
Prospective, Longitudinal Study on Specific Cellular Immune Responses after Vaccination with an Adjuvanted, Recombinant Zoster Vaccine in Kidney Transplant Recipients

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Volunteers
2.2. Vaccine
2.3. ELISpot Assay
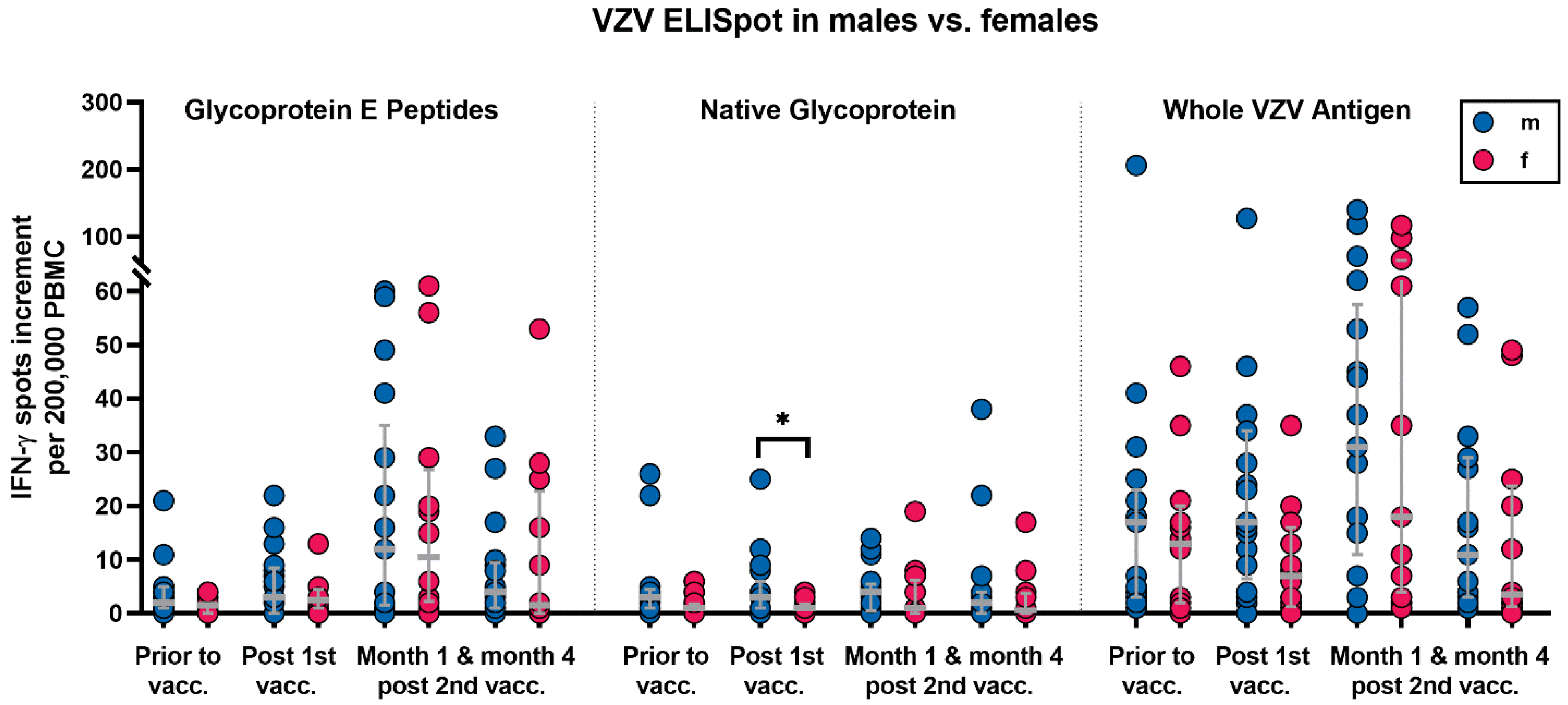
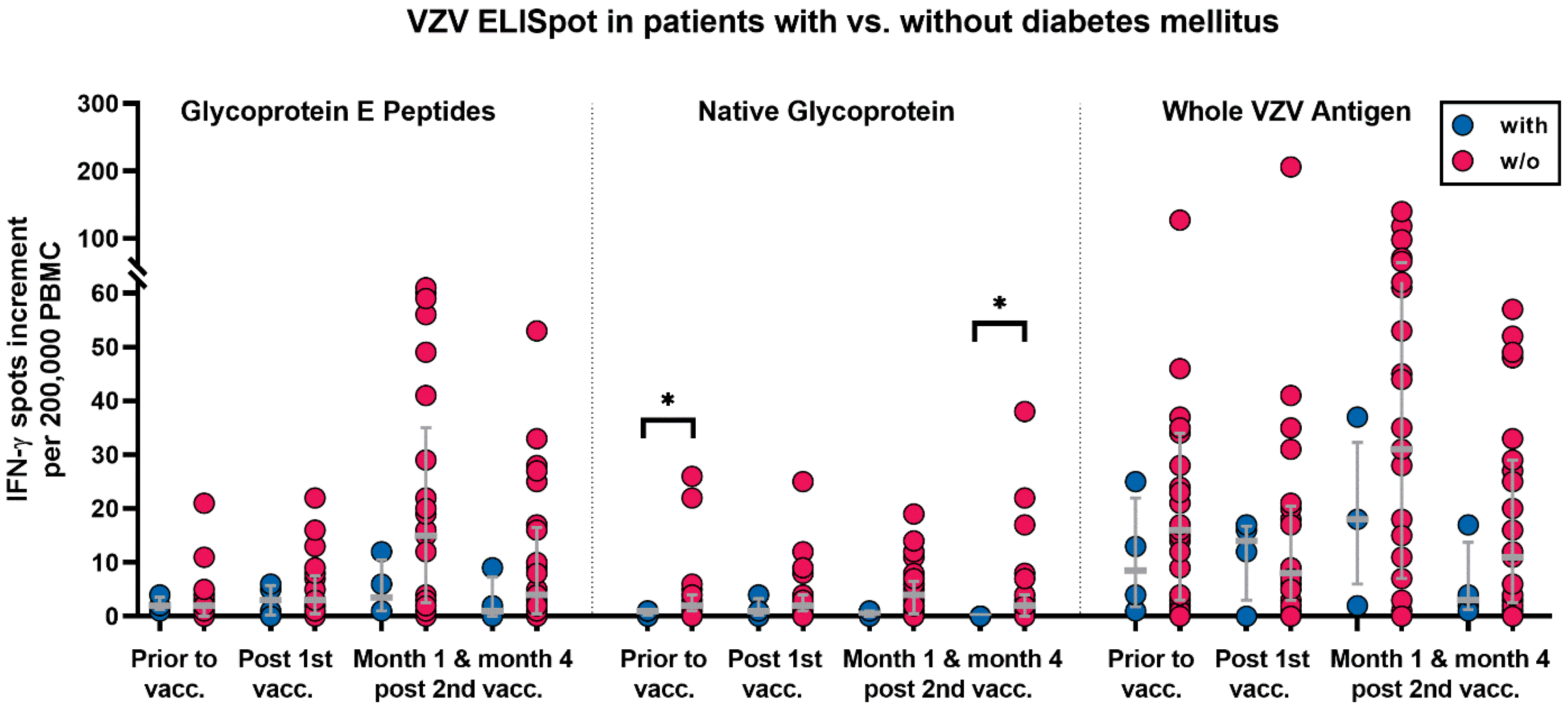
2.4. Parameters with Potential Influence on Vaccination Responses
2.5. Statistical Analysis
3. Results
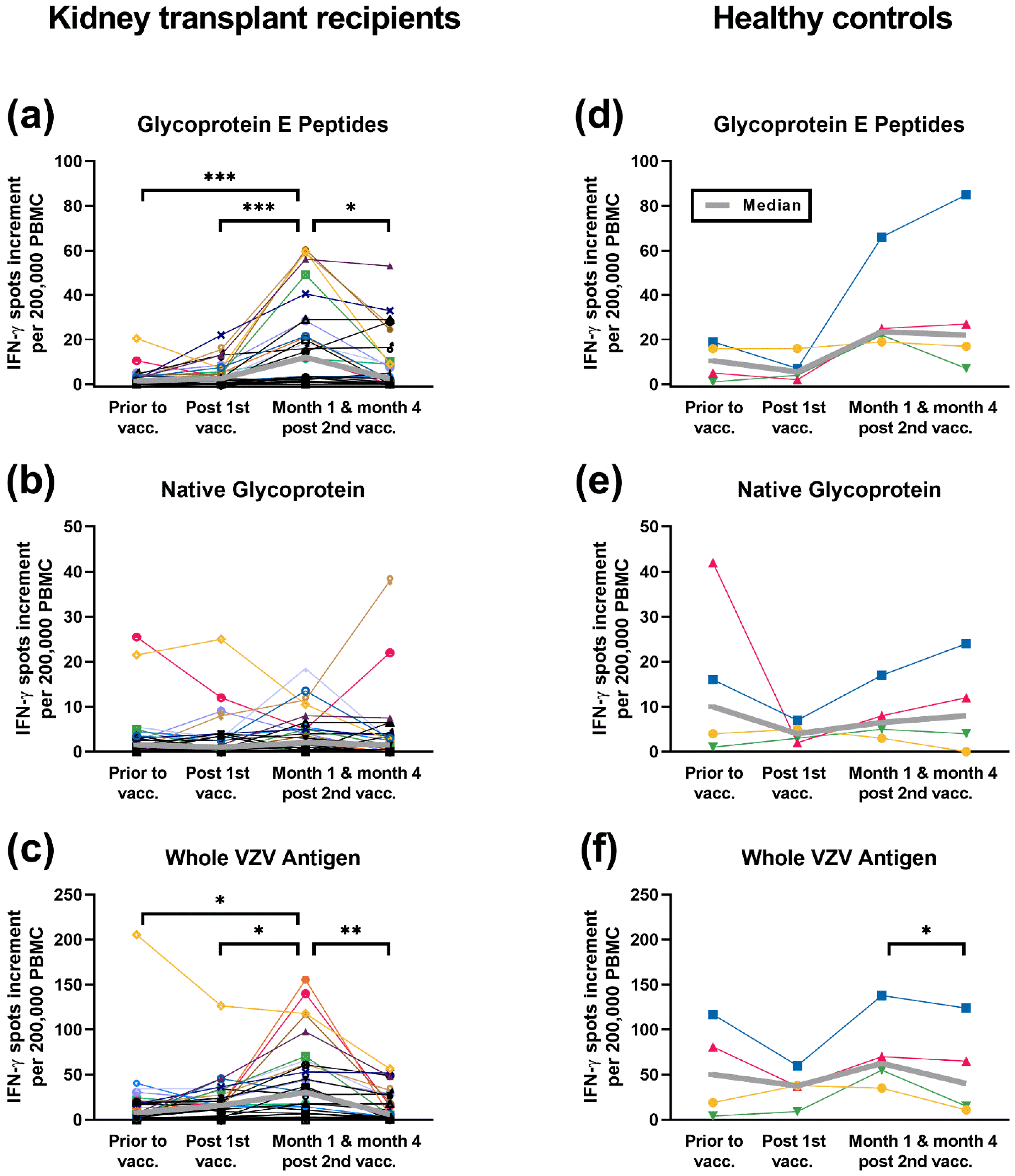
3.1. Time Course of ELISpot Responses to Three Different VZV Antigens
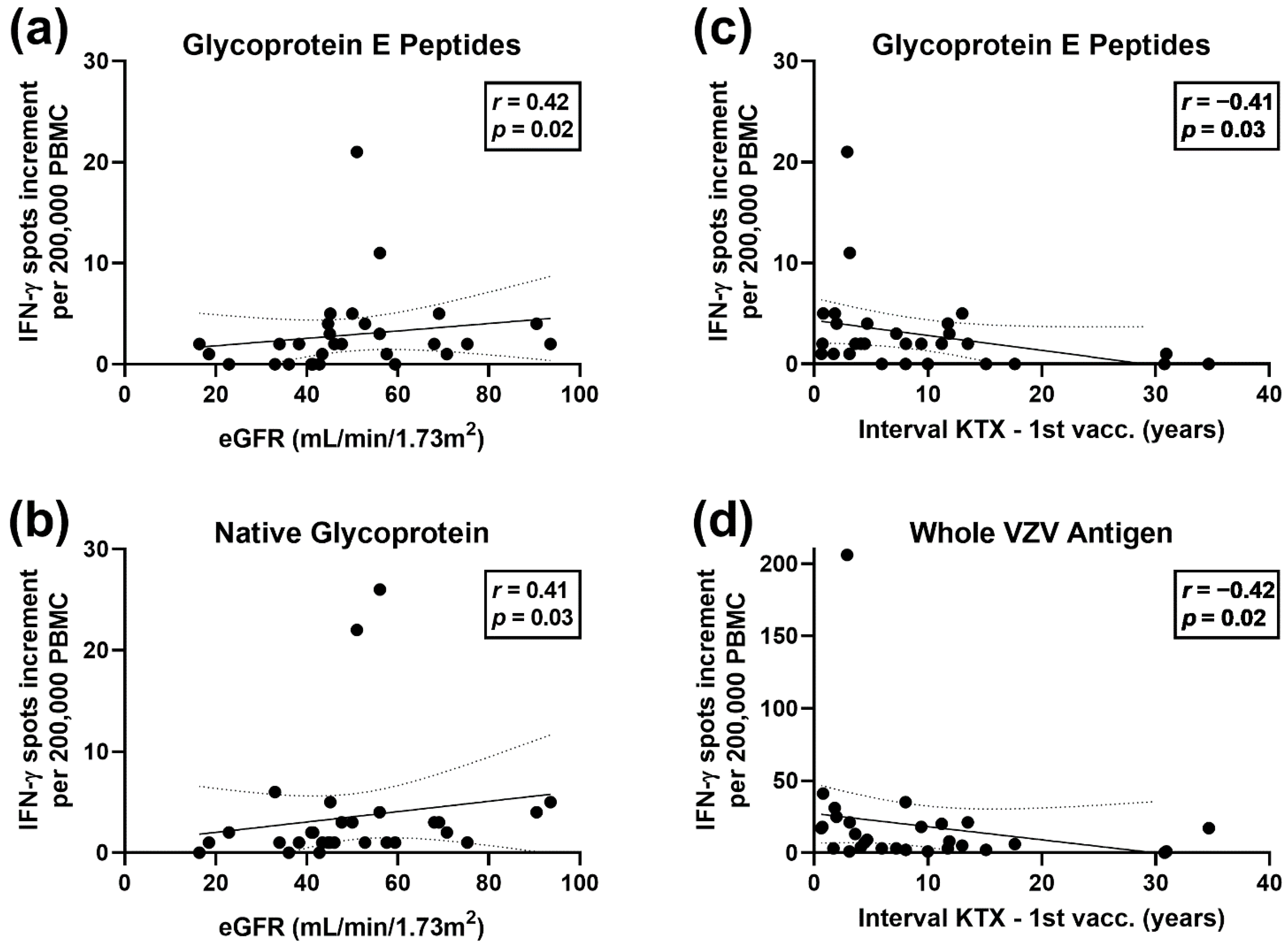
3.2. Correlation of VZV-Specific Cellular Immunity with Clinical Parameters
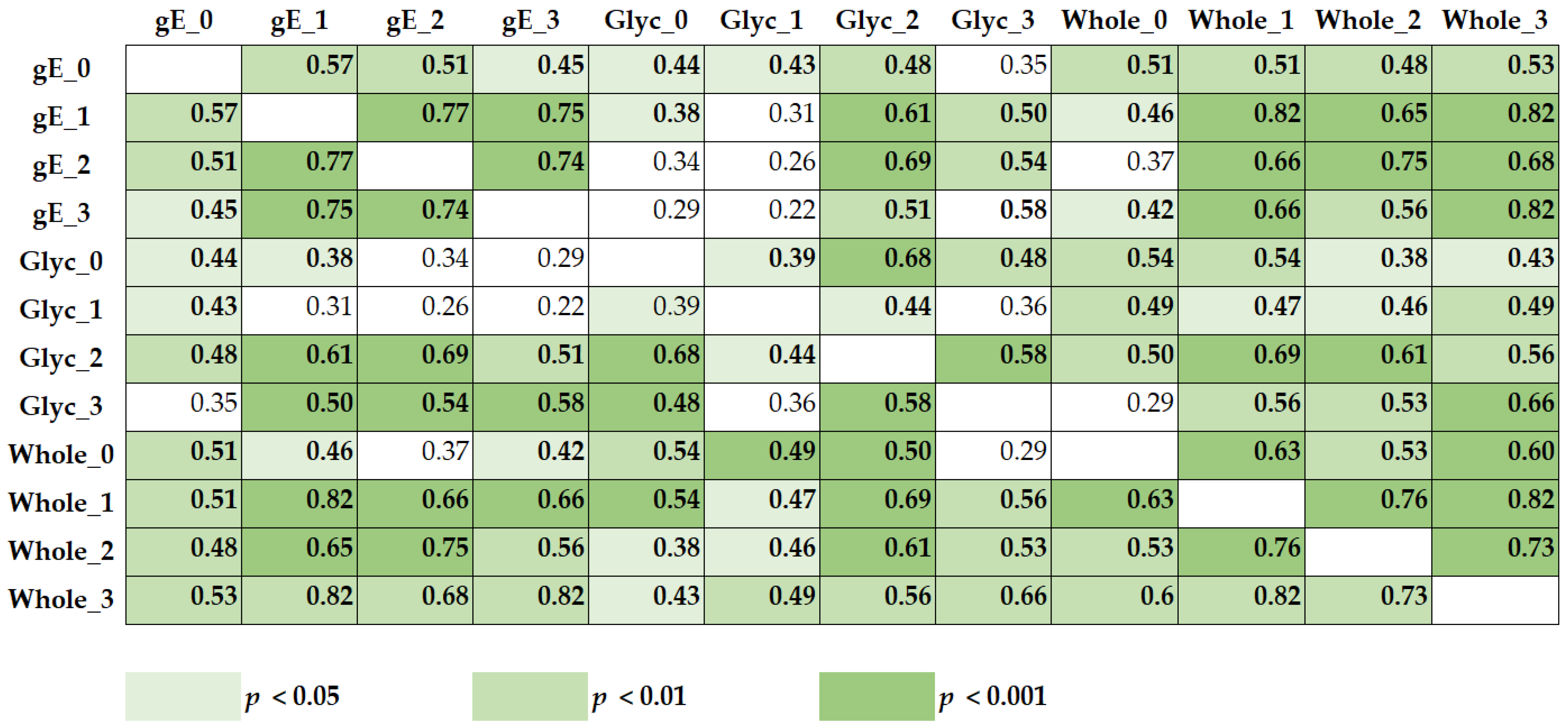
3.3. Correlation of VZV-Specific Immunity Measured with Various VZV Antigens and at Various Time Points
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sauerbrei, A. Diagnosis, antiviral therapy, and prophylaxis of varicella-zoster virus infections. Eur. J. Clin. Microbiol. Infect. Dis. 2016, 35, 723–734. [Google Scholar] [CrossRef] [PubMed]
- Hillebrand, K.; Bricout, H.; Schulze-Rath, R.; Schink, T.; Garbe, E. Incidence of herpes zoster and its complications in Germany, 2005–2009. J. Infect. 2015, 70, 178–186. [Google Scholar] [CrossRef] [PubMed]
- Gershon, A.A.; Mervish, N.; LaRussa, P.; Steinberg, S.; Lo, S.H.; Hodes, D.; Fikrig, S.; Bonagura, V.; Bakshi, S. Varicella-zoster virus infection in children with underlying human immunodeficiency virus infection. J. Infect. Dis. 1997, 176, 1496–1500. [Google Scholar] [CrossRef] [PubMed]
- Arvin, A. Aging, immunity, and the varicella-zoster virus. N. Engl. J. Med. 2005, 352, 2266–2267. [Google Scholar] [CrossRef]
- Vink, P.; Ramon Torrell, J.M.; Sanchez Fructuoso, A.; Kim, S.J.; Kim, S.I.; Zaltzman, J.; Ortiz, F.; Campistol Plana, J.M.; Fernandez Rodriguez, A.M.; Rebollo Rodrigo, H.; et al. Immunogenicity and Safety of the Adjuvanted Recombinant Zoster Vaccine in Chronically Immunosuppressed Adults Following Renal Transplant: A Phase 3, Randomized Clinical Trial. Clin. Infect. Dis. 2020, 70, 181–190. [Google Scholar] [CrossRef]
- Insinga, R.P.; Itzler, R.F.; Pellissier, J.M.; Saddier, P.; Nikas, A.A. The incidence of herpes zoster in a United States administrative database. J. Gen. Intern. Med. 2005, 20, 748–753. [Google Scholar] [CrossRef] [Green Version]
- Whitley, R.J.; Siebenhaar, F.; Sterry, W. Varicella-Zoster-Virus Infections. In Harrison’s Principles of Internal Medicine (Deutsche Ausgabe), 16th ed.; Dietel, M., Suttrop, N., Zeitz, M., Eds.; Union Druckerei: Weimar, Germany, 2005; Volume 1, pp. 1121–1124. [Google Scholar]
- Robert-Koch-Institut. Empfehlungen der Ständigen Impfkommission beim Robert Koch-Institut 2021. Epid. Bull. 2021, 34, 4–38. [Google Scholar]
- U.S. Department of Health & Human Services. HHS.gov Immunization. Available online: https://www.hhs.gov/immunization/diseases/chickenpox/index.html (accessed on 9 May 2022).
- Robert-Koch-Institut. Schutzimpfung Gegen Herpes Zoster (Gürtelrose). Available online: https://www.rki.de/DE/Content/Infekt/Impfen/Materialien/Faktenblaetter/Zoster.html;jsessionid=80F817FC1C302CB2CD1F2D9F5F5641D5.internet071?nn=2375548 (accessed on 9 May 2022).
- Cohen, J.I.; Straus, S.E.; Arvin, A.M. Varicella-zoster virus replication, pathogenesis, and management. In Fields Virology, 5th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2007; Volume 2, pp. 2773–2818. [Google Scholar]
- Czerkinsky, C.; Andersson, G.; Ekre, H.P.; Nilsson, L.A.; Klareskog, L.; Ouchterlony, O. Reverse ELISPOT assay for clonal analysis of cytokine production. I. Enumeration of gamma-interferon-secreting cells. J. Immunol. Methods 1988, 110, 29–36. [Google Scholar] [CrossRef]
- Murthy, K.; Stevens, L.A.; Stark, P.C.; Levey, A.S. Variation in the serum creatinine assay calibration: A practical application to glomerular filtration rate estimation. Kidney Int. 2005, 68, 1884–1887. [Google Scholar] [CrossRef] [Green Version]
- European Medicines Agency. Shingrix-EPAR-Product Information. Available online: https://www.ema.europa.eu/en/medicines/human/EPAR/shingrix (accessed on 9 May 2022).
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef]
- Lindemann, M.; Horn, P.A.; Koldehoff, M. Cellular Immune Response after Vaccination with an Adjuvanted, Recombinant Zoster Vaccine in Allogeneic Hematopoietic Stem Cell Transplant Recipients. Vaccines 2022, 10, 809. [Google Scholar] [CrossRef]
- Arvin, A.M. Humoral and cellular immunity to varicella-zoster virus: An overview. J. Infect. Dis. 2008, 197 (Suppl. S2), S58–S60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, C.J.; Kuter, B.J.; Ngai, A.; Hildebrand, C.S.; Isganitis, K.L.; Patterson, C.M.; Capra, A.; Miller, W.J.; Krah, D.L.; Provost, P.J.; et al. Modified cases of chickenpox after varicella vaccination: Correlation of protection with antibody response. Pediatr. Infect. Dis. J. 1992, 11, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Feyssaguet, M.; Berthold, V.; Helle, L.; Povey, M.; Ravault, S.; Carryn, S.; Gillard, P.; Di Paolo, E. Comparison of a glycoprotein E-based ELISA with a varicella-zoster whole-virus ELISA for the quantification of varicella vaccine immune responses in young children. Vaccine 2020, 38, 3300–3304. [Google Scholar] [CrossRef]
- Sauerbrei, A.; Schafler, A.; Hofmann, J.; Schacke, M.; Gruhn, B.; Wutzler, P. Evaluation of three commercial varicella-zoster virus IgG enzyme-linked immunosorbent assays in comparison to the fluorescent-antibody-to-membrane-antigen test. Clin. Vaccine Immunol. 2012, 19, 1261–1268. [Google Scholar] [CrossRef] [Green Version]
- Cassaniti, I.; Ferrari, A.; Comolli, G.; Sarasini, A.; Gregorini, M.; Rampino, T.; Lilleri, D.; Baldanti, F. Characterization of Varicella-Zoster (VZV) Specific T Cell Response in Healthy Subjects and Transplanted Patients by Using Enzyme Linked Immunospot (ELISpot) Assays. Vaccines 2021, 9, 875. [Google Scholar] [CrossRef]
- Syed-Ahmed, M.; Narayanan, M. Immune Dysfunction and Risk of Infection in Chronic Kidney Disease. Adv. Chronic Kidney Dis. 2019, 26, 8–15. [Google Scholar] [CrossRef]
- Lindemann, M.; Korth, J.; Sun, M.; Xu, S.; Struve, C.; Werner, K.; Dornieden, T.; Horn, P.A.; Witzke, O.; Wilde, B. The Cytomegalovirus-Specific IL-21 ELISpot Correlates with Allograft Function of Kidney Transplant Recipients. Int. J. Mol. Sci. 2018, 19, 3945. [Google Scholar] [CrossRef] [Green Version]
- Koc, Y.; Miller, K.B.; Schenkein, D.P.; Griffith, J.; Akhtar, M.; DesJardin, J.; Snydman, D.R. Varicella zoster virus infections following allogeneic bone marrow transplantation: Frequency, risk factors, and clinical outcome. Biol. Blood Marrow Transplant. 2000, 6, 44–49. [Google Scholar] [CrossRef] [Green Version]
- Lindemann, M.; Wilde, B.; Friebus-Kardash, J.; Gackler, A.; Witzke, O.; Dittmer, U.; Horn, P.A.; Kribben, A.; Mulling, N.; Eisenberger, U. Comparison of Humoral and Cellular CMV Immunity in Patients Awaiting Kidney Transplantation. Diagnostics 2021, 11, 1688. [Google Scholar] [CrossRef]
- Lindemann, M.; Schuett, P.; Moritz, T.; Ottinger, H.D.; Opalka, B.; Seeber, S.; Nowrousian, M.R.; Grosse-Wilde, H. Cellular in vitro immune function in multiple myeloma patients after high-dose chemotherapy and autologous peripheral stem cell transplantation. Leukemia 2005, 19, 490–492. [Google Scholar] [CrossRef] [PubMed]
- Dolff, S.; Korth, J.; Jahn, M.; Kribben, A.; Witzke, O.; Wilde, B. Anti-SARS-CoV-2 T-cell Responses After mRNA Vaccination in Belatacept-treated Renal Transplant Patients. Transplantation 2021, 105, e99. [Google Scholar] [CrossRef] [PubMed]
- Chavarot, N.; Ouedrani, A.; Marion, O.; Leruez-Ville, M.; Vilain, E.; Baaziz, M.; Del Bello, A.; Burger, C.; Sberro-Soussan, R.; Martinez, F.; et al. Poor Anti-SARS-CoV-2 Humoral and T-cell Responses after 2 Injections of mRNA Vaccine in Kidney Transplant Recipients Treated With Belatacept. Transplantation 2021, 105, e94–e95. [Google Scholar] [CrossRef]
- Liefeldt, L.; Glander, P.; Klotsche, J.; Straub-Hohenbleicher, H.; Budde, K.; Eberspacher, B.; Friedersdorff, F.; Halleck, F.; Hambach, P.; Hofmann, J.; et al. Predictors of Serological Response to SARS-CoV-2 Vaccination in Kidney Transplant Patients: Baseline Characteristics, Immunosuppression, and the Role of IMPDH Monitoring. J. Clin. Med. 2022, 11, 1697. [Google Scholar] [CrossRef] [PubMed]
- Klein, S.L. Sex influences immune responses to viruses, and efficacy of prophylaxis and treatments for viral diseases. Bioessays 2012, 34, 1050–1059. [Google Scholar] [CrossRef] [Green Version]
- Bernin, H.; Fehling, H.; Marggraff, C.; Tannich, E.; Lotter, H. The cytokine profile of human NKT cells and PBMCs is dependent on donor sex and stimulus. Med. Microbiol. Immunol. 2016, 205, 321–332. [Google Scholar] [CrossRef] [Green Version]
- Di Benedetto, S.; Derhovanessian, E.; Steinhagen-Thiessen, E.; Goldeck, D.; Muller, L.; Pawelec, G. Impact of age, sex and CMV-infection on peripheral T cell phenotypes: Results from the Berlin BASE-II Study. Biogerontology 2015, 16, 631–643. [Google Scholar] [CrossRef]
- Villacres, M.C.; Longmate, J.; Auge, C.; Diamond, D.J. Predominant type 1 CMV-specific memory T-helper response in humans: Evidence for gender differences in cytokine secretion. Hum. Immunol. 2004, 65, 476–485. [Google Scholar] [CrossRef]
- Klein, S.L.; Flanagan, K.L. Sex differences in immune responses. Nat. Rev. Immunol. 2016, 16, 626–638. [Google Scholar] [CrossRef]
- Boef, A.G.C.; van der Klis, F.R.M.; Berbers, G.A.M.; Buisman, A.M.; Sanders, E.A.M.; Kemmeren, J.M.; van der Ende, A.; de Melker, H.E.; Rots, N.Y.; Knol, M.J. Differences by sex in IgG levels following infant and childhood vaccinations: An individual participant data meta-analysis of vaccination studies. Vaccine 2018, 36, 400–407. [Google Scholar] [CrossRef]
- Giefing-Kroll, C.; Berger, P.; Lepperdinger, G.; Grubeck-Loebenstein, B. How sex and age affect immune responses, susceptibility to infections, and response to vaccination. Aging Cell 2015, 14, 309–321. [Google Scholar] [CrossRef] [PubMed]
- Lindemann, M.; Klisanin, V.; Thummler, L.; Fisenkci, N.; Tsachakis-Muck, N.; Ditschkowski, M.; Schwarzkopf, S.; Klump, H.; Reinhardt, H.C.; Horn, P.A.; et al. Humoral and Cellular Vaccination Responses against SARS-CoV-2 in Hematopoietic Stem Cell Transplant Recipients. Vaccines 2021, 9, 1075. [Google Scholar] [CrossRef] [PubMed]
- Gackler, A.; Mulling, N.; Volk, K.; Wilde, B.; Eisenberger, U.; Rohn, H.; Horn, P.A.; Witzke, O.; Lindemann, M. Establishment of an ELISpot Assay to Detect Cellular Immunity against S. pneumoniae in Vaccinated Kidney Transplant Recipients. Vaccines 2021, 9, 1438. [Google Scholar] [CrossRef] [PubMed]
- Berbudi, A.; Rahmadika, N.; Tjahjadi, A.I.; Ruslami, R. Type 2 Diabetes and its Impact on the Immune System. Curr. Diabetes Rev. 2020, 16, 442–449. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion | Exclusion |
|---|---|
| Age ≥ 45 years | Acute rejection 2 |
| Interval to kidney transplantation ≥ 6 months | Active shingles infection |
| Interval to shingles ≥ 2 months | Acute (other) infection (fever > 38.5 °C) |
| Stable kidney function 1 | Actual malignant tumor |
| Complete clinical dataset | Allergy against a component of the vaccine |
| Sequential ELISpot data at four time points | Pregnancy |
| Written informed consent | Inability to consent |
| Variable | Group | Absolute Number or Median (Range) |
|---|---|---|
| Sex | Female | 12 |
| Male | 17 | |
| Age (years) | 61 (45–79) | |
| Kidney transplantation, no. | First | 24 |
| Second | 5 | |
| eGFR | Prior to vacc. | 46 (16–94) |
| (mL/min/1.73 m2) | Post 1st vacc. | 49 (12–99) |
| M1 post 2nd vacc. | 51 (14–94) | |
| M4 post 2nd vacc. | 47 (15–88) | |
| Co-morbidities | Diabetes mellitus | 4 |
| Hypertension | 12 | |
| Coronary heart disease | 8 | |
| Previous malignant tumor | 11 | |
| COPD | 4 | |
| Anamnesis of | Cytomegalovirus | 11 |
| previous infection | Herpes simplex virus type 1 | 2 |
| with herpesviruses | VZV (chickenpox) | 29 |
| VZV (shingles) | 8 | |
| Previous | Aciclovir | 1 |
| antiviral treatment | Valganciclovir | 3 |
| Entecavir | 1 | |
| Cytotect | 1 | |
| Immunosuppressive | Tacrolimus | 25 |
| therapy | Mycophenolate | 20 |
| Corticosteroids | 26 | |
| Everolimus | 5 | |
| Azathioprine | 1 | |
| Ciclosporin | 1 | |
| Belatacept | 2 | |
| Allograft rejection | Total | 6 |
| Acute | 5 | |
| Acute and chronic | 1 | |
| Humoral | 2 | |
| Cellular | 3 | |
| Humoral and Cellular | 1 | |
| Interval transplantation– 1st vaccination | 7.2 years (8 months–34.7 years) | |
| Interval 1st vaccination– 2nd vaccination | 71 days (62–149) | |
| Interval 2nd vaccination– blood sampling | ||
| First follow-up | 1.2 months (0.9–1.9) | |
| Second follow-up | 4.2 months (3.7–9.6) |
| Antigen | Time Point | KTX | HC | p | ||||
|---|---|---|---|---|---|---|---|---|
| Median | MIN | MAX | Median | MIN | MAX | |||
| Glycoprotein E Peptides | Pre vacc. | 1.5 | −0.5 | 20.5 | 10.5 | 1 | 19 | 0.07 |
| post 1st vacc. | 2.5 | −1 | 22 | 5.5 | 2 | 16 | 0.11 | |
| M1 post 2nd vacc. | 12 | 0 | 60.5 | 23.5 | 19 | 66 | 0.09 | |
| M4 post 2nd vacc. | 2.5 | 0 | 53 | 22 | 7 | 85 | 0.04 * | |
| Native Glycoprotein | Pre vacc. | 1.5 | 0 | 25.5 | 10 | 1 | 42 | 0.10 |
| post 1st vacc. | 1 | 0 | 25 | 4 | 2 | 7 | 0.15 | |
| M1 post 2nd vacc. | 2 | 0 | 18.5 | 6.5 | 3 | 17 | 0.09 | |
| M4 post 2nd vacc. | 1.5 | 0 | 38 | 8 | 0 | 24 | 0.17 | |
| Whole VZV Antigen | Pre vacc. | 7.5 | 0 | 205.5 | 50 | 4 | 117 | 0.10 |
| post 1st vacc. | 16 | 0 | 126.5 | 37.5 | 9 | 60 | 0.09 | |
| M1 post 2nd vacc. | 30.5 | 0 | 155.5 | 62.5 | 35 | 138 | 0.08 | |
| M4 post 2nd vacc. | 6 | 0 | 56.5 | 40 | 11 | 124 | 0.07 |
| Variable | Antigen | Time Point | Treatment Received | Treatment Not Received | p | ||||
|---|---|---|---|---|---|---|---|---|---|
| Median | MIN | MAX | Median | MIN | MAX | ||||
| Tacrolimus | Glycoprotein E Peptides | Pre vacc. | 2 | 0 | 21 | 1.5 | 0 | 2 | 0.32 |
| post 1st vacc. | 3 | 0 | 22 | 1 | 0 | 2 | 0.06 | ||
| M1 post 2nd vacc. | 15 | 0 | 61 | 1.5 | 0 | 3 | 0.02 * | ||
| M4 post 2nd vacc. | 5 | 0 | 53 | 0.5 | 0 | 1 | 0.05 | ||
| Native Glycoprotein | Pre vacc. | 2 | 0 | 26 | 1.5 | 1 | 3 | 0.74 | |
| post 1st vacc. | 2 | 0 | 25 | 1 | 0 | 4 | 0.34 | ||
| M1 post 2nd vacc. | 4 | 0 | 19 | 0.5 | 0 | 2 | 0.12 | ||
| M4 post 2nd vacc. | 2 | 0 | 38 | 0 | 0 | 1 | 0.06 | ||
| Whole VZV Antigen | Pre vacc. | 8 | 0 | 206 | 9.5 | 1 | 21 | 0.55 | |
| post 1st vacc. | 16 | 0 | 127 | 6 | 2 | 17 | 0.21 | ||
| M1 post 2nd vacc. | 35 | 0 | 140 | 7 | 1 | 11 | 0.03 * | ||
| M4 post 2nd vacc. | 12 | 0 | 57 | 2 | 1 | 3 | 0.04 * | ||
| Mycophenolate | Glycoprotein E Peptides | Pre vacc. | 2 | 0 | 21 | 0 | 0 | 4 | 0.03 * |
| post 1st vacc. | 3 | 0 | 22 | 1 | 0 | 8 | 0.06 | ||
| M1 post 2nd vacc. | 13.5 | 0 | 60 | 3 | 0 | 61 | 0.33 | ||
| M4 post 2nd vacc. | 8.5 | 0 | 53 | 0 | 0 | 25 | 0.045 * | ||
| Native Glycoprotein | Pre vacc. | 2.5 | 1 | 26 | 1 | 0 | 4 | 0.03 * | |
| post 1st vacc. | 2.5 | 0 | 25 | 1 | 0 | 4 | 0.21 | ||
| M1 post 2nd vacc. | 4 | 0 | 19 | 1 | 0 | 14 | 0.08 | ||
| M4 post 2nd vacc. | 3 | 0 | 38 | 0 | 0 | 3 | 0.03 * | ||
| Whole VZV Antigen | Pre vacc. | 17.5 | 1 | 206 | 3 | 0 | 9 | 0.002 * | |
| post 1st vacc. | 19 | 0 | 127 | 3 | 0 | 46 | 0.01 * | ||
| M1 post 2nd vacc. | 36 | 1 | 140 | 18 | 0 | 117 | 0.24 | ||
| M4 post 2nd vacc. | 16.5 | 2 | 57 | 1 | 0 | 20 | 0.006 * | ||
| Antigen | Time Point | eGFR | Interval to KTX 1 | Sex | Diabetes Mellitus | Tacrolimus | Mycophenolate |
|---|---|---|---|---|---|---|---|
| Glycoprotein E Peptides | Pre vacc. | <0.0001 | 0.002 | <0.0001 | <0.0001 | ||
| post 1st vacc. | |||||||
| M1 post 2nd vacc. | <0.0001 | ||||||
| M4 post 2nd vacc. | <0.0001 | ||||||
| Native Glycoprotein | Pre vacc. | 0.02 | 0.01 | <0.0001 | 0.02 | ||
| post 1st vacc. | 0.046 | ||||||
| M1 post 2nd vacc. | <0.0001 | <0.0001 | |||||
| M4 post 2nd vacc. | <0.0001 | <0.0001 | 0.0001 | 0.006 | <0.0001 | ||
| Whole VZV Antigen | Pre vacc. | 0.01 | 0.02 | 0.001 | |||
| post 1st vacc. | 0.003 | <0.0001 | <0.0001 | ||||
| M1 post 2nd vacc. | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.01 | |
| M4 post 2nd vacc. | 0.04 | <0.0001 | <0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lindemann, M.; Baumann, C.; Wilde, B.; Gäckler, A.; Meller, L.; Horn, P.A.; Krawczyk, A.; Witzke, O. Prospective, Longitudinal Study on Specific Cellular Immune Responses after Vaccination with an Adjuvanted, Recombinant Zoster Vaccine in Kidney Transplant Recipients. Vaccines 2022, 10, 844. https://doi.org/10.3390/vaccines10060844
Lindemann M, Baumann C, Wilde B, Gäckler A, Meller L, Horn PA, Krawczyk A, Witzke O. Prospective, Longitudinal Study on Specific Cellular Immune Responses after Vaccination with an Adjuvanted, Recombinant Zoster Vaccine in Kidney Transplant Recipients. Vaccines. 2022; 10(6):844. https://doi.org/10.3390/vaccines10060844
Chicago/Turabian StyleLindemann, Monika, Charleen Baumann, Benjamin Wilde, Anja Gäckler, Lara Meller, Peter A. Horn, Adalbert Krawczyk, and Oliver Witzke. 2022. "Prospective, Longitudinal Study on Specific Cellular Immune Responses after Vaccination with an Adjuvanted, Recombinant Zoster Vaccine in Kidney Transplant Recipients" Vaccines 10, no. 6: 844. https://doi.org/10.3390/vaccines10060844