
Co-Administration of Adjuvanted Recombinant Ov-103 and Ov-RAL-2 Vaccines Confer Protection against Natural Challenge in A Bovine Onchocerca ochengi Infection Model of Human Onchocerciasis

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Recombinant Antigen Production
2.2. Calf Husbandry and Field Site
2.3. Immunisation Strategy
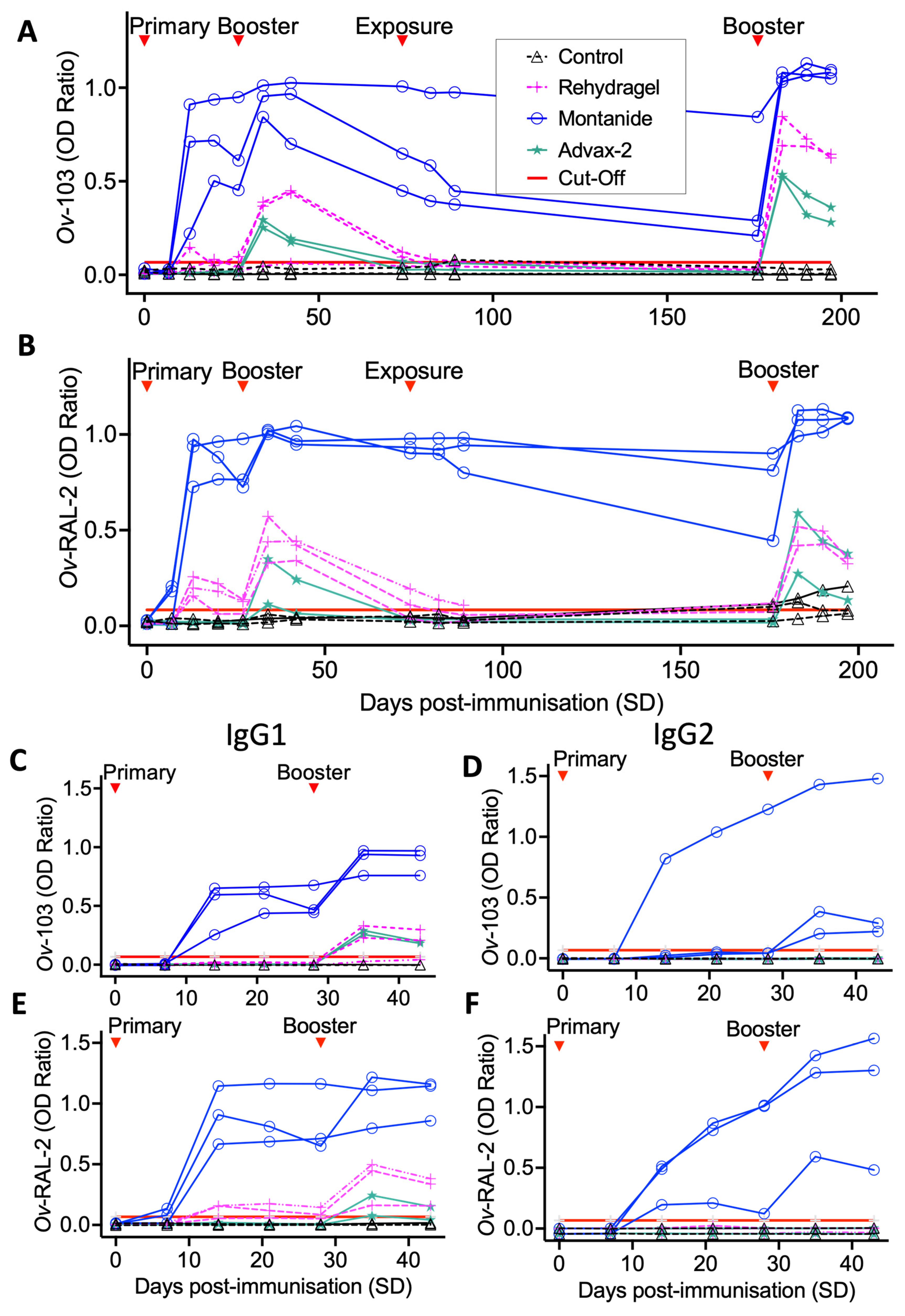
2.3.1. Immunogenicity Trial
2.3.2. Vaccine Efficacy Trial
2.4. Natural Exposure to O. ochengi
2.5. Routine Sampling and Clinical Observations
2.6. End-Point Sampling
2.7. Entomological Analyses and Transmission Potential
2.8. Measuring Anti-Ov-103 and Ov-Ral-2 IgG Responses
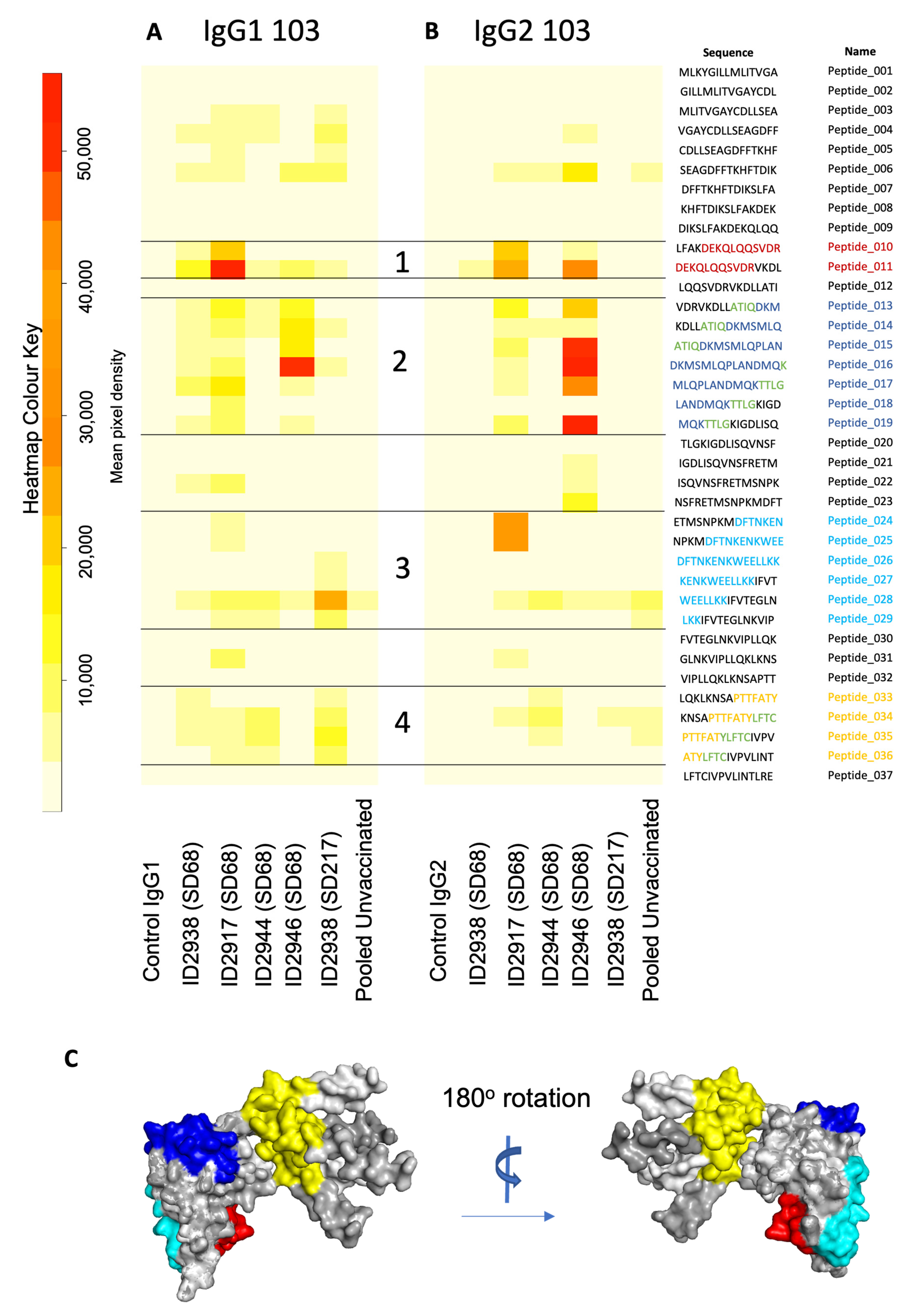
2.9. IgG Purification and Epitope Mapping
2.10. Protein Structure Prediction
2.11. PBMC Proliferation Assays
2.12. Statistical Data Analysis
2.12.1. Immunisation Period
2.12.2. Exposure Period
2.12.3. Assessing Immunological Correlates of Vaccine-Associated Protection
2.12.4. Final Parasitological Burden
3. Results
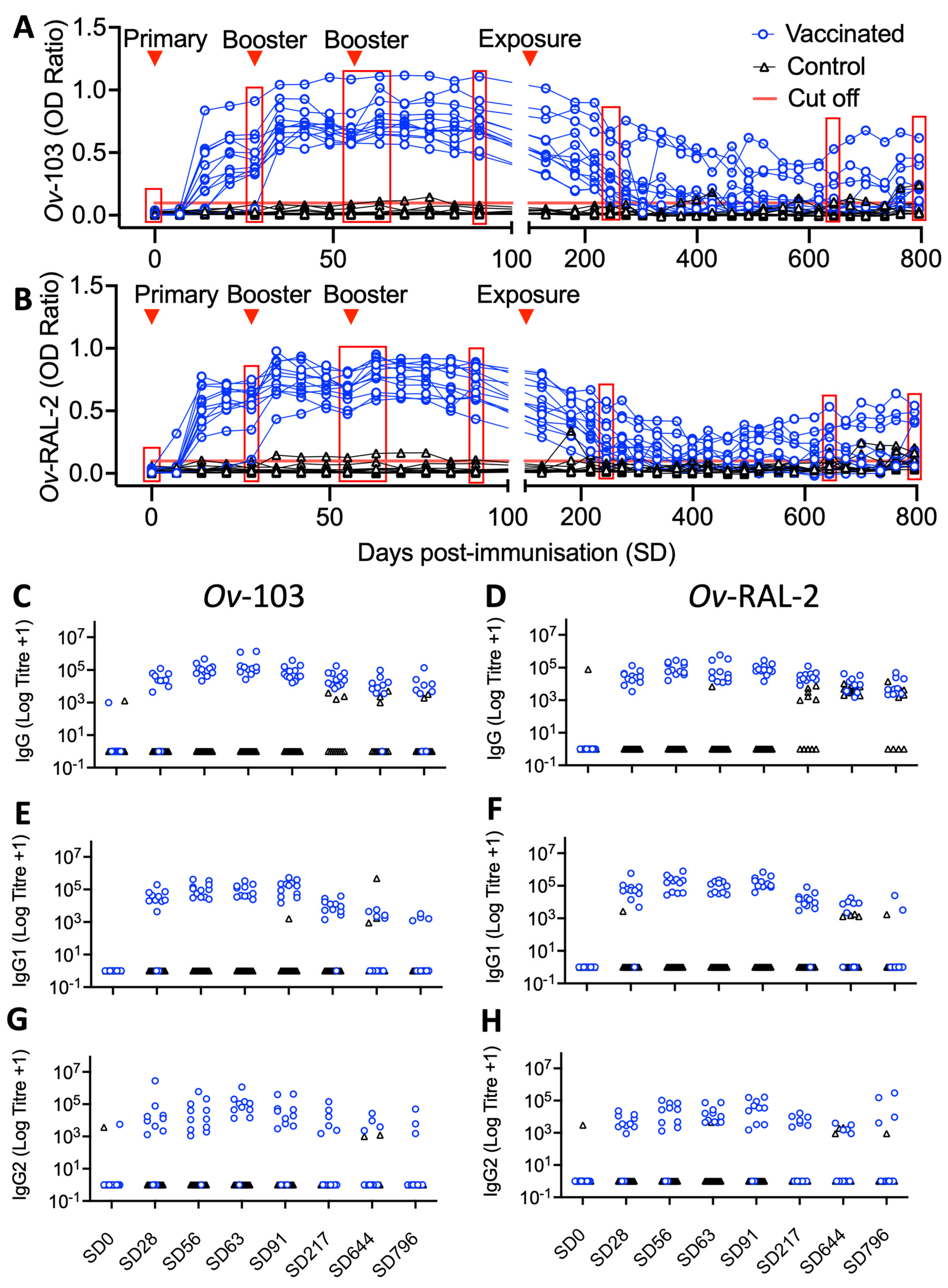
3.1. Ov-103 and Ov-RAL-2 Immunisation in Cattle with MontanideTM ISA 206VG Induces Pronounced, Rapid and Sustained Seroconversion
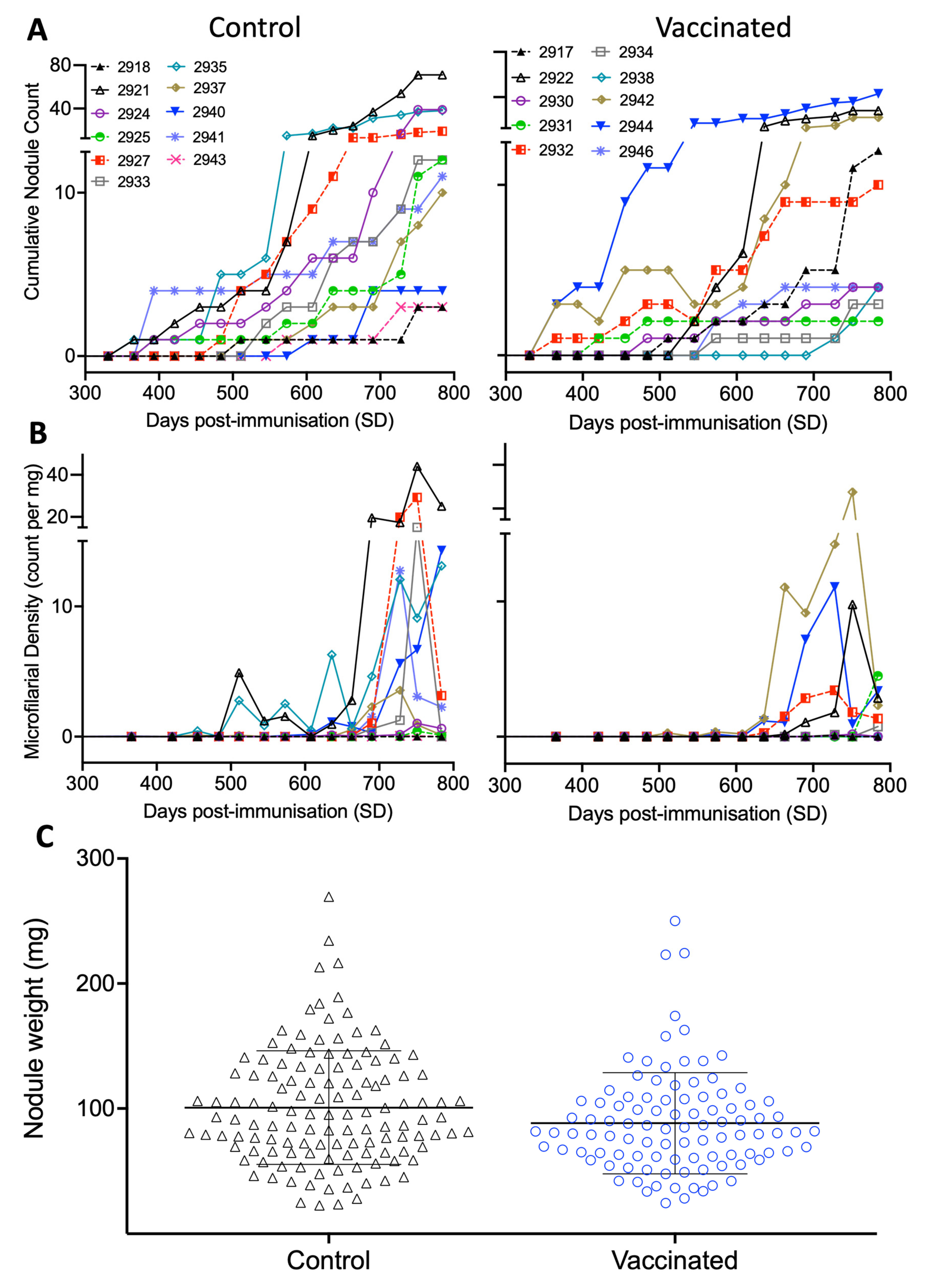
3.2. Ov-103 and Ov-RAL-2 Immunisation in Cattle Is Associated with Reduced O. ochengi Female and Microfilarial Worm Burdens following Natural Exposure
3.3. Vaccine-Induced Ov-103 and Ov-RAL-2-Specific Serum IgG Responses Are Associated with Reduced Adult O. ochengi Burdens following Natural Exposure
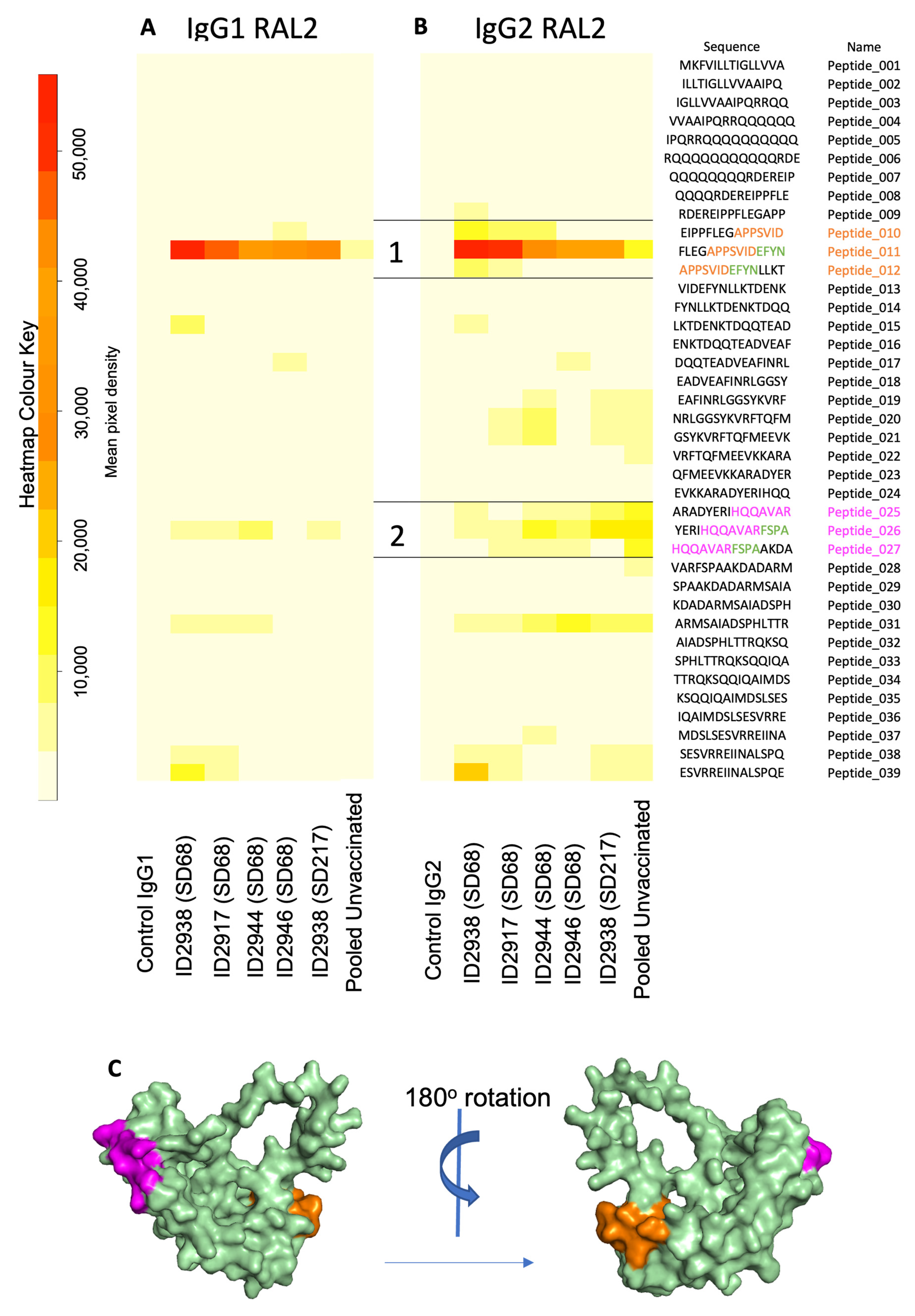
3.4. Epitope Mapping Identifies Immunogenic Surface Peptide Sequences of Ov-103 and Ov-RAL-2
3.5. Ov-103 and Ov-RAL-2 Immunisation in Cattle Is Associated with Altered Cellular Immune Responses
3.6. Calf Health and Diagnostics
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- WHO Onchocerciasis (River Blindness). Available online: https://www.who.int/health-topics/onchocerciasis (accessed on 23 March 2021).
- James, S.L.; Abate, D.; Abate, K.H.; Abay, S.M.; Abbafati, C.; Abbasi, N.; Abbastabar, H.; Abd-Allah, F.; Abdela, J.; Abdelalim, A.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 354 Diseases and Injuries for 195 Countries and Territories, 1990–2017: A Systematic Analysis for the Global Burden of Disease Study 2017. Lancet 2018, 392, 1789–1858. [Google Scholar] [CrossRef] [Green Version]
- Vos, T.; Allen, C.; Arora, M.; Barber, R.M.; Bhutta, Z.A.; Brown, A.; Carter, A.; Casey, D.C.; Charlson, F.J.; Chen, A.Z.; et al. Global, Regional, and National Incidence, Prevalence, and Years Lived with Disability for 310 Diseases and Injuries, 1990–2015: A Systematic Analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1545–1602. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization; African Programme for Onchocerciasis Control (APOC). Report of the Consultative Meetings on Strategic Options and Alternative Treatment Strategies for Accelerating Onchocerciasis Elimination in Africa; African Programme for Onchocerciasis Control. 2015. Available online: https://apps.who.int/iris/handle/10665/343028 (accessed on 24 May 2022).
- Gardon, J.; Gardon-Wendel, N.; Demanga-Ngangue; Kamgno, J.; Chippaux, J.-P.; Boussinesq, M. Serious Reactions after Mass Treatment of Onchocerciasis with Ivermectin in an Area Endemic for Loa Loa Infection. Lancet 1997, 350, 18–22. [Google Scholar] [CrossRef]
- Pion, S.D.S.; Nana-Djeunga, H.C.; Kamgno, J.; Tendongfor, N.; Wanji, S.; Njiokou, F.; Prichard, R.K.; Boussinesq, M. Dynamics of Onchocerca volvulus Microfilarial Densities after Ivermectin Treatment in an Ivermectin-Naïve and a Multiply Treated Population from Cameroon. PLoS Negl. Trop. Dis. 2013, 7, e2084. [Google Scholar] [CrossRef] [Green Version]
- Turner, H.C.; Walker, M.; Churcher, T.S.; Osei-Atweneboana, M.Y.; Biritwum, N.-K.; Hopkins, A.; Prichard, R.K.; Basáñez, M.-G. Reaching the London Declaration on Neglected Tropical Diseases Goals for Onchocerciasis: An Economic Evaluation of Increasing the Frequency of Ivermectin Treatment in Africa. Clin. Infect. Dis. 2014, 59, 923–932. [Google Scholar] [CrossRef]
- Keenan, J.D.; Hotez, P.J.; Amza, A.; Stoller, N.E.; Gaynor, B.D.; Porco, T.C.; Lietman, T.M. Elimination and Eradication of Neglected Tropical Diseases with Mass Drug Administrations: A Survey of Experts. PLoS Negl. Trop. Dis. 2013, 7, e2562. [Google Scholar] [CrossRef]
- Awadzi, K.; Opoku, N.O.; Attah, S.K.; Lazdins-Helds, J.; Kuesel, A.C. A Randomized, Single-Ascending-Dose, Ivermectin-Controlled, Double-Blind Study of Moxidectin in Onchocerca volvulus Infection. PLoS Negl. Trop. Dis. 2014, 8, e2953. [Google Scholar] [CrossRef]
- Milton, P.; Hamley, J.I.D.; Walker, M.; Basáñez, M.-G. Moxidectin: An Oral Treatment for Human Onchocerciasis. Expert Rev. Anti-Infect. Ther. 2020, 18, 1067–1081. [Google Scholar] [CrossRef]
- Opoku, N.O.; Bakajika, D.K.; Kanza, E.M.; Howard, H.; Mambandu, G.L.; Nyathirombo, A.; Nigo, M.M.; Kasonia, K.; Masembe, S.L.; Mumbere, M.; et al. Single Dose Moxidectin versus Ivermectin for Onchocerca volvulus Infection in Ghana, Liberia, and the Democratic Republic of the Congo: A Randomised, Controlled, Double-Blind Phase 3 Trial. Lancet 2018, 392, 1207–1216. [Google Scholar] [CrossRef] [Green Version]
- Ngwewondo, A.; Scandale, I.; Specht, S. Onchocerciasis Drug Development: From Preclinical Models to Humans. Parasitol. Res. 2021, 120, 3939–3964. [Google Scholar] [CrossRef]
- Johnston, K.L.; Hong, W.D.; Turner, J.D.; O’Neill, P.M.; Ward, S.A.; Taylor, M.J. Anti-Wolbachia Drugs for Filariasis. Trends Parasitol. 2021, 37, 1068–1081. [Google Scholar] [CrossRef]
- Lustigman, S.; Makepeace, B.L.; Klei, T.R.; Babayan, S.; Hotez, P.; Abraham, D. Bottazzi Onchocerca volvulus: The Road from Basic Biology to a Vaccine. Trends Parasitol. 2018, 34, 64–79. [Google Scholar] [CrossRef]
- Turner, H.C.; Walker, M.; Lustigman, S.; Taylor, D.W.; Basáñez, M.-G. Human Onchocerciasis: Modelling the Potential Long-Term Consequences of a Vaccination Programme. PLoS Negl. Trop. Dis. 2015, 9, e0003938. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, S.; Wei, J.; Liu, Z.; Abraham, D.; Bell, A.; Bottazzi, M.E.; Hotez, P.J.; Zhan, B.; Lustigman, S.; Klei, T.R. Vaccination of Gerbils with Bm-103 and Bm-RAL-2 Concurrently or as a Fusion Protein Confers Consistent and Improved Protection against Brugia malayi Infection. PLoS Negl. Trop. Dis. 2016, 10, 1–26. [Google Scholar] [CrossRef]
- George, J.P.; Hess, J.A.; Jain, S.; Patton, J.B.; Zhan, T.; Tricoche, N.; Zhan, B.; Bottazzi, M.E.; Hotez, P.J.; Abraham, D.; et al. Antibody Responses against the Vaccine Antigens Ov-103 and Ov-RAL-2 Are Associated with Protective Immunity to Onchocerca volvulus Infection in Both Mice and Humans. PLoS Negl. Trop. Dis. 2019, 13, e0007730. [Google Scholar] [CrossRef]
- Hess, J.A.; Zhan, B.; Bonne-Année, S.; Deckman, J.M.; Bottazzi, M.E.; Hotez, P.J.; Klei, T.R.; Lustigman, S.; Abraham, D. Vaccines to Combat River Blindness: Expression, Selection and Formulation of Vaccines against Infection with Onchocerca volvulus in a Mouse Model. Int. J. Parasitol. 2014, 44, 637–646. [Google Scholar] [CrossRef] [Green Version]
- Hess, J.A.; Zhan, B.; Torigian, A.R.; Patton, J.B.; Petrovsky, N.; Zhan, T.; Bottazzi, M.E.; Hotez, P.J.; Klei, T.R.; Lustigman, S.; et al. The Immunomodulatory Role of Adjuvants in Vaccines Formulated with the Recombinant Antigens Ov-103 and Ov-RAL-2 against Onchocerca volvulus in Mice. PLoS Negl. Trop. Dis. 2016, 10, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Ryan, N.M.; Hess, J.A.; de Villena, F.P.-M.; Leiby, B.E.; Shimada, A.; Yu, L.; Yarmahmoodi, A.; Petrovsky, N.; Zhan, B.; Bottazzi, M.E.; et al. Onchocerca volvulus Bivalent Subunit Vaccine Induces Protective Immunity in Genetically Diverse Collaborative Cross Recombinant Inbred Intercross Mice. NPJ Vaccines 2021, 6, 1–12. [Google Scholar] [CrossRef]
- Makepeace, B.L.; Jensen, S.A.; Laney, S.J.; Nfon, C.K.; Njongmeta, L.M.; Tanya, V.N.; Williams, S.A.; Bianco, A.E.; Trees, A.J. Immunisation with a Multivalent, Subunit Vaccine Reduces Patent Infection in a Natural Bovine Model of Onchocerciasis during Intense Field Exposure. PLoS Negl. Trop. Dis. 2009, 3, e544. [Google Scholar] [CrossRef] [Green Version]
- Estes, M.D.; Closser, N.M.; Allen, G.K. IFN-γ Stimulates IgG2 Production from Bovine B Cells Costimulated with Anti-μ and Mitogen. Cell. Microbiol. 1994, 154, 287–295. [Google Scholar] [CrossRef]
- Townsend, J.; Duffus, W.P.H. Antibody-Dependent Cellular Cytotoxicity of Trypanosoma theileri Mediated by Purified Bovine Isotypes and Subisotypes. Parasite Immunol. 1985, 7, 179–189. [Google Scholar] [CrossRef]
- Makepeace, B.L.; Tanya, V.N. 25 Years of the Onchocerca ochengi Model. Trends Parasitol. 2016, 32, 966–978. [Google Scholar] [CrossRef] [Green Version]
- Bah, G.S.; Ward, E.L.; Srivastava, A.; Trees, A.J.; Tanya, V.N.; Makepeace, B.L. Efficacy of Three-Week Oxytetracycline or Rifampin Monotherapy Compared with a Combination Regimen against the Filarial Nematode Onchocerca ochengi. Antimicrob. Agents Chemother. 2014, 58. [Google Scholar] [CrossRef] [Green Version]
- Eisenbarth, A.; Achukwi, M.D.; Renz, A. Ongoing Transmission of Onchocerca volvulus after 25 Years of Annual Ivermectin Mass Treatments in the Vina Du Nord River Valley, in North Cameroon. PLOS Negl. Trop. Dis. 2016, 10, e0004392. [Google Scholar] [CrossRef] [Green Version]
- Tchakouté, V.L.; Graham, S.P.; Jensen, S.A.; Makepeace, B.L.; Nfon, C.K.; Njongmeta, L.M.; Lustigman, S.; Enyong, P.A.; Tanya, V.N.; Bianco, A.E.; et al. In a Bovine Model of Onchocerciasis, Protective Immunity Exists Naturally, Is Absent in Drug-Cured Hosts, and Is Induced by Vaccination. Proc. Natl. Acad. Sci. USA 2006, 103, 5971–5976. [Google Scholar] [CrossRef] [Green Version]
- Renz, A.; Trees, A.J.; Achu-Kwi, D.; Edwards, G.; Wahl, G. Evaluation of Suramin, Ivermectin and CGP 20376 in a New Macrofilaricidal Drug Screen, Onchocerca ochengi in African Cattle. Trop. Med. Parasitol. 1995, 46, 31–37. [Google Scholar]
- Detinova, T.S. Age-Grouping Methods in Diptera of Medical Importance with Special Reference to Some Vectors of Malaria. Monogr. Ser. World Health Organ. 1962, 47, 13–191. [Google Scholar] [CrossRef]
- Wahl, G.; Ekale, D.; Schmitz, A. Onchocerca ochengi: Assessment of the Simulium Vectors in North Cameroon. Parasitology 1998, 116, 327–336. [Google Scholar] [CrossRef]
- Duke, B.O.L. Infective Filaria Larvae, Other than Onchocerca volvulus, in Simulium damnosum. Ann. Trop. Med. Parasitol. 1967, 61, 200–205. [Google Scholar] [CrossRef]
- Wahl, G.; Ekale, D.; Enyong, P.; Renz, A. The Development of Onchocerca dukei and O. ochengi Microfilariae to Infective-Stage Larvae in Simulium damnosum s.l. and in Members of the S. medusaeforme Group, Following Intra-Thoracic Injection. Ann. Trop. Med. Parasitol. 1991, 85, 329–337. [Google Scholar] [CrossRef]
- Wahl, G.; Schibel, J.M. Onchocerca ochengi: Morphological Identification of the L3 in Wild Simulium damnosum s.l., Verified by DNA Probes. Parasitology 1998, 116, 337–348. [Google Scholar] [CrossRef] [PubMed]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria. Available online: https://www.r-project.org/ (accessed on 24 May 2022).
- Stephenson, K.E.; Neubauer, G.H.; Reimer, U.; Pawlowski, N.; Knaute, T.; Zerweck, J.; Korber, B.T.; Barouch, D.H. Quantification of the Epitope Diversity of HIV-1-Specific Binding Antibodies by Peptide Microarrays for Global HIV-1 Vaccine Development. J. Immunol. Methods 2015, 416, 105–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jumper, J.; Evans, R.; Pritzel, A.; Green, T.; Figurnov, M.; Ronneberger, O.; Tunyasuvunakool, K.; Bates, R.; Žídek, A.; Potapenko, A.; et al. Highly Accurate Protein Structure Prediction with AlphaFold. Nature 2021, 596, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.; DiMaio, F.; Anishchenko, I.; Dauparas, J.; Ovchinnikov, S.; Lee, G.R.; Wang, J.; Cong, Q.; Kinch, L.N.; Schaeffer, R.D.; et al. Accurate Prediction of Protein Structures and Interactions Using a 3-Track Neural Network. Science 2021, 373, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Graham-Brown, J.; Hartley, C.; Clough, H.; Kadioglu, A.; Baylis, M.; Williams, D.J.L. Dairy Heifers Naturally Exposed to Fasciola hepatica Develop a Type 2 Immune Response and Concomitant Suppression of Leukocyte Proliferation. Infect. Immun. 2018, 88, e00607-17. [Google Scholar] [CrossRef] [Green Version]
- Pinheiro, J.; Bates, D.; Debroy, S.; Sarkar, D. R Core Team Nlme: Linear and Nonlinear Mixed Effects Models. R Package Verion 3.1-155. 2022. Available online: https://cran.r-project.org/web/packages/nlme/nlme.pdf (accessed on 24 May 2022).
- Achukwi, M.D.; Harnett, W.; Bradley, J.; Renz, A. Onchocerca ochengi Acquisition in Zebu Gudali Cattle Exposed to Natural Transmission: Parasite Population Dynamics and IgG Antibody Subclass Responses to Ov10/Ov11 Recombinant Antigens. Vet. Parasitol. 2004, 122, 35–49. [Google Scholar] [CrossRef]
- Duerr, H.P.; Dietz, K.; Schulz-Key, H.; Buttner, D.W.; Eichner, M. The Relationships between the Burden of Adult Parasites, Host Age and the Microfilarial Density in Human Onchocerciasis. Int. J. Parasitol. 2004, 34, 463–473. [Google Scholar] [CrossRef]
- Howard, C.J. Comparison of Bovine IgG1, IgG2 and IgM for Ability to Promote Killing of Mycoplasma bovis by Bovine Alveaolar Macrophages and Neutrophils. Vet. Immunol. Immunopathol. 1984, 6, 321–326. [Google Scholar] [CrossRef]
- MacDonald, A.J.; Turaga, P.S.D.; Harmon-Brown, C.; Tierney, T.J.; Bennett, K.E.; McCarthy, M.C.; Simonek, S.C.; Enyong, P.A.; Moukatte, D.W.; Lustigman, S. Differential Cytokine and Antibody Responses to Adult and Larval Stages of Onchocerca volvulus Consistent with the Development of Concomitant Immunity. Infect. Immun. 2002, 70, 2796–2804. [Google Scholar] [CrossRef] [Green Version]
- Turaga, P.S.D.; Tierney, T.J.; Bennett, K.E.; McCarthy, M.C.; Simonek, S.C.; Enyong, P.A.; Moukatte, D.W.; Lustigman, S. Immunity to Onchocerciasis: Cells from Putatively Immune Individuals Produce Enhanced Levels of Interleukin-5, Gamma Interferon, and Granulocyte-Macrophage Colony-Stimulating Factor in Response to Onchocerca volvulus Larval and Male Worm Antigens. Infect. Immun. 2000, 68, 1905–1911. [Google Scholar] [CrossRef] [Green Version]
- Johnson, E.H.; Irvine, M.; Kass, P.H.; Browne, J.; Abdullai, M.; Prince, A.M.; Lustigman, S. Onchocerca volvulus: In Vitro Cytotoxic Effects of Human Neutrophils and Serum on Third-Stage Larvae. Trop. Med. Parasitol. 1994, 45, 331–335. [Google Scholar] [PubMed]
- Johnson, E.H.; Lustigman, S.; Kass, P.H.; Irvine, M.; Browne, J.; Prince, A.M. Onchocerca volvulus: A Comparative Study of in Vitro Neutrophil Killing of Microfilariae and Humoral Responses in Infected and Endemic Normals. Exp. Parasitol. 1995, 81, 9–19. [Google Scholar] [CrossRef]
- Lustigman, S.; Brotman, B.; Johnson, E.H.; Smith, A.B.; Huima, T.; Prince, A.M. Identification and Characterization of an Onchocerca volvulus CDNA Clone Encoding a Microfilarial Surface-Associated Antigen. Mol. Biochem. Parasitol. 1992, 50, 79–93. [Google Scholar] [CrossRef]
- Bradley, J.E.; Elson, L.; Tree, T.I.M.; Stewart, G.; Guderian, R.; Calvopina, M.; Paredes, W.; Araujo, E.; Nutman, T.B. Resistance to Onchocerca volvulus: Differential Cellular and Humoral Responses to a Recombinant Antigen, OvMBP20/11. J. Infect. Dis. 1995, 172, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Lustigman, S.; MacDonald, A.J.; Abraham, D. CD4+-Depedent Immunity to Onchocerca volvulus Third-Stage Larvae in Humans and the Mouse Vaccination Model: Common Ground and Disinctions. Int. J. Parasitol. 2003, 33, 1161–1171. [Google Scholar] [CrossRef]
- Lustigman, S.; James, E.R.; Tawe, W.; Abraham, D. Towards a Recombinant Antigen Vaccine against Onchocerca volvulus. Trends Parasitol. 2002, 18, 135–141. [Google Scholar]
- Gupta, J.; Misra, S.; Misra-Bhattacharya, S. Immunization with Brugia malayi Myosin as Heterologous DNA Prime Protein Boost Induces Protective Immunity against B. malayi Infection in Mastomys coucha. PLoS ONE 2016, 11, e0164991. [Google Scholar] [CrossRef]
- Khatri, V.; Chauhan, N.; Kalyanasundaram, R. Fecundity of Adult Female Worms Were Affected When Brugia malayi Infected Mongolian Gerbils Were Immunized with a Multivalent Vaccine (RBmHAXT) against Human Lymphatic Filarial Parasite. Acta Trop. 2020, 208, 105487. [Google Scholar] [CrossRef]
- Hernández-Goenaga, J.; López-Abán, J.; Protasio, A.V.; Vicente Santiago, B.; del Olmo, E.; Vanegas, M.; Fernández-Soto, P.; Patarroyo, M.A.; Muro, A. Peptides Derived of Kunitz-Type Serine Protease Inhibitor as Potential Vaccine Against Experimental Schistosomiasis. Front. Immunol. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Turner, J.; Howell, A.; McCann, C.; Caminade, C.; Bowers, R.G.; Williams, D.; Baylis, M. A Model to Assess the Efficacy of Vaccines for Control of Liver Fluke Infection. Sci. Rep. 2016, 6, 23345. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, G.; Zhang, W.; Torben, W.; Damian, R.T.; Wolf, R.F.; White, G.L.; Chavez-Suarez, M.; Kennedy, R.C.; Siddiqui, A.A. Protective and Antifecundity Effects of Sm-P80-Based DNA Vaccine Formulation against Schistosoma mansoni in a Nonhuman Primate Model. Vaccine 2009, 27, 2830–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagatie, O.; Verheyen, A.; Van Dorst, B.; Batsa Debrah, L.; Debrah, A.; Stuyver, L.J. Linear Epitopes in Onchocerca volvulus Vaccine Candidate Proteins and Excretory-Secretory Proteins. Parasite Immunol. 2018, 40, e12587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shey, R.A.; Ghogomu, S.M.; Shintouo, C.M.; Nkemngo, F.N.; Nebangwa, D.N.; Esoh, K.; Yaah, N.E.; Manka’aFri, M.; Nguve, J.E.; Ngwese, R.A.; et al. Computational Design and Preliminary Serological Analysis of a Novel Multi-Epitope Vaccine Candidate Against Onchocerciasis and Related Filarial Diseases. Pathogens 2021, 10, 99. [Google Scholar] [CrossRef] [PubMed]
- Pastor, A.F.; Silva, M.R.; dos Santos, W.J.T.; Rego, T.; Brandão, E.; de-Melo-Neto, O.P.; Rocha, A. Recombinant Antigens Used as Diagnostic Tools for Lymphatic Filariasis. Parasites Vectors 2021, 14, 474. [Google Scholar] [CrossRef] [PubMed]
- Hansen, R.D.E.; Trees, A.J.; Bah, G.S.; Hetzel, U.; Martin, C.; Bain, O.; Tanya, V.N.; Makepeace, B.L. A Worm’s Best Friend: Recruitment of Neutrophils by Wolbachia Confounds Eosinophil Degranulation against the Filarial Nematode Onchocerca ochengi. Proc. R. Soc. B Biol. Sci. 2011, 278, 2293–2302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bah, G.S.; Schneckener, S.; Hahnel, S.R.; Bayang, N.H.; Fieseler, H.; Schmuck, G.M.; Krebber, R.; Sarr, A.; Terjung, C.; Ngangyung, H.F.; et al. Emodepside Targets SLO-1 Channels of Onchocerca ochengi and Induces Broad Anthelmintic Effects in a Bovine Model of Onchocerciasis. PLOS Pathog. 2021, 17, e1009601. [Google Scholar] [CrossRef]
- Griebel, P.J.; Brownlie, R.; Manuja, A.; Nichani, A.; Mookherjee, N.; Popowych, Y.; Mutwiri, G.; Hecker, R.; Babiuk, L.A. Bovine Toll-like Receptor 9: A Comparative Analysis of Molecular Structure, Function and Expression. Vet. Immunol. Immunopathol. 2005, 108, 11–16. [Google Scholar] [CrossRef]
- Golden, O.; Flynn, R.J.; Read, C.; Sekiya, M.; Donnelly, S.M.; Stack, C.; Dalton, J.P.; Mulcahy, G. Protection of Cattle against a Natural Infection of Fasciola hepatica by Vaccination with Recombinant Cathepsin L1 (RFhCL1). Vaccine 2010, 28, 5551–5557. [Google Scholar] [CrossRef]
- Flynn, R.J.; Mulcahy, G. The Roles of IL-10 and TGF-β in Controlling IL-4 and IFN-γ Production during Experimental Fasciola hepatica Infection. Int. J. Parasitol. 2008, 38, 1673–1680. [Google Scholar] [CrossRef]
- Mulcahy, G.; O’Connor, F.; McGonigle, S.; Down, A.; Clery, D.G.; Andrews, S.J.; Dalton, J.P. Correlation of Specific Antibody Titre and Avidity with Protection in Cattle Immunized against Fasciola hepatica. Vaccine 1998, 16, 932–939. [Google Scholar] [CrossRef]
- Kloor, M.; Reuschenbach, M.; Pauligk, C.; Karbach, J.; Rafiyan, M.-R.; Al-Batran, S.-E.; Tariverdian, M.; Jäger, E.; von Knebel Doeberitz, M. A Frameshift Peptide Neoantigen-Based Vaccine for Mismatch Repair-Deficient Cancers: A Phase I/IIa Clinical Trial. Clin. Cancer Res. 2020, 26, 4503–4510. [Google Scholar] [CrossRef] [PubMed]
- Heitmann, J.S.; Bilich, T.; Tandler, C.; Nelde, A.; Maringer, Y.; Marconato, M.; Reusch, J.; Jäger, S.; Denk, M.; Richter, M.; et al. A COVID-19 Peptide Vaccine for the Induction of SARS-CoV-2 T Cell Immunity. Nature 2022, 601, 617–622. [Google Scholar] [CrossRef] [PubMed]
- Aucouturier, J.; Dupuis, L.; Deville, S.; Ascarateil, S.; Ganne, V. Montanide ISA 720 and 51: A New Generation of Water in Oil Emulsions as Adjuvants for Human Vaccines. Expert Rev. Vaccines 2014, 1, 111–118. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ID | Ov-RAL-2 | Ov-103 | ||||
|---|---|---|---|---|---|---|---|
| IgG | IgG1 | IgG2 | IgG | IgG1 | IgG2 | ||
| Control | 2901 | 5800 | 0 | 0 | 2300 | 0 | 0 |
| 2904 | 0 | 0 | 0 | 0 | 0 | 0 | |
| 2911 | 0 | 0 | 0 | 900 | 0 | 0 | |
| Rehydragel | 2906 | 47,700 | 15,800 | 0 | 19,500 | 34,200 | 0 |
| 2908 | 18,300 | 15,100 | 0 | 9500 | 47,700 | 0 | |
| Advax-2 | 2910 | 12,800 | 11,900 | 0 | 5000 | 38,800 | 0 |
| 2912 | 24,000 | 28,300 | 0 | 8500 | 44,300 | 0 | |
| Montanide | 2907 | 944,300 | 669,900 | 1,086,800 | 373,900 | 233,300 | 27,300 |
| 2909 | 1,039,700 | 87,700 | 868,200 | 283,200 | 82,200 | 721,200 | |
| 2913 | 404,000 | 524,000 | 119,900 | 216,500 | 183,200 | 28,200 | |
| Control Group (n = 11) | Vaccine Group (n = 10) | |
|---|---|---|
| Nodule burden | 20.2 (±21.0); 3–79 | 13.2 (±13.6); 2–43 |
| Nodule diameter (mm) | 4.8 (0.8); 3.1–7.8 (nodules assessed = 147) | 4.7 (0.8); 3.0–7.0 (nodules assessed = 107) |
| Nodule age * (days) | 545 (±89); 293–694 (nodules assessed = 147) | 536 (±101); 266–688 (nodules assessed = 107) |
| Nodule weight (mg) | 100.6 (45.6); 22.7–269.4 (nodules assessed = 128) | 88.6 (40.3); 24.6–250.1 (nodules assessed = 100) |
| Male worms recovered per nodule | 0.5 (0.7); 0–3 (nodules assessed = 50) | 0.7 (1.2); 0–5 (nodules assessed = 38) |
| O. ochengi microfilarial density (per mg skin) | 4.7 (±7.9); 0–25.1 | 1.4 (±1.6); 0–4.5 |
| Response Variable (Y) | Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
|---|---|---|---|---|
| Female worm nodules | Time | 0.024 | 0.0017 | <0.0001 * |
| Sex (m) | 2.65 | 1.72 | 0.14 | |
| Vaccination:Time (interaction) | −0.0062 | 0.0022 | 0.0043 * | |
| Microfilarial burden | Time | 0.0080 | 0.0012 | <0.0001 * |
| Sex (m) | 0.76 | 0.78 | 0.34 | |
| Vaccination:Time (interaction) | −0.0031 | 0.0012 | 0.0085 * |
| Anti-Ov-103 IgG Responses | ||||
|---|---|---|---|---|
| Response Variable (Y) | Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
| Total IgG | Time | −0.00013 | 0.00038 | 0.74 |
| Sex (m) | 0.025 | 0.056 | 0.66 | |
| Vaccination:Time (interaction) | 0.0091 | 0.00051 | <0.0001 * | |
| IgG1 titre (logged) | Time | −0.013 | 0.0091 | 0.15 |
| Sex (m) | −0.15 | 0.43 | 0.74 | |
| Vaccination:Time (interaction) | 0.21 | 0.0090 | <0.0001 * | |
| IgG2 titre (logged) | Time | −0.016 | 0.011 | 0.17 |
| Sex (m) | −0.29 | 0.97 | 0.77 | |
| Vaccination:Time (interaction) | 0.16 | 0.014 | <0.0001 * | |
| Anti-Ov-RAL-2 IgG Responses | ||||
| Response Variable (Y) | Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
| Total IgG | Time | −0.00020 | 0.00041 | 0.62 |
| Sex (m) | 0.039 | 0.059 | 0.52 | |
| Vaccination:Time (interaction) | 0.0083 | 0.00055 | <0.0001 * | |
| IgG1 titre (logged) | Time | −0.018 | 0.010 | 0.094 |
| Sex (m) | 0.19 | 0.49 | 0.70 | |
| Vaccination:Time (interaction) | 0.21 | 0.010 | <0.0001 * | |
| IgG2 titre (logged) | Time | −0.0075 | 0.011 | 0.50 |
| Sex (m) | −0.13 | 0.71 | 0.85 | |
| Vaccination:Time (interaction) | 0.15 | 0.013 | <0.0001 * | |
| Anti-Ov-103 IgG Responses | ||||
|---|---|---|---|---|
| Response Variable (Y) | Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
| Total IgG | Time | 0.000031 | 0.000029 | 0.30 |
| Vaccination | 0.51 | 0.056 | <0.0001 * | |
| Sex (m) | −0.013 | 0.058 | 0.82 | |
| Vaccination:Time (interaction) | −0.00062 | 0.000042 | <0.0001 * | |
| IgG1 titre (logged) | Time | 0.0040 | 0.0020 | 0.053 |
| Vaccination | 12.21 | 1.17 | <0.0001 * | |
| Sex (m) | 0.69 | 0.87 | 0.44 | |
| Vaccination:Time (interaction) | −0.017 | 0.0029 | <0.0001 * | |
| IgG2 titre (logged) | Time | 0.0025 | 0.0019 | 0.19 |
| Vaccination | 8.34 | 1.46 | <0.0001 * | |
| Sex (m) | −0.56 | 1.34 | 0.68 | |
| Vaccination:Time (interaction) | −0.010 | 0.0027 | 0.00054 * | |
| Anti-Ov-RAL-2 IgG Responses | ||||
| Response Variable (Y) | Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
| Total IgG | Time | 0.000050 | 0.000031 | 0.10 |
| Vaccination | 0.43 | 0.043 | <0.0001 * | |
| Sex (m) | 0.047 | 0.042 | 0.28 | |
| Vaccination:Time (interaction) | −0.00055 | 0.000044 | <0.0001 * | |
| IgG1 titre (logged) | Time | 0.0052 | 0.0017 | 0.0042 * |
| Vaccination | 12.68 | 1.04 | <0.0001 * | |
| Sex (m) | 1.51 | 0.81 | 0.076 | |
| Vaccination:Time (interaction) | −0.016 | 0.0025 | <0.0001 * | |
| IgG2 titre (logged) | Time | 0.0039 | 0.0016 | 0.020 * |
| Vaccination | 8.62 | 1.37 | <0.0001 * | |
| Sex (m) | −0.97 | 1.31 | 0.46 | |
| Vaccination:Time (interaction) | −0.011 | 0.0023 | <0.0001 * | |
| Anti-Ov-103 IgG Responses | |||
|---|---|---|---|
| Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
| Total IgG | −8.01 | 1.83 | <0.0001 * |
| Sex (m) | 2.62 | 2.74 | 0.36 |
| Total IgG:Time (interaction) | 0.034 | 0.0059 | <0.0001 * |
| IgG1 titre (logged) | −0.52 | 0.17 | 0.0059 * |
| Sex (m) | 1.51 | 1.58 | 0.36 |
| IgG1 titre (logged):Time (interaction) | 0.0012 | 0.00045 | 0.015 * |
| IgG2 titre (logged) | −0.38 | 0.18 | 0.046 * |
| Sex (m) | 1.42 | 1.75 | 0.43 |
| IgG2 titre (logged):Time (interaction) | 0.00095 | 0.00051 | 0.073 |
| Anti-Ov-RAL-2 IgG Responses | |||
| Fixed-Effect Explanatory Variable (x) | Co-Efficient Value (β) | Standard Error | p-Value |
| Total IgG | −7.40 | 1.83 | <0.0001 * |
| Sex (m) | 0.42 | 2.94 | 0.89 |
| Total IgG:Time (interaction) | 0.049 | 0.0050 | <0.0001 * |
| IgG1 titre (logged) | −0.55 | 0.18 | 0.0057 * |
| Sex (m) | 1.66 | 1.61 | 0.33 |
| IgG1 titre (logged):Time (interaction) | 0.00094 | 0.00038 | 0.023 * |
| IgG2 titre (logged) | −0.40 | 0.19 | 0.043 * |
| Sex (m) | 1.08 | 1.75 | 0.55 |
| IgG2 titre (logged):Time (interaction) | 0.0011 | 0.00052 | 0.044 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luu, L.; Bah, G.S.; Okah-Nnane, N.H.; Hartley, C.S.; Glover, A.F.; Walsh, T.R.; Lian, L.-Y.; Zhan, B.; Bottazzi, M.E.; Abraham, D.; et al. Co-Administration of Adjuvanted Recombinant Ov-103 and Ov-RAL-2 Vaccines Confer Protection against Natural Challenge in A Bovine Onchocerca ochengi Infection Model of Human Onchocerciasis. Vaccines 2022, 10, 861. https://doi.org/10.3390/vaccines10060861
Luu L, Bah GS, Okah-Nnane NH, Hartley CS, Glover AF, Walsh TR, Lian L-Y, Zhan B, Bottazzi ME, Abraham D, et al. Co-Administration of Adjuvanted Recombinant Ov-103 and Ov-RAL-2 Vaccines Confer Protection against Natural Challenge in A Bovine Onchocerca ochengi Infection Model of Human Onchocerciasis. Vaccines. 2022; 10(6):861. https://doi.org/10.3390/vaccines10060861
Chicago/Turabian StyleLuu, Lisa, Germanus S. Bah, Ndode Herman Okah-Nnane, Catherine S. Hartley, Alexandra F. Glover, Tessa R. Walsh, Lu-Yun Lian, Bin Zhan, Maria Elena Bottazzi, David Abraham, and et al. 2022. "Co-Administration of Adjuvanted Recombinant Ov-103 and Ov-RAL-2 Vaccines Confer Protection against Natural Challenge in A Bovine Onchocerca ochengi Infection Model of Human Onchocerciasis" Vaccines 10, no. 6: 861. https://doi.org/10.3390/vaccines10060861