
Dermal Delivery of a SARS-CoV-2 Subunit Vaccine Induces Immunogenicity against Variants of Concern

 ,
,  , , , , , , and
, , , , , , and 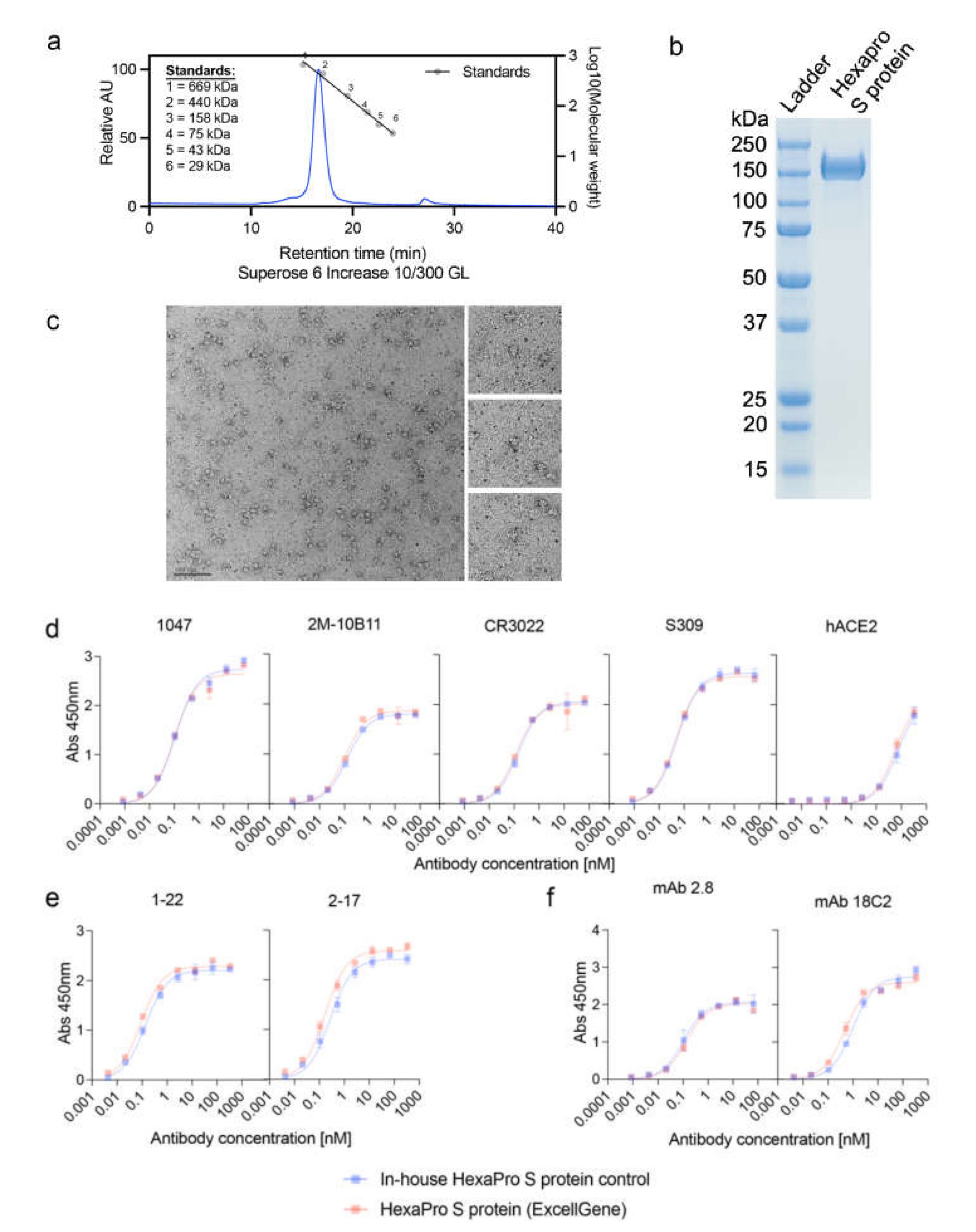{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. SARS CoV-2 HexaPro S Protein
2.2. SARS CoV-2 HexaPro S Protein Characterization
2.3. Transmission Electron Microscopy (TEM)
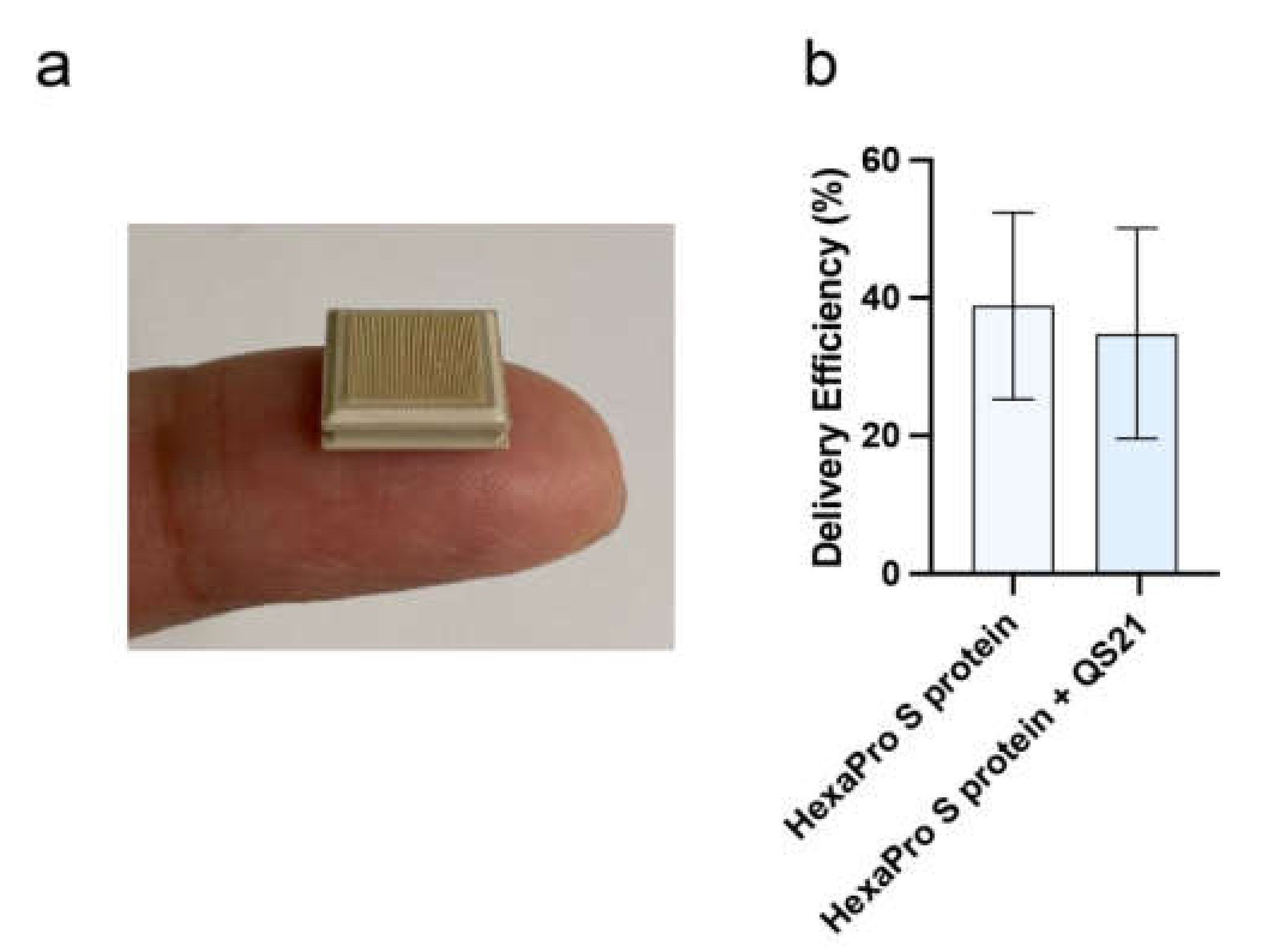
2.4. High-Density Microarray Patch (HD-MAP) Coating and Application
2.5. Vaccine Delivery Efficiency Using HD-MAP
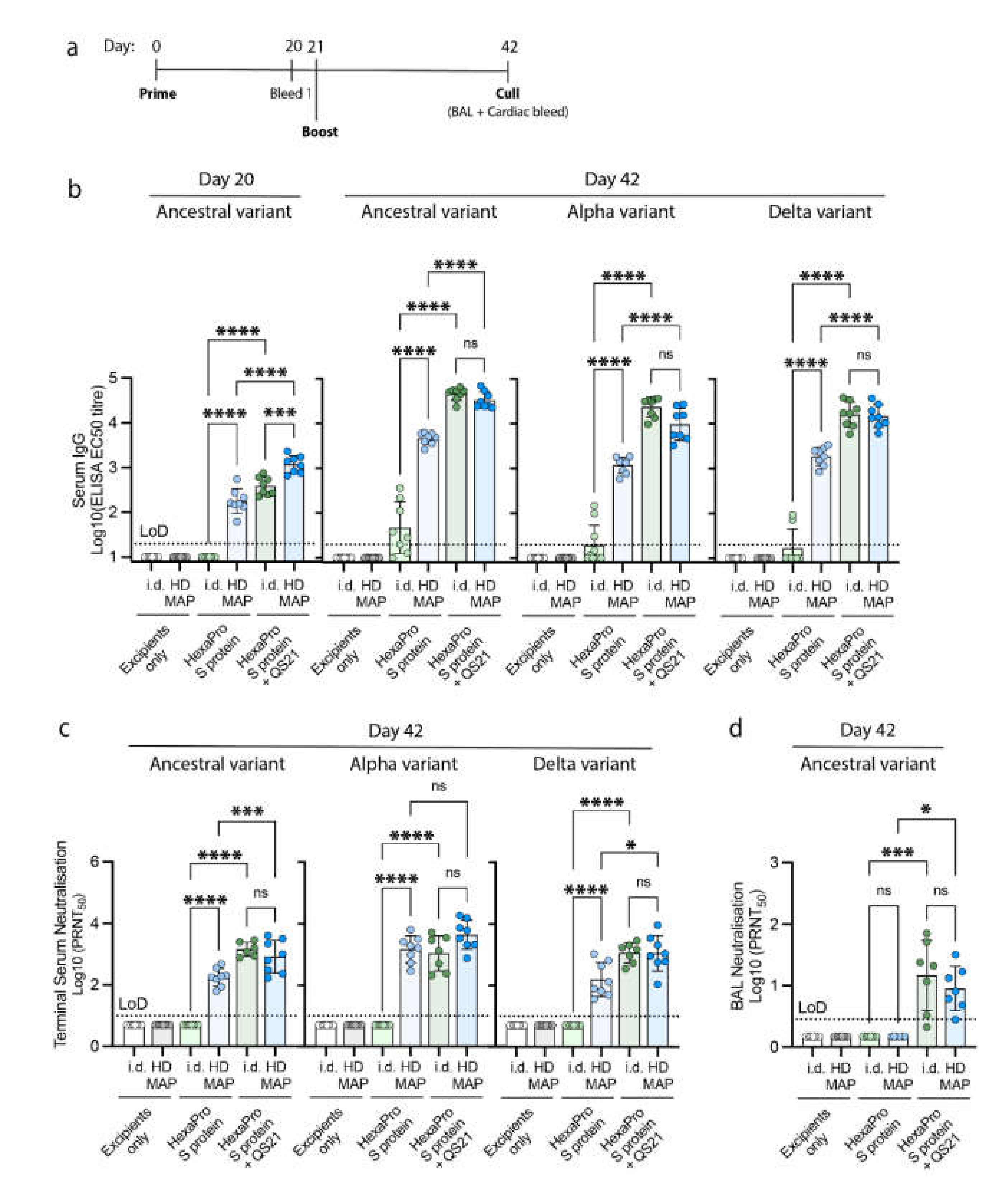
2.6. Immunization in an In Vivo Model
2.7. Collection of Serum and Bronchoalveolar Lavage (BAL)
2.8. Antibody Titer Detection by ELISA
2.9. Virus Neutralization by Serum and BALs
2.9.1. Virus Preparations
2.9.2. Plaque Reduction Neutralization Test (PRNT)
2.10. Ethics Statement
2.11. Statistical Analysis
3. Results
3.1. Characterization of SARS-CoV-2 HexaPro S Protein
3.2. Vaccine Delivery via HD-MAPs
3.3. Immune Responses Following HD-MAP Vaccination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- George, J.A.; Mayne, E.S. The Novel Coronavirus and Inflammation. In Clinical, Biological and Molecular Aspects of COVID-19; Guest, P.C., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 127–138. [Google Scholar] [CrossRef]
- Guo, Y.-R.; Cao, Q.-D.; Hong, Z.-S.; Tan, Y.-Y.; Chen, S.-D.; Jin, H.-J.; Tan, K.-S.; Wang, D.-Y.; Yan, Y. The origin, transmission and clinical therapies on coronavirus disease 2019 (COVID-19) outbreak—An update on the status. Mil. Med. Res. 2020, 7, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saxena, S.K.; Kumar, S.; Maurya, V.K.; Sharma, R.; Dandu, H.R.; Bhatt, M.L.B. Current Insight into the Novel Coronavirus Disease 2019 (COVID-19). In Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Saxena, S.K., Ed.; Springer Singapore: Singapore, 2020; pp. 1–8. [Google Scholar] [CrossRef]
- Van Vo, G.; Bagyinszky, E.; Park, Y.S.; Hulme, J.; An, S.S.A. SARS-CoV-2 (COVID-19): Beginning to Understand a New Virus. In Clinical, Biological and Molecular Aspects of COVID-19; Guest, P.C., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 3–19. [Google Scholar] [CrossRef]
- WHO. WHO Director-General’s Opening Remarks at the Media Briefing on COVID-19. 2020. Available online: https://www.who.int/director-general/speeches/detail/who-director-general-s-opening-remarks-at-the-media-briefing-on-covid-19---11-march-2020 (accessed on 6 February 2022).
- Licastro, D.; Rajasekharan, S.; Monego, S.D.; Segat, L.; D’Agaro, P.; Marcello, A.; Sandri-Goldin, R.M. Isolation and Full-Length Genome Characterization of SARS-CoV-2 from COVID-19 Cases in Northern Italy. J. Virol. 2020, 94, e00543-20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, N.; Zhang, D.; Wang, W.; Li, X.; Yang, B.; Song, J.; Zhao, X.; Huang, B.; Shi, W.; Lu, R.; et al. A Novel Coronavirus from Patients with Pneumonia in China, 2019. N. Engl. J. Med. 2020, 382, 727–733. [Google Scholar] [CrossRef] [PubMed]
- Caly, L.; Druce, J.; Roberts, J.; Bond, K.; Tran, T.; Kostecki, R.; Yoga, Y.; Naughton, W.; Taiaroa, G.; Seemann, T.; et al. Isolation and rapid sharing of the 2019 novel coronavirus (SARS-CoV-2) from the first patient diagnosed with COVID-19 in Australia. Med. J. Aust. 2020, 212, 459–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, W.B.; Kwon, N.-J.; Choi, S.-J.; Kang, C.K.; Choe, P.G.; Kim, J.Y.; Yun, J.; Lee, G.-W.; Seong, M.-W.; Kim, N.J.; et al. Virus Isolation from the First Patient with SARS-CoV-2 in Korea. J. Korean Med. Sci. 2020, 35, e84. [Google Scholar] [CrossRef] [PubMed]
- Gorbalenya, A.E.; Baker, S.C.; Baric, R.S.; de Groot, R.J.; Drosten, C.; Gulyaeva, A.A.; Haagmans, B.L.; Lauber, C.; Leontovich, A.M.; Neuman, B.W.; et al. The species Severe acute respiratory syndrome-related coronavirus: Classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 2020, 5, 536–544. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Morphology, Genome Organization, Replication, and Pathogenesis of Severe Acute Respiratory Syndrome Coronavirus 2 (SARS-CoV-2). In Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Saxena, S.K., Ed.; Springer Singapore: Singapore, 2020; pp. 23–31. [Google Scholar] [CrossRef]
- Dai, L.; Gao, G.F. Viral targets for vaccines against COVID-19. Nat. Rev. Immunol. 2021, 21, 73–82. [Google Scholar] [CrossRef]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef]
- Krammer, F. SARS-CoV-2 vaccines in development. Nature 2020, 586, 516–527. [Google Scholar] [CrossRef]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. NPJ Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Sternberg, A.; Naujokat, C. Structural features of coronavirus SARS-CoV-2 spike protein: Targets for vaccination. Life Sci. 2020, 257, 118056. [Google Scholar] [CrossRef] [PubMed]
- Walls, A.C.; Park, Y.-J.; Tortorici, M.A.; Wall, A.; McGuire, A.T.; Veesler, D. Structure, Function, and Antigenicity of the SARS-CoV-2 Spike Glycoprotein. Cell 2020, 181, 281–292.e6. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Nyodu, R.; Maurya, V.K.; Saxena, S.K. Host Immune Response and Immunobiology of Human SARS-CoV-2 Infection. In Coronavirus Disease 2019 (COVID-19): Epidemiology, Pathogenesis, Diagnosis, and Therapeutics; Saxena, S.K., Ed.; Springer Singapore: Singapore, 2020; pp. 43–53. [Google Scholar] [CrossRef]
- Shojaee, A.; Vahedian-Azimi, A.; Faizi, F.; Rahimi-Bashar, F.; Shahriary, A.; Galeh, H.E.G.; Nehrir, B.; Guest, P.C.; Sahebkar, A. Relationship between COVID-19 and Angiotensin-Converting Enzyme 2: A Scoping Review. In Clinical, Biological and Molecular Aspects of COVID-19; Guest, P.C., Ed.; Springer International Publishing: Cham, Switzerland, 2021; pp. 53–68. [Google Scholar] [CrossRef]
- Voysey, M.; Clemens, S.A.C.; Madhi, S.A.; Weckx, L.Y.; Folegatti, P.M.; Aley, P.K.; Angus, B.; Baillie, V.L.; Barnabas, S.L.; Bhorat, Q.E.; et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: An interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. Lancet 2021, 397, 99–111. [Google Scholar] [CrossRef]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 mRNA Covid-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Study of Safety and Immunogenicity of Adjuvanted SARS-CoV-2 (SCB-2019) Vaccine in Adults with Chronic Immune-Mediated Diseases. Available online: https://ClinicalTrials.gov/show/NCT05012787 (accessed on 6 February 2022).
- A Study to Evaluate the Efficacy, Immune Response, and Safety of a COVID-19 Vaccine in Adults ≥ 18 Years with a Pediatric Expansion in Adolescents (12 to <18 Years) at Risk for SARS-CoV-2. Available online: https://ClinicalTrials.gov/show/NCT04611802 (accessed on 6 February 2022).
- Study of Recombinant Protein Vaccines with Adjuvant as a Primary Series and as a Booster Dose against COVID-19 in Adults 18 Years of Age and Older. Available online: https://ClinicalTrials.gov/show/NCT04762680 (accessed on 6 February 2022).
- Immunogenicity and Safety Study of SK SARS-CoV-2 Recombinant Nanoparticle Vaccine (GBP510) Adjuvanted with AS03 (COVID-19). Available online: https://ClinicalTrials.gov/show/NCT05007951 (accessed on 6 February 2022).
- WHO. COVID-19 Vaccine Tracker and Landscape. 2021. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 6 February 2022).
- WHO. WHO Recommendation BioNtech Tozinameran—COVID-19 mRNA Vaccine (Nucleoside Modified)—COMIRNATY®. 2020. Available online: https://extranet.who.int/pqweb/vaccines/who-recommendation-covid-19-mrna-vaccine-nucleoside-modified-comirnaty (accessed on 6 February 2022).
- WHO. WHO Recommendation Moderna COVID-19 mRNA Vaccine (Nucleoside Modified). 2021. Available online: https://extranet.who.int/pqweb/vaccines/covid-19-mrna-vaccine-nucleoside-modified (accessed on 6 February 2022).
- WHO. WHO Recommendation AstraZeneca/SKBio—COVID-19 Vaccine (ChAdOx1-S [Recombinant]). 2021. Available online: https://extranet.who.int/pqweb/vaccines/covid-19-vaccine-chadox1-s-recombinant (accessed on 6 February 2022).
- WHO. WHO Recommendation Bharat Biotech International Ltd—COVID-19 Vaccine (Whole Virion Inactivated Corona Virus). 2021. Available online: https://extranet.who.int/pqweb/vaccines/who-recommendation-bharat-biotech-international-ltd-covid-19-vaccine-whole-virion (accessed on 6 February 2022).
- WHO. WHO Recommendation COVID-19 Vaccine BIBP/Sinopharm. 2021. Available online: https://extranet.who.int/pqweb/vaccines/who-recommendation-covid-19-vaccine-bibp (accessed on 6 February 2022).
- WHO. WHO Recommendation Janssen–Cilag International NV (Belgium) COVID-19 Vaccine (Ad26.COV2-S [Recombinant]). 2021. Available online: https://www.medbox.org/document/who-recommendation-janssencilag-international-nv-belgium-covid-19-vaccine-ad26cov2-s-recombinant (accessed on 6 February 2022).
- WHO. WHO Recommendation of Sinovac COVID-19 vaccine (Vero Cell [Inactivated])—CoronaVac. 2021. Available online: https://extranet.who.int/pqweb/vaccines/who-recommendation-sinovac-covid-19-vaccine-vero-cell-inactivated-coronavac (accessed on 6 February 2022).
- WHO. WHO Recommendation Serum Institute of India Pvt Ltd—COVID-19 Vaccine (ChAdOx1-S [Recombinant])—COVISHIELD™. 2021. Available online: https://extranet.who.int/pqweb/vaccines/covid-19-vaccine-chadox1-s-recombinant-covishield (accessed on 6 February 2022).
- Arya, J.; Prausnitz, M.R. Microneedle patches for vaccination in developing countries. J. Control Release 2016, 240, 135–141. [Google Scholar] [CrossRef] [Green Version]
- Marshall, S.; Sahm, L.J.; Moore, A.C. The success of microneedle-mediated vaccine delivery into skin. Hum. Vaccines Immunother. 2016, 12, 2975–2983. [Google Scholar] [CrossRef] [Green Version]
- Babiuk, S.; Baca-Estrada, M.; Babiuk, L.A.; Ewen, C.; Foldvari, M. Cutaneous vaccination: The skin as an immunologically active tissue and the challenge of antigen delivery. J. Control Release 2000, 66, 199–214. [Google Scholar] [CrossRef]
- Muller, D.A.; Henricson, J.; Baker, S.B.; Togö, T.; Jayashi Flores, C.M.; Lemaire, P.A.; Forster, A.; Anderson, C.D. Innate local response and tissue recovery following application of high density microarray patches to human skin. Sci. Rep. 2020, 10, 18468. [Google Scholar] [CrossRef]
- Ng, H.-I.; Tuong, Z.K.; Fernando, G.J.P.; Depelsenaire, A.C.I.; Meliga, S.C.; Frazer, I.H.; Kendall, M.A.F. Microprojection arrays applied to skin generate mechanical stress, induce an inflammatory transcriptome and cell death, and improve vaccine-induced immune responses. NPJ Vaccines 2019, 4, 41. [Google Scholar] [CrossRef] [Green Version]
- Forster, A.H.; Witham, K.; Depelsenaire, A.C.I.; Veitch, M.; Wells, J.W.; Wheatley, A.; Pryor, M.; Lickliter, J.D.; Francis, B.; Rockman, S.; et al. Safety, tolerability, and immunogenicity of influenza vaccination with a high-density microarray patch: Results from a randomized, controlled phase I clinical trial. PLoS Med. 2020, 17, e1003024. [Google Scholar] [CrossRef] [PubMed]
- McMillan, C.L.D.; Choo, J.J.Y.; Idris, A.; Supramaniam, A.; Modhiran, N.; Amarilla, A.A.; Isaacs, A.; Cheung, S.T.M.; Liang, B.; Bielefeldt-Ohmann, H.; et al. Complete protection by a single-dose skin patch-delivered SARS-CoV-2 spike vaccine. Sci. Adv. 2021, 7, eabj8065. [Google Scholar] [CrossRef] [PubMed]
- Hsieh, C.-L.; Goldsmith, J.A.; Schaub, J.M.; DiVenere, A.M.; Kuo, H.-C.; Javanmardi, K.; Le, K.C.; Wrapp, D.; Lee, A.G.; Liu, Y.; et al. Structure-based design of prefusion-stabilized SARS-CoV-2 spikes. Science 2020, 369, 1501–1505. [Google Scholar] [CrossRef]
- Pino, P.; Kint, J.; Kiseljak, D.; Agnolon, V.; Corradin, G.; Kajava, A.V.; Rovero, P.; Dijkman, R.; den Hartog, G.; McLellan, J.S.; et al. Trimeric SARS-CoV-2 Spike Proteins Produced from CHO Cells in Bioreactors Are High-Quality Antigens. Processes 2020, 8, 1539. [Google Scholar] [CrossRef]
- Schaub, J.M.; Chou, C.-W.; Kuo, H.-C.; Javanmardi, K.; Hsieh, C.-L.; Goldsmith, J.; DiVenere, A.M.; Le, K.C.; Wrapp, D.; Byrne, P.O.; et al. Expression and characterization of SARS-CoV-2 spike proteins. Nat. Protoc. 2021, 16, 5339–5356. [Google Scholar] [CrossRef]
- Seephetdee, C.; Buasri, N.; Bhukhai, K.; Srisanga, K.; Manopwisedjaroen, S.; Lertjintanakit, S.; Phueakphud, N.; Pakiranay, C.; Kangwanrangsan, N.; Srichatrapimuk, S.; et al. Mice Immunized with the Vaccine Candidate HexaPro Spike Produce Neutralizing Antibodies against SARS-CoV-2. Vaccines 2021, 9, 498. [Google Scholar] [CrossRef]
- Liu, L.; Wang, P.; Nair, M.S.; Yu, J.; Rapp, M.; Wang, Q.; Luo, Y.; Chan, J.F.W.; Sahi, V.; Figueroa, A.; et al. Potent neutralizing antibodies against multiple epitopes on SARS-CoV-2 spike. Nature 2020, 584, 450–456. [Google Scholar] [CrossRef]
- Martinez, D.R.; Schaefer, A.; Gobeil, S.; Li, D.; De la Cruz, G.; Parks, R.; Lu, X.; Barr, M.; Manne, K.; Mansouri, K.; et al. A broadly neutralizing antibody protects against SARS-CoV, pre-emergent bat CoVs, and SARS-CoV-2 variants in mice. bioRxiv 2021. [Google Scholar] [CrossRef]
- Pinto, D.; Park, Y.-J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A.; Bianchi, S.; Jaconi, S.; Culap, K.; Zatta, F.; De Marco, A.; et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef]
- Rattanapisit, K.; Shanmugaraj, B.; Manopwisedjaroen, S.; Purwono, P.B.; Siriwattananon, K.; Khorattanakulchai, N.; Hanittinan, O.; Boonyayothin, W.; Thitithanyanont, A.; Smith, D.R.; et al. Rapid production of SARS-CoV-2 receptor binding domain (RBD) and spike specific monoclonal antibody CR3022 in Nicotiana benthamiana. Sci. Rep. 2020, 10, 17698. [Google Scholar] [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef] [PubMed]
- Tian, X.; Li, C.; Huang, A.; Xia, S.; Lu, S.; Shi, Z.; Lu, L.; Jiang, S.; Yang, Z.; Wu, Y.; et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 2020, 9, 382–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez, D.R.; Schäfer, A.; Gobeil, S.; Li, D.; Cruz, G.D.l.; Parks, R.; Lu, X.; Barr, M.; Stalls, V.; Janowska, K.; et al. A broadly cross-reactive antibody neutralizes and protects against sarbecovirus challenge in mice. Sci. Transl. Med. 2022, 14, eabj7125. [Google Scholar] [CrossRef] [PubMed]
- Ter Meulen, J.; van den Brink, E.N.; Poon, L.L.; Marissen, W.E.; Leung, C.S.; Cox, F.; Cheung, C.Y.; Bakker, A.Q.; Bogaards, J.A.; van Deventer, E.; et al. Human monoclonal antibody combination against SARS coronavirus: Synergy and coverage of escape mutants. PLoS Med. 2006, 3, e237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuan, M.; Wu, N.C.; Zhu, X.; Lee, C.-C.D.; So, R.T.Y.; Lv, H.; Mok, C.K.P.; Wilson, I.A. A highly conserved cryptic epitope in the receptor binding domains of SARS-CoV-2 and SARS-CoV. Science 2020, 368, 630–633. [Google Scholar] [CrossRef] [Green Version]
- Amarilla, A.A.; Modhiran, N.; Setoh, Y.X.; Peng, N.Y.G.; Sng, J.D.J.; Liang, B.; McMillan, C.L.D.; Freney, M.E.; Cheung, S.T.M.; Chappell, K.J.; et al. An Optimized High-Throughput Immuno-Plaque Assay for SARS-CoV-2. Front. Microbiol. 2021, 12, 625136. [Google Scholar] [CrossRef]
- Katzelnick, L.C.; Coello Escoto, A.; McElvany, B.D.; Chávez, C.; Salje, H.; Luo, W.; Rodriguez-Barraquer, I.; Jarman, R.; Durbin, A.P.; Diehl, S.A.; et al. Viridot: An automated virus plaque (immunofocus) counter for the measurement of serological neutralizing responses with application to dengue virus. PLoS Negl. Trop. Dis. 2018, 12, e0006862. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.J.; Mansouri, K.; Stalls, V.; Manne, K.; Watts, B.; Parks, R.; Janowska, K.; Gobeil, S.M.C.; Kopp, M.; Li, D.; et al. Cold sensitivity of the SARS-CoV-2 spike ectodomain. Nat. Struct. Mol. Biol. 2021, 28, 128–131. [Google Scholar] [CrossRef]
- Kim, Y.C.; Reyes-Sandoval, A. Chapter 7—Viral-vectored vaccines against SARS-CoV-2. In Biomedical Innovations to Combat COVID-19; Rosales-Mendoza, S., Comas-Garcia, M., Gonzalez-Ortega, O., Eds.; Academic Press: London, UK, 2022; pp. 115–127. [Google Scholar] [CrossRef]
- Scheiblhofer, S.; Laimer, J.; Machado, Y.; Weiss, R.; Thalhamer, J. Influence of protein fold stability on immunogenicity and its implications for vaccine design. Expert Rev. Vaccines 2017, 16, 479–489. [Google Scholar] [CrossRef]
- Choo, J.J.Y.; Vet, L.J.; McMillan, C.L.D.; Harrison, J.J.; Scott, C.A.P.; Depelsenaire, A.C.I.; Fernando, G.J.P.; Watterson, D.; Hall, R.A.; Young, P.R.; et al. A chimeric dengue virus vaccine candidate delivered by high density microarray patches protects against infection in mice. NPJ Vaccines 2021, 6, 66. [Google Scholar] [CrossRef]
- Fernando, G.J.; Chen, X.; Primiero, C.A.; Yukiko, S.R.; Fairmaid, E.J.; Corbett, H.J.; Frazer, I.H.; Brown, L.E.; Kendall, M.A. Nanopatch targeted delivery of both antigen and adjuvant to skin synergistically drives enhanced antibody responses. J. Control Release 2012, 159, 215–221. [Google Scholar] [CrossRef] [PubMed]
- Fernando, G.J.; Chen, X.; Prow, T.W.; Crichton, M.L.; Fairmaid, E.J.; Roberts, M.S.; Frazer, I.H.; Brown, L.E.; Kendall, M.A. Potent immunity to low doses of influenza vaccine by probabilistic guided micro-targeted skin delivery in a mouse model. PLoS ONE 2010, 5, e10266. [Google Scholar] [CrossRef] [PubMed]
- Fernando, G.J.P.; Hickling, J.; Jayashi Flores, C.M.; Griffin, P.; Anderson, C.D.; Skinner, S.R.; Davies, C.; Witham, K.; Pryor, M.; Bodle, J.; et al. Safety, tolerability, acceptability and immunogenicity of an influenza vaccine delivered to human skin by a novel high-density microprojection array patch (Nanopatch™). Vaccine 2018, 36, 3779–3788. [Google Scholar] [CrossRef] [PubMed]
- Muller, D.A.; Depelsenaire, A.C.I.; Shannon, A.E.; Watterson, D.; Corrie, S.R.; Owens, N.S.; Agyei-Yeboah, C.; Cheung, S.T.M.; Zhang, J.; Fernando, G.J.P.; et al. Efficient Delivery of Dengue Virus Subunit Vaccines to the Skin by Microprojection Arrays. Vaccines 2019, 7, 189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muller, D.A.; Fernando, G.J.P.; Owens, N.S.; Agyei-Yeboah, C.; Wei, J.C.J.; Depelsenaire, A.C.I.; Forster, A.; Fahey, P.; Weldon, W.C.; Oberste, M.S.; et al. High-density microprojection array delivery to rat skin of low doses of trivalent inactivated poliovirus vaccine elicits potent neutralising antibody responses. Sci. Rep. 2017, 7, 12644. [Google Scholar] [CrossRef]
- Muller, D.A.; Pearson, F.E.; Fernando, G.J.; Agyei-Yeboah, C.; Owens, N.S.; Corrie, S.R.; Crichton, M.L.; Wei, J.C.; Weldon, W.C.; Oberste, M.S.; et al. Inactivated poliovirus type 2 vaccine delivered to rat skin via high density microprojection array elicits potent neutralising antibody responses. Sci. Rep. 2016, 6, 22094. [Google Scholar] [CrossRef]
- Moyle, P.M.; Toth, I. Modern Subunit Vaccines: Development, Components, and Research Opportunities. ChemMedChem 2013, 8, 360–376. [Google Scholar] [CrossRef]
- Sun, H.-X.; Xie, Y.; Ye, Y.-P. Advances in saponin-based adjuvants. Vaccine 2009, 27, 1787–1796. [Google Scholar] [CrossRef]
- Fontanet, A.; Autran, B.; Lina, B.; Kieny, M.P.; Karim, S.S.A.; Sridhar, D. SARS-CoV-2 variants and ending the COVID-19 pandemic. Lancet 2021, 397, 952–954. [Google Scholar] [CrossRef]
- Martínez-Flores, D.; Zepeda-Cervantes, J.; Cruz-Reséndiz, A.; Aguirre-Sampieri, S.; Sampieri, A.; Vaca, L. SARS-CoV-2 Vaccines Based on the Spike Glycoprotein and Implications of New Viral Variants. Front. Immunol. 2021, 12, 701501. [Google Scholar] [CrossRef]
- WHO. WHO Update on Omicron. Update on Omicron. 2021. Available online: https://www.who.int/news/item/28-11-2021-update-on-omicron (accessed on 6 February 2022).
- Araf, Y.; Akter, F.; Tang, Y.-D.; Fatemi, R.; Parvez, M.S.A.; Zheng, C.; Hossain, M.G. Omicron variant of SARS-CoV-2: Genomics, transmissibility, and responses to current COVID-19 vaccines. J. Med. Virol. 2022, 94, 1825–1832. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.-Y.; Wang, W.-B.; Gao, R.-D.; Zhou, A.-M. Omicron variant (B.1.1.529) of SARS-CoV-2: Mutation, infectivity, transmission, and vaccine resistance. World J. Clin. Cases 2022, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, M.; Krüger, N.; Schulz, S.; Cossmann, A.; Rocha, C.; Kempf, A.; Nehlmeier, I.; Graichen, L.; Moldenhauer, A.-S.; Winkler, M.S.; et al. The Omicron variant is highly resistant against antibody-mediated neutralization: Implications for control of the COVID-19 pandemic. Cell 2022, 185, 447–456.e11. [Google Scholar] [CrossRef] [PubMed]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McMillan, C.L.D.; Azuar, A.; Choo, J.J.Y.; Modhiran, N.; Amarilla, A.A.; Isaacs, A.; Honeyman, K.E.; Cheung, S.T.M.; Liang, B.; Wurm, M.J.; et al. Dermal Delivery of a SARS-CoV-2 Subunit Vaccine Induces Immunogenicity against Variants of Concern. Vaccines 2022, 10, 578. https://doi.org/10.3390/vaccines10040578
McMillan CLD, Azuar A, Choo JJY, Modhiran N, Amarilla AA, Isaacs A, Honeyman KE, Cheung STM, Liang B, Wurm MJ, et al. Dermal Delivery of a SARS-CoV-2 Subunit Vaccine Induces Immunogenicity against Variants of Concern. Vaccines. 2022; 10(4):578. https://doi.org/10.3390/vaccines10040578
Chicago/Turabian StyleMcMillan, Christopher L. D., Armira Azuar, Jovin J. Y. Choo, Naphak Modhiran, Alberto A. Amarilla, Ariel Isaacs, Kate E. Honeyman, Stacey T. M. Cheung, Benjamin Liang, Maria J. Wurm, and et al. 2022. "Dermal Delivery of a SARS-CoV-2 Subunit Vaccine Induces Immunogenicity against Variants of Concern" Vaccines 10, no. 4: 578. https://doi.org/10.3390/vaccines10040578