
Mycobacterium bovis Wild-Type BCG or Recombinant BCG Secreting Murine IL-18 (rBCG/IL-18) Strains in Driving Immune Responses in Immunocompetent or Immunosuppressed Mice

 , , ,
, , ,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Bacteria Growth Conditions
2.3. Infection and Immunosuppression of Mice
2.4. Isolation of Bone Marrow Cells and Cytospin Preparation
2.5. Fluorescent-Antibody Staining of Bone Marrow Cells
2.6. Cytokine Measurement
2.7. Isolation of Alveolar Macrophages and Assessment of Phagocytosis
2.8. Staining of Splenocytes and Flow Cytometry for the Assessment of Central Memory T Cells (TCM) and Effector Memory T Cells (TEM)
2.9. Statistical Analysis
3. Results
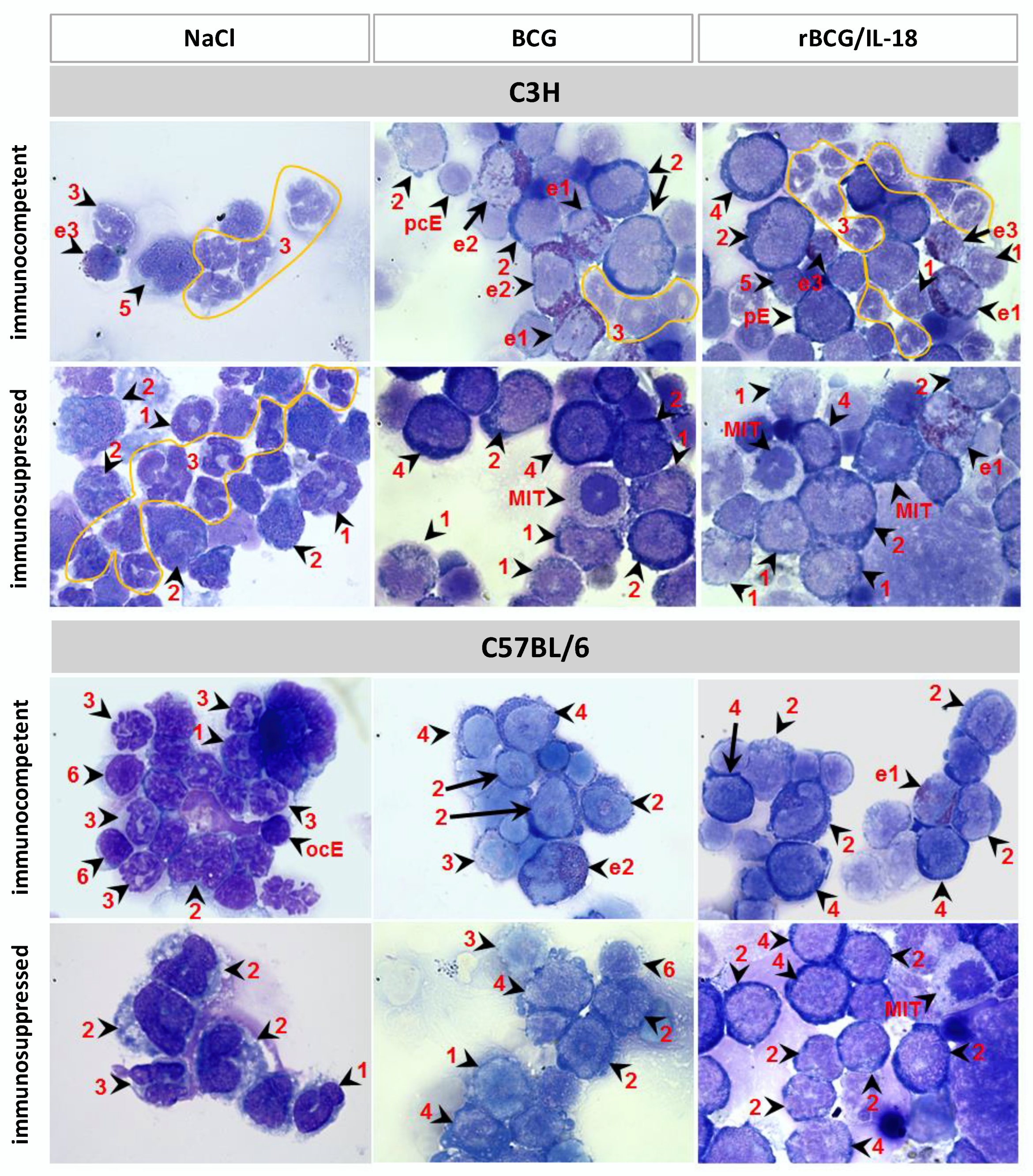
3.1. Composition of Leukocytes in Cytospins from Bone Marrow Cells
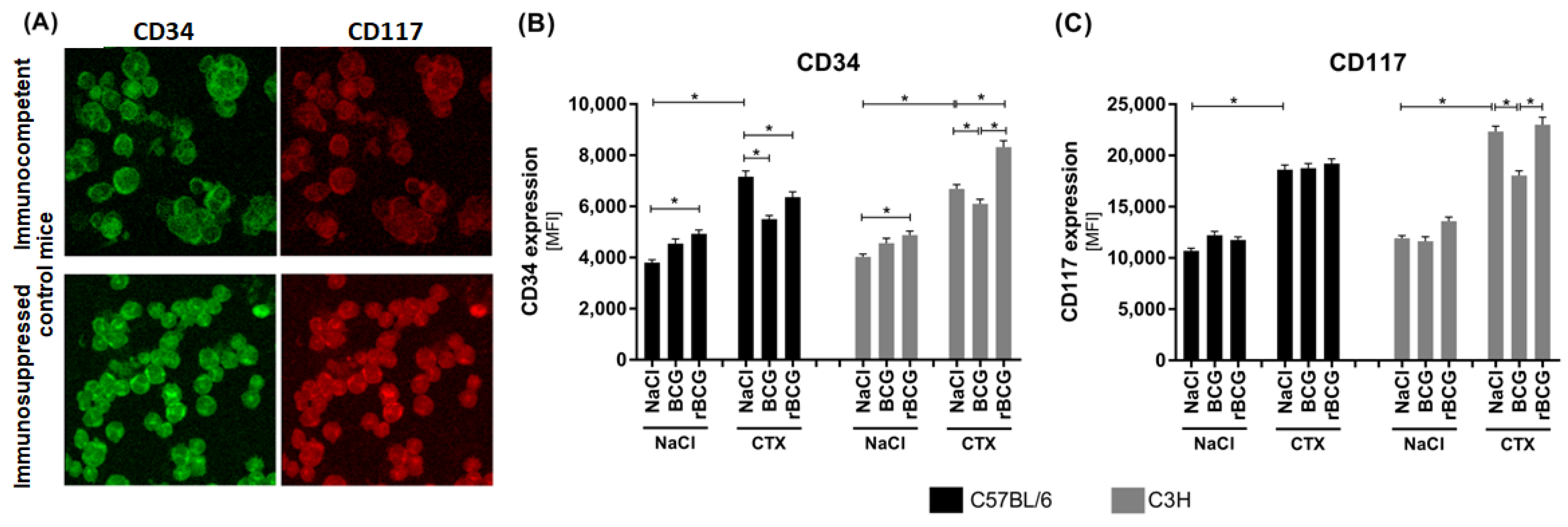
3.2. CD34 and CD117 Expression on Bone Marrow Cells
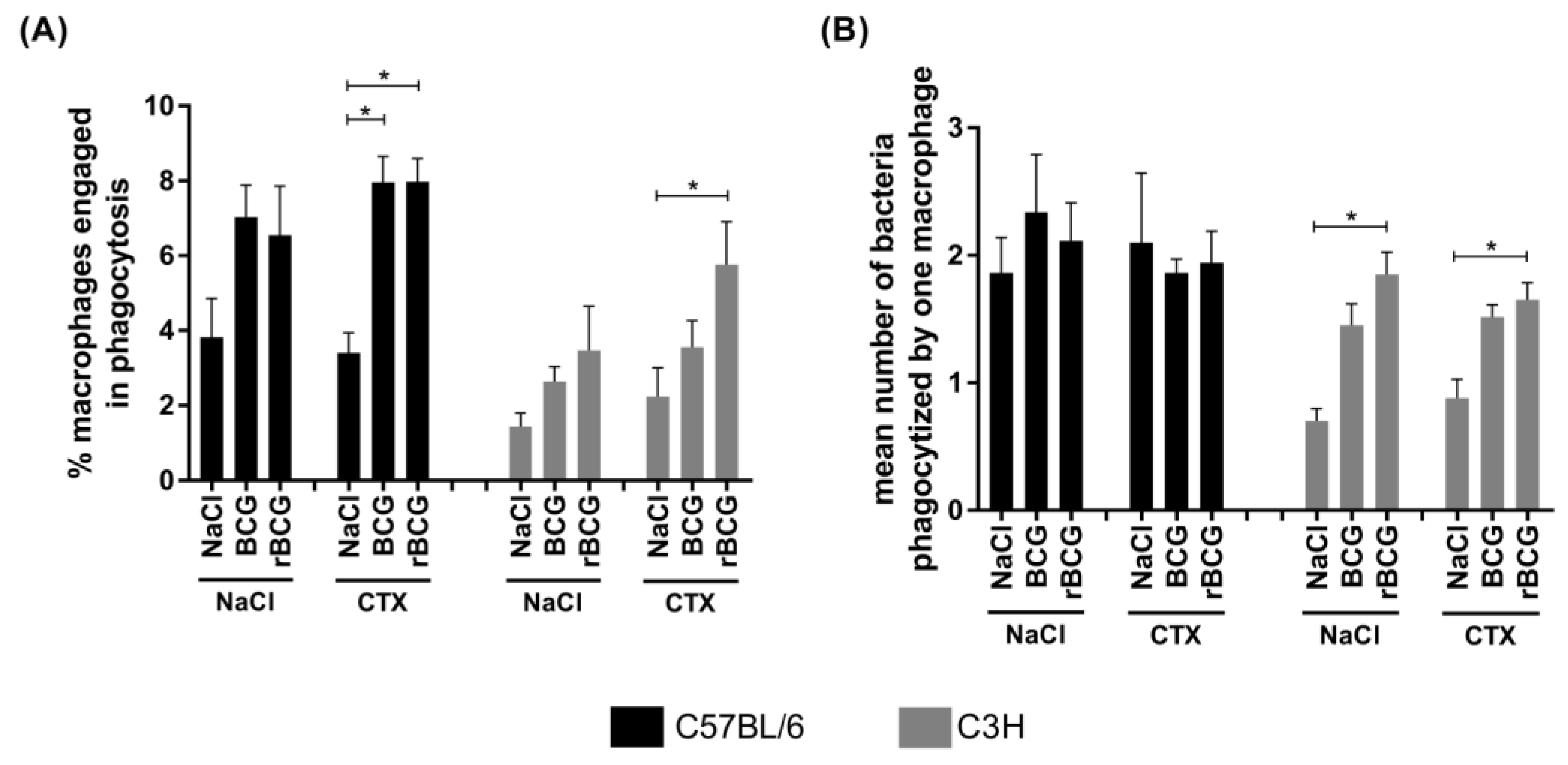
3.3. Phagocytic Activity of Alveolar Macrophages
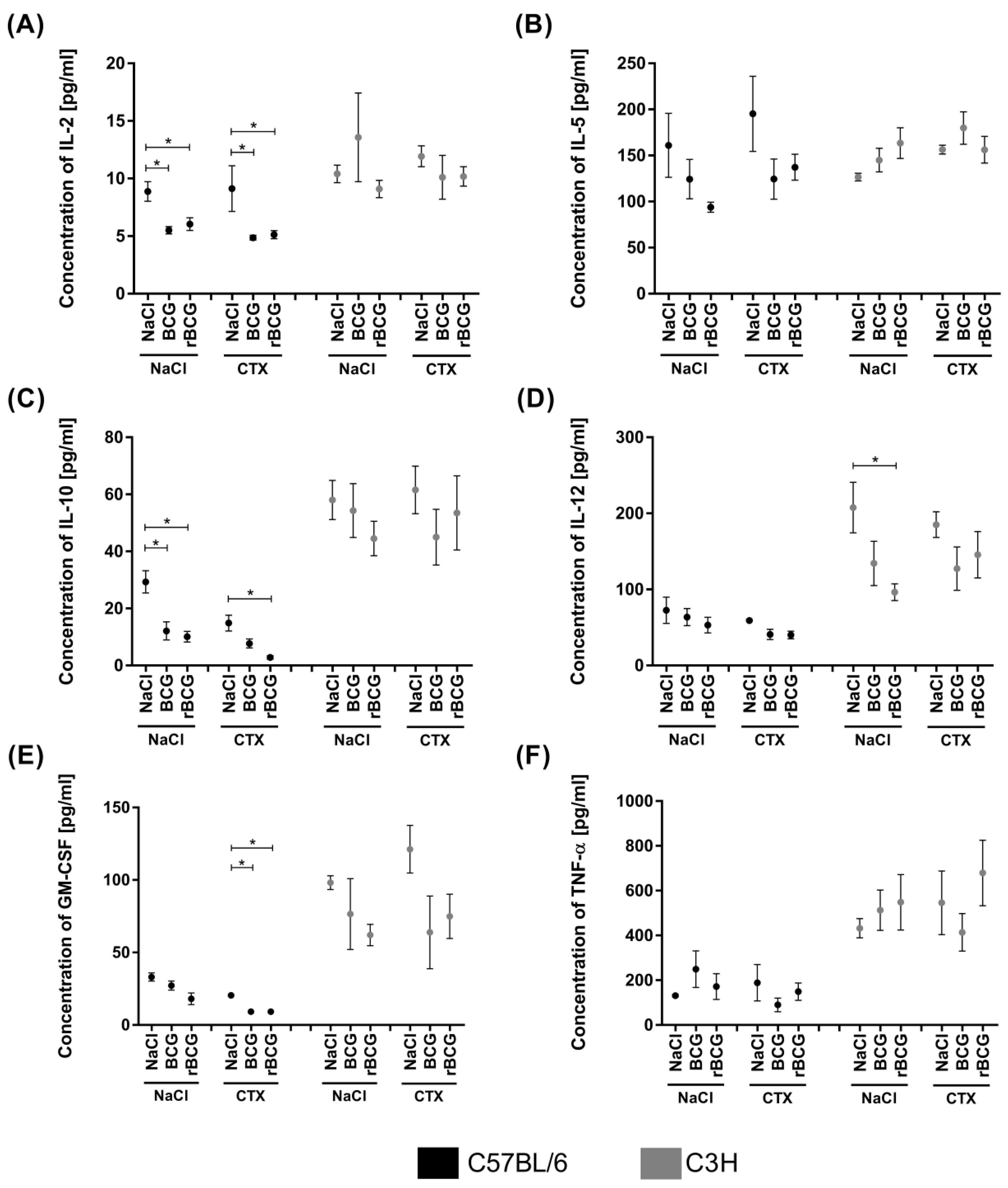
3.4. Serum Cytokine Concentration
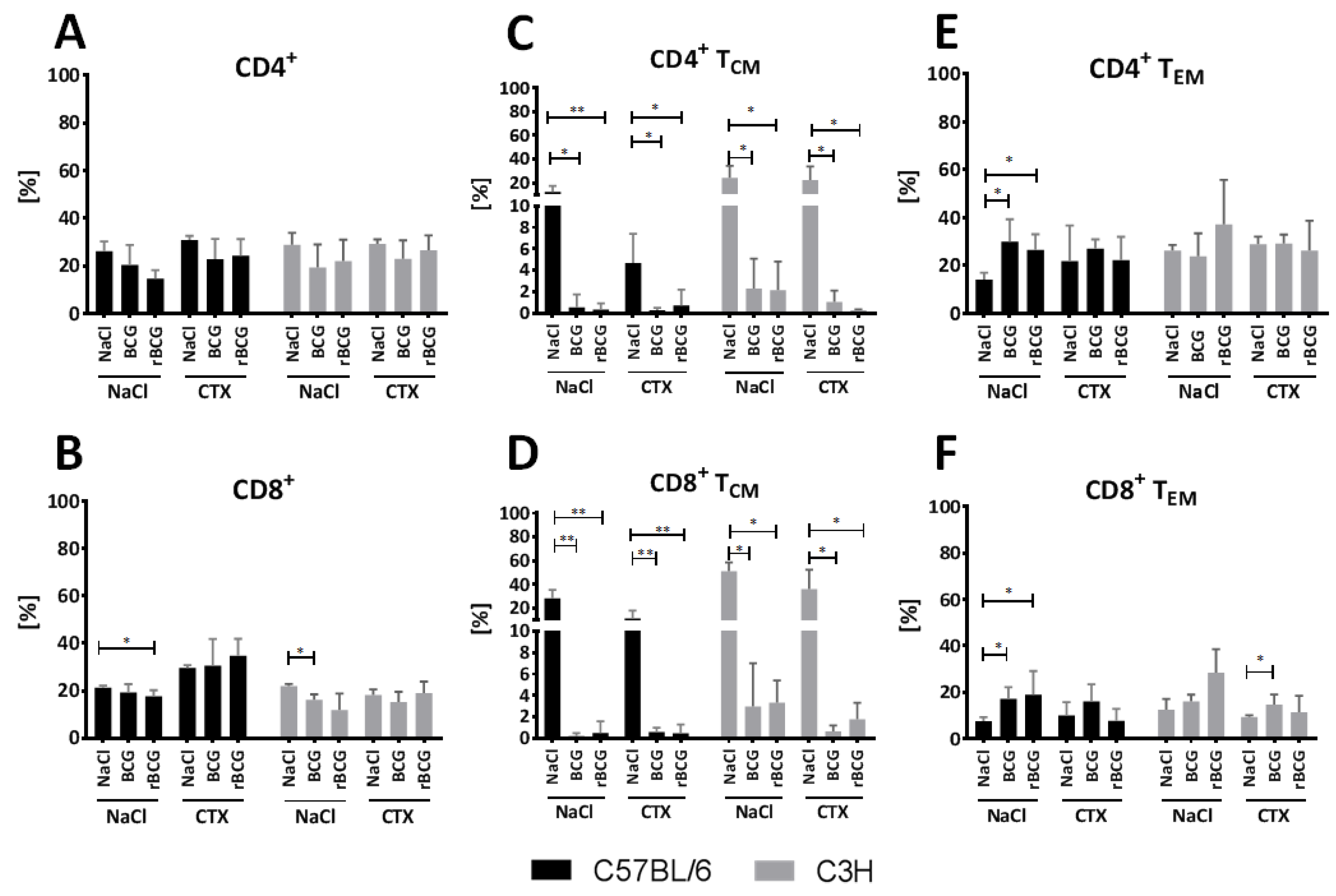
3.5. Effect of BCG and rBCG on the CD4+ and CD8+ TCM and TEM Lymphocytes Response
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Global Tuberculosis Report WHO 2021. Available online: https://www.who.int/teams/global-tuberculosis-programme/tb-reports (accessed on 17 February 2022).
- Houben, R.M.; Dodd, P.J. The global burden of latent tuberculosis infection: A re-estimation using mathematical modelling. PLoS Med. 2016, 13, e1002152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiazyk, S.; Ball, T.B. Latent tuberculosis infection: An overview. Can. Commun. Dis. Rep. 2017, 43, 62–66. [Google Scholar] [CrossRef] [PubMed]
- Ai, J.W.; Ruan, Q.L.; Liu, Q.H.; Zhang, W.H. Updates on the risk factors for latent tuberculosis reactivation and their managements. Emerg. Microbes Infect. 2016, 5, e10. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, L.; Stagg, H.R.; Abubakar, I. Diagnosis and management of latent tuberculosis infection. Cold Spring Harb. Perspect. Med. 2015, 5, a017830. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaufmann, S.H.; Hussey, G.; Lambert, P.H. New vaccines for tuberculosis. Lancet 2010, 375, 2110–2119. [Google Scholar] [CrossRef]
- Matsuo, K.; Yasutomi, Y. Mycobacterium bovis Bacille Calmette-Guérin as a vaccine vector for global infectious disease control. Tuberc. Res. Treat. 2011, 2011, 574591. [Google Scholar]
- Covián, C.; Fernández-Fierro, A.; Retamal-Díaz, A.; Díaz, F.E.; Vasquez, A.E.; Lay, M.K.; Riedel, C.A.; González, P.A.; Bueno, S.M.; Kalergis, A.M. BCG-induced cross-protection and development of trained immunity: Implication for vaccine design. Front. Immunol. 2019, 10, 2806. [Google Scholar] [CrossRef]
- Biet, F.; Kremer, L.; Wolowczuk, I.; Delacre, M.; Locht, C. Mycobacterium bovis BCG producing interleukin-18 increases antigen-specific gamma interferon production in mice. Infect. Immun. 2002, 70, 6549–6557. [Google Scholar] [CrossRef] [Green Version]
- Biet, F.; Duez, C.; Kremer, L.; Marquillies, P.; Amniai, L.; Tonnel, A.B.; Locht, C.; Pestel, J. Recombinant Mycobacterium bovis BCG producing IL-18 reduces IL-5 production and bronchoalveolar eosinophilia induced by an allergic reaction. Allergy 2005, 60, 1065–1072. [Google Scholar] [CrossRef]
- Gracie, J.A.; Robertson, S.E.; McInnes, I.B. Interleukin-18. J. Leukoc. Biol. 2003, 73, 213–224. [Google Scholar] [CrossRef]
- Dinarello, C.A.; Novick, D.; Kim, S.; Kaplanski, G. Interleukin-18 and IL-18 binding protein. Front. Immunol. 2013, 4, 289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swain, S.L. Interleukin 18: Tipping the balance towards a T helper cell 1 response. J. Exp. Med. 2001, 194, F11–F14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaplanski, G. Interleukin-18: Biological properties and role in disease pathogenesis. Immunol. Rev. 2018, 281, 138–153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Li, X.; Zhang, C.; Wu, T.; Li, Y.; Cheng, X. A eukaryotic expression plasmid carrying chicken interleukin-18 enhances the response to Newcastle disease virus vaccine. Clin. Vaccine Immunol. 2015, 22, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Wawrocki, S.; Druszczynska, M.; Kowalewicz-Kulbat, M.; Rudnicka, W. Interleukin 18 (IL-18) as a target for immune intervention. Acta Biochim. Pol. 2016, 63, 59–63. [Google Scholar] [CrossRef] [Green Version]
- Kinoshita, M.; Miyazaki, H.; Ono, S.; Seki, S. Immunoenhancing therapy with interleukin-18 against bacterial infection in immunocompromised hosts after severe surgical stress. J. Leukoc. Biol. 2013, 93, 689–698. [Google Scholar] [CrossRef]
- Pelletier, M.; Forget, A.; Bourassa, D.; Gros, P.; Skamene, E. Immunopathology of BCG infection in genetically resistant and susceptible mouse strains. J. Immunol. 1982, 129, 2179–2185. [Google Scholar]
- Mortatti, R.C.; Maia, L.C. Immune response to BCG-Moreau (Rio de Janeiro) strain. Spectrum of delayed hypersensitivity in genetically defined mice. FEMS Microbiol. Immunol. 1989, 1, 491–497. [Google Scholar] [CrossRef]
- Colvin, O.M. An overview of cyclophosphamide development and clinical applications. Curr. Pharm. Des. 1999, 5, 555–560. [Google Scholar]
- Ahlmann, M.; Hempel, G. The effect of cyclophosphamide on the immune system: Implications for clinical cancer therapy. Cancer Chemother. Pharmacol. 2016, 78, 661–671. [Google Scholar] [CrossRef]
- Neri, P.; Bahlis, N.J.; Lonial, S. New strategies in multiple myeloma: Immunotherapy as a novel approach to treat patients with multiple myeloma. Clin. Cancer Res. 2016, 22, 5959–5965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Włodarczyk, M.; Ograczyk, E.; Kowalewicz-Kulbat, M.; Druszczyńska, M.; Rudnicka, W.; Fol, M. Effect of cyclophosphamide treatment on central and effector memory T cells in mice. Int. J. Toxicol. 2018, 37, 373–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Quan, N. Immune cell isolation from mouse femur bone marrow. Bio-protocol 2015, 5, e1631. [Google Scholar] [CrossRef] [PubMed]
- Löffler, H.; Rastetter, J.; Haferlach, T. Atlas of Clinical Hematology, 6th ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2005; p. 8. [Google Scholar]
- Drew, E.; Merzaban, J.S.; Seo, W.; Ziltener, H.J.; McNagny, K.M. CD34 and CD43 inhibit mast cell adhesion and are required for optimal mast cell reconstitution. Immunity 2005, 22, 43–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saleh, M.; Shamsasanian, K.; Movassaghpourakbari, A.; Akbarzadehlaleh, P.; Molaeipour, Z. The impact of mesenchymal stem cells on differentiation of hematopoietic stem cells. Adv. Pharm. Bull. 2015, 5, 299. [Google Scholar] [CrossRef] [Green Version]
- Sperling, C.; Schwartz, S.; Buchner, T.; Thiel, E.; Ludwig, W.D. Expression of the stem cell factor receptor C-KIT (CD117) in acute leukemias. Haematologica 1997, 82, 617–621. [Google Scholar]
- Feng, L.; Huang, Q.; Huang, Z.; Li, H.; Qi, X.; Wang, Y.; Liu, Z.; Liu, X.; Lu, L. Optimized animal model of cyclophosphamide -induced bone marrow suppression. Basic Clin. Pharmacol. Toxicol. 2016, 119, 428–435. [Google Scholar] [CrossRef]
- Ogura, T.; Ueda, H.; Hosohara, K.; Tsuii, R.; Nagata, Y.; Kashiwamura, S.; Okamura, H. Interleukin 18 stimulates hematopoietic cytokine and growth factor formation and augments circulating granulocytes in mice. Blood 2001, 98, 2101–2107. [Google Scholar] [CrossRef] [Green Version]
- Deng, Y.; Ye, J.; Luo, Q.; Huang, Z.; Peng, Y.; Xiong, G.; Guo, Y.; Jiang, H.; Li, J. Low-density granulocytes are elevated in mycobacterial infection and associated with the severity of tuberculosis. PLoS ONE 2016, 11, e0153567. [Google Scholar] [CrossRef]
- Fackler, M.J.; Krause, D.S.; Smith, O.M.; Civin, C.I.; May, W.S. Full-length but not truncated CD34 inhibits hematopoietic cell differentiation of M1 cells. Blood 1995, 85, 3040–3047. [Google Scholar] [CrossRef] [Green Version]
- McGarry, M.P.; Protheroe, C.A.; Lee, J.J. Mouse hematology: A laboratory manual, Har/DVD La Edition; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2009; pp. 1–99. [Google Scholar]
- Shihan, M.H.; Novo, S.G.; Le Marchand, S.J.; Wang, Y.; Duncan, M.K. A simple method for quantifying confocal fluorescent images. Biochem. Biophys. Rep. 2021, 25, 100916. [Google Scholar]
- Liu, C.; Huang, M.; Liu, F.; Xu, X.; Feng, W.; Han, G.; Liu, X.; Geng, L.; Fu, T. The role of surgical management of BCG vaccine-induced regional suppurative lymphadenitis in children: A 7 years’ experience from one medical center. BMC Infect. Dis. 2021, 21, 801. [Google Scholar] [CrossRef] [PubMed]
- Cirovic, B.; de Bree, L.C.J.; Groh, L.; Blok, B.A.; Chan, J.; van der Velden, W.J.F.M.; Bremmers, M.E.J.; van Creve, R.; Handler, K.; Picelli, S.; et al. BCG vaccination in humans elicits trained immunity via the hematopoietic progenitor compartment. Cell Host Microbe 2020, 28, 322–334. [Google Scholar] [CrossRef] [PubMed]
- Huyan, X.H.; Lin, Y.P.; Gao, T.; Chen, R.Y.; Fan, Y.M. Immunosuppressive effect of cyclophosphamide on white blood cells and lymphocyte subpopulations from peripheral blood of Balb/c mice. Int. Immunopharmacol. 2011, 11, 1293–1297. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Waxman, D.J. Immunogenic chemotherapy: Dose and schedule dependence and combination with immunotherapy. Cancer Lett. 2018, 419, 210–221. [Google Scholar] [CrossRef]
- Buisman, A.M.; Van Zwet, T.L.; Langermans, J.A.M.; Geertsma, M.F.; Leenen, P.J.M.; Van Furth, R. Different effect of granulocyte colony-stimulating factor or bacterial infection on bone-marrow cells of cyclophosphamide-treated or irradiated mice. Immunology 1999, 97, 601–610. [Google Scholar] [CrossRef]
- Roberst, A.W.; Metcalf, D. Noncycling state of peripheral blood progenitor cells mobilized by granulocyte colony-stimulating factor and other cytokines. Blood 1995, 86, 1600–1605. [Google Scholar]
- Croockewit, A.J.; Raymakers, R.A.P.; Smeets, M.E.P.; vd Bosch, G.; Pennings, A.H.M.; de Witte, T.J.M. The low cycling status of mobilized peripheral blood CD34+ cells is not restricted to the more primitive subfraction. Leukemia 1998, 12, 571–577. [Google Scholar] [CrossRef] [Green Version]
- Miettinen, M.; Lasota, J. KIT (CD117): A review on expression in normal and neoplastic tissues, and mutations and their clinicopathologic correlation. Appl. Immunohistochem. Mol. Morphol. 2005, 13, 205–220. [Google Scholar] [CrossRef]
- Sitsigu, A.; Viaud, S.; Chaput, N.; Bracci, L.; Proietti, E.; Zitvogel, L. Immunomodulatory effects of cyclophosphamide and implementations for vaccine design. Semin. Immunopathol. 2011, 33, 369–383. [Google Scholar]
- Lis, M.; Obmińska-Mrukowicz, B. Effects of bestatin on phagocytic cells in cyclophosphamide-treated mice. Pharmacol. Rep. 2011, 63, 1481–1490. [Google Scholar] [CrossRef]
- Kinoshita, M.; Seki, S.; Ono, S.; Shinomiya, N.; Hiraide, H. Paradoxical effect of IL-18 therapy on the severe and mild Escherichia coli infections in burn-injured mice. Ann. Surg. 2004, 240, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, M.; Ceballos-Olvera, I.; del Barrio, L.; Re, F. Role of inflammasome, IL-1β and IL-18 in bacterial infections. Sci. World J. 2011, 11, 2037–2050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Steenwinkel, J.E.; de Knegt, G.J.; ten Kate, M.T.; Verbrugh, H.A.; Ottenhoff, T.H.; Bakker-Wounderberg, I.A. Dynamics of interferon gamma release assay and cytokine profiles in blood and respiratory tract specimens from mice with tuberculosis and the effect of therapy. Eur. J. Clin. Microbiol. Infect. Dis. 2012, 6, 1195–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shafiani, S.; Tucker-Heard, G.; Kariyone, A.; Takatsu, K.; Urdahl, K.B. Pathogen-specific regulatory T cells delay the arrival of effector T cells in the lung during early tuberculosis. J. Exp. Med. 2010, 207, 1409–1420. [Google Scholar] [CrossRef] [Green Version]
- Olobo, J.O.; Geletu, M.; Demissie, A.; Eguale, T.; Hiwot, K.; Aderaye, G.; Britton, S. Circulating TNF-α, TGF-β, and IL-10 in tuberculosis patients and healthy contacts. Scand. J. Immunol. 2001, 53, 85–91. [Google Scholar] [CrossRef]
- Liu, X.; Li, F.; Niu, H.; Ma, L.; Chen, J.; Zhang, Y.; Peng, L.; Gan, C.; Ma, X.; Zhu, B. IL-2 restores T-cell dysfunction induced by persistent Mycobacterium tuberculosis antigen stimulation. Front. Immunol. 2019, 10, 2350. [Google Scholar] [CrossRef]
- Inoue, Y.; Abe, K.; Onozaki, K.; Hayashi, H. TGF-β decreases the stability of IL-18—Induced IFN-gamma mRNA through the expression of TGF-β induced tristetraptolin in KG-1 cells. Biol. Pharm. Bull. 2015, 38, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Luo, Y.; Yamada, H.; Chen, X.; Ryan, A.A.; Evatoff, D.P.; Triccas, J.A.; O’Donnel, M.A. Recombinant Mycobacterium bovis bacillus Calmette-Guérin (BCG) expressing mouse IL-18 augments Th1 immunity and macrophage cytotoxicity. Clin. Exp. Immunol. 2004, 137, 24–34. [Google Scholar] [CrossRef]
- Schultz, M.J.; Knapps, S.; Florguin, S.; Pater, J.; Takeda, K.; Akira, S.; van der Poll, T. Interleukin-18 impairs the pulmonary host response to Pseudomonas aeruginosa. Infect. Immun. 2003, 71, 1630–1634. [Google Scholar] [CrossRef] [Green Version]
- Ghose, P.; Ali, A.Q.; Fang, R.; Forbes, D.; Ballard, B.; Ismaiul, N. The interaction between IL-18 and IL-18 receptor limits the magnitude of protective immunity and enhances pathogenic responses following infection with intracellular bacteria. J. Immunol. 2011, 187, 1333–1346. [Google Scholar] [CrossRef] [PubMed]
- Vahedi, G.C.; Poholek, A.; Hand, T.W.; Laurence, A.; Kanno, Y.; O’Shea, J.J.; Hirahara, K. Helper T-cell identity and evolution of differential transcriptomes and epigenomes. Immunol. Rev. 2013, 252, 24–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vahedi, G.; Kanno, Y.; Sartorelli, V.; O’Shea, J.J. Transcription factors and CD4 T cells seeking identity: Masters, minions, setters and spikers. Immunology 2013, 139, 294–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yagi, J.; Arimura, Y.; Takatori, H.; Nakajima, H.; Iwamoto, I.; Uchiyama, T. Genetic background influences Th cell differentiation by controlling the capacity for IL-2-induced IL-4 production by naive CD4+ T cells. Int. Immunol. 2006, 18, 1681–1690. [Google Scholar] [CrossRef] [Green Version]
- Garcia-Pelayo, M.C.; Bachy, V.S.; Kaveh, D.A.; Hogarth, P.J. BALB/c mice display more enhanced BCG vaccine induced Th1 and Th17 response than C57BL/6 mice but have equivalent protection. Tuberculosis 2015, 95, 48–53. [Google Scholar] [CrossRef] [Green Version]
- Kremer, L.; Estaquier, J.; Wolowczuk, I.; Biet, F.; Ameisen, J.C.; Locht, C. Ineffective cellular immune response associated with t-cell apoptosis in susceptible Mycobacterium bovis BCG-infected mice. Infect. Immun. 2000, 68, 4264–4273. [Google Scholar] [CrossRef] [Green Version]
- Gourley, T.S.; Wherry, E.J.; Masopust, D.; Ahmed, R. Generation and maintenance of immunological memory. Semin. Immunol. 2004, 16, 323–333. [Google Scholar] [CrossRef]
- McKinstry, K.K.; Strutt, T.M.; Swain, S.L. The effector to memory transition of CD4 T cells. Immunol. Res. 2008, 40, 114–127. [Google Scholar] [CrossRef]
- Lanzavecchia, A.; Sallusto, F. Understanding the generation and function of memory T cell subsets. Curr. Opin. Immunol. 2005, 17, 326–332. [Google Scholar] [CrossRef]
- Sallusto, F.; Geginat, J.; Lanzavecchia, A. Central memory and effector memory T cell subsets: Function, generation, and maintenance. Annu. Rev. Immunol. 2004, 22, 745–763. [Google Scholar] [CrossRef]
- Reinhardt, R.L.; Khoruts, A.; Merica, R.; Zell, T.; Jenkins, M.K. Visualizing the generation of memory CD4 T cells in the whole body. Nature 2001, 410, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Masopust, D.; Vezys, V.; Marzo, A.L.; Lefrancois, L. Preferential localization of effector memory cells in nonlymphoid tissue. Science 2001, 291, 2413–2417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henao-Tamayo, M.I.; Ordway, D.J.; Irwin, S.M.; Shang, S.; Shanley, C.; Orme, I.M. Phenotypic definition of effector and memory T-lymphocyte subsets in mice chronically infected with Mycobacterium tuberculosis. Clin. Vaccine Immunol. 2010, 17, 618–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| C57BL/6 Mice | C3H Mice | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Immunocompetent | Immunosuppressed | Immunocompetent | Immunosuppressed | ||||||||||
| Nacl | BCG | rBCG/IL-18 | Nacl | BCG | rBCG/IL-18 | Nacl | BCG | rBCG/IL-18 | Nacl | BCG | rBCG/IL-18 | ||
| Granulocytes | Myeloblast | 3.0 ± 0.6 | 10.4 ± 0.5 | 5.7 ± 1.7 | 5.7 ± 1.8 | 10.2 ± 2.6 | 18.6 ± 0.6 | 1.1 ± 1.1 | 4.8 ± 0.9 | 5.5 ± 1.4 | 6.8 ± 1.3 | 25.5 ± 4.6 | 18.4 ± 1.5 |
| Promyelocyte | 11.4 ± 5.3 | 12.4 ± 2.7 | 16.0 ± 4.1 | 15.7 ± 1.9 | 43.8 ± 1.4 | 42.4 ± 0.7 | 7.7 ± 1.0 | 18.6 ± 0.7 | 12.4 ± 2.7 | 29.2 ± 8.6 | 43.9 ± 7.5 | 44.9 ± 3.5 | |
| Myelocyte | 13.6 ± 1.8 | 7.7 ± 0.3 | 11.4 ± 1.8 | 17.2 ± 2.4 | 14.1 ± 3.1 | 19.0 ± 0.3 | 8.2 ± 0.9 | 10.9 ± 1.8 | 12.6 ± 2.6 | 12.2 ± 4.0 | 14.1 ± 7.0 | 19.0 ± 1.7 | |
| Metamyelocyte | 38.7 ± 5.5 | 39.6 ± 2.5 | 45.7 ± 9.9 | 59.0 ± 2.9 | 12.9 ± 6.1 | 10.4 ± 0.7 | 61.6 ± 4.5 | 41.5 ± 1.5 | 44.9 ± 2.9 | 43.0 ± 15.9 | 4.0 ± 2.7 | 10.1 ± 1.6 | |
| ∑ | 66.7 | 70.1 | 78.8 | 97.6 | 81.0 | 90.4 | 78.6 | 75.8 | 75.4 | 91.2 | 87.5 | 92.4 | |
| Lymphocytes | 5.7 | 3.2 | 3.5 | 0.0 | 1.6 | 0.0 | 0.0 | 2.5 | 2.6 | 2.0 | 0.9 | 0.0 | |
| Monocytes | 0.9 | 0.0 | 1.0 | 0.0 | 0.0 | 0.1 | 1.3 | 0.0 | 0.3 | 0.0 | 0.0 | 0.0 | |
| ∑ | 73.3 | 73.3 | 83.3 | 97.6 | 82.6 | 90.5 | 79.9 | 78.3 | 78.0 | 93.2 | 88.4 | 92.4 | |
| C57BL/6 mice | C3H mice | ||||||||||||
| Myeloblast | NaCl/CTX vs. rBCG/CTX p = 0.001 | NaCl/CTX vs. BCG/CTX p = 0.002 | |||||||||||
| Promyelocyte | NaCl/CTX vs. BCG/CTX p = 0.003 | NaCl/CTX vs. rBCG/CTX p = 0.04 | |||||||||||
| NaCl/CTX vs. rBCG/CTX p = 0.02 | |||||||||||||
| Metamyelocyte | NaCl/CTX vs. BCG/CTX p = 0.02 | NaCl/CTX vs. BCG/CTX p = 0.002 | |||||||||||
| NaCl/CTX vs. rBCG/CTX p = 0.003 | |||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fol, M.; Włodarczyk, M.; Kowalewicz-Kulbat, M.; Druszczyńska, M.; Krawczyk, K.T.; Wawrocki, S.; Rudnicka, W.; Chmiela, M. Mycobacterium bovis Wild-Type BCG or Recombinant BCG Secreting Murine IL-18 (rBCG/IL-18) Strains in Driving Immune Responses in Immunocompetent or Immunosuppressed Mice. Vaccines 2022, 10, 615. https://doi.org/10.3390/vaccines10040615
Fol M, Włodarczyk M, Kowalewicz-Kulbat M, Druszczyńska M, Krawczyk KT, Wawrocki S, Rudnicka W, Chmiela M. Mycobacterium bovis Wild-Type BCG or Recombinant BCG Secreting Murine IL-18 (rBCG/IL-18) Strains in Driving Immune Responses in Immunocompetent or Immunosuppressed Mice. Vaccines. 2022; 10(4):615. https://doi.org/10.3390/vaccines10040615
Chicago/Turabian StyleFol, Marek, Marcin Włodarczyk, Magdalena Kowalewicz-Kulbat, Magdalena Druszczyńska, Krzysztof T. Krawczyk, Sebastian Wawrocki, Wiesława Rudnicka, and Magdalena Chmiela. 2022. "Mycobacterium bovis Wild-Type BCG or Recombinant BCG Secreting Murine IL-18 (rBCG/IL-18) Strains in Driving Immune Responses in Immunocompetent or Immunosuppressed Mice" Vaccines 10, no. 4: 615. https://doi.org/10.3390/vaccines10040615