2022, 71, 177. [Google Scholar] [CrossRef]
- Kozlov, M. Does Omicron Hit Kids Harder? Scientists Are Trying to Find out. Available online: https://www.nature.com/articles/d41586-022-00309-x (accessed on 6 February 2022).
- Bloomberg. COVID-19: Toddlers Make up 10% of Hospital Cases in South Africa’s Omicron Epicentre. Available online: https://gulfnews.com/world/africa/covid-19-toddlers-make-up-10-of-hospital-cases-in-south-africas-omicron-epicentre-1.84075071 (accessed on 31 January 2022).
- WHO. COVID-19 Disease in Children and Adolescents: Scientific Brief. Available online: https://www.who.int/publications/i/item/WHO-2019-nCoV-Sci_Brief-Children_and_adolescents-2021.1 (accessed on 22 February 2022).
- Munro, A.P.; Faust, S.N. Children are not COVID-19 super spreaders: Time to go back to school. Arch. Dis. Child. 2020, 105, 618–619. [Google Scholar] [CrossRef] [PubMed]
- Viner, R.M.; Mytton, O.T.; Bonell, C.; Melendez-Torres, G.; Ward, J.; Hudson, L.; Waddington, C.; Thomas, J.; Russell, S.; Van Der Klis, F. Susceptibility to SARS-CoV-2 infection among children and adolescents compared with adults: A systematic review and meta-analysis. JAMA Pediatr. 2021, 175, 143–156. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, P.; Curtis, N. Why is COVID-19 less severe in children? A review of the proposed mechanisms underlying the age-related difference in severity of SARS-CoV-2 infections. Arch. Dis. Child. 2021, 106, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Gallo Marin, B.; Aghagoli, G.; Lavine, K.; Yang, L.; Siff, E.J.; Chiang, S.S.; Salazar-Mather, T.P.; Dumenco, L.; Savaria, M.C.; Aung, S.N. Predictors of COVID-19 severity: A literature review. Rev. Med. Virol. 2021, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; McGoogan, J.M. Characteristics of and important lessons from the coronavirus disease 2019 (COVID-19) outbreak in China: Summary of a report of 72 314 cases from the Chinese Center for Disease Control and Prevention. JAMA 2020, 323, 1239–1242. [Google Scholar] [CrossRef]
- Zhou, F.; Yu, T.; Du, R.; Fan, G.; Liu, Y.; Liu, Z.; Xiang, J.; Wang, Y.; Song, B.; Gu, X. Clinical course and risk factors for mortality of adult inpatients with COVID-19 in Wuhan, China: A retrospective cohort study. Lancet 2020, 395, 1054–1062. [Google Scholar] [CrossRef]
- Wiersinga, W.J.; Rhodes, A.; Cheng, A.C.; Peacock, S.J.; Prescott, H.C. Pathophysiology, transmission, diagnosis, and treatment of coronavirus disease 2019 (COVID-19): A review. JAMA 2020, 324, 782–793. [Google Scholar] [CrossRef] [PubMed]
- Vestergaard, L.S.; Nielsen, J.; Richter, L.; Schmid, D.; Bustos, N.; Braeye, T.; Denissov, G.; Veideman, T.; Luomala, O.; Möttönen, T. Excess all-cause mortality during the COVID-19 pandemic in Europe–preliminary pooled estimates from the EuroMOMO network, March to April 2020. Eurosurveillance 2020, 25, 2001214. [Google Scholar] [CrossRef]
- Du, P.; Li, D.; Wang, A.; Shen, S.; Ma, Z.; Li, X. A systematic review and meta-analysis of risk factors associated with severity and death in COVID-19 patients. Can. J. Infect. Dis. Med. Microbiol. 2021, 2021, 6660930. [Google Scholar] [CrossRef]
- Grint, D.J.; Wing, K.; Williamson, E.; McDonald, H.I.; Bhaskaran, K.; Evans, D.; Evans, S.J.; Walker, A.J.; Hickman, G.; Nightingale, E. Case fatality risk of the SARS-CoV-2 variant of concern B. 1.1. 7 in England, 16 November to 5 February. Eurosurveillance 2021, 26, 2100256. [Google Scholar] [CrossRef] [PubMed]
- Hu, K.; Lin, L.; Liang, Y.; Shao, X.; Hu, Z.; Luo, H.; Lei, M. COVID-19: Risk factors for severe cases of the Delta variant. Aging 2021, 13, 23459. [Google Scholar] [CrossRef] [PubMed]
- Scobie, H.M.; Johnson, A.G.; Suthar, A.B.; Severson, R.; Alden, N.B.; Balter, S.; Bertolino, D.; Blythe, D.; Brady, S.; Cadwell, B. Monitoring incidence of COVID-19 cases, hospitalizations, and deaths, by vaccination status—13 US jurisdictions, April 4–July 17, 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 1284. [Google Scholar] [CrossRef] [PubMed]
- Budhiraja, S.; Indrayan, A.; Aggarwal, M.; Jha, V.; Jain, D.; Tarai, B.; Das, P.; Aggarwal, B.; Mishra, R.; Bali, S. Differentials in the characteristics of COVID-19 cases in Wave-1 and Wave-2 admitted to a network of hospitals in North India. medRxiv 2021. [Google Scholar] [CrossRef]
- Wang, L.; Berger, N.A.; Kaelber, D.C.; Davis, P.B.; Volkow, N.D.; Xu, R. Comparison of outcomes from COVID infection in pediatric and adult patients before and after the emergence of Omicron. medRxiv 2022. [Google Scholar] [CrossRef]
- Grasselli, G.; Zangrillo, A.; Zanella, A.; Antonelli, M.; Cabrini, L.; Castelli, A.; Cereda, D.; Coluccello, A.; Foti, G.; Fumagalli, R. Baseline characteristics and outcomes of 1591 patients infected with SARS-CoV-2 admitted to ICUs of the Lombardy Region, Italy. JAMA 2020, 323, 1574–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaeuffer, C.; Le Hyaric, C.; Fabacher, T.; Mootien, J.; Dervieux, B.; Ruch, Y.; Hugerot, A.; Zhu, Y.-J.; Pointurier, V.; Clere-Jehl, R. Clinical characteristics and risk factors associated with severe COVID-19: Prospective analysis of 1,045 hospitalised cases in North-Eastern France, March 2020. Eurosurveillance 2020, 25, 2000895. [Google Scholar] [CrossRef] [PubMed]
- de Souza, C.D.; de Arruda Magalhães, A.J.; Lima, A.J.; Nunes, D.N.; de Fátima Machado Soares, É.; de Castro Silva, L.; Santos, L.G.; dos Santos Cardoso, V.I.; Nobre, Y.V.; do Carmo, R.F. Clinical manifestations and factors associated with mortality from COVID-19 in older adults: Retrospective population-based study with 9807 older Brazilian COVID-19 patients. Geriatr. Gerontol. Int. 2020, 20, 1177–1181. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, N.T.; Chinn, J.; De Ferrante, M.; Kirby, K.A.; Hohmann, S.F.; Amin, A. Male gender is a predictor of higher mortality in hospitalized adults with COVID-19. PLoS ONE 2021, 16, e0254066. [Google Scholar] [CrossRef]
- WebMD. Omicron Affects Men and Women Differently, Research Indicates. Available online: https://playcrazygame.com/2022/01/26/omicron-affects-men-and-women-differently-research-indicates/ (accessed on 6 February 2022).
- Coopersmith, C.M.; Antonelli, M.; Bauer, S.R.; Deutschman, C.S.; Evans, L.E.; Ferrer, R.; Hellman, J.; Jog, S.; Kesecioglu, J.; Kissoon, N. The surviving sepsis campaign: Research priorities for coronavirus disease 2019 in critical illness. Crit. Care Med. 2021, 49, 598–622. [Google Scholar] [CrossRef]
- Cascella, M.; Rajnik, M.; Aleem, A.; Dulebohn, S.C.; Di Napoli, R. Features, evaluation, and treatment of coronavirus (COVID-19). In Statpearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Gandhi, R.T.; Lynch, J.B.; Del Rio, C. Mild or moderate COVID-19. N. Engl. J. Med. 2020, 383, 1757–1766. [Google Scholar] [CrossRef]
- Mølhave, M.; Agergaard, J.; Wejse, C. Clinical management of COVID-19 patients—An update. Semin. Nucl. Med. 2022, 52, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Group, R.C. Effect of hydroxychloroquine in hospitalized patients with COVID-19. N. Engl. J. Med. 2020, 383, 2030–2040. [Google Scholar]
- Zhang, R.; Mylonakis, E. In inpatients with COVID-19, none of remdesivir, hydroxychloroquine, lopinavir, or interferon β-1a differed from standard care for in-hospital mortality. Ann. Intern. Med. 2021, 174, JC17. [Google Scholar] [CrossRef]
- Mitjà, O.; Corbacho-Monné, M.; Ubals, M.; Alemany, A.; Suñer, C.; Tebé, C.; Tobias, A.; Peñafiel, J.; Ballana, E.; Pérez, C.A. A cluster-randomized trial of hydroxychloroquine for prevention of COVID-19. N. Engl. J. Med. 2020, 384, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Boulware, D.R.; Pullen, M.F.; Bangdiwala, A.S.; Pastick, K.A.; Lofgren, S.M.; Okafor, E.C.; Skipper, C.P.; Nascene, A.A.; Nicol, M.R.; Abassi, M. A randomized trial of hydroxychloroquine as postexposure prophylaxis for COVID-19. N. Engl. J. Med. 2020, 383, 517–525. [Google Scholar] [CrossRef]
- Beigel, J.; Tomashek, K.M.; Dodd, L.E.; Mehta, A.K.; Zingman, B.S.; Kalil, A.C.; Hohmann, E.; Chu, H.Y.; Luetkemeyer, A.; Kline, S. ACTT-1 Study Group Members. Remdesivir for the treatment of COVID-19—Final report. N. Engl. J. Med. 2020, 383, 1813–1826. [Google Scholar] [CrossRef] [PubMed]
- Goldman, J.D.; Lye, D.C.; Hui, D.S.; Marks, K.M.; Bruno, R.; Montejano, R.; Spinner, C.D.; Galli, M.; Ahn, M.-Y.; Nahass, R.G. Remdesivir for 5 or 10 days in patients with severe COVID-19. N. Engl. J. Med. 2020, 383, 1827–1837. [Google Scholar] [CrossRef]
- Spinner, C.D.; Gottlieb, R.L.; Criner, G.J.; López, J.R.A.; Cattelan, A.M.; Viladomiu, A.S.; Ogbuagu, O.; Malhotra, P.; Mullane, K.M.; Castagna, A. Effect of remdesivir vs standard care on clinical status at 11 days in patients with moderate COVID-19: A randomized clinical trial. JAMA 2020, 324, 1048–1057. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, R.L.; Vaca, C.E.; Paredes, R.; Mera, J.; Webb, B.J.; Perez, G.; Oguchi, G.; Ryan, P.; Nielsen, B.U.; Brown, M. Early remdesivir to prevent progression to severe COVID-19 in outpatients. N. Engl. J. Med. 2022, 386, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.K.; Singh, A.; Singh, R.; Misra, A. Molnupiravir in COVID-19: A systematic review of literature. Diabetes Metab. Syndr. Clin. Res. Rev. 2021, 15, 102329. [Google Scholar] [CrossRef]
- Joyner, M.J.; Senefeld, J.W.; Klassen, S.A.; Mills, J.R.; Johnson, P.W.; Theel, E.S.; Wiggins, C.C.; Bruno, K.A.; Klompas, A.M.; Lesser, E.R. Effect of convalescent plasma on mortality among hospitalized patients with COVID-19: Initial three-month experience. medrxiv 2020. [Google Scholar] [CrossRef]
- Simonovich, V.A.; Burgos Pratx, L.D.; Scibona, P.; Beruto, M.V.; Vallone, M.G.; Vázquez, C.; Savoy, N.; Giunta, D.H.; Pérez, L.G.; Sánchez, M.d.L. A randomized trial of convalescent plasma in COVID-19 severe pneumonia. N. Engl. J. Med. 2021, 384, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Zhang, W.; Hu, Y.; Tong, X.; Zheng, S.; Yang, J.; Kong, Y.; Ren, L.; Wei, Q.; Mei, H. Effect of convalescent plasma therapy on time to clinical improvement in patients with severe and life-threatening COVID-19: A randomized clinical trial. JAMA 2020, 324, 460–470. [Google Scholar] [CrossRef]
- Agarwal, A.; Mukherjee, A.; Kumar, G.; Chatterjee, P.; Bhatnagar, T.; Malhotra, P. Convalescent plasma in the management of moderate covid-19 in adults in India: Open label phase II multicentre randomised controlled trial (PLACID Trial). BMJ 2020, 371, m3939. [Google Scholar] [CrossRef] [PubMed]
- Weinreich, D.; Sivapalasingam, S.; Norton, T.; Ali, S.; Gao, H.; Bhore, R.; Musser, B.; Soo, Y.; Rofail, D.; Im, J.; et al. REGN-COV2, a Neutralizing Antibody Cocktail in Outpatients with COVID-19. N. Engl. J. Med. 2021, 384, 238–251. [Google Scholar] [PubMed]
- Gupta, A.; Gonzalez-Rojas, Y.; Juarez, E.; Crespo Casal, M.; Moya, J.; Falci, D.R.; Sarkis, E.; Solis, J.; Zheng, H.; Scott, N. Early treatment for COVID-19 with SARS-CoV-2 neutralizing antibody sotrovimab. N. Engl. J. Med. 2021, 385, 1941–1950. [Google Scholar] [CrossRef]
- WHO. Corticosteroids for COVID-19: Living Guidance, 2 September 2020; World Health Organization: Geneva, Switzerland, 2020. [Google Scholar]
- Group, R.C. Dexamethasone in hospitalized patients with COVID-19. N. Engl. J. Med. 2021, 384, 693–704. [Google Scholar]
- Stone, J.H.; Frigault, M.J.; Serling-Boyd, N.J.; Fernandes, A.D.; Harvey, L.; Foulkes, A.S.; Horick, N.K.; Healy, B.C.; Shah, R.; Bensaci, A.M. Efficacy of tocilizumab in patients hospitalized with COVID-19. N. Engl. J. Med. 2020, 383, 2333–2344. [Google Scholar] [CrossRef]
- Salvarani, C.; Dolci, G.; Massari, M.; Merlo, D.F.; Cavuto, S.; Savoldi, L.; Bruzzi, P.; Boni, F.; Braglia, L.; Turrà, C. Effect of tocilizumab vs standard care on clinical worsening in patients hospitalized with COVID-19 pneumonia: A randomized clinical trial. JAMA Intern. Med. 2021, 181, 24–31. [Google Scholar] [CrossRef]
- Salama, C.; Han, J.; Yau, L.; Reiss, W.G.; Kramer, B.; Neidhart, J.D.; Criner, G.J.; Kaplan-Lewis, E.; Baden, R.; Pandit, L. Tocilizumab in patients hospitalized with COVID-19 pneumonia. N. Engl. J. Med. 2021, 384, 20–30. [Google Scholar] [CrossRef]
- Gordon, A.C.; Mouncey, P.R.; Al-Beidh, F.; Rowan, K.M.; Nichol, A.D.; Arabi, Y.M.; Annane, D.; Beane, A.; van Bentum-Puijk, W.; Berry, L.R. Interleukin-6 receptor antagonists in critically ill patients with COVID-19. N. Engl. J. Med. 2021, 385, 1147–1149. [Google Scholar] [PubMed]
- Abani, O.; Abbas, A.; Abbas, F.; Abbas, M.; Abbasi, S.; Abbass, H.; Abbott, A.; Abdallah, N.; Abdelaziz, A.; Abdelfattah, M. Tocilizumab in patients admitted to hospital with COVID-19 (RECOVERY): A randomised, controlled, open-label, platform trial. Lancet 2021, 397, 1637–1645. [Google Scholar]
- Monk, P.D.; Marsden, R.J.; Tear, V.J.; Brookes, J.; Batten, T.N.; Mankowski, M.; Gabbay, F.J.; Davies, D.E.; Holgate, S.T.; Ho, L.-P. Safety and efficacy of inhaled nebulised interferon beta-1a (SNG001) for treatment of SARS-CoV-2 infection: A randomised, double-blind, placebo-controlled, phase 2 trial. Lancet Respir. Med. 2021, 9, 196–206. [Google Scholar] [CrossRef]
- Ranieri, V.M.; Pettilä, V.; Karvonen, M.K.; Jalkanen, J.; Nightingale, P.; Brealey, D.; Mancebo, J.; Ferrer, R.; Mercat, A.; Patroniti, N. Effect of intravenous interferon β-1a on death and days free from mechanical ventilation among patients with moderate to severe acute respiratory distress syndrome: A randomized clinical trial. JAMA 2020, 323, 725–733. [Google Scholar] [CrossRef] [PubMed]
- Davoudi-Monfared, E.; Rahmani, H.; Khalili, H.; Hajiabdolbaghi, M.; Salehi, M.; Abbasian, L.; Kazemzadeh, H.; Yekaninejad, M.S. A randomized clinical trial of the efficacy and safety of interferon β-1a in treatment of severe COVID-19. Antimicrob. Agents Chemother. 2020, 64, e01061-20. [Google Scholar] [CrossRef]
- Huet, T.; Beaussier, H.; Voisin, O.; Jouveshomme, S.; Dauriat, G.; Lazareth, I.; Sacco, E.; Naccache, J.-M.; Bézie, Y.; Laplanche, S. Anakinra for severe forms of COVID-19: A cohort study. Lancet Rheumatol. 2020, 2, e393–e400. [Google Scholar] [CrossRef]
- Yuen, C.-K.; Lam, J.-Y.; Wong, W.-M.; Mak, L.-F.; Wang, X.; Chu, H.; Cai, J.-P.; Jin, D.-Y.; To, K.K.-W.; Chan, J.F.-W. SARS-CoV-2 nsp13, nsp14, nsp15 and orf6 function as potent interferon antagonists. Emerg. Microb. Infect. 2020, 9, 1418–1428. [Google Scholar] [CrossRef]
- Zhou, H.; Yang, J.; Zhou, C.; Chen, B.; Fang, H.; Chen, S.; Zhang, X.; Wang, L.; Zhang, L. A Review of SARS-CoV2: Compared With SARS-CoV and MERS-CoV. Front. Med. 2021, 8, 628370. [Google Scholar] [CrossRef] [PubMed]
- Goujon, C.; Rebendenne, A.; Roy, P.; Bonaventure, B.; Valadao, A.C.; Desmarets, L.; Rouillé, Y.; Tauziet, M.; Arnaud-Arnould, M.; Giovannini, D. Bidirectional genome-wide CRISPR screens reveal host factors regulating SARS-CoV-2, MERS-CoV and seasonal HCoVs. Res. Sq. 2021, rs. 3, rs-555275. [Google Scholar]
- Ovsyannikova, I.G.; Haralambieva, I.H.; Crooke, S.N.; Poland, G.A.; Kennedy, R.B. The role of host genetics in the immune response to SARS-CoV-2 and COVID-19 susceptibility and severity. Immunol. Rev. 2020, 296, 205–219. [Google Scholar] [CrossRef]
- Riou, J.; Althaus, C. Pattern of early human-to-human transmission of Wuhan 2019 novel coronavirus (2019-nCoV), December 2019 to January 2020. Eurosurveillance 2020, 25, 2000058. [Google Scholar] [CrossRef]
- Abdelrahman, Z.; Li, M.; Wang, X. Comparative review of SARS-CoV-2, SARS-CoV, MERS-CoV, and influenza a respiratory viruses. Front. Immunol. 2020, 11, 55290. [Google Scholar] [CrossRef] [PubMed]
- WHO. Consensus Document on the Epidemiology of Severe Acute Respiratory Syndrome (SARS). Available online: https://www.who.int/csr/sars/WHOconsensus.pdf?ua=1 (accessed on 5 February 2022).
- Alfaraj, S.H.; Al-Tawfiq, J.A.; Assiri, A.Y.; Alzahrani, N.A.; Alanazi, A.A.; Memish, Z.A. Clinical predictors of mortality of Middle East Respiratory Syndrome Coronavirus (MERS-CoV) infection: A cohort study. Travel Med. Infect. Dis. 2019, 29, 48–50. [Google Scholar] [CrossRef] [PubMed]
- Al Kahlout, R.A.; Nasrallah, G.K.; Farag, E.A.; Wang, L.; Lattwein, E.; Müller, M.A.; El Zowalaty, M.E.; Al Romaihi, H.E.; Graham, B.S.; Al Thani, A.A. Comparative serological study for the prevalence of anti-MERS coronavirus antibodies in high-and low-risk groups in Qatar. J. Immunol. Res. 2019, 2019, 1386740. [Google Scholar] [CrossRef] [Green Version]
- Oboho, I.K.; Tomczyk, S.M.; Al-Asmari, A.M.; Banjar, A.A.; Al-Mugti, H.; Aloraini, M.S.; Alkhaldi, K.Z.; Almohammadi, E.L.; Alraddadi, B.M.; Gerber, S.I. 2014 MERS-CoV outbreak in Jeddah—A link to health care facilities. N. Engl. J. Med. 2015, 372, 846–854. [Google Scholar] [CrossRef]
- Hunter, J.C.; Nguyen, D.; Aden, B.; Al Bandar, Z.; Al Dhaheri, W.; Elkheir, K.A.; Khudair, A.; Al Mulla, M.; El Saleh, F.; Imambaccus, H. Transmission of Middle East respiratory syndrome coronavirus infections in healthcare settings, Abu Dhabi. Emerg. Infect. Dis. 2016, 22, 647. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.; Yang, T.; Jeong, Y.; Park, J.; Lee, K.; Kim, K. Middle East respiratory syndrome coronavirus outbreak in the Republic of Korea, 2015. Osong Public Health Res. Perspect 2016, 6, 269–278. [Google Scholar]
- Kim, J.Y.; Song, J.Y.; Yoon, Y.K.; Choi, S.-H.; Song, Y.G.; Kim, S.-R.; Son, H.-J.; Jeong, S.-Y.; Choi, J.-H.; Kim, K.M. Middle East respiratory syndrome infection control and prevention guideline for healthcare facilities. Infect. Chemother. 2015, 47, 278–302. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.Y.; Kang, J.-M.; Ha, Y.E.; Park, G.E.; Lee, J.Y.; Ko, J.-H.; Lee, J.Y.; Kim, J.M.; Kang, C.-I.; Jo, I.J. MERS-CoV outbreak following a single patient exposure in an emergency room in South Korea: An epidemiological outbreak study. Lancet 2016, 388, 994–1001. [Google Scholar] [CrossRef] [Green Version]
- Willman, M.; Kobasa, D.; Kindrachuk, J. A comparative analysis of factors influencing two outbreaks of Middle Eastern respiratory syndrome (MERS) in Saudi Arabia and South Korea. Viruses 2019, 11, 1119. [Google Scholar] [CrossRef] [Green Version]
- Kang, C.K.; Song, K.-H.; Choe, P.G.; Park, W.B.; Bang, J.H.; Kim, E.S.; Park, S.W.; Kim, H.B.; Kim, N.J.; Cho, S.-i. Clinical and epidemiologic characteristics of spreaders of Middle East respiratory syndrome coronavirus during the 2015 outbreak in Korea. J. Korean Med. Sci. 2017, 32, 744–749. [Google Scholar] [CrossRef] [PubMed]
- Payne, D.C.; Biggs, H.M.; Al-Abdallat, M.M.; Alqasrawi, S.; Lu, X.; Abedi, G.R.; Haddadin, A.; Iblan, I.; Alsanouri, T.; Al Nsour, M. Multihospital outbreak of a Middle East respiratory syndrome coronavirus deletion variant, Jordan: A molecular, serologic, and epidemiologic investigation. Open Forum Infect. Dis. 2018, 28, ofy095. [Google Scholar] [CrossRef] [Green Version]
- Hemida, M.G.; Alnaeem, A.; Chu, D.K.; Perera, R.A.; Chan, S.M.; Almathen, F.; Yau, E.; Ng, B.C.; Webby, R.J.; Poon, L.L. Longitudinal study of Middle East respiratory syndrome coronavirus infection in dromedary camel herds in Saudi Arabia, 2014–2015. Emerg. Microb. Infect. 2017, 6, e56. [Google Scholar] [CrossRef] [Green Version]
- Raj, V.S.; Farag, E.A.; Reusken, C.B.; Lamers, M.M.; Pas, S.D.; Voermans, J.; Smits, S.L.; Osterhaus, A.D.; Al-Mawlawi, N.; Al-Romaihi, H.E. Isolation of MERS coronavirus from a dromedary camel, Qatar, 2014. Emerg. Infect. Dis. 2014, 20, 1339. [Google Scholar] [CrossRef] [PubMed]
- Haagmans, B.L.; Al Dhahiry, S.H.; Reusken, C.B.; Raj, V.S.; Galiano, M.; Myers, R.; Godeke, G.-J.; Jonges, M.; Farag, E.; Diab, A. Middle East respiratory syndrome coronavirus in dromedary camels: An outbreak investigation. Lancet Infect. Dis. 2014, 14, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Liya, G.; Yuguang, W.; Jian, L.; Huaiping, Y.; Xue, H.; Jianwei, H.; Jiaju, M.; Youran, L.; Chen, M.; Yiqing, J. Studies on viral pneumonia related to novel coronavirus SARS-CoV-2, SARS-CoV, and MERS-CoV: A literature review. Apmis 2020, 128, 423–432. [Google Scholar] [CrossRef]
- Petersen, E.; Koopmans, M.; Go, U.; Hamer, D.H.; Petrosillo, N.; Castelli, F.; Storgaard, M.; Al Khalili, S.; Simonsen, L. Comparing SARS-CoV-2 with SARS-CoV and influenza pandemics. Lancet Infect. Dis. 2020, 20, e238–e244. [Google Scholar] [CrossRef]
- Bell, D.M. Public health interventions and SARS spread, 2003. Emerg. Infect. Dis. 2004, 10, 1900. [Google Scholar] [CrossRef] [Green Version]
- Chowell, G.; Castillo-Chavez, C.; Fenimore, P.W.; Kribs-Zaleta, C.M.; Arriola, L.; Hyman, J.M. Model parameters and outbreak control for SARS. Emerg. Infect. Dis. 2004, 10, 1258. [Google Scholar] [CrossRef] [PubMed]
- Lipsitch, M.; Cohen, T.; Cooper, B.; Robins, J.M.; Ma, S.; James, L.; Gopalakrishna, G.; Chew, S.K.; Tan, C.C.; Samore, M.H. Transmission dynamics and control of severe acute respiratory syndrome. Science 2003, 300, 1966–1970. [Google Scholar] [CrossRef] [Green Version]
- Breban, R.; Riou, J.; Fontanet, A. Interhuman transmissibility of Middle East respiratory syndrome coronavirus: Estimation of pandemic risk. Lancet 2013, 382, 694–699. [Google Scholar] [CrossRef] [Green Version]
- Cauchemez, S.; Fraser, C.; Van Kerkhove, M.D.; Donnelly, C.A.; Riley, S.; Rambaut, A.; Enouf, V.; van der Werf, S.; Ferguson, N.M. Middle East respiratory syndrome coronavirus: Quantification of the extent of the epidemic, surveillance biases, and transmissibility. Lancet Infect. Dis. 2014, 14, 50–56. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; Diao, M.; Yu, W.; Pei, L.; Lin, Z.; Chen, D. Estimation of the reproductive number of novel coronavirus (COVID-19) and the probable outbreak size on the Diamond Princess cruise ship: A data-driven analysis. Int. J. Infect. Dis. 2020, 93, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Jansen, L.; Tegomoh, B.; Lange, K.; Showalter, K.; Figliomeni, J.; Abdalhamid, B.; Iwen, P.C.; Fauver, J.; Buss, B.; Donahue, M. Investigation of a sars-cov-2 b. 1.1. 529 (omicron) variant cluster—Nebraska, november–december 2021. Morb. Mortal. Wkly. Rep. 2021, 70, 1782. [Google Scholar] [CrossRef] [PubMed]
- Grant, R.; Charmet, T.; Schaeffer, L.; Galmiche, S.; Madec, Y.; Von Platen, C.; Chény, O.; Omar, F.; David, C.; Rogoff, A. Impact of SARS-CoV-2 Delta variant on incubation, transmission settings and vaccine effectiveness: Results from a nationwide case-control study in France. Lancet Reg. Health-Eur. 2021, 13, 100278. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-E.; Jung, S.; Kim, A. MERS transmission and risk factors: A systematic review. BMC Public Health 2018, 18, 574. [Google Scholar] [CrossRef] [Green Version]
- Chathappady House, N.N.; Palissery, S.; Sebastian, H. Corona viruses: A review on SARS, MERS and COVID-19. Microbiol. Insights 2021, 14, 11786361211002481. [Google Scholar] [CrossRef]
- Hui, D.; Chan, M.; Wu, A.; Ng, P. Severe acute respiratory syndrome (SARS): Epidemiology and clinical features. Postgrad. Med. J. 2004, 80, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Al-Romaihi, H.E.; Smatti, M.K.; Ganesan, N.; Nadeem, S.; Farag, E.; Coyle, P.V.; Nader, J.D.; Al-Khatib, H.A.; Elmagboul, E.B.; Al Dhahry, S. Epidemiology of respiratory infections among adults in Qatar (2012–2017). PLoS ONE 2019, 14, e0218097. [Google Scholar] [CrossRef]
- Wolfel, R.; Corman, V.M.; Guggemos, W.; Seilmaier, M.; Zange, S.; Muller, M.A.; Niemeyer, D.; Jones, T.C.; Vollmar, P.; Rothe, C. Virological assessment of hospitalized patients with COVID-2019. Nature 2020, 581, 465–469, Erratum in Nature 2020, 588, E35. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.-S.; Zhang, C. What to do next to control the 2019-nCoV epidemic? Lancet 2020, 395, 391–393. [Google Scholar] [CrossRef] [Green Version]
- Cheng, P.K.; Wong, D.A.; Tong, L.K.; Ip, S.-M.; Lo, A.C.; Lau, C.-S.; Yeung, E.Y.; Lim, W.W. Viral shedding patterns of coronavirus in patients with probable severe acute respiratory syndrome. Lancet 2004, 363, 1699–1700. [Google Scholar] [CrossRef] [Green Version]
- Pan, Y.; Zhang, D.; Yang, P.; Poon, L.L.; Wang, Q. Viral load of SARS-CoV-2 in clinical samples. Lancet Infect. Dis. 2020, 20, 411–412. [Google Scholar] [CrossRef]
- Feikin, D.R.; Alraddadi, B.; Qutub, M.; Shabouni, O.; Curns, A.; Oboho, I.K.; Tomczyk, S.M.; Wolff, B.; Watson, J.T.; Madani, T.A. Association of higher MERS-CoV virus load with severe disease and death, Saudi Arabia, 2014. Emerg. Infect. Dis. 2015, 21, 2029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, M.-D.; Park, W.B.; Choe, P.G.; Choi, S.-J.; Kim, J.-I.; Chae, J.; Park, S.S.; Kim, E.-C.; Oh, H.S.; Kim, E.J. Viral load kinetics of MERS coronavirus infection. N. Engl. J. Med. 2016, 375, 1303–1305. [Google Scholar] [CrossRef] [PubMed]
- Bager, P.; Wohlfahrt, J.; Fonager, J.; Albertsen, M.; Yssing Michaelsen, T.; Holten Møller, C.; Ethelberg, S.; Legarth, R.; Fischer Button, M.S.; Gubbels, S.M. Increased risk of hospitalisation associated with infection with SARS-CoV-2 lineage B. 1.1. 7 in Denmark. Lancet Infect. Dis. 2021, 21, 1507–1517. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoteit, R.; Yassine, H.M. Biological Properties of SARS-CoV-2 Variants: Epidemiological Impact and Clinical Consequences. Vaccines 2022, 10, 919. https://doi.org/10.3390/vaccines10060919
Hoteit R, Yassine HM. Biological Properties of SARS-CoV-2 Variants: Epidemiological Impact and Clinical Consequences. Vaccines. 2022; 10(6):919. https://doi.org/10.3390/vaccines10060919
Chicago/Turabian StyleHoteit, Reem, and Hadi M. Yassine. 2022. "Biological Properties of SARS-CoV-2 Variants: Epidemiological Impact and Clinical Consequences" Vaccines 10, no. 6: 919. https://doi.org/10.3390/vaccines10060919