
Live-Attenuated Oral Vaccines to Reduce Campylobacter Colonization in Poultry



Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Culture Conditions
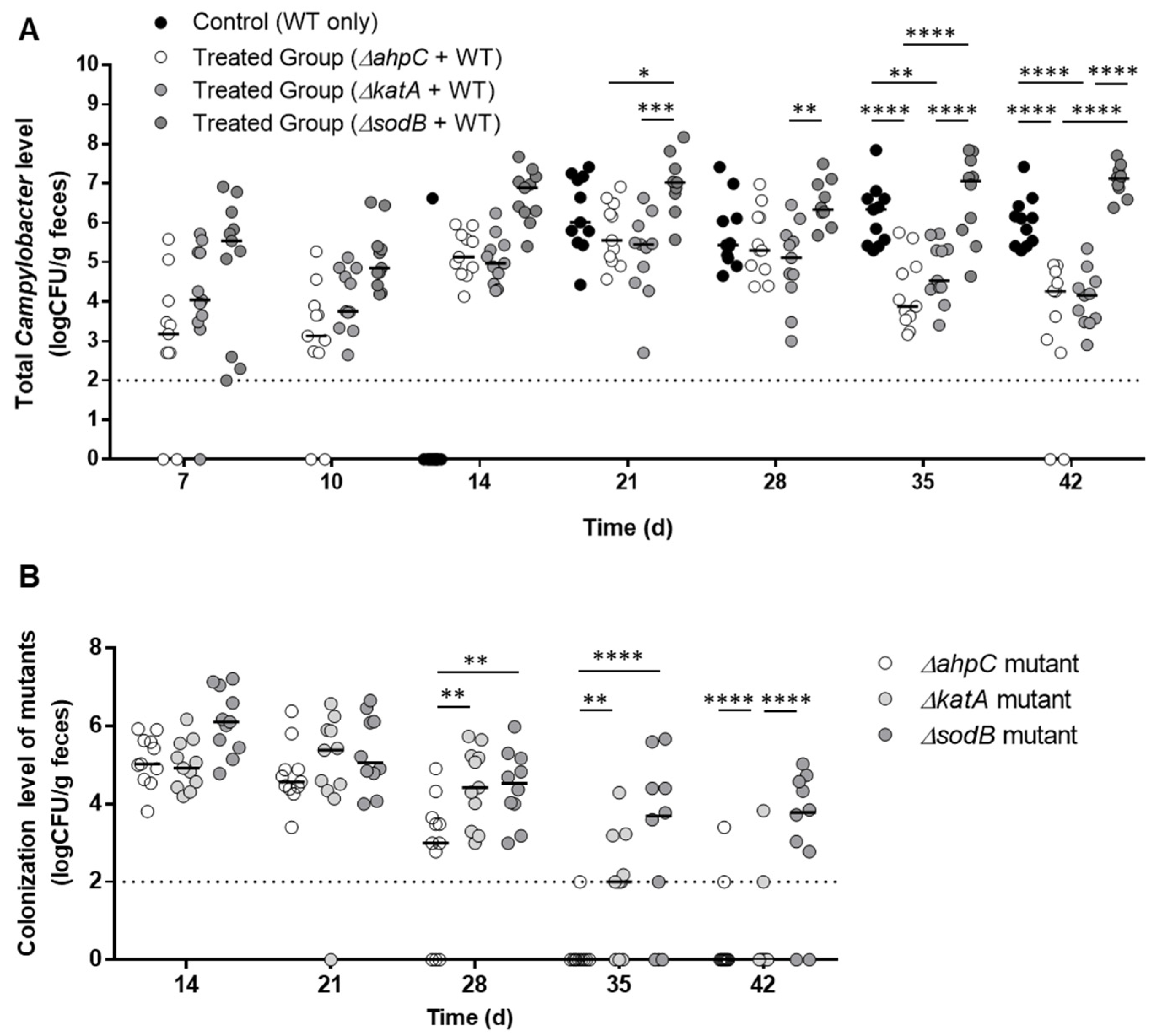
2.2. Evaluation of Efficacy in Preventing Campylobacter Colonization by Pre-Colonization with Oxidative Stress Defense Mutants
3. Results
3.1. Prevention of C. jejuni Colonization by Pre-Colonization with Oxidative Stress Defense Mutants
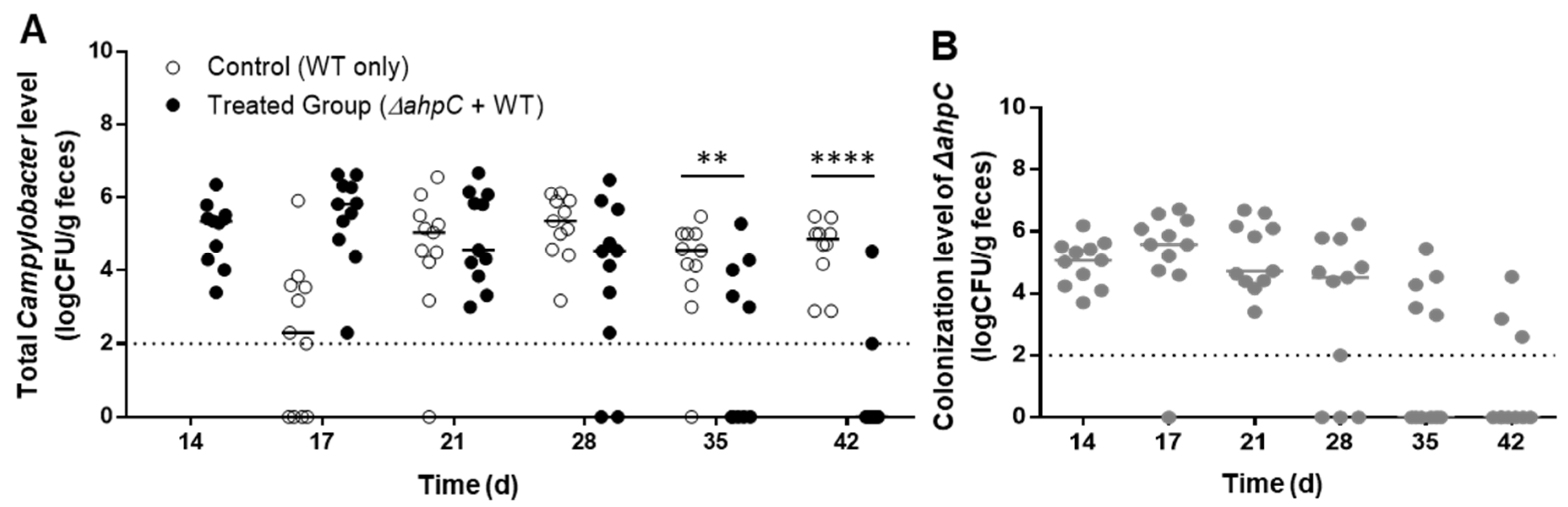
3.2. C. jejuni Reduction by Challenge with the ΔahpC Mutant A High Dose
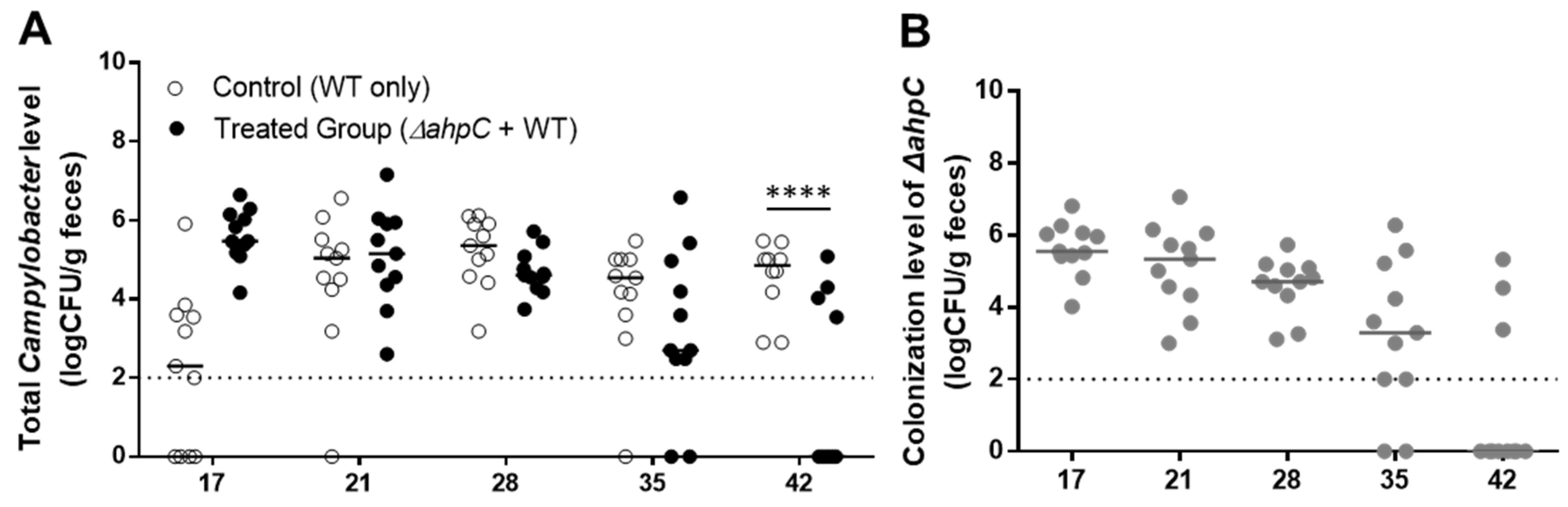
3.3. C. jejuni Reduction by Challenge with the ΔahpC Mutant A Low Dose
3.4. Increase in Chicken Body Weight after Pre-Colonization with the Δahpc Mutant
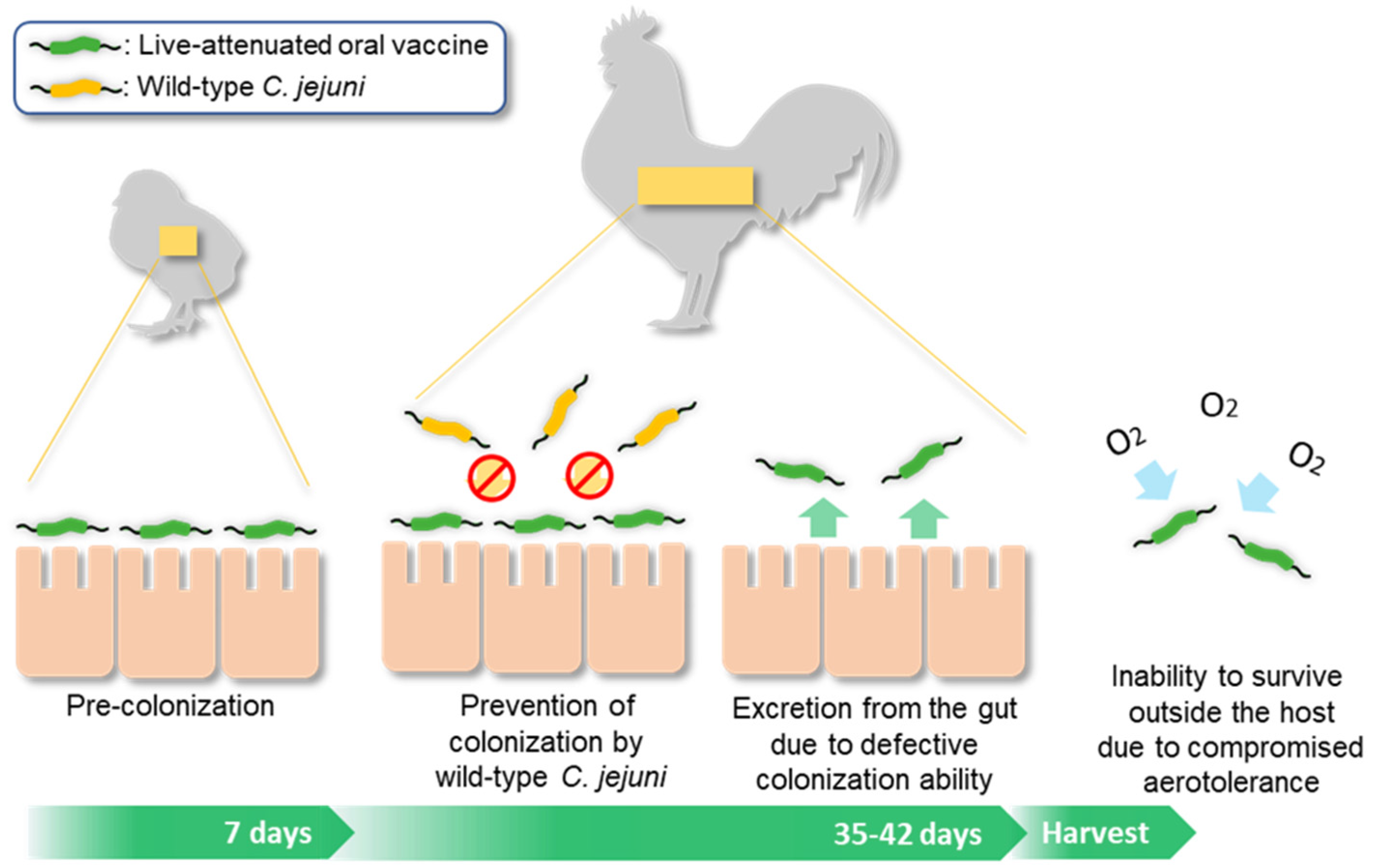
4. Discussion
5. Conclusions
6. Patents
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Kirk, M.D.; Pires, S.M.; Black, R.E.; Caipo, M.; Crump, J.A.; Devleesschauwer, B.; Dopfer, D.; Fazil, A.; Fischer-Walker, C.L.; Hald, T.; et al. World health organization estimates of the global and regional disease burden of 22 foodborne bacterial, protozoal, and viral diseases, 2010: A data synthesis. PLoS Med. 2015, 12, e1001921. [Google Scholar] [CrossRef] [Green Version]
- Poropatich, K.O.; Walker, C.L.; Black, R.E. Quantifying the association between Campylobacter infection and Guillain-Barre syndrome: A systematic review. J. Health Popul. Nutr. 2010, 28, 545–552. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heimesaat, M.M.; Backert, S.; Alter, T.; Bereswill, S. Human campylobacteriosis-a serious infectious threat in a one health perspective. Curr. Top. Microbiol. Immunol. 2021, 431, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Jeon, B.; Muraoka, W.T.; Zhang, Q. Advances in Campylobacter biology and implications for biotechnological applications. Microb. Biotechnol. 2010, 3, 242–258. [Google Scholar] [CrossRef] [Green Version]
- Davis, L.; DiRita, V. Growth and laboratory maintenance of Campylobacter jejuni. Curr. Protocols Microbiol. 2008, 8, 8A.1.1–8A.1.7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasschaert, G.; De Zutter, L.; Herman, L.; Heyndrickx, M. Campylobacter contamination of broilers: The role of transport and slaughterhouse. Int. J. Food Microbiol. 2020, 322, 108564. [Google Scholar] [CrossRef]
- Nauta, M.J.; Johannessen, G.; Laureano Adame, L.; Williams, N.; Rosenquist, H. The effect of reducing numbers of Campylobacter in broiler intestines on human health risk. Microbial. Risk Anal. 2016, 2–3, 68–77. [Google Scholar] [CrossRef]
- Rosenquist, H.; Nielsen, N.L.; Sommer, H.M.; Norrung, B.; Christensen, B.B. Quantitative risk assessment of human campylobacteriosis associated with thermophilic Campylobacter species in chickens. Int. J. Food Microbiol. 2003, 83, 87–103. [Google Scholar] [CrossRef]
- Lin, I.Y.; Van, T.T.; Smooker, P.M. Live-attenuated bacterial vectors: Tools for vaccine and therapeutic agent delivery. Vaccines 2015, 3, 940–972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krieg, N.R.; Hoffman, P.S. Microaerophily and oxygen toxicity. Annu. Reviron. Microbiol. 1986, 40, 107–130. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.J.; Kelly, D.J. The function, biogenesis and regulation of the electron transport chains in Campylobacter jejuni: New insights into the bioenergetics of a major food-borne pathogen. Adv. Microbial. Physiol. 2019, 74, 239–329. [Google Scholar]
- Imlay, J.A. Cellular defenses against superoxide and hydrogen peroxide. Annu. Rev. Biochem. 2008, 77, 755–776. [Google Scholar] [CrossRef] [Green Version]
- Parkhill, J.; Wren, B.; Mungall, K.; Ketley, J.; Churcher, C.; Basham, D.; Chillingworth, T.; Davies, R.; Feltwell, T.; Holroyd, S. The genome sequence of the food-borne pathogen Campylobacter jejuni reveals hypervariable sequences. Nature 2000, 403, 665–668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hébrard, M.; Viala, J.P.M.; Méresse, S.; Barras, F.; Aussel, L. Redundant hydrogen peroxide scavengers contribute to Salmonella virulence and oxidative stress resistance. J. Bacteriol. 2009, 191, 4605–4614. [Google Scholar] [CrossRef] [Green Version]
- Oh, E.; McMullen, L.; Jeon, B. Impact of oxidative stress defense on bacterial survival and morphological change in Campylobacter jejuni under aerobic conditions. Front. Microbiol. 2015, 6, 295. [Google Scholar] [CrossRef] [PubMed]
- Baillon, M.L.; van Vliet, A.H.; Ketley, J.M.; Constantinidou, C.; Penn, C.W. An iron-regulated alkyl hydroperoxide reductase (AhpC) confers aerotolerance and oxidative stress resistance to the microaerophilic pathogen Campylobacter jejuni. J. Bacteriol. 1999, 181, 4798–4804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atack, J.M.; Kelly, D.J. Oxidative stress in Campylobacter jejuni: Responses, resistance and regulation. Future Microbiol. 2009, 4, 677–690. [Google Scholar] [CrossRef] [PubMed]
- Day, W.A., Jr.; Sajecki, J.L.; Pitts, T.M.; Joens, L.A. Role of catalase in Campylobacter jejuni intracellular survival. Infect. Immun. 2000, 68, 6337–6345. [Google Scholar] [CrossRef] [PubMed]
- Purdy, D.; Cawthraw, S.; Dickinson, J.H.; Newell, D.G.; Park, S.F. Generation of a superoxide dismutase (SOD)-deficient mutant of Campylobacter coli: Evidence for the significance of SOD in Campylobacter survival and colonization. Appl. Environ. Microbiol. 1999, 65, 2540–2546. [Google Scholar] [CrossRef] [Green Version]
- Hermans, D.; Van Deun, K.; Martel, A.; Van Immerseel, F.; Messens, W.; Heyndrickx, M.; Haesebrouck, F.; Pasmans, F. Colonization factors of Campylobacter jejuni in the chicken gut. Vet. Res. 2011, 42, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palyada, K.; Sun, Y.-Q.; Flint, A.; Butcher, J.; Naikare, H.; Stintzi, A. Characterization of the oxidative stress stimulon and PerR regulon of Campylobacter jejuni. BMC Genom. 2009, 10, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hwang, S.; Ryu, S.; Jeon, B. Roles of the superoxide dismutase SodB and the catalase KatA in the antibiotic resistance of Campylobacter jejuni. J. Antibiot. 2013, 66, 351–353. [Google Scholar] [CrossRef] [PubMed]
- Oh, E.; Jeon, B. Role of alkyl hydroperoxide reductase (AhpC) in the biofilm formation of Campylobacter jejuni. PLoS ONE 2014, 9, e87312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, R.I. Considerations for development of whole cell bacterial vaccines to prevent diarrheal diseases in children in developing countries. Vaccine 2005, 23, 3369–3385. [Google Scholar] [CrossRef]
- Okamura, M.; Tominaga, A.; Ueda, M.; Ohshima, R.; Kobayashi, M.; Tsukada, M.; Yokoyama, E.; Takehara, K.; Deguchi, K.; Honda, T.; et al. Irrelevance between the induction of anti-Campylobacter humoral response by a bacterin and the lack of protection against homologous challenge in Japanese Jidori chickens. J. Vet. Med. Sci. 2012, 74, 75–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, B.E.; Rollins, D.M.; Mallinson, E.T.; Carr, L.; Joseph, S.W. Campylobacter jejuni in broiler chickens: Colonization and humoral immunity following oral vaccination and experimental infection. Vaccine 1997, 15, 1922–1932. [Google Scholar] [CrossRef]
- Sizemore, D.R.; Warner, B.; Lawrence, J.; Jones, A.; Killeen, K.P. Live, attenuated Salmonella typhimurium vectoring Campylobacter antigens. Vaccine 2006, 24, 3793–3803. [Google Scholar] [CrossRef] [PubMed]
- Layton, S.L.; Morgan, M.J.; Cole, K.; Kwon, Y.M.; Donoghue, D.J.; Hargis, B.M.; Pumford, N.R. Evaluation of Salmonella-vectored Campylobacter peptide epitopes for reduction of Campylobacter jejuni in broiler chickens. Clin. Vaccine Immunol. 2011, 18, 449–454. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buckley, A.M.; Wang, J.; Hudson, D.L.; Grant, A.J.; Jones, M.A.; Maskell, D.J.; Stevens, M.P. Evaluation of live-attenuated Salmonella vaccines expressing Campylobacter antigens for control of C. jejuni in poultry. Vaccine 2010, 28, 1094–1105. [Google Scholar] [CrossRef] [PubMed]
- Alemka, A.; Nothaft, H.; Zheng, J.; Szymanski, C.M. N-glycosylation of Campylobacter jejuni surface proteins promotes bacterial fitness. Infect. Immun. 2013, 81, 1674–1682. [Google Scholar] [CrossRef] [Green Version]
- van Sorge, N.M.; Bleumink, N.M.; van Vliet, S.J.; Saeland, E.; van der Pol, W.L.; van Kooyk, Y.; van Putten, J.P. N-glycosylated proteins and distinct lipooligosaccharide glycoforms of Campylobacter jejuni target the human C-type lectin receptor MGL. Cell Microbiol. 2009, 11, 1768–1781. [Google Scholar] [CrossRef] [PubMed]
- Nothaft, H.; Davis, B.; Lock, Y.Y.; Perez-Munoz, M.E.; Vinogradov, E.; Walter, J.; Coros, C.; Szymanski, C.M. Engineering the Campylobacter jejuni N-glycan to create an effective chicken vaccine. Sci. Rep. 2016, 6, 26511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battersby, T.; Whyte, P.; Bolton, D.J. The pattern of Campylobacter contamination on broiler farms; external and internal sources. J. Appl. Microbiol. 2016, 120, 1108–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, O.; Luo, N.; Huang, S.; Zhang, Q. Effect of Campylobacter-specific maternal antibodies on Campylobacter jejuni colonization in young chickens. Appl. Environ. Microbiol. 2003, 69, 5372–5379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahin, O.; Zhang, Q.; Meitzler, J.C.; Harr, B.S.; Morishita, T.Y.; Mohan, R. Prevalence, antigenic specificity, and bactericidal activity of poultry anti-Campylobacter maternal antibodies. Appl. Environ. Microbiol. 2001, 67, 3951–3957. [Google Scholar] [CrossRef] [Green Version]
- Mortada, M.; Cosby, D.E.; Akerele, G.; Ramadan, N.; Oxford, J.; Shanmugasundaram, R.; Ng, T.T.; Selvaraj, R.K. Characterizing the immune response of chickens to Campylobacter jejuni (Strain A74C). PLoS ONE 2021, 16, e0247080. [Google Scholar] [CrossRef] [PubMed]
- Connerton, P.L.; Richards, P.J.; Lafontaine, G.M.; O’Kane, P.M.; Ghaffar, N.; Cummings, N.J.; Smith, D.L.; Fish, N.M.; Connerton, I.F. The effect of the timing of exposure to Campylobacter jejuni on the gut microbiome and inflammatory responses of broiler chickens. Microbiome 2018, 6, 88. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.K.; Abuoun, M.; Cawthraw, S.A.; Humphrey, T.J.; Rothwell, L.; Kaiser, P.; Barrow, P.A.; Jones, M.A. Campylobacter colonization of the chicken induces a proinflammatory response in mucosal tissues. FEMS Microbiol. Immunol. 2008, 54, 114–121. [Google Scholar] [CrossRef] [Green Version]
- Lacharme-Lora, L.; Chaloner, G.; Gilroy, R.; Humphrey, S.; Gibbs, K.; Jopson, S.; Wright, E.; Reid, W.; Ketley, J.; Humphrey, T.; et al. B lymphocytes play a limited role in clearance of Campylobacter jejuni from the chicken intestinal tract. Sci. Rep. 2017, 7, 45090. [Google Scholar] [CrossRef]
- Meunier, M.; Guyard-Nicodème, M.; Dory, D.; Chemaly, M. Control strategies against Campylobacter at the poultry production level: Biosecurity measures, feed additives and vaccination. J. Appl. Microbiol. 2016, 120, 1139–1173. [Google Scholar] [CrossRef] [Green Version]
- Hendrixson, D.R.; Akerley, B.J.; DiRita, V.J. Transposon mutagenesis of Campylobacter jejuni identifies a bipartite energy taxis system required for motility. Mol. Microbiol. 2001, 40, 214–224. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day | 0 d | 21 d | 42 d | |
|---|---|---|---|---|
| 1st Experiment | Control (PBS; n = 11) | 47.06 ± 2.92 g | 890.36 ± 70.29 g | 2114.55 ± 204.76 g |
| Control (WT; n = 11) | 44.09 ± 3.40 g | 981.00 ± 104.05 g | 2101.50 ± 363.49 g | |
| Treated group (ΔahpC + WT; n = 11) 1 | 45.91 ± 3.02 g | 993.18 ± 91.17 g | 2289.55 ± 246.95 g | |
| 2nd Experiment | Control (PBS; n = 11) | 26.27 ± 4.45 g | 980.00 ± 27.93 g | 2400.00 ± 163.30 g |
| Control (WT; n = 11) | 38.64 ± 6.36 g | 940.00 ± 84.38 g | 2150.00 ± 190.03 g | |
| Treated group (ΔahpC + WT; n = 11) 2 | 42.91 ± 6.01 g | 910.91 ± 126.29 g | 2218.18 ± 348.76 g | |
| Treated group (ΔahpC + WT; n = 11) 3 | 39.55 ± 4.72 g | 906.36 ± 60.71 g | 2245.45 ± 136.85 g | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, B.; Saisom, T.; Sasipreeyajan, J.; Luangtongkum, T. Live-Attenuated Oral Vaccines to Reduce Campylobacter Colonization in Poultry. Vaccines 2022, 10, 685. https://doi.org/10.3390/vaccines10050685
Jeon B, Saisom T, Sasipreeyajan J, Luangtongkum T. Live-Attenuated Oral Vaccines to Reduce Campylobacter Colonization in Poultry. Vaccines. 2022; 10(5):685. https://doi.org/10.3390/vaccines10050685
Chicago/Turabian StyleJeon, Byeonghwa, Tunchanok Saisom, Jiroj Sasipreeyajan, and Taradon Luangtongkum. 2022. "Live-Attenuated Oral Vaccines to Reduce Campylobacter Colonization in Poultry" Vaccines 10, no. 5: 685. https://doi.org/10.3390/vaccines10050685