
Chitosan-Based Nanomaterial as Immune Adjuvant and Delivery Carrier for Vaccines

Abstract
:1. Introduction
2. Vaccine Adjuvant and Delivery System
2.1. Vaccine Adjuvant
2.2. Vaccine Delivery System
2.2.1. Viral Carrier for the Delivery of Vaccine
2.2.2. Nonviral Carriers for the Delivery of Vaccines
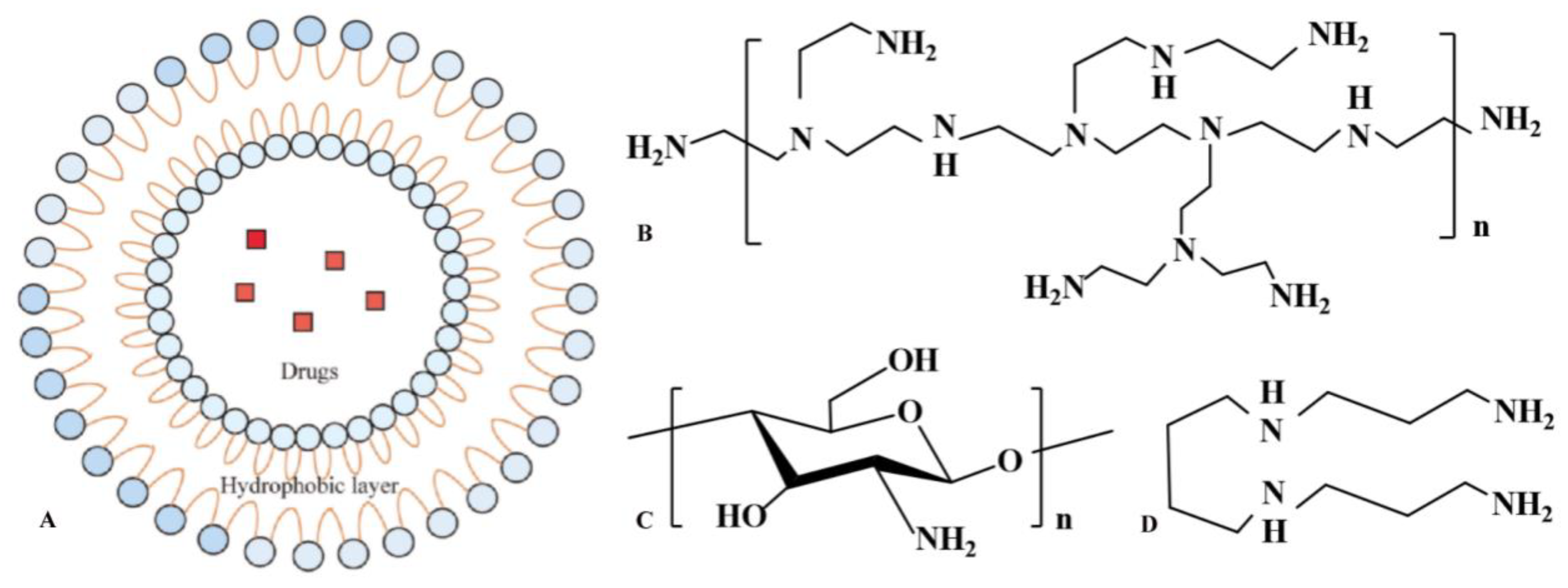
3. Preparation of Chitosan-Based NPs for Vaccine Delivery
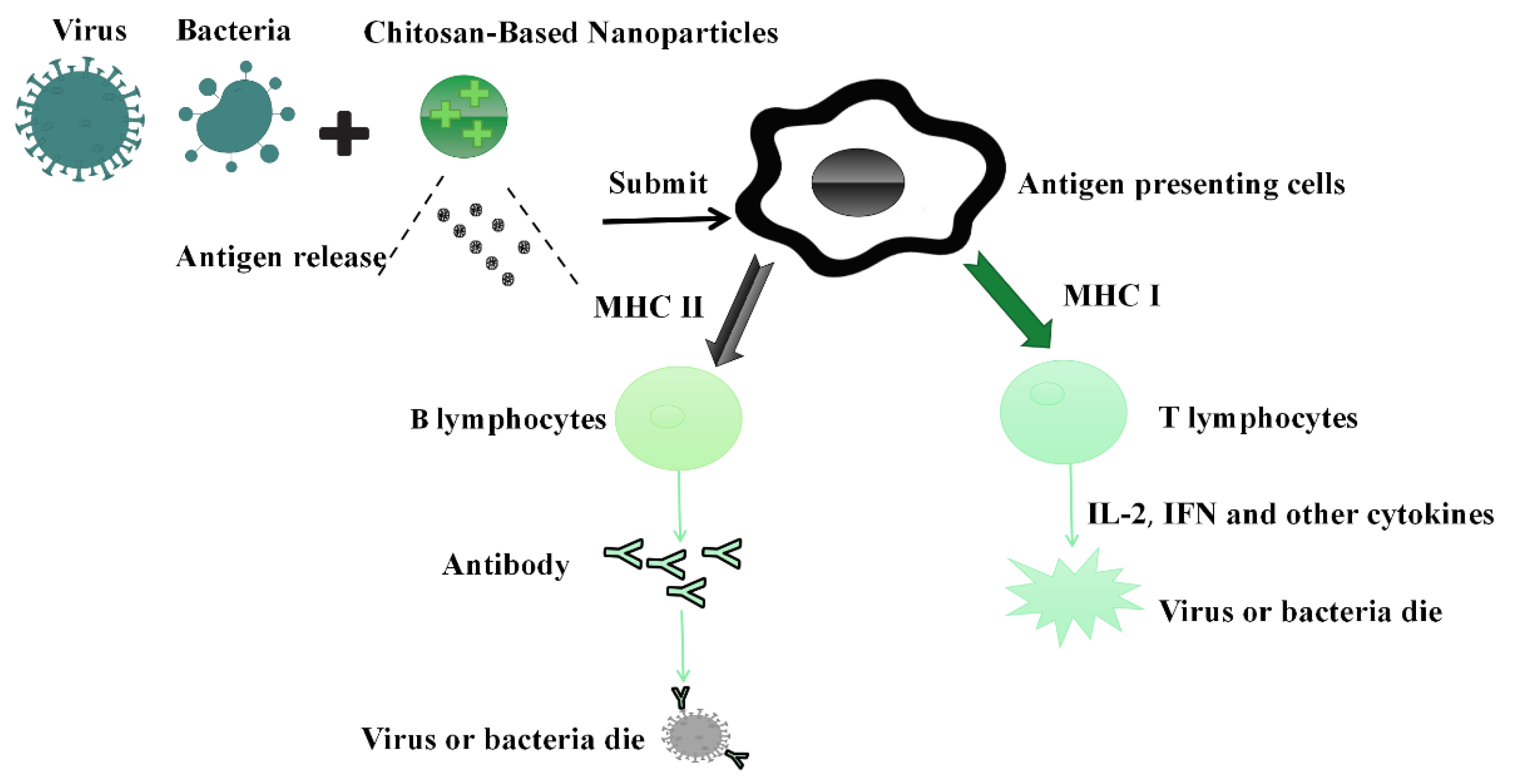
4. Immune Mechanism of Chitosan and Its Nanocomposites as the Vaccine Adjuvants/Delivery System
5. Application of Chitosan and Its Nanocomposites in Vaccine Delivery
5.1. Application in Protein Vaccine Delivery
5.2. Application in Polypeptide Vaccine Delivery
5.3. Application in Nucleic Acid Vaccine Delivery
5.3.1. DNA Vaccines
5.3.2. RNA Vaccines
6. Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wan, X.; Yin, Y.; Zhou, C.; Hou, L.; Cui, Q.; Zhang, X.; Cai, X.; Wang, Y.; Wang, L.; Tian, J. Polysaccharides derived from Chinese medicinal herbs: A promising choice of vaccine adjuvants. Carbohydr. Polym. 2022, 276, 118739. [Google Scholar] [CrossRef]
- Sun, B.; Yu, S.; Zhao, D.; Guo, S.; Wang, X.; Zhao, K. Polysaccharides as vaccine adjuvants. Vaccine 2018, 36, 5226–5234. [Google Scholar] [CrossRef]
- Tlaxca, J.L.; Ellis, S.; Remmele, R.L. Live attenuated and inactivated viral vaccine formulation and nasal delivery: Potential and challenges. Adv. Drug Deliv. Rev. 2015, 93, 56–78. [Google Scholar] [CrossRef]
- Guan, L.J.; Pei, S.X.; Song, J.J.; Zhan, P.F.; Han, Y.N.; Xue, Y.; Ding, K.; Zhao, Z.Q. Screening immune adjuvants for an inactivated vaccine against Erysipelothrix rhusiopathiae. Front Vet. Sci. 2022, 9, 922867. [Google Scholar] [CrossRef]
- Seo, S.U.; Seong, B.L. Prospects on repurposing a live attenuated vaccine for the control of unrelated infections. Front Immunol. 2022, 13, 877845. [Google Scholar] [CrossRef]
- Van Herck, S.; Feng, B.; Tang, L. Delivery of sting agonists for adjuvanting subunit vaccines. Adv. Drug Deliv. Rev. 2021, 179, 114020. [Google Scholar] [CrossRef]
- Bose, R.J.; Kim, M.; Chang, J.H.; Paulmurugan, R.; Moon, J.J.; Koh, W.-G.; Lee, S.-H.; Park, H. Biodegradable polymers for modern vaccine development. J. Ind. Eng. Chem. 2019, 77, 12–24. [Google Scholar] [CrossRef]
- Pirahmadi, S.; Afzali, S.; Zargar, M.; Zakeri, S.; Mehrizi, A.A. How can we develop an effective subunit vaccine to achieve successful malaria eradication? Microb. Pathog. 2021, 160, 105203. [Google Scholar] [CrossRef]
- Mohebbi, A.; Ebrahimzadeh, M.S.; Baghban-Rahimi, S.; Saeidi, M.; Tabarraei, A.; Mohebbi, S.R.; Shirian, S.; Gorji, A.; Ghaemi, A. Non-replicating newcastle disease virus as an adjuvant for DNA vaccine enhances antitumor efficacy through the induction of TRAIL and granzyme B expression. Virus Res. 2019, 261, 72–80. [Google Scholar] [CrossRef]
- Gumina, E.; Hall, J.W.; Vecchi, B.; Hernandez-Velasco, X.; Lumpkins, B.; Mathis, G.; Layton, S. Evaluation of a subunit vaccine candidate (Biotech Vac Cox) against Eimeria spp. in broiler chickens. Poult. Sci. 2021, 100, 101329. [Google Scholar] [CrossRef]
- Eusébio, D.; Neves, A.R.; Costa, D.; Biswas, S.; Sousa, N. Methods to improve the immunogenicity of plasmid DNA vaccines. Drug Discov. Today. 2021, 26, 2575–2592. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, Y.; Takada, Y.; Kotake, M.; Zenke, K.; Itoh, N.; Yoshinaga, T. Evaluation of the protective effects of DNA vaccines encoding an infection-related cysteine protease from Cryptocaryon irritans, the pathogenic organism of cryptocaryoniasis. Aquaculture 2021, 548, 737641. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhang, X.; Li, Q.; Fu, L.; Wang, M.; Liu, S.; Wu, J.; Nie, J.; Zhang, L.; Zhao, C.; et al. Unmethylated CpG motif-containing genomic DNA fragments of Bacillus Calmette-guerin improves immune response towards a DNA vaccine for COVID-19. Vaccine. 2021, 39, 6050–6056. [Google Scholar] [CrossRef]
- Gerhardt, A.; Voigt, E.; Archer, M.; Reed, S.; Larson, E.; Van Hoeven, N.; Kramer, R.; Fox, C.; Casper, C. A flexible, thermostable nanostructured lipid carrier platform for RNA vaccine delivery. Mol. Ther. Methods Clin. Dev. 2022, 25, 205–214. [Google Scholar] [CrossRef]
- He, Q.; Gao, H.; Tan, D.; Zhang, H.; Wang, J.-Z. mRNA cancer vaccines: Advances, trends and challenges. Acta Pharm. Sin. B 2022, 12, 107020. [Google Scholar] [CrossRef]
- Maruggi, G.; Chiarot, E.; Giovani, C.; Buccato, S.; Bonacci, S.; Frigimelica, E.; Margarit, I.; Geall, A.; Bensi, G.; Maione, D. Immunogenicity and protective efficacy induced by self-amplifying mRNA vaccines encoding bacterial antigens. Vaccine 2017, 35, 361–368. [Google Scholar] [CrossRef]
- Park, J.W.; Lagniton, P.N.; Liu, Y.; Xu, R.-H. mRNA vaccines for COVID-19: What, why and how. Int. J. Biol. Sci. 2021, 17, 1446–1460. [Google Scholar] [CrossRef]
- Zafar, S.; Arshad, M.S.; Fatima, S.; Ali, A.; Zaman, A.; Sayed, E.; Chang, M.W.; Ahmad, Z. COVID-19: Current developments and further opportunities in drug delivery and therapeutics. Pharmaceutics 2020, 12, 945. [Google Scholar] [CrossRef]
- Kim, H.J.; Seo, S.K.; Park, H.Y. Physical and chemical advances of synthetic delivery vehicles to enhance mRNA vaccine efficacy. J. Control. Release. 2022, 345, 405–416. [Google Scholar] [CrossRef]
- Li, D.; Fu, D.; Kang, H.; Rong, G.; Jin, Z.; Wang, X.; Zhao, K. Advances and potential applications of chitosan nanoparticles as a delivery carrier for the mucosal immunity of vaccine. Curr. Drug Deliv. 2017, 14, 27–35. [Google Scholar] [CrossRef]
- Bento, D.; Jesus, S.; Lebre, F.; Gonçalves, T.; Borges, O. Chitosan plus compound 48/80: Formulation and preliminary evaluation as a hepatitis B vaccine adjuvant. Pharmaceutics. 2019, 11, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malik, A.; Gupta, M.; Gupta, V.; Gogoi, H.; Bhatnagar, R. Novel application of trimethyl chitosan as an adjuvant in vaccine delivery. Int. J. Nanomed. 2018, 13, 7959–7970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dmour, I.; Islam, N. Recent advances on chitosan as an adjuvant for vaccine delivery. Int. J. Biol. Macromol. 2022, 200, 498–519. [Google Scholar] [CrossRef]
- Paula, H.; Silva, R.; Santos, C.M.; Dantas, F.; de Paula, R.; de Lima, L.; de Oliveira, E.; Figueiredo, E.; Dias, F. Eco-friendly synthesis of an alkyl chitosan derivative. Int. J. Biol. Macromol. 2020, 163, 1591–1598. [Google Scholar] [CrossRef]
- Wang, W.; Meng, Q.; Li, Q.; Liu, J.; Zhou, M.; Jin, Z.; Zhao, K. Chitosan derivatives and yheir application in biomedicine. Int. J. Mol. Sci. 2020, 21, 487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Xing, R.; Xu, C.; Liu, S.; Qin, Y.; Li, K.; Yu, H.; Li, P. Immunostimulatory effect of chitosan and quaternary chitosan: A review of potential vaccine adjuvants. Carbohydr. Polym. 2021, 264, 118050. [Google Scholar] [CrossRef]
- Das, S.S.; Kar, S.; Singh, S.K.; Verma, P.; Hussain, A.; Beg, S. Chitosan-based systems for oral drug delivery applications. In Chitosan in Drug Delivery; Elsevier: India, 2021; pp. 23–53. [Google Scholar]
- Song, F.; Kong, Y.; Shao, C.; Cheng, Y.; Lu, J.; Tao, Y.; Du, J.; Wang, H. Chitosan-based multifunctional flexible hemostatic bio-hydrogel. Acta Biomater. 2021, 136, 170–183. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, C.; Li, C.; Qin, Y.; Wang, Z.; Yang, F.; Li, Z.; Wang, J. A functional chitosan-based hydrogel as a wound dressing and drug delivery system in the treatment of wound healing. RSC Adv. 2018, 8, 7533–7549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, J.; Li, J.; Jiang, Z.; Tong, R.; Duan, X.; Bai, L.; Shi, J. Chitosan, N,N,N-trimethyl chitosan (TMC) and 2-hydroxypropyltrimethyl ammonium chloride chitosan (HTCC): The potential immune adjuvants and nano carriers. Int. J. Biol. Macromol. 2020, 154, 339–348. [Google Scholar] [CrossRef]
- Norpi, A.; Nordin, M.L.; Ahmad, N.; Katas, H.; Fuaad, A.; Sukri, A.; Marasini, N.; Azmi, F. New modular platform based on multi-adjuvanted amphiphilic chitosan nanoparticles for efficient lipopeptide vaccine delivery against group A streptococcus. Asian J. Pharm. Sci. 2022, 17, 435–446. [Google Scholar] [CrossRef]
- Şenel, S. Current status and future of chitosan in drug and vaccine delivery. React. Funct. Polym. 2020, 147, 104452. [Google Scholar] [CrossRef]
- Yang, Y.; Xing, R.; Liu, S.; Qin, Y.; Li, K.; Yu, H.; Li, P. Chitosan, hydroxypropyltrimethyl ammonium chloride chitosan and sulfated chitosan nanoparticles as adjuvants for inactivated newcastle disease vaccine. Carbohydr. Polym. 2020, 229, 115423. [Google Scholar] [CrossRef] [PubMed]
- Rizeq, B.R.; Younes, N.N.; Rasool, K.; Nasrallah, G.K. Synthesis, bioapplications, and toxicity evaluation of chitosan-based nanoparticles. Int. J. Mol. Sci. 2019, 20, 5776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrabi, M.; Montazeri, H.; Mohamadpour Dounighi, N.; Rashti, A.; Vakili-Ghartavol, R. Chitosan based nanoparticles in mucosal vaccine delivery. Arch. Razi. Inst. 2018, 73, 165–176. [Google Scholar] [CrossRef]
- Quiñones, J.P.; Peniche, H.; Peniche, C. Chitosan based self-assembled nanoparticles in drug delivery. Polym. Basel 2018, 10, 235. [Google Scholar] [CrossRef] [Green Version]
- Samah, H.M.; Abdel, S.A.; Satar, A.; Wesam, H.M.; Hanan, A.F.; Lamiaa, M.O.; Rania, E.M. Preparation and immunological evaluation of inactivated avian influenza virus vaccine encapsulated in chitosan nanoparticles. Biologicals 2018, 51, 46–53. [Google Scholar] [CrossRef]
- Jin, Z.; Li, D.; Dai, C.; Cheng, G.; Wang, X.; Zhao, K. Response of live newcastle disease virus encapsulated in N -2-hydroxypropyl dimethylethyl ammonium chloride chitosan nanoparticles. Carbohydr. Polym. 2017, 171, 267–280. [Google Scholar] [CrossRef]
- Zhao, K.; Gao, Y.; Hu, G.; Wang, L.; Cui, S.; Jin, Z. N-2-Hydroxypropyl Trimethyl ammonium chloride chitosan as adjuvant enhances the immunogenicity of a VP2 subunit vaccine against porcine parvovirus infection in sows. Vaccines Basel 2021, 9, 1027. [Google Scholar] [CrossRef]
- Zhao, K.; Han, J.; Zhang, Y.; Wei, L.; Yu, S.; Wang, X.; Jin, Z.; Wang, Y. Enhancing mucosal immune response of newcastle disease virus DNA vaccine using N-2-Hydroxypropyl Trimethylammonium chloride chitosan and N,O-Carboxymethyl chitosan nanoparticles as delivery carrier. Mol. Pharm. 2018, 15, 226–237. [Google Scholar] [CrossRef]
- Zhao, K.; Li, S.; Li, W.; Yu, L.; Duan, X.; Han, J.; Wang, X.; Jin, Z. Quaternized chitosan nanoparticles loaded with the combined attenuated live vaccine against newcastle disease and infectious bronchitis elicit immune response in chicken after intranasal administration. Drug Deliv. 2017, 24, 1574–1586. [Google Scholar] [CrossRef]
- Jesus, S.; Soares, E.; Borchard, G.; Borges, O. Poly-ϵ-caprolactone/chitosan nanoparticles provide strong adjuvant effect for hepatitis B antigen. Nanomed. Lond 2017, 12, 2335–2348. [Google Scholar] [CrossRef] [PubMed]
- Hoseinpur, R.; Hasani, A.; Baradaran, B.; Abdolalizadeh, J.; Amini, Y.; Salehi, R.; Kafil, H.S.; Azizian, K.; Memar, M.Y.; Gholizadeh, P.; et al. Chitosan nanoparticles containing fusion protein (Hspx–PPE44–EsxV) and resiquimod adjuvant (HPERC) as a novel booster vaccine for Mycobacterium tuberculosis. J. Biomater. Appl. 2022, 37, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Waeckerle-Men, Y.; Groettrup, M. PLGA microspheres for improved antigen delivery to dendritic cells as cellular vaccines. Adv. Drug Deliv. Rev. 2005, 57, 475–482. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Zou, Y.; Hu, Z. Advances in aluminum hydroxide-based adjuvant research and its mechanism. Hum. Vaccines Immunother. 2015, 11, 477–488. [Google Scholar] [CrossRef] [PubMed]
- Korkmaz, E.; Balmert, S.C.; Carey, C.D.; Erdos, G.; Falo, L.D., Jr. Emerging skin-targeted drug delivery strategies to engineer immunity: A focus on infectious diseases. Expert Opin. Drug Deliv. 2021, 18, 151–167. [Google Scholar] [CrossRef]
- Jin, Z.; Gao, S.; Cui, X.; Sun, D.; Zhao, K. Adjuvants and delivery systems based on polymeric nanoparticles for mucosal vaccines. Int. J. Pharm. 2019, 572, 118731. [Google Scholar] [CrossRef]
- Li, H.; Ren, H.; Zhou, Y.; Zhang, Y.; Cao, L.; Xu, W. HRSV prefusion-F protein with Adju-Phos adjuvant induces long-lasting Th2-biased immunity in mice. PLoS ONE 2022, 17, e0262231. [Google Scholar] [CrossRef]
- Lin, Y.; Wang, X.; Huang, X.; Zhang, J.; Xia, N.; Zhao, Q. Calcium phosphate nanoparticles as a new generation vaccine adjuvant. Expert Rev. Vaccines. 2017, 16, 895–906. [Google Scholar] [CrossRef]
- Song, T.; Liao, Y.; Zuo, Q.; Liu, N.; Liu, Z. MnO2 nanoparticles as a minimalist multimode vaccine adjuvant/delivery system to regulate antigen presenting cells for tumor immunotherapy. J. Mater. Chem. B 2022, 10, 3474–3490. [Google Scholar] [CrossRef]
- Ko, E.-J.; Kang, S.-M. Immunology and efficacy of MF59-adjuvanted vaccines. Hum. Vaccines Immunother. 2018, 14, 3041–3045. [Google Scholar] [CrossRef]
- Rastakhiz, S.; Yazdani, M.; Shariat, S.; Arab, A.; Momtazi-Borojeni, A.A.; Barati, N.; Mansourian, M.; Amin, M.; Abbasi, A.; Saberi, Z.; et al. Preparation of nanoliposomes linked to HER2/neu-derived (P5) peptide containing MPL adjuvant as vaccine against breast cancer. J. Cell. Biochem. 2018, 120, 1294–1303. [Google Scholar] [CrossRef] [PubMed]
- Carter, D.; van Hoeven, N.; Baldwin, S.; Levin, Y.; Kochba, E.; Magill, A.; Charland, N.; Landry, N.; Nu, K.; Frevol, A.; et al. The adjuvant GLA-AF enhances human intradermal vaccine responses. Sci. Adv. 2018, 4, eaas9930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kazzaz, J.; Singh, M.; Ugozzoli, M.; Chesko, J.; Soenawan, E.; O’Hagan, D.T. Encapsulation of the immune potentiators MPL and RC529 in PLG microparticles enhances their potency. J. Control. Release. 2006, 110, 566–573. [Google Scholar] [CrossRef] [PubMed]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev. Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clements, J.D.; Norton, E.B. The mucosal vaccine adjuvant LT(R192G/L211A) or dmLT. mSphere. 2018, 3, e00215–e00218. [Google Scholar] [CrossRef] [Green Version]
- Qu, W.; Li, N.; Yu, R.; Zuo, W.; Fu, T.; Fei, W.; Hou, Y.; Liu, Y.; Yang, J. Cationic DDA/TDB liposome as a mucosal vaccine adjuvant for uptake by dendritic cells in vitro induces potent humoural immunity. Artif. Cells Nanomed. Biotechnol. 2018, 46 (Suppl. 1), 852–860. [Google Scholar] [CrossRef] [Green Version]
- Mohanraj, M.; Sekar, P.; Liou, H.-H.; Chang, S.-F.; Lin, W.-W. The mycobacterial adjuvant analogue TDB attenuates neuroinflammation via mincle-independent PLC-γ1/PKC/ERK signaling and microglial polarization. Mol. Neurobiol. 2019, 56, 1167–1187. [Google Scholar] [CrossRef]
- Cui, B.; Liu, X.; Fang, Y.; Zhou, P.; Zhang, Y.; Wang, Y. Flagellin as a vaccine adjuvant. Expert Rev. Vaccines 2018, 17, 335–349. [Google Scholar] [CrossRef]
- Hafner, A.M.; Corthésy, B.; Merkle, H.P. Particulate formulations for the delivery of poly(I:C) as vaccine adjuvant. Adv. Drug Deliv. Rev. 2013, 65, 1386–1399. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Laupeze, B.; Di Pasquale, A.; Hergli, N.; Collignon, C.; Garçon, N. Adjuvant system AS01: Helping to overcome the challenges of modern vaccines. Expert Rev. Vaccines. 2017, 16, 55–63. [Google Scholar] [CrossRef]
- Murugappan, S.; Frijlink, H.W.; Petrovsky, N.; Hinrichs, W.L. Enhanced pulmonary immunization with aerosolized inactivated influenza vaccine containing delta inulin adjuvant. Eur. J. Pharm. Sci. 2015, 66, 118–122. [Google Scholar] [CrossRef] [PubMed]
- Rizza, P.; Capone, I.; Moretti, F.; Proietti, E.; Belardelli, F. IFN-α as a vaccine adjuvant: Recent insights into the mechanisms and perspectives for its clinical use. Expert Rev. Vaccines. 2011, 10, 487–498. [Google Scholar] [CrossRef] [PubMed]
- Sciullo, P.D.; Menay, F.; Cocozza, F.; Gravisaco, M.J.; Waldner, C.I.; Mongini, C. Systemic administration of imiquimod as an adjuvant improves immunogenicity of a tumor-lysate vaccine inducing the rejection of a highly aggressive T-cell lymphoma. Clin. Immunol. 2019, 203, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Clemens, E.A.; Holbrook, B.C.; Kanekiyo, M.; Yewdell, J.W.; Graham, B.S.; Alexander-Miller, M.A. An R848-conjugated influenza virus vaccine elicits robust immunoglobulin G to hemagglutinin stem in a newborn nonhuman primate model. J. Infect. Dis. 2021, 224, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Garçon, N.; Morel, S.; Didierlaurent, A.; Descamps, D.; Wettendorff, M.; Van Mechelen, M. Development of an AS04-adjuvanted HPV vaccine with the adjuvant system approach. BioDrugs. 2011, 25, 217–226. [Google Scholar] [CrossRef]
- Zheng, Y.; Bian, L.; Zhao, H.; Liu, Y.; Lu, J.; Liu, D.; Zhang, K.; Song, Y.; Luo, Y.; Jiang, C.; et al. Respiratory syncytial virus F subunit vaccine with AS02 adjuvant elicits balanced, robust humoral and cellular immunity in BALB/c Mice. Front. Immunol. 2020, 11, 526965. [Google Scholar] [CrossRef]
- Didierlaurent, A.M.; Collignon, C.; Bourguignon, P.; Wouters, S.; Fierens, K.; Fochesato, M.; Dendouga, N.; Langlet, C.; Malissen, B.; Lambrecht, B.N.; et al. Enhancement of adaptive immunity by the human vaccine adjuvant AS01 depends on activated dendritic cells. J. Immunol. 2014, 193, 1920–1930. [Google Scholar] [CrossRef] [Green Version]
- Carroll, E.C.; Jin, L.; Mori, A.; Muñoz-Wolf, N.; Oleszycka, E.; Moran, H.B.; Mansouri, S.; McEntee, C.; Lambe, E.; Agger, E.M.; et al. The vaccine adjuvant chitosan promotes cellular immunity via DNA sensor cGAS-STING-dependent induction of Type I interferons. Immunity. 2016, 44, 597–608. [Google Scholar] [CrossRef] [Green Version]
- Mallakpour, S.; Azadi, E.; Hussain, C.M. Chitosan, alginate, hyaluronic acid, gums, and β-glucan as potent adjuvants and vaccine delivery systems for viral threats including SARS-CoV-2: A review. Int. J. Biol. Macromol. 2021, 182, 1931–1940. [Google Scholar] [CrossRef]
- He, J.; Liu, Z.; Jiang, W.; Zhu, T.; Wusiman, A.; Gu, P.; Liu, J.; Wang, D. Immune-adjuvant activity of lentinan-modified calcium carbonate microparticles on a H5N1 vaccine. Int. J. Biol. Macromol. 2020, 163, 1384–1392. [Google Scholar] [CrossRef]
- Ediriweera, G.R.; Chen, L.; Yerbury, J.J.; Thurecht, K.J.; Vine, K.L. Non-viral vector-mediated gene therapy for ALS: Challenges and future perspectives. Mol. Pharm. 2021, 18, 2142–2160. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Zhu, X.; Yi, H.; Gu, J.; Liu, S.; Izenwasser, S.; Lemmon, V.P.; Roy, S.; Hao, S. Viral vector-mediated gene therapy for opioid use disorders. Exp. Neurol. 2021, 341, 113710. [Google Scholar] [CrossRef] [PubMed]
- Abdala, A.; Alvarez, I.; Brossel, H.; Calvinho, L.; Carignano, H.; Franco, L.; Gazon, H.; Gillissen, C.; Hamaidia, M.; Hoyos, C.; et al. BLV: Lessons on vaccine development. Retrovirology. 2019, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.A.; Stanfield, B.A.; Chouljenko, V.N.; Naidu, S.; Langohr, I.; Del Piero, F.; Ferracone, J.; Roy, A.A.; Kousoulas, K.G. Intramuscular immunization of mice with the live-attenuated herpes simplex virus 1 vaccine strain VC2 expressing equine herpesvirus 1 (EHV-1) glycoprotein D generates anti-EHV-1 immune responses in mice. J. Virol. 2017, 91, e02445-16. [Google Scholar] [CrossRef] [Green Version]
- Santana-Armas, M.L.; de Ilarduya, C.T. Strategies for cancer gene-delivery improvement by non-viral vectors. Int. J. Pharm. 2021, 596, 120291. [Google Scholar] [CrossRef]
- Sharma, O.; Sultan, A.A.; Ding, H.; Triggle, C.R. A review of the progress and challenges of developing a vaccine for COVID-19. Front. Immunol. 2020, 11, 585354. [Google Scholar] [CrossRef]
- Ura, T.; Yamashita, A.; Mizuki, N.; Okuda, K.; Shimada, M. New vaccine production platforms used in developing SARS-CoV-2 vaccine candidates. Vaccine. 2021, 39, 197–201. [Google Scholar] [CrossRef]
- Khan, W.H.; Hashmi, Z.; Goel, A.; Ahmad, R.; Gupta, K.; Khan, N.; Alam, I.; Ahmed, F.; Ansari, M.A. COVID-19 pandemic and vaccines update on challenges and resolutions. Front. Cell. Infect. Microbiol. 2021, 11, 690621. [Google Scholar] [CrossRef]
- Boroumand, H.; Badie, F.; Mazaheri, S.; Seyedi, Z.S.; Nahand, J.S.; Nejati, M.; Baghi, H.B.; Abbasi-Kolli, M.; Badehnoosh, B.; Ghandali, M.; et al. Chitosan-based nanoparticles against viral infections. Front. Cell. Infect. Microbiol. 2021, 11, 643953. [Google Scholar] [CrossRef]
- Segura, M.M.; Mangion, M.; Gaillet, B.; Garnier, A. New developments in lentiviral vector design, production and purification. Expert Opin. Biol. Ther. 2013, 13, 987–1011. [Google Scholar] [CrossRef]
- Luis, A. The Old and the New: Prospects for non-Integrating lentiviral vector technology. Viruses 2020, 12, 1103. [Google Scholar] [CrossRef]
- Humphreys, I.R.; Sebastian, S. Novel viral vectors in infectious diseases. Immunology 2018, 153, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Kasala, D.; Yoon, A.-R.; Hong, J.; Kim, S.W.; Yun, C.-O. Evolving lessons on nanomaterial-coated viral vectors for local and systemic gene therapy. Nanomed. Lond. 2016, 11, 1689–1713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kochhar, S.; Excler, J.-L.; Bok, K.; Gurwith, M.; McNeil, M.M.; Seligman, S.J.; Khuri-Bulos, N.; Klug, B.; Laderoute, M.; Robertson, J.S.; et al. Defining the interval for monitoring potential adverse events following immunization (AEFIs) after receipt of live viral vectored vaccines. Vaccine. 2019, 37, 5796–5802. [Google Scholar] [CrossRef] [PubMed]
- Miller, A.D. Retroviral vectors: From cancer viruses to therapeutic tools. Hum. Gene Ther. 2014, 25, 989–994. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, V.K.; Kumar, S.; Dhaked, R.K.; Ansari, A.S.; Lohiya, N.K.; Tapryal, S. Generation of oligomers of subunit vaccine candidate glycoprotein D of Herpes Simplex Virus-2 expressed in fusion with IgM Fc domain(s) in Escherichia coli: A strategy to enhance the immunogenicity of the antigen. 3 Biotech. 2020, 10, 463. [Google Scholar] [CrossRef]
- Chentoufi, A.A.; Dhanushkodi, N.R.; Srivastava, R.; Prakash, S.; Coulon, P.A.; Zayou, L.; Vahed, H.; Chentoufi, H.A.; Hormi-Carver, K.K.; BenMohamed, L. Combinatorial herpes simplex vaccine strategies: From bedside to bench and back. Front Immunol. 2022, 13, 849515. [Google Scholar] [CrossRef]
- Sakurai, F.; Tachibana, M.; Mizuguchi, H. Adenovirus vector-based vaccine for infectious diseases. Drug Metab. Pharmacokinet. 2022, 42, 100432. [Google Scholar] [CrossRef]
- Mitani, K.; Kubo, S. Adenovirus as an integrating vector. Curr. Gene Ther. 2002, 2, 135–144. [Google Scholar] [CrossRef]
- Shimizu, K.; Sakurai, F.; Iizuka, S.; Ono, R.; Tsukamoto, T.; Nishimae, F.; Nakamura, S.-I.; Nishinaka, T.; Terada, T.; Fujio, Y.; et al. Adenovirus vector–induced IL-6 promotes leaky adenoviral gene expression, leading to acute hepatotoxicity. J. Immunol. 2021, 206, 410–421. [Google Scholar] [CrossRef]
- Milone, M.C.; O’Doherty, U. Clinical use of lentiviral vectors. Leukemia. 2018, 32, 1529–1541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morante, V.; Borghi, M.; Farina, I.; Michelini, Z.; Grasso, F.; Gallinaro, A.; Cecchetti, S.; Di Virgilio, A.; Canitano, A.; Pirillo, M.; et al. Integrase-defective lentiviral vector is an efficient vaccine platform for cancer immunotherapy. Viruses. 2021, 13, 355. [Google Scholar] [CrossRef] [PubMed]
- Ndwandwe, D.; Wiysonge, C.S. COVID-19 vaccines. Curr. Opin. Immunol. 2021, 71, 111–116. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.Y.; Cho, M.-H.; Kim, S. Recent advances in aerosol gene delivery systems using non-viral vectors for lung cancer therapy. Expert Opin. Drug Deliv. 2019, 16, 757–772. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Li, T.; Zhang, J.; Zhao, L.; Zhai, G. Advances in liver-directed gene therapy for hepatocellular carcinoma by non-viral delivery systems. Curr. Gene Ther. 2012, 12, 92–102. [Google Scholar] [CrossRef]
- Zhao, H.; Wang, H.; Hu, Y.; Xu, D.; Yin, C.; Han, Q.; Zhang, J. Chitosan nanovaccines as efficient carrier adjuvant system for IL-12 with enhanced protection against HBV. Int. J. Nanomed. 2021, 16, 4913–4928. [Google Scholar] [CrossRef]
- Chuang, C.-C.; Tsai, M.-H.; Yen, H.-J.; Shyu, H.-F.; Cheng, K.-M.; Chen, X.-A.; Chen, C.-C.; Young, J.-J.; Kau, J.-H. A fucoidan-quaternary chitosan nanoparticle adjuvant for anthrax vaccine as an alternative to CpG oligodeoxynucleotides. Carbohydr. Polym. 2020, 229, 115403. [Google Scholar] [CrossRef]
- Yüksel, S.; Pekcan, M.; Puralı, N.; Esendağlı, G.; Tavukçuoğlu, E.; Rivero-Arredondo, V.; Ontiveros-Padilla, L.; López-Macías, C.; Şenel, S. Development and in vitro evaluation of a new adjuvant system containing Salmonella Typhi porins and chitosan. Int. J. Pharm. 2020, 578, 119129. [Google Scholar] [CrossRef]
- Lebre, F.; de Lima, M.P.; Lavelle, E.; Borges, O. Mechanistic study of the adjuvant effect of chitosan-aluminum nanoparticles. Int. J. Pharm. 2018, 552, 7–15. [Google Scholar] [CrossRef]
- Rabiee, N.; Bagherzadeh, M.; Ghadiri, A.M.; Kiani, M.; Ahmadi, S.; Jajarmi, V.; Fatahi, Y.; Aldhaher, A.; Tahriri, M.; Webster, T.J.; et al. Calcium-based nanomaterials and their interrelation with chitosan: Optimization for pCRISPR delivery. J. Nanostructure Chem. 2022, 12, 919–932. [Google Scholar] [CrossRef]
- Haryono, A.; Salsabila, K.; Restu, W.K.; Harmami, S.B.; Safari, D. Effect of chitosan and liposome nanoparticles as adjuvant codelivery on the immunoglobulin G subclass distribution in a mouse model. J. Immunol. Res. 2017, 2017, 9125048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van der Lubben, I.M.; Verhoef, J.; Borchard, G.; Junginger, H.E. Chitosan and its derivatives in mucosal drug and vaccine delivery. Eur. J. Pharm. Sci. 2001, 14, 201–207. [Google Scholar] [CrossRef]
- Wu, X.; Farooq, M.A.; Li, T.; Geng, T.; Kutoka, P.T.; Wang, B. Cationic chitosan-modified silica nanoparticles for oral delivery of protein vaccine. J. Biomed. Mater. Res. Part A 2021, 109, 2111–2119. [Google Scholar] [CrossRef] [PubMed]
- Tian, J.; Yu, J.; Sun, X. Chitosan microspheres as candidate plasmid vaccine carrier for oral immunisation of Japanese flounder (Paralichthys olivaceus). Vet. Immunol. Immunopathol. 2008, 126, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Pei, M.; Liang, J.; Zhang, C.; Wang, X.; Zhang, C.; Ma, G.; Sun, H. Chitosan/calcium phosphates nanosheet as a vaccine carrier for effective cross-presentation of exogenous antigens. Carbohydr. Polym. 2019, 224, 115172. [Google Scholar] [CrossRef]
- Cai, J.; Zhang, W.; Xu, J.; Xue, W.; Liu, Z. Evaluation of N-phosphonium chitosan as a novel vaccine carrier for intramuscular immunization. J. Biomater. Appl. 2017, 32, 677–685. [Google Scholar] [CrossRef]
- Li, Y.; Wang, C.; Sun, Z.; Xiao, J.; Yan, X.; Chen, Y.; Yu, J.; Wu, Y. Simultaneous intramuscular and intranasal administration of chitosan nanoparticles-adjuvanted chlamydia vaccine elicits elevated protective responses in the lung. Int. J. Nanomed. 2019, 14, 8179–8193. [Google Scholar] [CrossRef] [Green Version]
- Boulbria, G.; Brilland, S.; Teixeira-Costa, C.; Brissonnier, M.; Charles, M.; Capdevielle, N.; Normand, V.; Bouchet, F.; Berton, P.; Krejci, R.; et al. Effectiveness of two intramuscular combined vaccines for the control of Mycoplasma hyopneu-moniae and porcine circovirus type 2 in growing pigs: A randomized field trial. Porc. Health Manag. 2021, 7, 41. [Google Scholar] [CrossRef]
- Peachman, K.K.; Rao, M.; Alving, C.R.; Burge, R.; Leppla, S.H.; Rao, V.B.; Matyas, G.R. Correlation between lethal toxin-neutralizing antibody titers and protection from intranasal challenge with Bacillus Anthracis Ames strain spores in mice after transcutaneous immunization with recombinant anthrax protective antigen. Infect. Immun. 2006, 74, 794–797. [Google Scholar] [CrossRef] [Green Version]
- Thakur, A.; Pinto, F.E.; Hansen, H.S.; Andersen, P.; Christensen, D.; Janfelt, C.; Foged, C. Intrapulmonary (i.pulmon.) pull immunization with the tuberculosis subunit vaccine candidate H56/CAF01 after intramuscular (i.m.) priming elicits a distinct innate myeloid response and activation of antigen-presenting cells than i.m. or i.pulmon. prime immunization alone. Front. Immunol. 2020, 11, 803. [Google Scholar] [CrossRef]
- Viegas, E.O.; Tembe, N.; Nilsson, C.; Meggi, B.; Maueia, C.G.; Augusto, O.; Stout, R.; Scarlatti, G.; Ferrari, G.; Earl, P.L.; et al. Intradermal HIV-1 DNA immunization using needle-free zetajet injection followed by HIV-modified vaccinia virus ankara vaccination is safe and immunogenic in nozambican young adults: A phase I randomized controlled trial. AIDS Res. Hum. Retrovir. 2018, 34, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhao, T.; Zeng, T.; Wu, N.; Xiao, Y.; Liu, S.; Yu, J.; Jiang, C.; Gan, L.; Deng, M.; et al. Intramuscular primary immunization by nucleic acid vaccine pcDNA/Gpd-IL-2 and enhanced immunization with mucosal adjuvant CpG-ODN and Gpd-IL-2 recombinant protein effectively induced strong mucosal immune responses and immune protective effects against Treponema pallidum skin infection. Exp. Ther. Med. 2018, 15, 2533–2540. [Google Scholar] [CrossRef] [PubMed]
- Zhai, Y.Z.; Li, X.M.; Zhou, Y.; Ma, L.; Feng, G.H. Intramuscular immunization with a plasmid DNA vaccine encoding prM-E protein from Japanese encephalitis virus: Enhanced immunogenicity by co-administration of GM-CSF gene and genetic fusions of prM-E protein and GM-CSF. Intervirology 2009, 52, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Westrop, S.J.; Moyle, G.; Jackson, A.; Nelson, M.; Mandalia, S.; Imami, N. CCR5 Antagonism impacts vaccination response and immune profile in HIV-1 Infection. Mol. Med. 2012, 18, 1240–1248. [Google Scholar] [CrossRef]
- Xiao, J.; Chen, T.; Wang, Q.; Liu, Q.; Wang, X.; Lv, Y.; Wu, H.; Zhang, Y. Search for live attenuated vaccine candidate against edwardsiellosis by mutating virulence-related genes of fish pathogen Edwardsiella tarda. Lett. Appl. Microbiol. 2011, 53, 430–437. [Google Scholar] [CrossRef]
- Huang, J.-L.; Yin, Y.-X.; Pan, Z.-M.; Zhang, G.; Zhu, A.-P.; Liu, X.-F.; Jiao, X.-A. Intranasal immunization with chitosan/pCAGGS-flaA Nanoparticles inhibits Campylobacter jejuni in a white leghorn model. J. Biomed. Biotechnol. 2010, 2010, 589476. [Google Scholar] [CrossRef] [Green Version]
- De Leeuw, P.W.; van Bekkum, J.G.; Tiessink, J.W.A. Excretion of foot-and-mouth disease virus in oesophageal-pharyngeal fluid and milk of cattle after intranasal infection. Epidemiol. Infect. 1978, 81, 415–426. [Google Scholar] [CrossRef] [Green Version]
- Vasilyev, K.; Shurygina, A.-P.; Sergeeva, M.; Stukova, M.; Egorov, A. intranasal immunization with the influenza A virus encoding truncated NS1 protein protects mice from heterologous challenge by restraining the inflammatory response in the lungs. Microorganisms 2021, 9, 690. [Google Scholar] [CrossRef]
- Mills, K.H.G.; Cosgrove, C.; McNeela, E.A.; Sexton, A.; Giemza, R.; Jabbal-Gill, I.; Church, A.; Lin, W.; Illum, L.; Podda, A.; et al. Protective levels of diphtheria-neutralizing antibody induced in healthy volunteers by unilateral priming-boosting in-tranasal immunization associated with restricted ipsilateral mucosal secretory immunoglobulin a. Infect. Immun. 2003, 71, 726–732. [Google Scholar] [CrossRef] [Green Version]
- Van Beek, L.F.; Langereis, J.D.; van Saparoea, H.V.D.B.; Gillard, J.; Jong, W.S.P.; van Opzeeland, F.J.; Mesman, R.; van Niftrik, L.A.M.P.; Joosten, I.; Diavatopoulos, D.A.; et al. Intranasal vaccination with protein bodies elicit strong protection against Streptococcus pneumoniae colonization. Vaccine. 2021, 39, 6920–6929. [Google Scholar] [CrossRef]
- Zhao, H.; Yang, J.; Qian, Q.; Wu, M.; Li, M.; Xu, W. Mesenteric CD103+DCs initiate switched coxsackievirus B3 VP1-Specific IgA response to intranasal chitosan-DNA vaccine through secreting BAFF/IL-6 and promoting Th17/Tfh differentiation. Front Immunol. 2018, 9, 2986. [Google Scholar] [CrossRef] [PubMed]
- Schild, S.; Nelson, E.J.; Camilli, A. Immunization with Vibrio cholerae outer membrane vesicles induces protective immunity in mice. Infect Immun. 2008, 76, 4554–4563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abkar, M.; Fasihi-Ramandi, M.; Kooshki, H.; Sahebghadam Lotfi, A. Oral immunization of mice with Omp31-loaded N-trimethyl chitosan nanoparticles induces high protection against Brucella melitensis infection. Int. J. Nanomed. 2017, 12, 8769–8778. [Google Scholar] [CrossRef] [Green Version]
- Zhang, F.; Hao, C.; Zhang, S.; Li, A.; Zhang, Q.; Wu, W.; Liu, L.; Li, C.; Liang, M.; Li, X.; et al. Oral immunization with recombinant enterovirus 71 VP1 formulated with chitosan protects mice against lethal challenge. Virol. J. 2014, 11, 80. [Google Scholar] [CrossRef] [Green Version]
- Huang, X.; Ma, Y.; Wang, Y.; Niu, C.; Liu, Z.; Yao, X.; Jiang, X.; Pan, R.; Jia, S.; Li, D.; et al. Oral probiotic vaccine expressing koi herpesvirus (KHV) ORF81 protein delivered by chitosan-alginate capsules is a promising strategy for mass oral vaccination of carps against KHV infection. J. Virol. 2021, 95, e00415-21. [Google Scholar] [CrossRef]
- Oliveira, C.R.; Rezende, C.M.F.; Silva, M.R.; Borges, O.M.; Pêgo, A.P.; Goes, A.M. Oral vaccination based on DNA-chitosan nanoparticles against Schistosoma mansoni infection. Sci. World J. 2012, 2012, 938457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milligan, R.; Paul, M.; Richardson, M.; Neuberger, A. Vaccines for preventing typhoid fever. Cochrane Database Syst. Rev. 2018, 5, CD001261. [Google Scholar] [CrossRef] [PubMed]
- Chudina, T.; Labyntsev, A.; Manoilov, K.; Kolybo, D.; Komisarenko, S. Cellobiose-coated poly (lactide-co-glycolide) particles loaded with diphtheria toxoid for per os immunization. Croat. Med. J. 2015, 56, 85–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, C.R.; Rezende, C.M.F.; Silva, M.R.; Pêgo, A.P.; Borges, O.; Goes, A.M. A new strategy based on Smrho Protein loaded chitosan nanoparticles as a candidate oral vaccine against Schistosomiasis. PLOS Negl. Trop. Dis. 2012, 6, e1894. [Google Scholar] [CrossRef] [Green Version]
- Periman, L.M.; Mah, F.S.; Karpecki, P.M. A review of the mechanism of action of cyclosporine A: The role of cyclosporine A in dry eye disease and recent formulation developments. Clin. Ophthalmol. 2020, 14, 4187–4200. [Google Scholar] [CrossRef]
- Irie, H.; Shimeld, C.; Williams, N.; Hill, T. Protection against ocular and cutaneous infection with herpes simplex virus type 1 by intragastric immunization with live virus. J. Gen. Virol. 1993, 74, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Yao, X.; Yuan, C.; Liu, X.; Xia, R.; He, J.; Li, R.; Yao, Y. The investigation of hepatitis B vaccine immune responses in occult hepatitis B virus-infected patients. Front. Immunol. 2022, 13, 903685. [Google Scholar] [CrossRef] [PubMed]
- Mishra, H.; Mishra, D.; Mishra, P.K.; Nahar, M.; Dubey, V.; Jain, N.K. Evaluation of solid lipid nanoparticles as carriers for delivery of hepatitis B surface antigen for vaccination using subcutaneous route. J. Pharm. Pharm. Sci. 2010, 13, 495–509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, H.; Helms, C.M.; Grizzard, M.B.; James, W.D.; Horswood, R.L.; Chanock, R.M. Immunoprophylaxis of experimental Mycoplasma pneumoniae disease: Effect of route of administration on the immunogenicity and protective effect of inactivated M. pneumoniae vaccine. Infect. Immun. 1977, 16, 88–92. [Google Scholar] [CrossRef] [Green Version]
- Boland, K.G.; Hayles, A.N.; Miller, C.B.; Kerr, T.; Brown, W.C.; Lahmers, K.K. Regional immune response to immunization with Escherichia coli O157:H7-Derived intimin in cattle. Clin. Vaccine Immunol. 2013, 20, 562–571. [Google Scholar] [CrossRef] [Green Version]
- Barisani-Asenbauer, T.; Inić-Kanada, A.; Belij, S.; Marinković, E.; Stojićević, I.; Montanaro, J.; Stein, E.; Bintner, N.; Stojanović, M. The ocular conjunctiva as a mucosal immunization route: A profile of the immune response to the model antigen tetanus toxoid. PLoS ONE 2013, 8, e60682. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Zhou, T.; Luo, Y.; Dong, L.; Li, C.; Liu, J.; Luo, J.; Yan, R.; Xu, L.; Song, X.; et al. Modulation effects of Toxoplasma gondii histone H2A1 on murine macrophages and encapsulation with polymer as a vaccine candidate. Vaccines 2020, 8, 731. [Google Scholar] [CrossRef]
- Zhu, M.; Gao, F.; Li, Z.; Wang, X.; Wang, H.; Wang, Z.; Zhao, J.; Zhao, W. Immunoprotection of recombinant Eg.myophilin against Echinococcus granulosus infection in sheep. Exp. Ther. Med. 2016, 12, 1585–1590. [Google Scholar] [CrossRef] [Green Version]
- Beyrodt, C.G.; Pinto, A.R.; Freymüller, E.; Barbiéri, C.L. Characterization of an antigen from Leishmania amazonensis amastigotes able to elicit protective responses in a murine model. Infect. Immun. 1997, 65, 2052–2059. [Google Scholar] [CrossRef] [Green Version]
- Muflihah, H.; Flórido, M.; Lin, L.C.W.; Xia, Y.; Triccas, J.A.; Stambas, J.; Britton, W.J. Boosting BCG with recombinant influenza A virus tuberculosis vaccines increases pulmonary T cell responses but not protection against Mycobacterium tuberculosis infection. PLoS ONE 2021, 16, e0259829. [Google Scholar] [CrossRef]
- Yang, J.; Liang, G.; Xiang, T.; Situ, W. Effect of crosslinking processing on the chemical structure and biocompatibility of a chitosan-based hydrogel. Food Chem. 2021, 354, 129476. [Google Scholar] [CrossRef] [PubMed]
- Deng, X.; Zhao, J.; Liu, K.; Wu, C.; Liang, F. Stealth PEGylated chitosan polyelectrolyte complex nanoparticles as drug delivery carrier. J. Biomater. Sci. Polym. Ed. 2021, 32, 1387–1405. [Google Scholar] [CrossRef]
- Bravo-Anaya, L.M.; Fernández-Solís, K.G.; Rosselgong, J.; Nano-Rodríguez, J.; Carvajal, F.; Rinaudo, M. Chitosan-DNA polyelectrolyte complex: Influence of chitosan characteristics and mechanism of complex formation. Int. J. Biol. Macromol. 2019, 126, 1037–1049. [Google Scholar] [CrossRef] [PubMed]
- Chandra Hembram, K.; Prabha, S.; Chandra, R.; Ahmed, B.; Nimesh, S. Advances in preparation and characterization of chitosan nanoparticles for therapeutics. Artif. Cells Nanomed. Biotechnol. 2016, 44, 305–314. [Google Scholar] [CrossRef] [PubMed]
- Tahara, K.; Yamamoto, H.; Hirashima, N.; Kawashima, Y. Chitosan-modified poly(D,L-lactide-co-glycolide) nanospheres for improving siRNA delivery and gene-silencing effects. Eur. J. Pharm. Biopharm. 2010, 74, 421–426. [Google Scholar] [CrossRef]
- Tahara, K.; Sakai, T.; Yamamoto, H.; Takeuchi, H.; Hirashima, N.; Kawashima, Y. Improvements in transfection efficiency with chitosan modified poly(DL-lactide-co-glycolide) nanospheres prepared by the emulsion solvent diffusion method, for gene delivery. Chem. Pharm. Bull Tokyo. 2011, 59, 298–301. [Google Scholar] [CrossRef] [Green Version]
- Gentile, P.; Nandagiri, V.K.; Pabari, R.; Daly, J.; Tonda-Turo, C.; Ciardelli, G.; Ramtoola, Z. Influence of parathyroid hormone-loaded PLGA nanoparticles in porous scaffolds for bone regeneration. Int. J. Mol. Sci. 2015, 16, 20492–20510. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Ye, W.; Zheng, Y.; Liu, X.; Tong, Z. Fabrication of drug-loaded biodegradable microcapsules for controlled release by combination of solvent evaporation and layer-by-layer self-assembly. Int. J. Pharm. 2007, 338, 165–173. [Google Scholar] [CrossRef]
- Zheng, K.; Xiao, S.; Li, W.; Wang, W.; Chen, H.; Yang, F.; Qin, C. Chitosan-acorn starch-eugenol edible film: Physico-chemical, barrier, antimicrobial, antioxidant and structural properties. Int. J. Biol. Macromol. 2019, 135, 344–352. [Google Scholar] [CrossRef]
- Zhang, Z.-L.; Li, L.-J.; Sun, D.; Wang, M.; Shi, J.-R.; Yang, D.; Wang, L.-H.; Zou, S.-C. Preparation and properties of chitosan-based microspheres by spray drying. Food Sci. Nutr. 2020, 8, 1933–1941. [Google Scholar] [CrossRef]
- Wei, Y.; Huang, Y.-H.; Cheng, K.-C.; Song, Y.-L. Investigations of the influences of processing conditions on the properties of spray dried chitosan-tripolyphosphate particles loaded with theophylline. Sci. Rep. 2020, 10, 1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garg, U.; Chauhan, S.; Nagaich, U.; Jain, N. Current advances in chitosan nanoparticles based drug delivery and targeting. Adv. Pharm. Bull. 2019, 9, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saeed, R.M.; Dmour, I.; Taha, M.O. Stable Chitosan-based nanoparticles using polyphosphoric acid or hexametaphosphate for tandem ionotropic/covalent crosslinking and subsequent investigation as novel vehicles for drug delivery. Front. Bioeng. Biotechnol. 2020, 8, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orellano, M.S.; Porporatto, C.; Silber, J.J.; Falcone, R.D.; Correa, N.M. AOT reverse micelles as versatile reaction media for chitosan nanoparticles synthesis. Carbohydr. Polym. 2017, 171, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Mikušová, V.; Mikuš, P. Advances in chitosan-based nanoparticles for drug delivery. Int. J. Mol. Sci. 2021, 22, 9652. [Google Scholar] [CrossRef]
- Koppolu, B.; Zaharoff, D.A. The effect of antigen encapsulation in chitosan particles on uptake, activation and presentation by antigen presenting cells. Biomaterials. 2013, 34, 2359–2369. [Google Scholar] [CrossRef] [Green Version]
- Imam, S.S.; Alshehri, S.; Ghoneim, M.M.; Zafar, A.; Alsaidan, O.A.; Alruwaili, N.K.; Gilani, S.J.; Rizwanullah, M. Recent advancement in chitosan-based nanoparticles for improved oral bioavailability and bioactivity of phytochemicals: Challenges and perspectives. Polym. Basel. 2021, 13, 4036. [Google Scholar] [CrossRef]
- Kim, T.H.; Nah, J.W.; Cho, M.-H.; Park, T.G.; Cho, C.S. Receptor-mediated gene delivery into antigen presenting cells using mannosylated chitosan/DNA nanoparticles. J. Nanosci. Nanotechnol. 2006, 6, 2796–2803. [Google Scholar] [CrossRef]
- Kojarunchitt, T.; Baldursdottir, S.; Dong, Y.-D.; Boyd, B.J.; Rades, T.; Hook, S. Modified thermoresponsive Poloxamer 407 and chitosan sol–gels as potential sustained-release vaccine delivery systems. Eur. J. Pharm. Biopharm. 2015, 89, 74–81. [Google Scholar] [CrossRef]
- Karthick Raja Namasivayam, S.; Nishanth, A.N.; Arvind Bharani, R.S.; Nivedh, K.; Syed, N.H.; Samuel, R.R. Hepatitis B-surface antigen (HBsAg) vaccine fabricated chitosan-polyethylene glycol nanocomposite (HBsAg-CS-PEG- NC) preparation, immunogenicity, controlled release pattern, biocompatibility or non-target toxicity. Int. J. Biol. Macromol. 2020, 144, 978–994. [Google Scholar] [CrossRef]
- Ishii-Mizuno, Y.; Umeki, Y.; Onuki, Y.; Watanabe, H.; Takahashi, Y.; Takakura, Y.; Nishikawa, M. Improved sustained release of antigen from immunostimulatory DNA hydrogel by electrostatic interaction with chitosan. Int. Pharm. 2017, 516, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Fan, Q.; Miao, C.; Huang, Y.; Yue, H.; Wu, A.; Wu, J.; Wu, J.; Ma, G. Hydroxypropyltrimethyl ammonium chloride chitosan-based hydrogel as the split H5N1 mucosal adjuvant: Structure-activity relationship. Carbohydr. Polym. 2021, 266, 118139. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Huang, S.; Lu, L.; Song, X.; Li, P.; Wang, F. Curdlan sulfate-O-linked quaternized chitosan nanoparticles: Potential adjuvants to improve the immunogenicity of exogenous antigens via intranasal vaccination. Int. J. Nanomed. 2018, 13, 2377–2394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, G.; Jia, P.; Liu, H.; Hu, T.; Du, Y. Conjugation of chitosan oligosaccharides enhances immune response to porcine circovirus vaccine by activating macrophages. Immunobiology. 2018, 223, 663–670. [Google Scholar] [CrossRef]
- Shim, S.; Soh, S.H.; Im, Y.B.; Park, H.-E.; Cho, C.-S.; Kim, S.; Yoo, H.S. Elicitation of Th1/Th2 related responses in mice by chitosan nanoparticles loaded with Brucella abortus malate dehydrogenase, outer membrane proteins 10 and 19. Int. J. Med. Microbiol. 2020, 310, 151362. [Google Scholar] [CrossRef]
- Wen, Z.-S.; Xu, Y.-L.; Zou, X.-T.; Xu, Z.-R. Chitosan nanoparticles act as an adjuvant to promote both Th1 and Th2 immune responses induced by ovalbumin in mice. Mar. Drugs. 2011, 9, 1038–1055. [Google Scholar] [CrossRef]
- Svindland, S.C.; Pedersen, G.K.; Pathirana, R.D.; Bredholt, G.; Nøstbakken, J.K.; Jul-Larsen, Å.; Guzmán, C.A.; Montomoli, E.; Lapini, G.; Piccirella, S.; et al. A study of chitosan and c-di-GMP as mucosal adjuvants for intranasal influenza H5N1 vaccine. Influ. Other Respir. Viruses 2013, 7, 1181–1193. [Google Scholar] [CrossRef] [Green Version]
- Ghendon, Y.; Markushin, S.; Vasiliev, Y.; Akopova, I.; Koptiaeva, I.; Krivtsov, G.; Borisova, O.; Ahmatova, N.; Kurbatova, E.; Mazurina, S.; et al. Evaluation of properties of chitosan as an adjuvant for inactivated influenza vaccines administered parenterally. J. Med. Virol. 2009, 81, 494–506. [Google Scholar] [CrossRef]
- Chua, B.Y.; Al Kobaisi, M.; Zeng, W.; Mainwaring, D.; Jackson, D.C. Chitosan microparticles and nanoparticles as biocompatible delivery vehicles for peptide and protein-based immunocontraceptive vaccines. Mol. Pharm. 2012, 9, 81–90. [Google Scholar] [CrossRef]
- Boraschi, D.; Italiani, P.; Palomba, R.; Decuzzi, P.; Duschl, A.; Fadeel, B.; Moghimi, S.M. Nanoparticles and innate immunity: New perspectives on host defence. Semin. Immunol. 2017, 34, 33–51. [Google Scholar] [CrossRef]
- Yu, X.; Wen, T.; Cao, P.; Shan, L.; Li, L. Alginate-chitosan coated layered double hydroxide nanocomposites for enhanced oral vaccine delivery. J. Colloid Interface Sci. 2019, 556, 258–265. [Google Scholar] [CrossRef] [PubMed]
- Xu, B.; Zhang, W.; Chen, Y.; Xu, Y.; Wang, B.; Zong, L. Eudragit® L100-coated mannosylated chitosan nanoparticles for oral protein vaccine delivery. Int. J. Biol. Macromol. 2018, 113, 534–542. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Miyaji, E.N.; Gonçalves, V.M.; Ferreira, D.M.; Solórzano, C.; MacLoughlin, R.; Saleem, I. Evaluation of polymer choice on immunogenicity of chitosan coated PLGA NPs with surface-adsorbed pneumococcal protein antigen PspA4Pro. Int. J. Pharm. 2021, 599, 120407. [Google Scholar] [CrossRef]
- Renu, S.; Han, Y.; Dhakal, S.; Lakshmanappa, Y.S.; Ghimire, S.; Feliciano-Ruiz, N.; Senapati, S.; Narasimhan, B.; Selvaraj, R.; Renukaradhya, G.J. Chitosan-adjuvanted Salmonella subunit nanoparticle vaccine for poultry delivered through drinking water and feed. Carbohydr. Polym. 2020, 243, 116434. [Google Scholar] [CrossRef] [PubMed]
- Sadeghi, Z.; Fasihi-Ramandi, M.; Azizi, M.; Bouzari, S. Mannosylated chitosan nanoparticles loaded with FliC antigen as a novel vaccine candidate against Brucella melitensis and Brucella abortus infection. J. Biotechnol. 2020, 310, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Koppolu, B.P.; Smith, S.G.; Ravindranathan, S.; Jayanthi, S.; Suresh-Kumar, T.K.; Zaharoff, D.A. Controlling chitosan-based encapsulation for protein and vaccine delivery. Biomaterials. 2014, 35, 4382–4389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nevagi, R.J.; Khalil, Z.G.; Hussein, W.M.; Powell, J.; Batzloff, M.R.; Capon, R.J.; Good, M.F.; Skwarczynski, M.; Toth, I. Polyglutamic acid-trimethyl chitosan-based intranasal peptide nano-vaccine induces potent immune responses against group A streptococcus. Acta Biomater. 2018, 80, 278–287. [Google Scholar] [CrossRef] [Green Version]
- Junior, S.R.D.S.; Da Silva, F.K.L.; Dias, L.S.; Souza, A.C.O.; De Araujo, M.V.; Da Silva, L.B.R.; Travassos, L.R.; Amaral, A.C.; Taborda, C.P. Intranasal vaccine using P10 peptide complexed within chitosan polymeric nanoparticles as experimental therapy for paracoccidioidomycosis in murine model. J. Fungi Basel. 2020, 6, 160. [Google Scholar] [CrossRef]
- Jung, S.-N.; Kang, S.-K.; Yeo, G.-H.; Li, H.-Y.; Jiang, T.; Nah, J.-W.; Bok, J.-D.; Cho, C.-S.; Choi, Y.-J. Targeted delivery of vaccine to dendritic cells by chitosan nanoparticles conjugated with a targeting peptide ligand selected by phage display technique. Macromol. Biosci. 2015, 15, 395–404. [Google Scholar] [CrossRef]
- Robles-Planells, C.; Barrera-Avalos, C.; Rojo, L.E.; Spencer, E.; Martin, M.C.-S.; Matiacevich, S.; Pavez, J.; Milla, L.A.; Navarro, F.D.; Martínez, B.A.; et al. Chitosan-based delivery of avian reovirus fusogenic protein p10 gene: In vitro and in vivo studies towards a new vaccine against melanoma. Biomed. Res. Int. 2020, 2020, 4045760. [Google Scholar] [CrossRef]
- Renu, S.; Feliciano-Ruiz, N.; Patil, V.; Schrock, J.; Han, Y.; Ramesh, A.; Dhakal, S.; Hanson, J.; Krakowka, S.; Renukaradhya, G.J. Immunity and protective efficacy of mannose conjugated chitosan-based influenza nanovaccine in maternal antibody positive pigs. Front. Immunol. 2021, 12, 584299. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Xu, Q.; Su, L.; Xu, J.; Liu, Z.; Yang, Y.; Tang, H.; Li, Y.; Fan, M. The combinations chitosan-Pam3CSK4 and chitosan-monophosphoryl lipid A: Promising immune-enhancing adjuvants for anticaries vaccine PAc. Infect. Immun. 2019, 87, e00651-19. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Xue, J.; Jiang, M.; Lin, S.; Huang, Y.; Deng, K.; Shu, L.; Xu, H.; Li, Z.; Yao, J.; et al. A multiepitope peptide, rOmp22, encapsulated in chitosan-PLGA nanoparticles as a candidate vaccine against Acinetobacter baumannii infection. Int. J. Nanomed. 2021, 16, 1819–1836. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Singh, B.; Maharjan, S.; Li, H.S.; Kang, S.K.; Bok, J.D.; Cho, C.S.; Choi, Y.J. Oral delivery of probiotic expressing M cell homing peptide conjugated BmpB vaccine encapsulated into alginate/chitosan/alginate microcapsules. Eur. J. Pharm. Biopharm. 2014, 88, 768–777. [Google Scholar] [CrossRef]
- Büyüktimkin, B.; Wang, Q.; Kiptoo, P.; Stewart, J.M.; Berkland, C.; Siahaan, T.J. Vaccine-like controlled-release delivery of an immunomodulating peptide to treat experimental autoimmune encephalomyelitis. Mol. Pharm. 2012, 9, 979–985. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Jin, W.; Cruz, J.G.; Marasini, N.; Khalil, Z.G.; Capon, R.J.; Hussein, W.M.; Skwarczynski, M.; Toth, I. Development of polyelectrolyte complexes for the delivery of peptide-based subunit vaccines against group A Streptococcus. Nanomater. Basel. 2020, 10, 823. [Google Scholar] [CrossRef]
- Xu, Y.; Zhou, Z.; Brooks, B.; Ferguson, T.; Obliosca, J.; Huang, J.; Kaneko, I.; Iwanaga, S.; Yuda, M.; Tsuji, Y.; et al. Layer-by-layer delivery of multiple antigens using trimethyl chitosan nanoparticles as a malaria vaccine candidate. Front. Immunol. 2022, 13, 900080. [Google Scholar] [CrossRef]
- Nguyen, K.G.; Mantooth, S.M.; Vrabel, M.R.; Zaharoff, D.A. Intranasal delivery of thermostable subunit vaccine for cross-reactive mucosal and systemic antibody responses against SARS-CoV-2. Front. Immunol. 2022, 13, 858904. [Google Scholar] [CrossRef]
- Tabynov, K.; Babayeva, M.; Nurpeisov, T.; Fomin, G.; Nurpeisov, T.; Saltabayeva, U.; Renu, S.; Renukaradhya, G.J.; Petrovsky, N.; Tabynov, K. Evaluation of a novel adjuvanted vaccine for ultrashort regimen therapy of artemisia pollen-induced allergic bronchial asthma in a mouse model. Front. Immunol. 2022, 13, 828690. [Google Scholar] [CrossRef]
- Silva-Marrero, J.I.; Villasante, J.; Rashidpour, A.; Palma, M.; Fàbregas, A.; Almajano, M.P.; Viegas, I.; Jones, J.G.; Miñarro, M.; Ticó, J.R.; et al. The administration of chitosan-tripolyphosphate-DNA nanoparticles to express exogenous SREBP1a enhances conversion of dietary carbohydrates into lipids in the liver of sparus aurata. Biomolecules 2019, 9, 297. [Google Scholar] [CrossRef]
- Tahamtan, A.; Barati, M.; Tabarraei, A.; Mohebbi, S.R.; Shirian, S.; Gorji, A.; Ghaemi, A. Antitumor immunity induced by genetic immunization with chitosan nanoparticle formulated adjuvanted for HPV-16 E7 DNA vaccine. Iran. J. Immunol. 2018, 15, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Kumar, U.S.; Afjei, R.; Ferrara, K.; Massoud, T.F.; Paulmurugan, R. Gold-nanostar-chitosan-mediated delivery of SARS-CoV-2 DNA vaccine for respiratory mucosal immunization: Development and proof-of-principle. ACS Nano. 2021, 15, 17582–17601. [Google Scholar] [CrossRef] [PubMed]
- Thirumalaikumar, E.; Lelin, C.; Sathishkumar, R.; Vimal, S.; Anand, S.B.; Babu, M.M.; Citarasu, T. Oral delivery of pVAX-OMP and pVAX-hly DNA vaccine using Chitosan-Tripolyphosphate (Cs-TPP) nanoparticles in Rohu, (Labeo rohita) for protection against Aeromonas hydrophila infection. Fish Shellfish Immunol. 2021, 115, 189–197. [Google Scholar] [CrossRef]
- Leya, T.; Ahmad, I.; Sharma, R.; Tripathi, G.; Kurcheti, P.P.; Rajendran, K.V.; Bedekar, M.K. Bicistronic DNA vaccine macromolecule complexed with poly lactic-co-glycolic acid-chitosan nanoparticles enhanced the mucosal immunity of Labeo rohita against Edwardsiella tarda infection. Int. J. Biol. Macromol. 2020, 156, 928–937. [Google Scholar] [CrossRef] [PubMed]
- Bande, F.; Arshad, S.S.; Bejo, M.H.; Omar, A.R.; Moeini, H.; Khadkodaei, S.; Wei, T.S.; Keong, Y.S.; Abba, Y.; Anka, I.A. Development and immunogenic potentials of chitosan-saponin encapsulated DNA vaccine against avian infectious bronchitis coronavirus. Microb. Pathog. 2020, 149, 104560. [Google Scholar] [CrossRef] [PubMed]
- Dubey, S.; Avadhani, K.; Mutalik, S.; Sivadasan, S.M.; Maiti, B.; Girisha, S.K.; Venugopal, M.N.; Mutoloki, S.; Evensen, Ø.; Karunasagar, I.; et al. Edwardsiella tarda OmpA encapsulated in chitosan nanoparticles shows superior protection over inactivated whole cell vaccine in orally vaccinated fringed-lipped peninsula carp (Labeo fimbriatus). Vaccines Basel. 2016, 4, 40. [Google Scholar] [CrossRef] [Green Version]
- Wu, M.; Zhao, H.; Li, M.; Yue, Y.; Xiong, S.; Xu, W. Intranasal vaccination with mannosylated chitosan formulated DNA vaccine enables robust IgA and cellular response induction in the lungs of mice and improves protection against pulmonary mycobacterial challenge. Front. Cell. Infect. Microbiol. 2017, 7, 445. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Li, X.; Jiang, J.; Cheng, A.; Wang, M.; Zhu, D.; Jia, R.; Chen, S.; Zhou, Y.; Chen, X.; et al. Distribution characteristics of DNA vaccine encoded with glycoprotein C from anatid herpesvirus 1 with chitosan and liposome as deliver carrier in ducks. Virol. J. 2013, 10, 89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lebre, F.; Borchard, G.; Faneca, H.; Pedroso de Lima, M.C.; Borges, O. Intranasal administration of novel chitosan nanoparticle/DNA complexes induces antibody response to Hepatitis B surface antigen in mice. Mol. Pharm. 2016, 13, 472–482. [Google Scholar] [CrossRef]
- Chehelgerdi, M.; Doosti, A. Effect of the cagW-based gene vaccine on the immunologic properties of BALB/c mouse: An efficient candidate for Helicobacter pylori DNA vaccine. J. Nanobiotechnol. 2020, 18, 63. [Google Scholar] [CrossRef]
- Nanda, R.K.; Hajam, I.A.; Edao, B.M.; Ramya, K.; Rajanga, M.; Sekar, S.C.; Ganesh, K.; Bhanuprakash, V.; Kishore, S. Immunological evaluation of mannosylated chitosan nanoparticles based foot and mouth disease virus DNA vaccine, pVAC FMDV VP1-OmpA in guinea pigs. Biologicals. 2014, 42, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Chen, R.; Zhang, H.; Bryers, J.D. Injectable biodegradable chitosan-alginate 3D porous gel scaffold for mRNA vaccine delivery. Macromol. Biosci. 2019, 19, e1800242. [Google Scholar] [CrossRef] [PubMed]
- Crommelin, D.J.; Anchordoquy, T.J.; Volkin, D.B.; Jiskoot, W.; Mastrobattista, E. Addressing the cold reality of mRNA vaccine stability. J. Pharm. Sci. 2021, 110, 997–1001. [Google Scholar] [CrossRef] [PubMed]
- Hajam, I.A.; Senevirathne, A.; Hewawaduge, C.; Kim, J.; Lee, J.H. Intranasally administered protein coated chitosan nanoparticles encapsulating influenza H9N2 HA2 and M2e mRNA molecules elicit protective immunity against avian influenza viruses in chickens. Veter-Res. 2020, 51, 37. [Google Scholar] [CrossRef] [Green Version]
- Démoulins, T.; Englezou, P.C.; Milona, P.; Ruggli, N.; Tirelli, N.; Pichon, C.; Sapet, C.; Ebensen, T.; Guzmán, C.A.; McCullough, K.C. Self-Replicating RNA vaccine delivery to dendritic cells. Methods Mol. Biol. 2017, 1499, 37–75. [Google Scholar] [CrossRef]
- Steinle, H.; Ionescu, T.-M.; Schenk, S.; Golombek, S.; Kunnakattu, S.-J.; Özbek, M.T.; Schlensak, C.; Wendel, H.P.; Avci-Adali, M. Incorporation of synthetic mRNA in injectable chitosan-alginate hybrid hydrogels for local and sustained expression of exogenous proteins in cells. Int. J. Mol. Sci. 2018, 19, 1313. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Wang, X.; Jiang, D.; Pei, Y.; Wang, Z.; Zhou, X.G.; Wu, G.L.; Mo, X.M.; Wang, H.S. Delivery of mRNA vaccines and anti-PDL1 siRNA through non-invasive transcutaneous route effectively inhibits tumor growth. Compos. B Eng. 2022, 233, 109648. [Google Scholar] [CrossRef]
- Zhang, N.N.; Li, X.F.; Deng, Y.Q.; Zhao, H.; Huang, Y.J.; Yang, G.; Huang, W.J.; Gao, P.; Zhou, C.; Zhang, R.R.; et al. A thermostable mRNA vaccine against COVID-19. Cell 2020, 182, 1271–1283. [Google Scholar] [CrossRef]
- Kremsner, P.G.; Mann, P.; Kroidl, A.; Leroux-Roels, I.; Schindler, C.; Gabor, J.J.; Schunk, M.; Leroux-Roels, G.; Bosch, J.J.; Fendel, R.; et al. Safety and immunogenicity of an mRNA-lipid nanoparticle vaccine candidate against SARS-CoV-2: A phase 1 randomized clinical trial. Wien. Klin. Wochenschr. 2021, 133, 931–941. [Google Scholar] [CrossRef]
- Linares-Fernández, S.; Lacroix, C.; Exposito, J.-Y.; Verrier, B. Tailoring mRNA vaccine to balance innate/adaptive immune response. Trends Mol. Med. 2020, 26, 311–323. [Google Scholar] [CrossRef]
- Han, Y.; Renu, S.; Patil, V.; Schrock, J.; Feliciano-Ruiz, N.; Selvaraj, R.; Renukaradhya, G.J. Immune response to Salmonella enteritidis infection in broilers immunized orally with chitosan-based salmonella subunit nanoparticle vaccine. Front. Immunol. 2020, 11, 935. [Google Scholar] [CrossRef] [PubMed]
- Safarzadeh, M.; Sadeghi, S.; Azizi, M.; Rastegari-Pouyani, M.; Pouriran, R.; Haji Molla Hoseini, M. Chitin and chitosan as tools to combat COVID-19: A triple approach. Int. J. Biol. Macromol. 2021, 183, 235–244. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.S.; Sung, S.H.; Chiang, C.E. Chitosan potentiation of warfarin effect. Ann. Pharmacother. 2007, 41, 1912–1914. [Google Scholar] [CrossRef]
- Srinivas, N.R. The interesting case of acyclovir delivered using chitosan in humans: Is it a drug issue or formulation Issue? Pharm. Res. 2016, 33, 543–547. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.C.; Vieira, A.; Huang, K.L.; Yeh, M.K.; Chiang, C.H. Pulmonary inflammation caused by chitosan microparticles. J. Biomed. Mater. Res. A. 2005, 75, 283–287. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Existing Vaccine | Preparation | Advantages | Disadvantages | References |
|---|---|---|---|---|
| Attenuated vaccines | Produced from pathogenic isolates via serial passage in embryonated, specific, pathogen-free eggs | Cause a strong immune response; long-lasting immune response; can survive the low-pH enzymatic environment of the stomach | Risk of toxic recovery in immunocompromised individuals; low safety profile | [3,4] |
| Inactivated vaccines | Formaldehyde inactivates live virus production | Compared with live attenuated vaccines, the safety is improved, and the production process is mature | Weak immune response; short duration; destruction or alteration of antigens; high cost; multiple vaccinations required | [4] |
| Subunit vaccines | Antigenic proteins from pathogens | High safety; modulated immune response; clear target antigen | Low immunogenicity | [4,5,6,7] |
| DNA vaccines | Transfection of DNA encoding a certain antigenic protein into animal cells | Simple to build; easy to mass-produce; high security and high stability | Cannot be used in humans; naked-pelleted DNA has low immunogenicity | [8,9,10,11] |
| mRNA vaccines | Transfection of RNA encoding a certain antigenic protein into animal cells | High transfection rate and simple process | Unstable; lack of effective delivery vehicles in vivo | [8,14,15,16,17,18,19] |
| Classification | Subdivision | Example | Mechanism |
|---|---|---|---|
| Inorganic | Aluminum salt | Aluminum hydroxide; Alhydrogel; Aluminum phosphate; Adju-Phos; Merck aluminum; AAHSA; Nano aluminum | Prolonged interactions between antigens and immune cells; prophagocytosis; induction of Th2 immune-biased responses; activation of the pro-inflammatory NLRP3 pathway [45,48] |
| Calcium salt | Calcium phosphate; Calcium phosphate NPs | Induces a more balanced Th1 and Th2 immune response [49] | |
| Other inorganic substances | Mn2+ | Enhancement of immune responses through the STING pathway in dendritic cells [50] | |
| Organic matter | Oil-in-water emulsion | MF59; AS03; AF03; SE | Induction of a Th2 immune bias response; local activation of the innate immune system [51] |
| Bacterially derived molecules | Monophosphoryl lipid A (MPL); Pyranose-based lipid adjuvant (GLA); RC529; CpG; stable toxin (LT); Cholera toxin (CT); TDB; Flagellin | Activates Toll-like receptor 4; triggers cells expressing Toll-like receptor 9 (including human plasmacytoid dendritic cells and B cells); induces Th1/Th2/Th17 responses; promotes dendritic cell uptake and maturation; activates the ERK1/2 pathway via PLC-γ1/PKC signaling [52,53,54,55,56,57,58,59] | |
| Virus-derived molecules | Poly I: C | Activation of TLR3; melanoma differentiation-associated gene 5 (MDA-5); cellular immunity and type I interferon response [60] | |
| Plant-derived molecules | QS21; Delta inulin | Promotes CD4+ T cell-mediated immune responses; induces a more balanced Th1/Th2 response [61,62]. | |
| Endogenous molecules | Cytokine | Activates dendritic cells [63] | |
| Synthesis of artificial small molecules | Imiquimod; R848; Cyclosporine | Activates TLR7/8 receptors and T cell co-stimulator CD40 [64,65] | |
| Composite adjuvant | Aluminum-based composite adjuvant | AS04 | Rapidly triggers local and transient cytokine responses to enhance humoral and cell-mediated responses, resulting in increased activation of antigen-presenting cells and promoting antigen presentation in CD4+ T lymphocytes [66] |
| Emulsion-based compound adjuvant | AS02 (AS03 + MPL+QS21), GLA-SE | Activation of TLR-4 regulates immune homeostasis and promotes Th1-type immune responses [67] | |
| Lipid group compound adjuvant | AS01 (liposome+MPL+QS21); AS15 (AS01+CpG); ISCOMs | Effectively promotes CD4+ T lymphocyte-mediated immune response [68] | |
| Polymer particles | CaCO3-LNT; Chitosan | Enhances the expression of MHC-II and CD86 in dendritic cells and increases the ratio of CD4+ to CD8+ T lymphocytes; provides a depot for the antigen to slowly release; facilitates the antigen’s targeting of immune cells; improves phagocytosis, modulation, and enhancement of the immune response induced by the antigen alone [69,70,71]. | |
| Virus Vectors | Advantages | Disadvantages | References |
|---|---|---|---|
| Retroviral-vector-based vaccines | Stable integration into the host genome; high gene transduction efficiency | Once infected, the likelihood of eliminating the virus is low | [86] |
| Herpes simplex virus vector vaccine | Enhances cellular and humoral immunity; Induces lasting immune response. | High cytotoxicity | [75,88,89] |
| Adenovirus vector vaccine | Induces moderate innate immunity; high thermal stability | Cause acute hepatotoxicity | [90,91,92] |
| Lentiviral vector vaccine | Induces durable humoral immunity | Insertional mutagenesis; yields low viral titers; easily degradable | [93,94] |
| Immunization Pathway | Pathogens |
|---|---|
| Intramuscular immunization | Chlamydia [108], Mycoplasma [109], Influenza virus [65], Bacillus anthracis [110], Mycobacterium tuberculosis [111], HIV [112] Treponema pallidum [113], Herpes virus [75], Japanese encephalitis virus [114], Cholera virus [115], Avian influenza virus [37], Edwardsiella tarda [116] |
| Intranasal immunization | Campylobacter jejuni [117], Foot-and-mouth disease [118], PR8 influenza virus [119], Escherichia coli [87], Diphtheria toxin [120], Influenza A virus [119], Newcastle disease virus [9], Streptococcus pneumoniae [121], Coxsackievirus [122], Vibrio cholerae [123] |
| Oral immunization | Salmonella [99], Brucella [124], Hepatitis B virus [21], Recombinant enterovirus [125], Koi herpesvirus [126], Schisoma mansoni [127], Vibrio cholerae [123], tetanus toxoid [128], diphtheria toxin [129], Schistosomiasis [130] |
| Ocular immunization | Cyclosporin A [131], Herpes simplex virus [132] |
| Subcutaneous immunization | Hepatitis B virus [133], Hepatitis B surface antigen [134], Mycoplasma pneumoniae [135], E. Coli [136], Tetanus toxoid [137], Toxoplasma gondii histones [138], Echinococcus granulosus [139], Influenza virus [37], Leishmania antigen [140], Mycobacterium tuberculosis [141] |
| Preparation Methods | Mechanism | Advantages | Disadvantages |
|---|---|---|---|
| Ion crosslinking [142] | Interaction between crosslinking agent and the amino or carboxyl groups of chitosan NPs. | The preparation process itself has no organic solvent; the reaction conditions are simple, mild, and controllable. | Not completely immune to gastric acid degradation; low solubility. |
| Polyelectrolyte complexation [143,144] | Interactions between oppositely charged polyelectrolytes. | Two- or three-step process; the equipment requirements are not demanding, and the conditions are mild. | Stability is susceptible to pH. |
| Desolvation [145] | Insolubility of chitosan in alkaline media. | High nanoparticle formation rate; improved physical stability. | Inhomogeneous distribution of NPs; difficulty in synthesizing smaller-sized chitosan NPs. |
| Emulsification [146,147,148,149,150] | The oil and water phases are emulsified; then, the solvent is removed. | Two- to three-step process; no need for sonication or homogenizers. | Causes significant toxicity to tissues or cells; poor stability. |
| Spray drying [151,152] | Amino groups can be protonated by acids. | Re-dispersibility; enables easier synthesis of smaller sizes compared other methods; low toxicity. | The particle size is not easy to control; the particles are irregularly shaped and sticky. |
| Covalent crosslinking [153,154] | Formation of covalent bonds between chitosan NPs and crosslinking agents. | Controllable drug release. | Cytotoxic. |
| Reverse micelle [153,155] | Trans-interaction between chitosan NPs and crosslinking agent. | One-step process; good dispersion. | Causes significant toxicity in tissues or cells. |
| Chitosan Complex | Protein/Polypeptide | Immune Mode | Target Disease |
|---|---|---|---|
| Chitosan-modified silica NPs [104] | Bovine serum albumin | Oral | Not mentioned |
| Chitosan NPs [175] | Salmonella enteritidis outer membrane proteins (OMPs) and flagellin proteins | Oral | Brucellosis |
| Chitosan NPs [181] | Fusogenic protein p10 of avian reovirus (ARV-p10) | Intramuscular injection | Melanoma |
| Mannosylated chitosan NPs [176] | FliC antigen | Subcutaneous injection | Brucellosis |
| N-2-Hydroxypropyl trimethyl ammonium chloride chitosan [39] | Porcine parvovirusVP2 protein | Intramuscular injection | Porcine parvovirus disease |
| Mannose-conjugated chitosan [182] | SwIAV antigen | Intranasal | Swine influenza |
| Chitosan [183] | Recombinant protein Pac | Oral | Dental caries |
| Chitosan polymeric NPs [179] | P10 peptide | Intranasal | Paracoccidioidomycosis |
| Chitosan–PLGA NPs [184] | rOmp22 peptide | Subcutaneous injection | Acinetobacter baumannii |
| Trimethyl chitosan [178] | A. streptococcus peptide | Intranasal | Group A Streptococcus |
| Alginate/chitosan/alginate microcapsules [185] | Probiotic expressing M cell homing peptide | Oral | Not mentioned |
| Alginate–chitosan–PLGA complex [186] | Ac-PLP-BPI-NH2-2 peptide | Subcutaneous injection | Autoimmune encephalomyelitis |
| Alginate and trimethyl chitosan [187] | Lipopeptide | Oral | Group A Streptococcus |
| Trimethyl Chitosan NPs [188] | Malaria antigens | Intramuscular injection | Malaria |
| Chitosan [189] | Receptor-binding domain (RBD) polypeptides | Subcutaneous injection | SARS-CoV-2 |
| Chitosan-mannose NPs [190] | Recombinant Art v 1 wormwood pollen protein | Intramuscular, subcutaneous, subcutaneous injection | Allergic bronchial asthma |
| Chitosan Complex | Causative Agent | Immune Animal or Model | Immunization Mode |
|---|---|---|---|
| Chitosan–saponin [196] | Avian infectious bronchitis virus | Chicken | Intramuscular injection |
| Chitosan–trimeric phosphate [194] | pVAX-OMP and pVAX-hly | Carp | Oral |
| Polylactic-co-glycolic acid-chitosan NPs [197] | Late-onset Edwardian spp. | Fish | Immunization by immersion |
| N-2-Hydroxypropyl dimethylethyl ammonium chloride chitosan NPs [38] | Newcastle disease virus | Chicken | Intranasal |
| Mannosylated chitosan [198] | Mycobacterium pneumonia | Mouse | Intranasal |
| N-2-HACC/CMCS NPs [40] | Newcastle disease virus | Chicken | Intranasal |
| Chitosan NPs [199] | Herpes simplex virus | Duck | Intramuscular injection |
| Chitosan NPs [200] | Hepatitis B antigen | Mouse | Intranasal |
| Chitosan NPs [201] | Helicobacter pylori | Mouse | Oral |
| Mannosylated chitosan NPs [202] | Mouth disease virus | Guinea pig | Intranasal |
| Chitosan Complex | Causative Agent | Diseases |
|---|---|---|
| Chitosan NPs [205] | Influenza H9N2 HA2 and M2e mRNA | Influenza |
| Chitosan NPs [206] | Influenza virus | Influenza |
| Chitosan–alginate hybrid hydrogel [207] | Synthetic mRNAs for tissue-engineering applications | Tissue-engineering applications |
| Mannosylated-chitosan-modified ethosomes [208] | Tyrosinase-related protein 2 | Melanoma |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gong, X.; Gao, Y.; Shu, J.; Zhang, C.; Zhao, K. Chitosan-Based Nanomaterial as Immune Adjuvant and Delivery Carrier for Vaccines. Vaccines 2022, 10, 1906. https://doi.org/10.3390/vaccines10111906
Gong X, Gao Y, Shu J, Zhang C, Zhao K. Chitosan-Based Nanomaterial as Immune Adjuvant and Delivery Carrier for Vaccines. Vaccines. 2022; 10(11):1906. https://doi.org/10.3390/vaccines10111906
Chicago/Turabian StyleGong, Xiaochen, Yuan Gao, Jianhong Shu, Chunjing Zhang, and Kai Zhao. 2022. "Chitosan-Based Nanomaterial as Immune Adjuvant and Delivery Carrier for Vaccines" Vaccines 10, no. 11: 1906. https://doi.org/10.3390/vaccines10111906