
Thermophilic Filamentous Fungus C1-Cell-Cloned SARS-CoV-2-Spike-RBD-Subunit-Vaccine Adjuvanted with Aldydrogel®85 Protects K18-hACE2 Mice against Lethal Virus Challenge
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Construction of Spike-S1333–527RBD-C-Tag Production Strains
2.2. PCR DNA Analysis of Recombinant Cell Clones Showed the Expected Correct RBD Expression Cassette Integration and Clone Purity
2.3. Fungal C1 Fermentation Recombinant Subunit Antigen Manufacturing Process Development in Low-Cost Glucose Fermentation Media
Off-Line Analytics of the Fermentations
2.4. Production of the RBD-C-Tag in 10-Liter Fermenters
2.5. Purification of S1333–527RBD-C-Tag Glycoprotein Antigen Using Capture-Select™ C-Tag Affinity Matrix
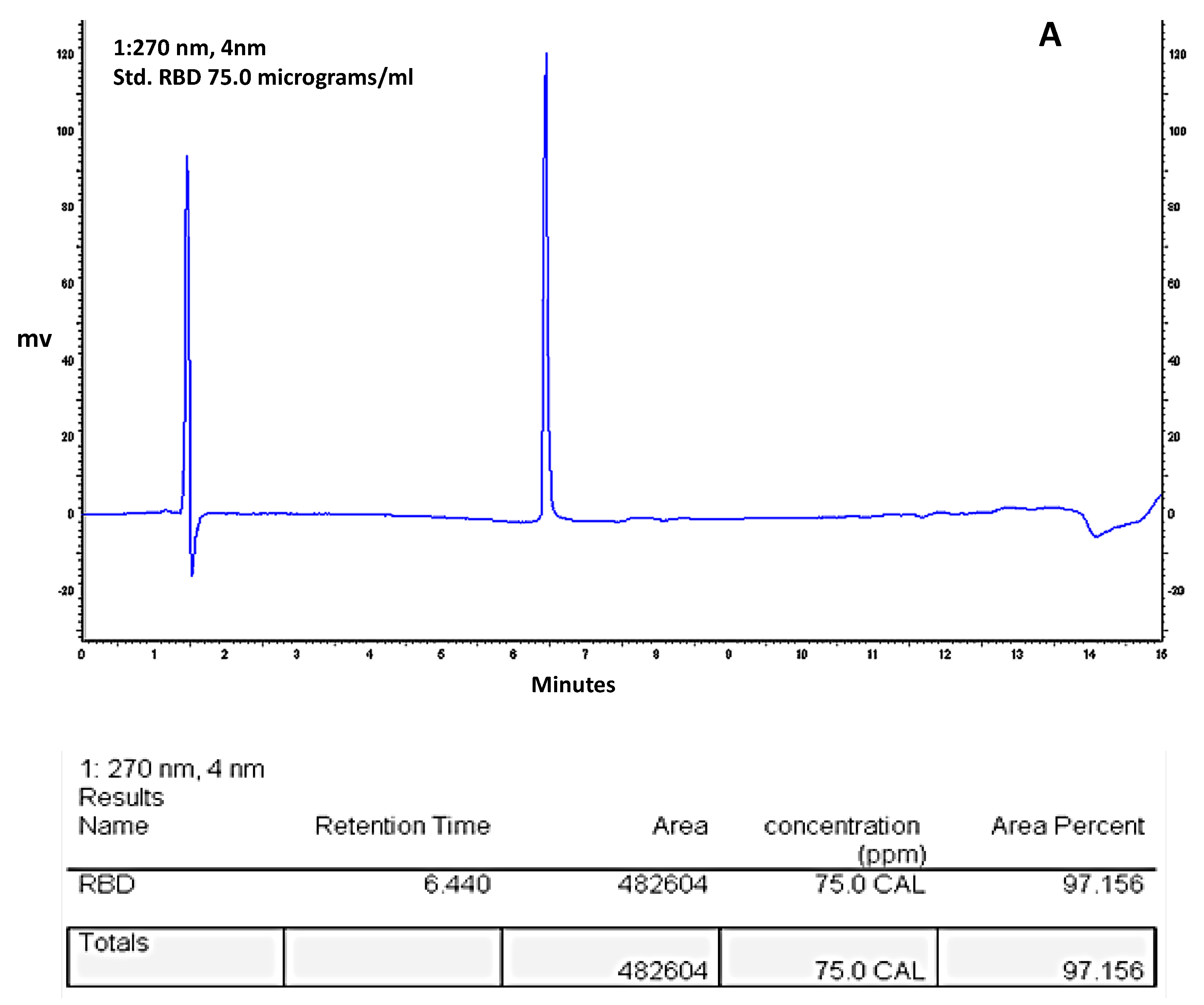
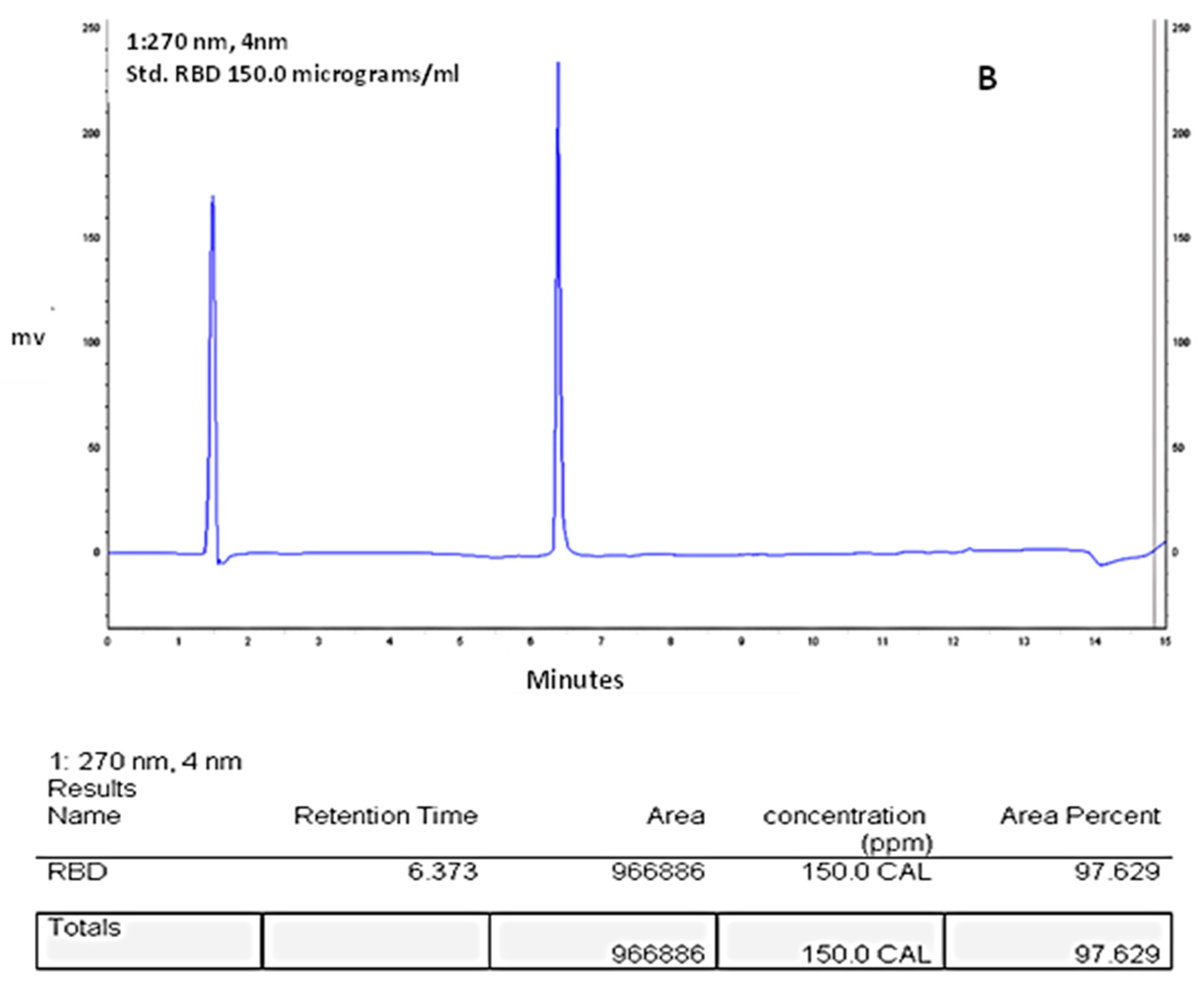
2.6. RBD-C-Tag Initial Purity Assessment by Reverse Phase HPLC
2.7. Glycoprotein Analyses and RBD-C-Tag Antigen Characterizations by SDS-PAGE and Anti-RBD IgG Western Immunoblotting
2.8. RBD-C-Tag Sample Preparation, LC-MS and Data Processing
2.9. SARS-CoV-2 Virion Specific Antibody Monitored by Plaque Reduction Neutralization Test (PRNT)
2.10. RBD-C-Tag Antigen Vaccine Candidate Preparation and RBD-C-Tag Alhydrogel®‘85’ Gel Binding
2.11. RBD Immunogenicity Assessments in Experimental Mice
2.12. Alhydrogel®‘85’, Aluminum Hydroxide Oxide Gel [AlO(OH)], Adjuvant Dose Ranging
2.13. K18-hACE2 Transgenic C57BL/6J Mouse Challenge Tests with Lethal Live SARS-CoV-2 Virus
2.14. Antibody Iso-Type Immunoglobulin G (IgG) Class Subtyping of DYAI-100A85 Vaccinated K18-hACE2 Transgenic C57BL/6J Mice
2.15. Statistical Analysis
3. Results
3.1. Production of the SARS-CoV-2-Spike-S1333–527RBD-C-Tag Glycoprotein Recombinant Subunit Antigen in Thermothelomyces Heterothallica
3.2. Integrity and Purity of RBD-C-Tag Subunit Recombinant Antigen Vaccine Harvested at 90-h EFT, as Monitored by Protein SDS-PAGE
3.3. LC-MS-MS Analysis of the Purity of the Affinity Purified S1333–527RBD-C-Tag Glycoprotein Subunit Antigen
3.4. HPLC-Reverse Phase Chromatography Analysis of Purity of the Affinity Purified S1333–527RBD-C-Tag Glycoprotein Subunit Antigen
3.5. RBD-C-Tag Antigen Binding to Recombinant Human ACE2 (Angiotensin Converting Enzyme 2)
3.6. S1333–527RBD-C-Tag Recombinant Subunit Antigen Binding to Aluminum Oxide Hydroxide, Alhydrogel®‘85’ Adjuvant
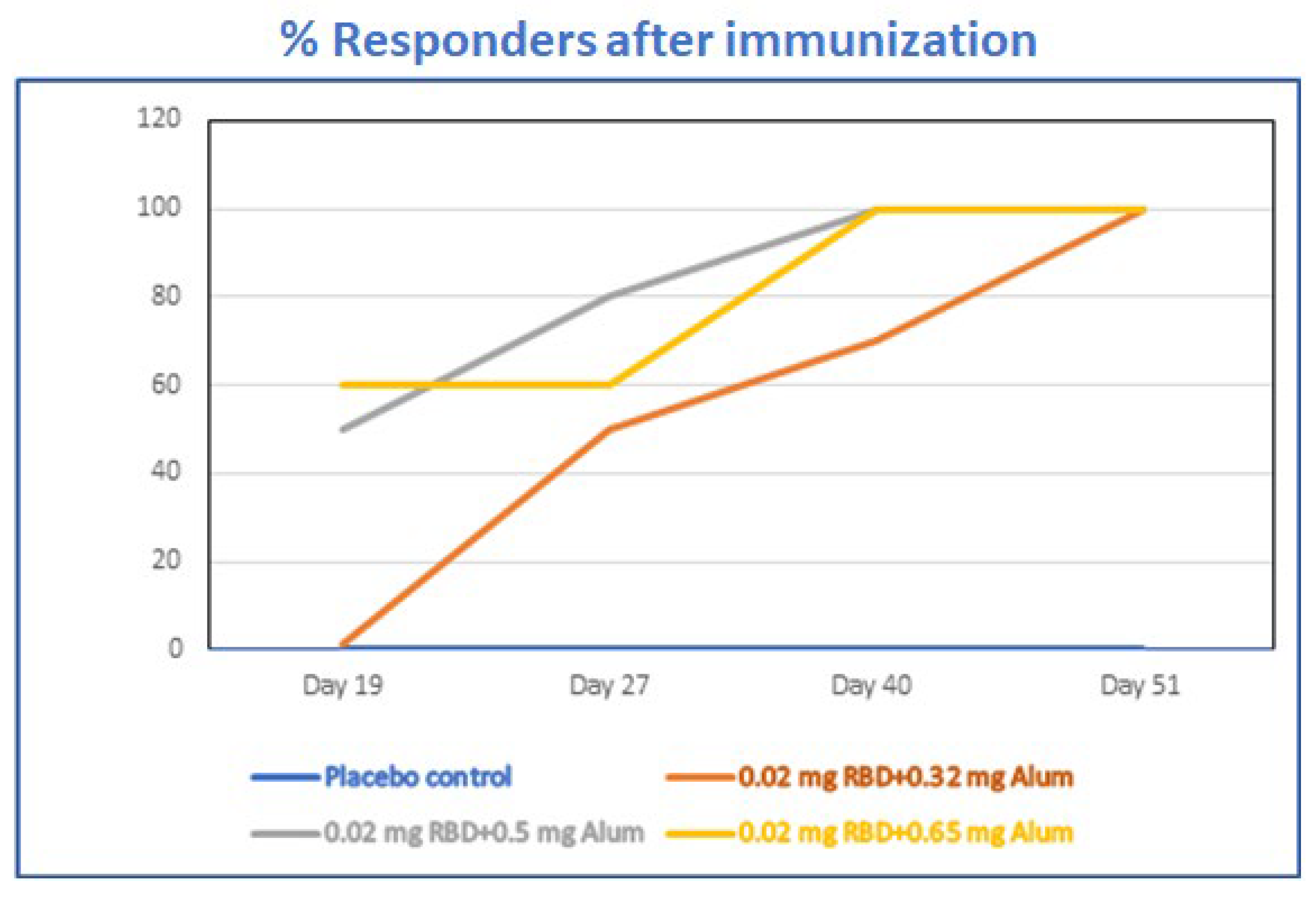
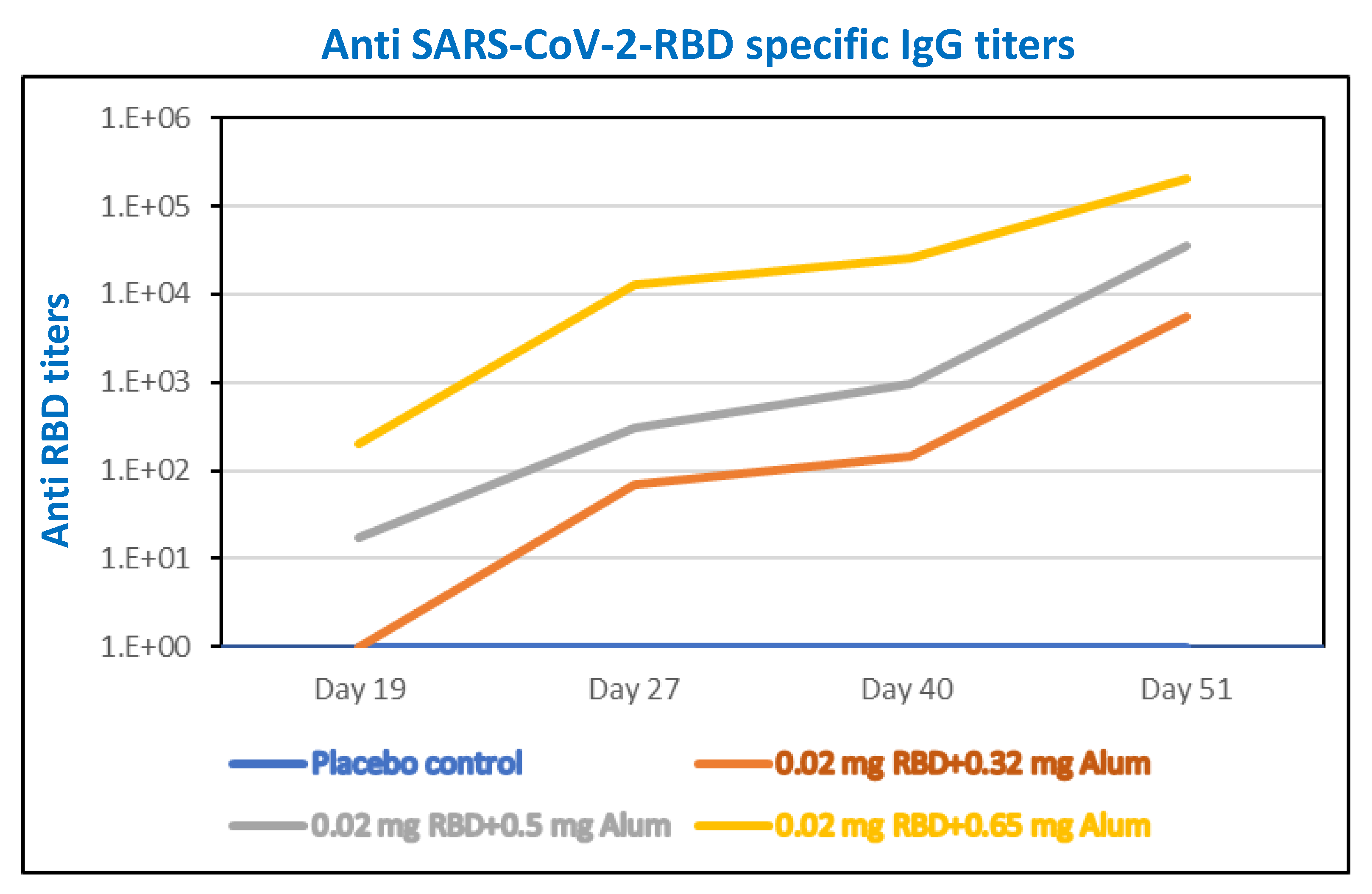
3.7. Alhydrogel®‘85’ Adjuvant Dose Range Screening of Adjuvanted-RBD Subunit Antigen Vaccine Immunogenicity in Mice
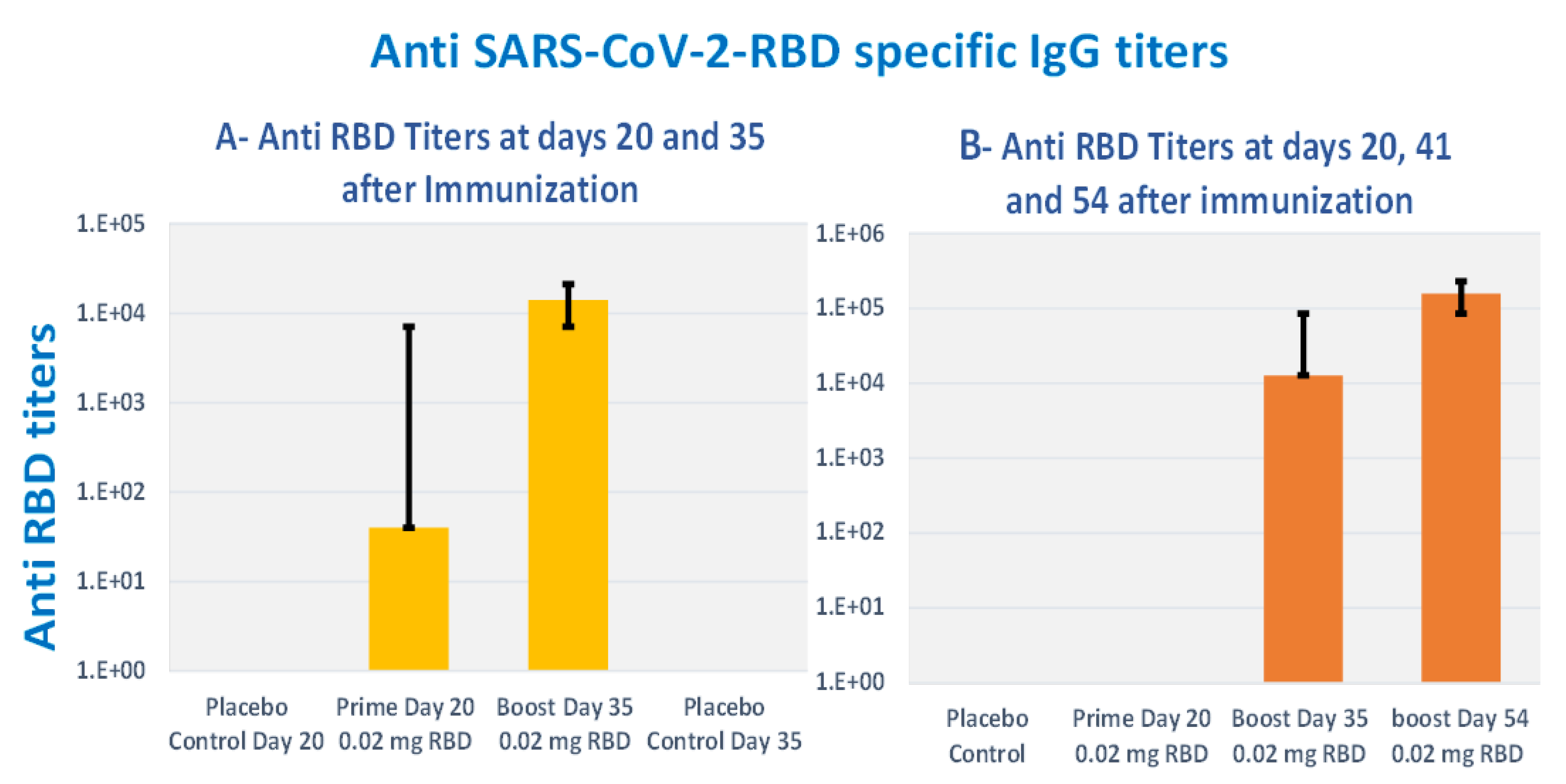
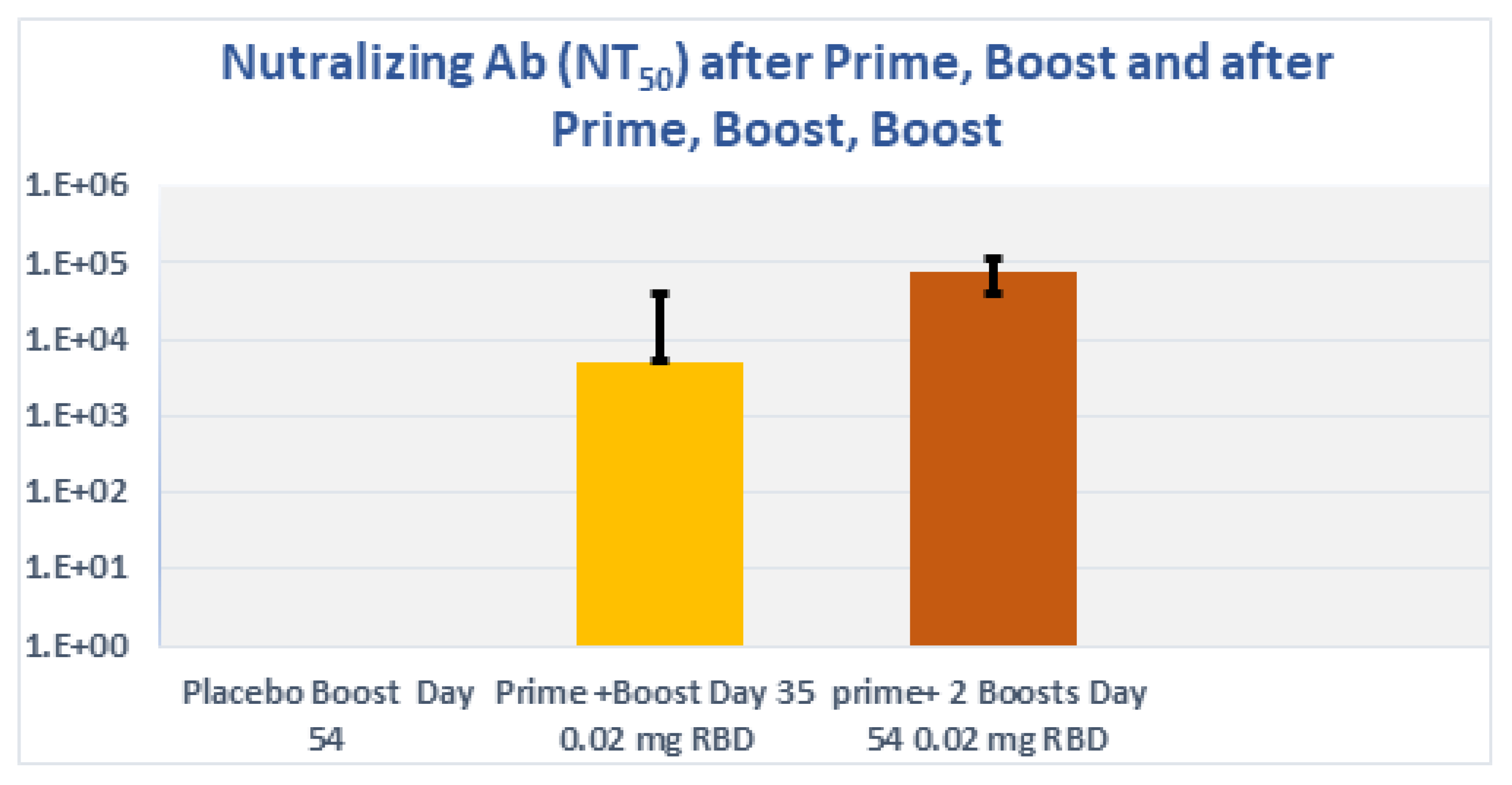
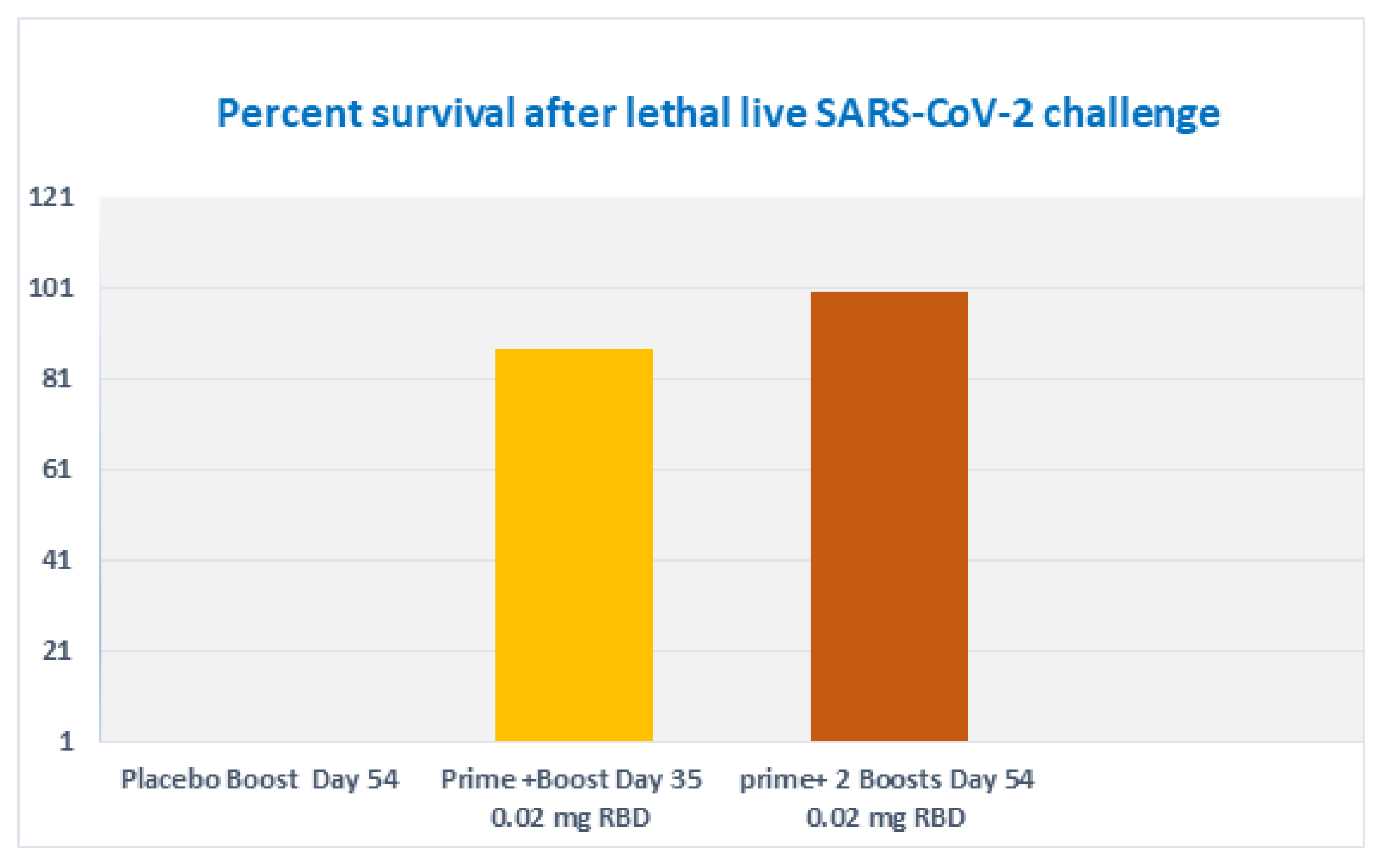
3.8. Subcutaneous Prime and Boost Vaccine Inoculations of Alhydrogel®‘85’ Adjuvanted RBD-C-Tag Antigen (DYAI-100A85) Protect k18-hACE2 Transgenic Mice against Intra-Nasal Challenge with Lethal SARS-CoV-2 Live Virus
3.9. Serum IgG Subtyping of DYAI-100A85 Vaccinated K18-huACE2 Transgenic C57BL/6J Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization. Tracking SARS-CoV-2 Variants. 2022. Available online: https://www.who.int/en/activities/tracking-SARS-CoV-2-variants/ (accessed on 5 August 2022).
- Khateeb, J.; Li, Y.; Zhang, H. Emerging SARS-CoV-2 variants of concern and potential intervention approaches. Crit. Care 2021, 25, 244. [Google Scholar] [CrossRef] [PubMed]
- Peacock, T.P.; Brown, J.C.; Zhou, J.; Thakur, N.; Newman, J.; Kugathasan, R.; Sukhova, K.; Kaforou, M.; Bailey, D.; Barclay, W.S. The SARS-CoV-2 variant, Omicron, shows rapid replication in human primary nasal epithelial cultures and efficiently uses the endosomal route of entry. bioRxiv 2022. [Google Scholar] [CrossRef]
- Willett, B.J.; Grove, J.; MacLean, O.A.; Wilkie, C.; Logan, N.; De Lorenzo, G.; Furnon, W.; Scott, S.; Manali, M.; Szemiel, A.; et al. The hyper-transmissible SARS-CoV-2 Omicron variant exhibits significant antigenic change, vaccine escape and a switch in cell entry mechanism. medRxiv 2022. [Google Scholar] [CrossRef]
- Mallapaty, S.; Callaway, E.; Kozlov, M.; Ledford, H.; Pickrell, J.; Van Noorden, R. How COVID vaccines shaped 2021 in eight powerful charts. Nature 2021, 600, 580–583. [Google Scholar] [CrossRef] [PubMed]
- Kyriakidis, N.C.; López-Cortés, A.; González, E.V.; Grimaldos, A.B.; Prado, E.O. SARS-CoV-2 vaccines strategies: A comprehensive review of phase 3 candidates. npj Vaccines 2021, 6, 28. [Google Scholar] [CrossRef]
- Premkumar, L.; Segovia-Chumbez, B.; Jadi, R.; Martinez, D.R.; Raut, R.; Markmann, A.; Cornaby, C.; Bartelt, L.; Weiss, S.; Park, Y.; et al. The receptor binding domain of the viral spike protein is an immunodominant and highly specific target of antibodies in SARS-CoV-2 patients. Sci. Immunol. 2020, 5, eabc8413. [Google Scholar] [CrossRef]
- Shi, R.; Shan, C.; Duan, X.; Chen, Z.; Liu, P.; Song, J.; Song, T.; Bi, X.; Han, C.; Wu, L.; et al. A human neutralizing antibody targets the receptor-binding site of SARS-CoV-2. Nature 2020, 584, 120–124. [Google Scholar] [CrossRef]
- Du, Y.; Shi, R.; Zhang, Y.; Duan, X.; Li, L.; Wang, F.; Zhang, R.; Shen, H.; Wang, Y.; Wu, Z.; et al. A broadly neutralizing humanized ACE2-targeting antibody against SARS-CoV-2 variants. Nat. Commun. 2021, 12, 5000. [Google Scholar] [CrossRef]
- Li, W.; Moore, M.J.; Vasilieva, N.; Sui, J.; Wong, S.K.; Berne, M.A.; Somasundaran, M.; Sullivan, J.L.; Luzuriaga, K.; Greenough, T.C.; et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 2003, 426, 450–454. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Li, W.; Farzan, M.; Harrison, S.C. Structure of SARS Coronavirus Spike Receptor-Binding Domain Complexed with Receptor. Science 2005, 309, 1864–1868. [Google Scholar] [CrossRef]
- Piccoli, L.; Park, Y.-J.; Tortorici, M.A.; Czudnochowski, N.; Walls, A.C.; Beltramello, M.; Silacci-Fregni, C.; Pinto, D.; Rosen, L.E.; Bowen, J.E.; et al. Mapping Neutralizing and Immunodominant Sites on the SARS-CoV-2 Spike Receptor-Binding Domain by Structure-Guided High-Resolution Serology. Cell 2020, 183, 1024–1042.e21. [Google Scholar] [CrossRef]
- Edara, V.V.; Norwood, C.; Floyd, K.; Lai, L.; Davis-Gardner, M.E.; Hudson, W.H.; Mantus, G.; Nyhoff, L.E.; Adelman, M.W.; Fineman, R.; et al. Infection- and vaccine-induced antibody binding and neutralization of the B.1.351 SARS-CoV-2 variant. Cell Host Microbe 2021, 29, 516–521.e3. [Google Scholar] [CrossRef]
- Starr, T.N.; Greaney, A.J.; Addetia, A.; Hannon, W.W.; Choudhary, M.C.; Dingens, A.S.; Li, J.Z.; Bloom, J.D. Prospective mapping of viral mutations that escape antibodies used to treat COVID-19. Science 2021, 371, 850–854. [Google Scholar] [CrossRef]
- Huang, Y.; Yang, C.; Xu, X.-F.; Xu, W.; Liu, S.-W. Structural and functional properties of SARS-CoV-2 spike protein: Potential antivirus drug development for COVID-19. Acta Pharmacol. Sin. 2020, 41, 1141–1149. [Google Scholar] [CrossRef]
- Yi, C.; Sun, X.; Ye, J.; Ding, L.; Liu, M.; Yang, Z.; Lu, X.; Zhang, Y.; Ma, L.; Gu, W.; et al. Key residues of the receptor binding motif in the spike protein of SARS-CoV-2 that interact with ACE2 and neutralizing antibodies. Cell. Mol. Immunol. 2020, 17, 621–630. [Google Scholar] [CrossRef]
- Lan, J.; Ge, J.; Yu, J.; Shan, S.; Zhou, H.; Fan, S.; Zhang, Q.; Shi, X.; Wang, Q.; Zhang, L.; et al. Structure of the SARS-CoV-2 spike receptor-binding domain bound to the ACE2 receptor. Nature 2020, 581, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Shang, J.; Ye, G.; Shi, K.; Wan, Y.; Luo, C.; Aihara, H.; Geng, Q.; Auerbach, A.; Li, F. Structural basis of receptor recognition by SARS-CoV-2. Nature 2020, 581, 221–224. [Google Scholar] [CrossRef] [Green Version]
- Robbiani, D.F.; Gaebler, C.; Muecksch, F.; Lorenzi, J.C.C.; Wang, Z.; Cho, A.; Agudelo, M.; Barnes, C.O.; Gazumyan, A.; Finkin, S.; et al. Convergent antibody responses to SARS-CoV-2 in convalescent individuals. Nature 2020, 584, 437–442. [Google Scholar] [CrossRef]
- Wang, Q.; Zhang, Y.; Wu, L.; Niu, S.; Song, C.; Zhang, Z.; Lu, G.; Qiao, C.; Hu, Y.; Yuen, K.Y.; et al. Structural and Functional Basis of SARS-CoV-2 Entry by Using Human ACE2. Cell 2020, 181, 894–904.e889. [Google Scholar] [CrossRef]
- Huo, J.; Zhao, Y.; Ren, J.; Zhou, D.; Duyvesteyn, H.M.; Ginn, H.M.; Carrique, L.; Malinauskas, T.; Ruza, R.R.; Shah, P.; et al. Neutralization of SARS-CoV-2 by Destruction of the Prefusion Spike. Cell Host Microbe 2020, 28, 445–454. [Google Scholar] [CrossRef]
- Pinto, D.; Park, Y.-J.; Beltramello, M.; Walls, A.C.; Tortorici, M.A.; Bianchi, S.; Jaconi, S.; Culap, K.; Zatta, F.; De Marco, A.; et al. Cross-neutralization of SARS-CoV-2 by a human monoclonal SARS-CoV antibody. Nature 2020, 583, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Rogers, T.F.; Zhao, F.; Huang, D.; Beutler, N.; Burns, A.; He, W.-T.; Limbo, O.; Smith, C.; Song, G.; Woehl, J.; et al. Isolation of potent SARS-CoV-2 neutralizing antibodies and protection from disease in a small animal model. Science 2020, 369, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Wang, F.; Shen, C.; Peng, W.; Li, D.; Zhao, C.; Li, Z.; Li, S.; Bi, Y.; Yang, Y.; et al. A noncompeting pair of human neutralizing antibodies block COVID-19 virus binding to its receptor ACE2. Science 2020, 368, 1274–1278. [Google Scholar] [CrossRef] [PubMed]
- Farnós, O.; Venereo-Sánchez, A.; Xu, X.; Chan, C.; Dash, S.; Chaabane, H.; Sauvageau, J.; Brahimi, F.; Saragovi, U.; Leclerc, D.; et al. Rapid High-Yield Production of Functional SARS-CoV-2 Receptor Binding Domain by Viral and Non-Viral Transient Expression for Pre-Clinical Evaluation. Vaccines 2020, 8, 654. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Zheng, Q.; Yu, H.; Wu, D.; Xue, W.; Xiong, H.; Huang, X.; Nie, M.; Yue, M.; Rong, R.; et al. SARS-CoV-2 spike produced in insect cells elicits high neutralization titers in non-human primates. Emerg. Microbes Infect. 2020, 9, 2076–2090. [Google Scholar] [CrossRef]
- Yang, J.; Wang, W.; Chen, Z.; Lu, S.; Yang, F.; Bi, Z.; Bao, L.; Mo, F.; Li, X.; Huang, Y.; et al. A vaccine targeting the RBD of the S protein of SARS-CoV-2 induces protective immunity. Nature 2020, 586, 572–577. [Google Scholar] [CrossRef]
- Dalvie, N.C.; Biedermann, A.M.; Rodriguez-Aponte, S.A.; Naranjo, C.A.; Rao, H.D.; Rajurkar, M.P.; Lothe, R.R.; Shaligram, U.S.; Johnston, R.S.; Crowell, L.E.; et al. Scalable, methanol-free manufacturing of the SARS-CoV-2 receptor binding domain in engineered Komagataella phaffii. bioRxiv 2021. [Google Scholar] [CrossRef]
- Zang, J.; Gu, C.; Zhou, B.; Zhang, C.; Yang, Y.; Xu, S.; Bai, L.; Zhang, R.; Deng, Q.; Yuan, Z.; et al. Immunization with the receptor-binding domain of SARS-CoV-2 elicits antibodies cross-neutralizing SARS-CoV-2 and SARS-CoV without antibody-dependent enhancement. Cell Discov. 2020, 6, 61. [Google Scholar] [CrossRef]
- COVID19 Vaccine Tracker. Available online: https://covid19.trackvaccines.org/ (accessed on 13 October 2022).
- Kleanthous, H.; Silverman, J.M.; Makar, K.W.; Yoon, I.-K.; Jackson, N.; Vaughn, D.W. Scientific rationale for developing potent RBD-based vaccines targeting COVID-19. npj Vaccines 2021, 6, 128. [Google Scholar] [CrossRef]
- World Health Organization. Draft Landscape of COVID-19 Candidate Vaccines. 2022. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 13 October 2022).
- COVID-19 Vaccine Market Dashboard. 2022. Available online: https://www.unicef.org/supply/covid-19-vaccine-market-dashboard (accessed on 13 October 2022).
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Zhou, H.; et al. Phase 1–2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef]
- Bangaru, S.; Ozorowski, G.; Turner, H.L.; Antanasijevic, A.; Huang, D.; Wang, X.; Torres, J.L.; Diedrich, J.K.; Tian, J.-H.; Portnoff, A.D.; et al. Structural analysis of full-length SARS-CoV-2 spike protein from an advanced vaccine candidate. Science 2020, 370, 1089–1094. [Google Scholar] [CrossRef]
- Dong, Y.; Dai, T.; Wei, Y.; Zhang, L.; Zheng, M.; Zhou, F. A systematic review of SARS-CoV-2 vaccine candidates. Signal Transduct. Target. Ther. 2020, 5, 237. [Google Scholar] [CrossRef]
- Yadav, T.; Srivastava, N.; Mishra, G.; Dhama, K.; Kumar, S.; Puri, B.; Saxena, S.K. Recombinant vaccines for COVID-19. Hum. Vaccines Immunother. 2020, 16, 2905–2912. [Google Scholar] [CrossRef]
- Dai, L.; Zheng, T.; Xu, K.; Han, Y.; Xu, L.; Huang, E.; An, Y.; Cheng, Y.; Li, S.; Liu, M.; et al. A Universal Design of Betacoronavirus Vaccines against COVID-19, MERS, and SARS. Cell 2020, 182, 722–733.e11. [Google Scholar] [CrossRef]
- Malladi, S.K.; Singh, R.; Pandey, S.; Gayathri, S.; Kanjo, K.; Ahmed, S.; Khan, M.S.; Kalita, P.; Girish, N.; Upadhyaya, A.; et al. Design of a highly thermotolerant, immunogenic SARS-CoV-2 spike fragment. J. Biol. Chem. 2021, 296, 100025. [Google Scholar] [CrossRef]
- Argentinian AntiCovid Consortium. Structural and functional comparison of SARS-CoV-2-spike receptor binding domain produced in Pichia pastoris and mammalian cells. Sci. Rep. 2020, 10, 21779. [Google Scholar] [CrossRef]
- Dalvie, N.C.; Tostanoski, L.H.; Rodriguez-Aponte, S.A.; Kaur, K.; Bajoria, S.; Kumru, O.S.; Martinot, A.J.; Chandrashekar, A.; McMahan, K.; Mercado, N.B.; et al. A modular protein subunit vaccine candidate produced in yeast confers protection against SARS-CoV-2 in non-human primates. bioRxiv 2021. [Google Scholar] [CrossRef]
- Chen, W.-H.; Chag, S.M.; Poongavanam, M.V.; Biter, A.B.; Ewere, E.A.; Rezende, W.; Seid, C.A.; Hudspeth, E.M.; Pollet, J.; McAtee, C.P.; et al. Optimization of the Production Process and Characterization of the Yeast-Expressed SARS-CoV Recombinant Receptor-Binding Domain (RBD219-N1), a SARS Vaccine Candidate. J. Pharm. Sci. 2017, 106, 1961–1970. [Google Scholar] [CrossRef] [Green Version]
- Pollet, J.; Chen, W.-H.; Versteeg, L.; Keegan, B.; Zhan, B.; Wei, J.; Liu, Z.; Lee, J.; Kundu, R.; Adhikari, R.; et al. SARS-CoV-2 RBD219-N1C1: A yeast-expressed SARS-CoV-2 recombinant receptor-binding domain candidate vaccine stimulates virus neutralizing antibodies and T-cell immunity in mice. Hum. Vaccines Immunother. 2021, 17, 2356–2366. [Google Scholar] [CrossRef]
- Chen, W.-H.; Wei, J.; Kundu, R.T.; Adhikari, R.; Liu, Z.; Lee, J.; Versteeg, L.; Poveda, C.; Keegan, B.; Villar, M.J.; et al. Genetic modification to design a stable yeast-expressed recombinant SARS-CoV-2 receptor binding domain as a COVID-19 vaccine candidate. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2021, 1865, 129893. [Google Scholar] [CrossRef]
- Quinlan, B.D.; Mou, H.; Zhang, L.; Guo, Y.; He, W.; Ojha, A.; Parcells, M.S.; Luo, G.; Li, W.; Zhong, G.; et al. The SARS-CoV-2 receptor-binding domain elicits a potent neutralizing response without antibody-dependent enhancement. bioRxiv 2020. [Google Scholar] [CrossRef]
- Limonta, M.; Chinea, G.; Martín, A.; Gonzalez, D.; Bequet, M.; Marquez, G.; González-Moya, I.; Canaan-Haden-Ayala, C.; Cabrales-Rico, A.; Espinosa-Rodríguez, L.A.; et al. The SARS-CoV-2 Receptor-Binding Domain Expressed in Pichia Pastoris as a Candidate Vaccine Antigen. medRxiv 2021. Available online: https://www.medrxiv.org/content/10.1101/2021.06.29.21259605v1.full.pdf (accessed on 9 July 2022).
- Reardon, S. Cuba’s bet on home-grown COVID vaccines is paying off. Nature 2021, 600, 15–16. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Li, Y.; Dai, L.; Wang, J.; He, P.; Li, C.; Fang, X.; Wang, C.; Zhao, X.; Huang, E.; et al. Safety and immunogenicity of a recombinant tandem-repeat dimeric RBD-based protein subunit vaccine (ZF2001) against COVID-19 in adults: Two randomised, double-blind, placebo-controlled, phase 1 and 2 trials. Lancet Infect. Dis. 2021, 21, 1107–1119. [Google Scholar] [CrossRef] [PubMed]
- Novavax: NVX-CoV2373—COVID19 Vaccine Tracker. 2022. Available online: https://covid19.trackvaccines.org/vaccines/25/ (accessed on 13 July 2022).
- Guebre-Xabier, M.; Patel, N.; Tian, J.-H.; Zhou, B.; Maciejewski, S.; Lam, K.; Portnoff, A.D.; Massare, M.J.; Frieman, M.B.; Piedra, P.A.; et al. NVX-CoV2373 vaccine protects cynomolgus macaque upper and lower airways against SARS-CoV-2 challenge. Vaccine 2020, 38, 7892–7896. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.-H.; Patel, N.; Haupt, R.; Zhou, H.; Weston, S.; Hammond, H.; Logue, J.; Portnoff, A.D.; Norton, J.; Guebre-Xabier, M.; et al. SARS-CoV-2 spike glycoprotein vaccine candidate NVX-CoV2373 immunogenicity in baboons and protection in mice. Nat. Commun. 2021, 12, 372. [Google Scholar] [CrossRef]
- Visser, H.; Joosten, V.; Punt, P.J.; Gusakov, A.V.; Olson, P.T.; Joosten, R.; Bartels, J.; Visser, J.; Sinitsyn, A.P.; Emalfarb, M.A.; et al. Development of a mature fungal technology and production platform for industrial enzymes based on a Myceliophthora thermophila isolate, previously known as Chrysosporium lucknowense C1. Ind. Biotechnol. 2011, 7, 214–223. [Google Scholar] [CrossRef]
- Keresztes, G.; Baer, M.; Alfenito, M.R.; Verwoerd, T.C.; Kovalchuk, A.; Wiebe, M.G.; Andersen, T.K.; Saloheimo, M.; Tchelet, R.; Kensinger, R.; et al. The Highly Productive Thermothelomyces heterothallica C1 Expression System as a Host for Rapid Development of Influenza Vaccines. Vaccines 2022, 10, 148. [Google Scholar] [CrossRef]
- Ramot, Y.; Kronfeld, N.; Ophir, Y.; Ezov, N.; Friedman, S.; Saloheimo, M.; Vitikainen, M.; Ben-Artzi, H.; Avigdor, A.; Tchelet, R.; et al. Toxicity and Local Tolerance of a Novel Spike Protein RBD Vaccine Against SARS-CoV-2, Produced Using the C1 Thermothelomyces heterothallica Protein Expression Platform. Toxicol. Pathol. 2022, 50, 294–307. [Google Scholar] [CrossRef]
- Available online: https://www.gurufocus.com/news/1879347/dyadic-receives-regulatory-approval-to-initiate-phase-1-clinical-trial-to-demonstrate-clinical-safety-and-efficacy-in-humans-for-dyai100-covid19-recombinant-protein-rbd-booster-vaccine-candidate.https://www.stocktitan.net/news/DYAI/dyadic-receives-regulatory-approval-to-initiate-phase-1-clinical-b9qggmq4vmp4.html (accessed on 13 October 2022).
- Reed, S.G.; Orr, M.T.; Fox, C.B. Key roles of adjuvants in modern vaccines. Nat. Med. 2013, 19, 1597–1608. [Google Scholar] [CrossRef]
- Hornung, V.; Bauernfeind, F.; Halle, A.; Samstad, E.O.; Kono, H.; Rock, K.L.; Fitzgerald, K.; Latz, E. Silica crystals and aluminum salts activate the NALP3 inflammasome through phagosomal destabilization. Nat. Immunol. 2008, 9, 847–856. [Google Scholar] [CrossRef]
- Matsiko, A. Alum Adjuvant Discovery and Potency. Nature Research. 2020. Available online: https://www.nature.com/articles/d42859-020-00011-w (accessed on 9 July 2022).
- Shirodkar, S.; Hutchinson, R.L.; Perry, D.L.; White, J.L.; Hem, S.L. Aluminum Compounds Used as Adjuvants in Vaccines. Pharm. Res. 1990, 7, 1282–1288. [Google Scholar] [CrossRef]
- Croda Healthcare. Alhydrogel® ‘85’ 2/19 HTCDS113v2 EN. 2021. Available online: https://www.crodahealthcare.com/en-gb/product-finder/product/4373-Alhydrogel (accessed on 9 July 2022).
- Jiang, R.-D.; Liu, M.-Q.; Chen, Y.; Shan, C.; Zhou, Y.-W.; Shen, X.-R.; Li, Q.; Zhang, L.; Zhu, Y.; Si, H.-R.; et al. Pathogenesis of SARS-CoV-2 in Transgenic Mice Expressing Human Angiotensin-Converting Enzyme 2. Cell 2020, 182, 50–58.e8. [Google Scholar] [CrossRef]
- Zheng, J.; Wong, L.-Y.R.; Li, K.; Verma, A.K.; Ortiz, M.E.; Wohlford-Lenane, C.; Leidinger, M.R.; Knudson, C.M.; Meyerholz, D.K.; McCray, P.B., Jr.; et al. COVID-19 treatments and pathogenesis including anosmia in K18-hACE2 mice. Nature 2021, 589, 603–607. [Google Scholar] [CrossRef]
- Oladunni, F.S.; Park, J.-G.; Pino, P.A.; Gonzalez, O.; Akhter, A.; Allué-Guardia, A.; Olmo-Fontánez, A.; Gautam, S.; Garcia-Vilanova, A.; Ye, C.; et al. Lethality of SARS-CoV-2 infection in K18 human angiotensin-converting enzyme 2 transgenic mice. Nat. Commun. 2020, 11, 6122. [Google Scholar] [CrossRef]
- Fornefett, J.; Krause, J.; Klose, K.; Fingas, F.; Hassert, R.; Benga, L.; Grunwald, T.; Müller, U.; Schrödl, W.; Baums, C.G. Comparative analysis of humoral immune responses and pathologies of BALB/c and C57BL/6 wildtype mice experimentally infected with a highly virulent Rodentibacter pneumotropicus (Pasteurella pneumotropica) strain. BMC Microbiol. 2018, 18, 45. [Google Scholar] [CrossRef]
- Lazo, L.; Bequet-Romero, M.; Lemos, G.; Musacchio, A.; Cabrales, A.; Bruno, A.J.; Espinosa, L.A.; Saloheimo, M.; Vitikainen, M.; Hernández, A.; et al. A recombinant SARS-CoV-2 receptor-binding domain expressed in an engineered fungal strain of Thermothelomyces heterothallica induces a functional immune response in mice. Vaccine 2022, 40, 1162–1169. [Google Scholar] [CrossRef]
- Aebischer, A.; Wernike, K.; König, P.; Franzke, K.; Schreur, P.W.; Kortekaas, J.; Vitikainen, M.; Wiebe, M.; Saloheimo, M.; Tchelet, R.; et al. Development of a Modular Vaccine Platform for Multimeric Antigen Display Using an Orthobunyavirus Model. Vaccines 2021, 9, 651. [Google Scholar] [CrossRef]
- Espinosa, L.A.; Ramos, Y.; Andújar, I.; Torres, E.O.; Cabrera, G.; Martín, A.; Roche, D.; Chinea, G.; Becquet, M.; González, I.; et al. In-solution buffer-free digestion allows full-sequence coverage and complete characterization of post-translational modifications of the receptor-binding domain of SARS-CoV-2 in a single ESI–MS spectrum. Anal. Bioanal. Chem. 2021, 413, 7559–7585. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccine | Manufacturer | Production Platform | Antigen/ Adjuvant | Current Approvals |
|---|---|---|---|---|
| ZIFIVAX (ZF2001) | Anhui Zhifei Longcom Biopharmaceutical | CHO K1 cells | RBD-Dimer Alum * | 4 EUA * [30] |
| CIGB-66, ABDALA | Center for Genetic Engineering and Biotechnology | Pichia pastoris | RBD Alum | 4 EUA [30] |
| SOBERANA-02 | Finlay Institute de Vaccines | CHO cells | RBD conjugated to Tetanus Toxoid antigen Alum | 4 EUA [30] |
| SOBERANA PLUS | Finlay Institute de Vaccines | CHO cells | RBD-Dimer conjugated to Tetanus Toxoid antigen Alum | 1 EUA [30] |
| CORBEVAXTM | Biological E Limited | Pichia pastoris | RBD CpG 1018 Alum | 1 EUA [30] |
| NVX-CoV2373 | Novavax | Sf9 insect-cells | Full-Spike Matrix-M1 | EUA [30] 31 countries authorizatios |
| COVOVAX | Serum Institute of India | Sf9 insect-cells | Full-Spike Matrix-M1 | EUA [30] 31 countries authorizations |
| EpiVaCorona Aurora-CoV | Vektor State Research Center of Virology and Biotechnology | Synthesized | Spike protein oligopeptide Alum | 4 EUA [30] |
| EpiVacCorona-N | Vektor State Research Center of Virology and Biotechnology | Synthesized | Spike protein oligopeptide Alum | 1 EUA [30] |
| Noora vaccine | Bagheiat-allah University of Medical Sciences | E. coli BL21 DE3 | RBD Alum | 1 EUA [30] |
| V-01 | Livzon Mabpharm Inc. | CHO cells | RBD-Fc dimer Alum | 1 EUA [30] |
| MVC-COV1901 | Medigen | CHO cells | S-2P Spike CpG 1018 Alum | 1 EUA [30] |
| NVSI-06-08 | National Vaccine and Serum Institute | CHO cells | trimeric RBD Alum | 2 EUA [30] |
| IndoVac | PT Bio Farma | Pichia | RBD CpG 1018 Alum | 1 EUA [30] |
| Razi Cov Pars | Razi Vaccine and Serum Research Institute | Expi293F Cells | Spike RAS-01 | 1 EUA [30] |
| CoV2 preS dTM D614/B.1.351 | Sanofi/GSK | Sf9 insect-cells | spike trimers ASO3 | 30 EUA [30] |
| SKYCovione GBP510 | SK Bioscience Co., Ltd. | Expi293F Cells | RBD Nanoparticles ASO3 | 1 EUA [30] |
| TAK-019 | Takeda | Sf9 insect-cells | Full-Spike Matrix-M1 | 1 EUA [30] |
| SpikoGen, COVAX-19 | Vaxine/CinnaGen Co. | Sf9 insect-cells | Spike Advax-CpG55.2 | 1 EUA [30] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nechooshtan, R.; Ehrlich, S.; Vitikainen, M.; Makovitzki, A.; Dor, E.; Marcus, H.; Hefetz, I.; Pitel, S.; Wiebe, M.; Huuskonen, A.; et al. Thermophilic Filamentous Fungus C1-Cell-Cloned SARS-CoV-2-Spike-RBD-Subunit-Vaccine Adjuvanted with Aldydrogel®85 Protects K18-hACE2 Mice against Lethal Virus Challenge. Vaccines 2022, 10, 2119. https://doi.org/10.3390/vaccines10122119
Nechooshtan R, Ehrlich S, Vitikainen M, Makovitzki A, Dor E, Marcus H, Hefetz I, Pitel S, Wiebe M, Huuskonen A, et al. Thermophilic Filamentous Fungus C1-Cell-Cloned SARS-CoV-2-Spike-RBD-Subunit-Vaccine Adjuvanted with Aldydrogel®85 Protects K18-hACE2 Mice against Lethal Virus Challenge. Vaccines. 2022; 10(12):2119. https://doi.org/10.3390/vaccines10122119
Chicago/Turabian StyleNechooshtan, Ram, Sharon Ehrlich, Marika Vitikainen, Arik Makovitzki, Eyal Dor, Hadar Marcus, Idan Hefetz, Shani Pitel, Marilyn Wiebe, Anne Huuskonen, and et al. 2022. "Thermophilic Filamentous Fungus C1-Cell-Cloned SARS-CoV-2-Spike-RBD-Subunit-Vaccine Adjuvanted with Aldydrogel®85 Protects K18-hACE2 Mice against Lethal Virus Challenge" Vaccines 10, no. 12: 2119. https://doi.org/10.3390/vaccines10122119