
Immunomodulatory Role of Thioredoxin Interacting Protein in Cancer’s Impediments: Current Understanding and Therapeutic Implications
 ,
,  , ,
, , 
Abstract
:1. Introduction
2. TXNIP
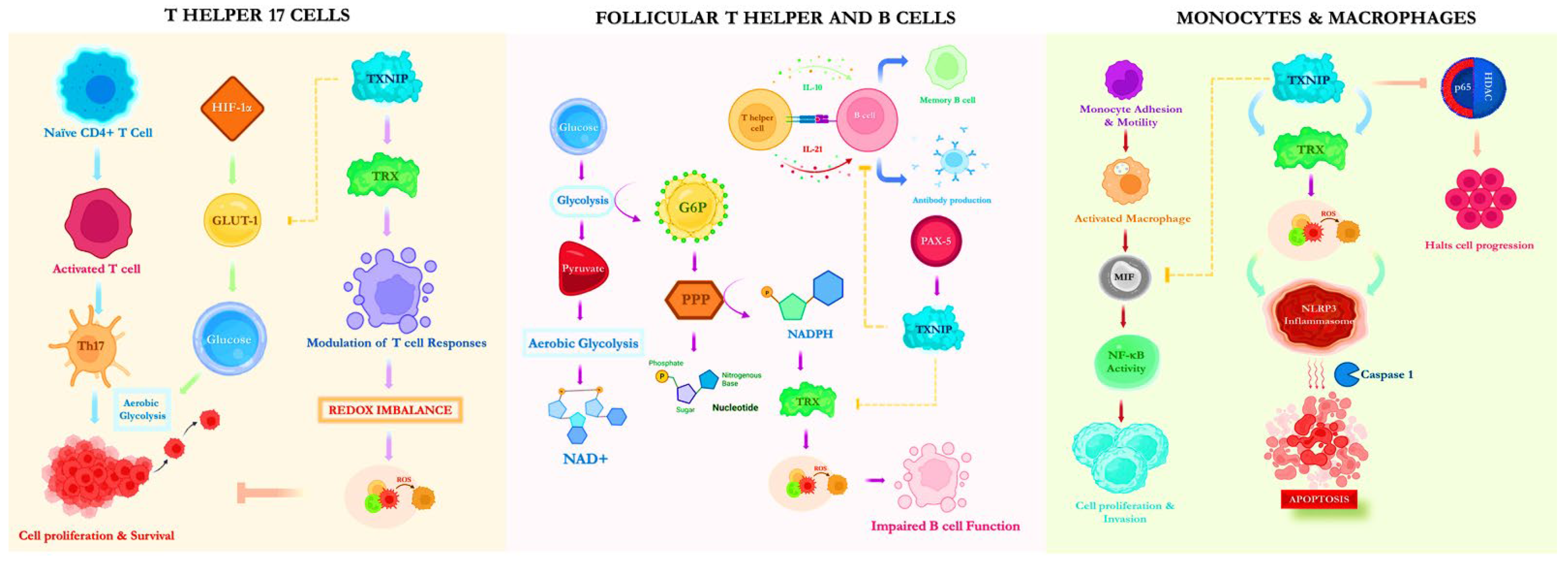
3. The Action of TXNIP on Immune Cells
3.1. T Helper 17 Cells
3.2. Follicular T Helper and B Cells
3.3. Monocytes and Macrophages
4. TXNIP and Inflammation and Immune Reaction
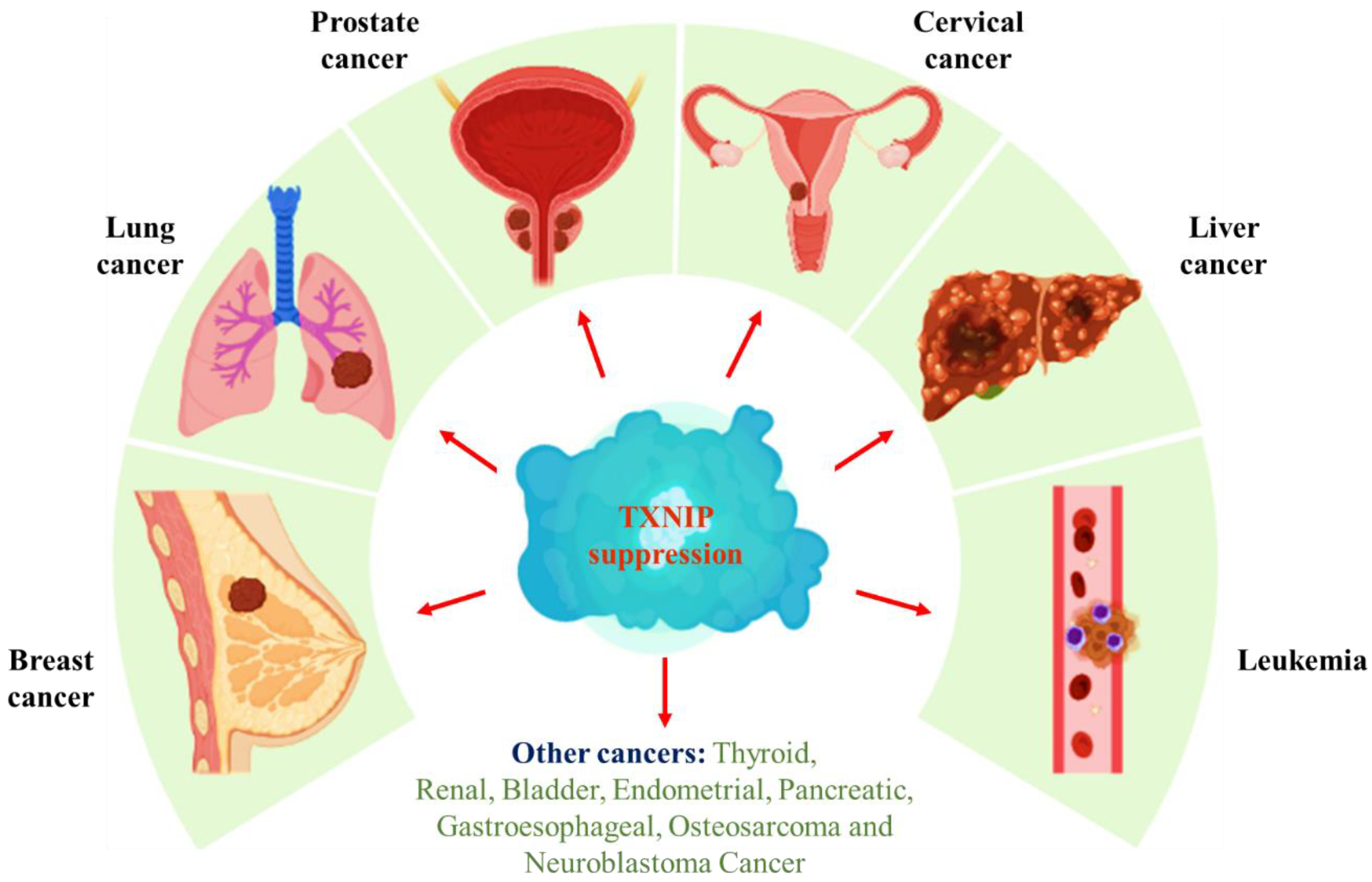
5. Role of TXNIP in Cancer
5.1. TXNIP and Breast Cancer
5.2. TXNIP and Lung Cancer
5.3. TXNIP and Prostate Cancer
5.4. TXNIP and Cervical Cancer
5.5. TXNIP and Liver Cancer
5.6. TXNIP and Leukemia
5.7. TXNIP and Other Cancer
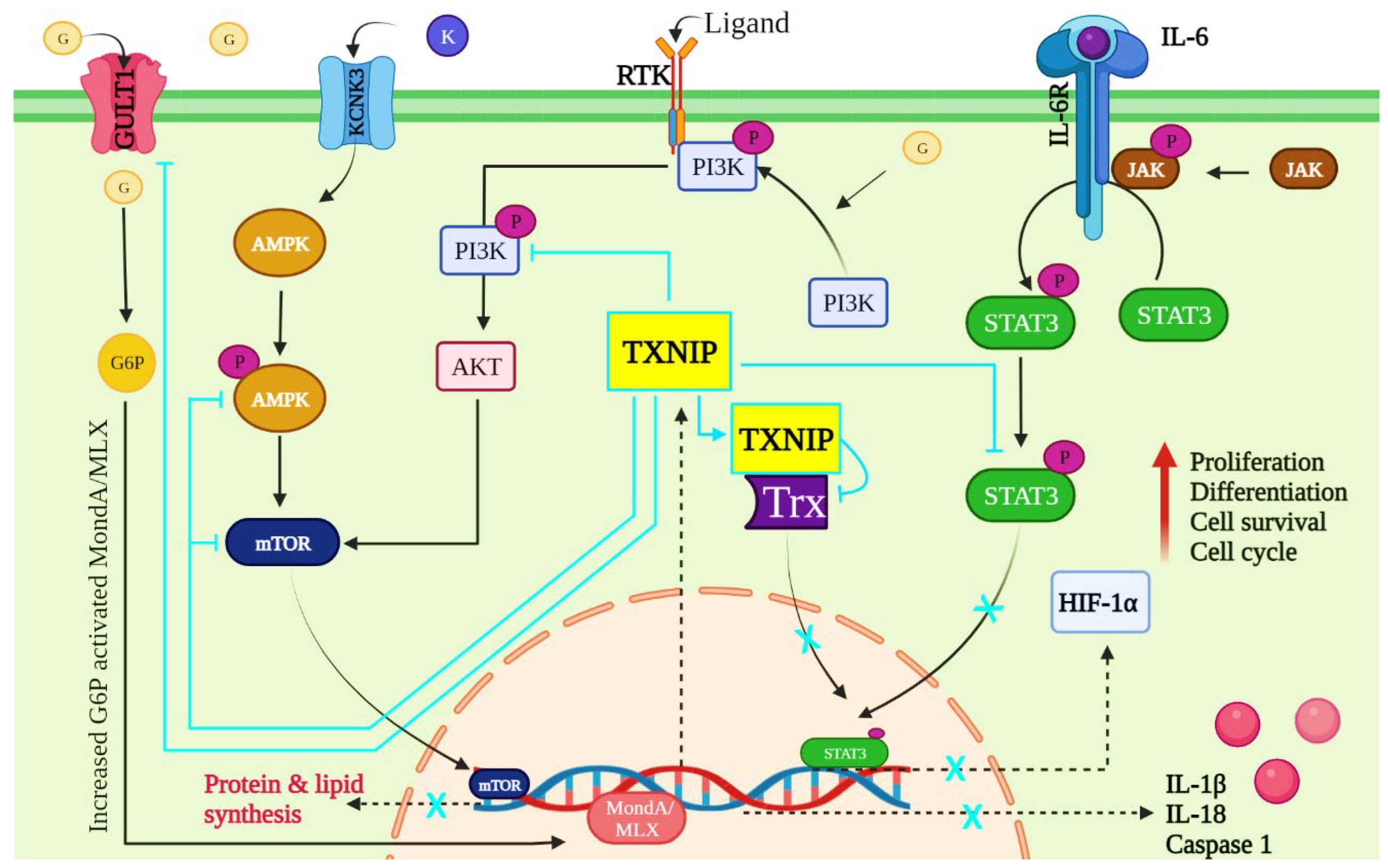
6. TXNIP-Dependent Signaling Pathways in Cancer
6.1. JAK/STAT Pathway
6.2. AMPK/mTOR Signaling Pathway
6.3. PI3K/AKT Signaling Pathway
6.4. Other Pathways
7. Therapeutic Targeting of TXNIP in Cancer
8. Future Perspective
9. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Marklew, R.E.; Jackson, A.A.; Wiseman, M.J.; Wootton, S.A. ICONIC: An International Task Force Supporting Collaboration in Nutrition and Cancer Globally. Trends Food Sci. Technol. 2022, 130, 3–10. [Google Scholar] [CrossRef]
- Debela, D.T.; Muzazu, S.G.; Heraro, K.D.; Ndalama, M.T.; Mesele, B.W.; Haile, D.C.; Kitui, S.K.; Manyazewal, T. New Approaches and Procedures for Cancer Treatment: Current Perspectives. SAGE Open Med. 2021, 9, 205031212110343. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Zhang, M.; Wang, Y. The Roles of Histone Modifications in Tumorigenesis and Associated Inhibitors in Cancer Therapy. J. Natl. Cancer Center 2022. [Google Scholar] [CrossRef]
- Wondafrash, D.Z.; Nire’a, A.T.; Tafere, G.G.; Desta, D.M.; Berhe, D.A.; Zewdie, K.A. Thioredoxin-Interacting Protein as a Novel Potential Therapeutic Target in Diabetes Mellitus and Its Underlying Complications. Diab. Metab. Syndr. Obes. 2020, 13, 43. [Google Scholar] [CrossRef] [Green Version]
- Schröder, J.; Schumacher, U.; Böckelmann, L.C. Thioredoxin Interacting Protein (TXNIP) Is Differentially Expressed in Human Tumor Samples but Is Absent in Human Tumor Cell Line Xenografts: Implications for Its Use as an Immunosurveillance Marker. Cancers 2020, 12, 3028. [Google Scholar] [CrossRef]
- Chen, L.; Deng, H.; Cui, H.; Fang, J.; Zuo, Z.; Deng, J.; Li, Y.; Wang, X.; Zhao, L. Inflammatory Responses and Inflammation-Associated Diseases in Organs. Oncotarget 2018, 9, 7204. [Google Scholar] [CrossRef] [Green Version]
- Pan, M.; Zhang, F.; Qu, K.; Liu, C.; Zhang, J. TXNIP: A Double-Edged Sword in Disease and Therapeutic Outlook. Oxid Med. Cell Longev. 2022, 2022, 7805115. [Google Scholar] [CrossRef]
- Wu, N.; Zheng, B.; Shaywitz, A.; Dagon, Y.; Tower, C.; Bellinger, G.; Shen, C.H.; Wen, J.; Asara, J.; McGraw, T.E.; et al. AMPK-Dependent Degradation of TXNIP upon Energy Stress Leads to Enhanced Glucose Uptake via GLUT1. Mol. Cell 2013, 49, 1167–1175. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Yu, Y. The Function of Thioredoxin-Binding Protein-2 (TBP-2) in Different Diseases. Oxid Med. Cell Longev. 2018, 2018, 4582130. [Google Scholar] [CrossRef] [Green Version]
- Hong, S.Y.; Hagen, T. 2-Deoxyglucose Induces the Expression of Thioredoxin Interacting Protein (TXNIP) by Increasing O-GlcNAcylation–Implications for Targeting the Warburg Effect in Cancer Cells. Biochem. Biophys. Res. Commun. 2015, 465, 838–844. [Google Scholar] [CrossRef]
- Yoshihara, E.; Masaki, S.; Matsuo, Y.; Chen, Z.; Tian, H.; Yodoi, J. Thioredoxin/Txnip: Redoxisome, as a Redox Switch for the Pathogenesis of Diseases. Front. Immunol 2013, 4, 514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Seco-Cervera, M.; González-Cabo, P.; Pallardó, F.v.; Romá-Mateo, C.; García-Giménez, J.L. Thioredoxin and Glutaredoxin Systems as Potential Targets for the Development of New Treatments in Friedreich’s Ataxia. Antioxidants 2020, 9, 1275. [Google Scholar] [CrossRef] [PubMed]
- Tsubaki, H.; Tooyama, I.; Walker, D.G. Thioredoxin-Interacting Protein (TXNIP) with Focus on Brain and Neurodegenerative Diseases. Int J. Mol. Sci 2020, 21, 9375. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Ning, J.; Cao, W.; Wang, S.; Du, T.; Jiang, J.; Feng, X.; Zhang, B. Research Progress of TXNIP as a Tumor Suppressor Gene Participating in the Metabolic Reprogramming and Oxidative Stress of Cancer Cells in Various Cancers. Front. Oncol. 2020, 10, 568574. [Google Scholar] [CrossRef]
- Spindel, O.N.; World, C.; Berk, B.C. Thioredoxin Interacting Protein: Redox Dependent and Independent Regulatory Mechanisms. Antioxid Redox Signal. 2012, 16, 587. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Chng, W.J. Roles of Thioredoxin Binding Protein (TXNIP) in Oxidative Stress, Apoptosis and Cancer. Mitochondrion 2013, 13, 163–169. [Google Scholar] [CrossRef]
- Pahl, M.v.; Vaziri, N.D. Immune Function in Chronic Kidney Disease. Chronic Renal Disease 2020, 503–519. [Google Scholar] [CrossRef]
- Charles A Janeway, J.; Travers, P.; Walport, M.; Shlomchik, M.J. T Cell-Mediated Immunity. Immunobiology: The Immune System in Health and Disease; Garland Science: New York, NY, USA, 2001. [Google Scholar]
- Mahmud, S.A.; Manlove, L.S.; Schmitz, H.M.; Xing, Y.; Wang, Y.; Owen, D.L.; Schenkel, J.M.; Boomer, J.S.; Green, J.M.; Yagita, H.; et al. Tumor Necrosis Factor Receptor Superfamily Costimulation Couples T Cell Receptor Signal Strength to Thymic Regulatory T Cell Differentiation. Nat. Immunol. 2014, 15, 473. [Google Scholar] [CrossRef]
- Palmer, C.S.; Ostrowski, M.; Balderson, B.; Christian, N.; Crowe, S.M. Glucose Metabolism Regulates T Cell Activation, Differentiation, and Functions. Front. Immunol. 2015, 6, 1. [Google Scholar] [CrossRef] [Green Version]
- Zuo, H.; Yuan, J.; Yang, L.; Liang, Z.; Weng, S.; He, J.; Xu, X. Characterization and Immune Function of the Thioredoxin-Interacting Protein (TXNIP) from Litopenaeus Vannamei. Fish Shellfish Immunol. 2019, 84, 20–27. [Google Scholar] [CrossRef]
- Levring, T.B.; Kongsbak-Wismann, M.; Rode, A.K.O.; Al-Jaberi, F.A.H.; Lopez, D.v.; Met, Ö.; Woetmann, A.; Bonefeld, C.M.; Ødum, N.; Geisler, C. Tumor Necrosis Factor Induces Rapid Down-Regulation of TXNIP in Human T Cells. Sci. Rep. 2019, 9, 16725. [Google Scholar] [CrossRef] [PubMed]
- Luckheeram, R.V.; Zhou, R.; Verma, A.D.; Xia, B. CD4+T Cells: Differentiation and Functions. Clin. Dev. Immunol. 2012, 2012, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghanbari Movahed, Z.; Rastegari-Pouyani, M.; Mohammadi, M.h.; Mansouri, K. Cancer Cells Change Their Glucose Metabolism to Overcome Increased ROS: One Step from Cancer Cell to Cancer Stem Cell? Biomed. Pharmacother. 2019, 112, 108690. [Google Scholar] [CrossRef]
- Dang, E.v.; Barbi, J.; Yang, H.Y.; Jinasena, D.; Yu, H.; Zheng, Y.; Bordman, Z.; Fu, J.; Kim, Y.; Yen, H.R.; et al. Control of TH17/Treg Balance by Hypoxia-Inducible Factor 1. Cell 2011, 146, 772. [Google Scholar] [CrossRef] [Green Version]
- Zhong, X.; He, X.; Wang, Y.; Hu, Z.; Huang, H.; Zhao, S.; Wei, P.; Li, D. Warburg Effect in Colorectal Cancer: The Emerging Roles in Tumor Microenvironment and Therapeutic Implications. J. Hematol. Oncol. 2022, 15, 1–29. [Google Scholar] [CrossRef]
- Cicalese, M.P.; Salek-Ardakani, S.; Fousteri, G. Editorial: Follicular Helper T Cells in Immunity and Autoimmunity. Front. Immunol. 2020, 11, 1042. [Google Scholar] [CrossRef] [PubMed]
- Crotty, S. T Follicular Helper Cell Differentiation, Function, and Roles in Disease. Immunity 2014, 41, 529. [Google Scholar] [CrossRef] [Green Version]
- Sage, P.T.; Tan, C.L.; Freeman, G.J.; Haigis, M.; Sharpe, A.H. Defective TFH Cell Function and Increased TFR Cells Contribute to Defective Antibody Production in Aging. Cell Rep. 2015, 12, 163. [Google Scholar] [CrossRef] [Green Version]
- Castellano, F.; Molinier-Frenkel, V. Control of T-Cell Activation and Signaling by Amino-Acid Catabolizing Enzymes. Front. Cell Dev. Biol. 2020, 8, 613416. [Google Scholar] [CrossRef]
- Muri, J.; Thut, H.; Heer, S.; Krueger, C.C.; Bornkamm, G.W.; Bachmann, M.F.; Kopf, M. The Thioredoxin-1 and Glutathione/Glutaredoxin-1 Systems Redundantly Fuel Murine B-Cell Development and Responses. Eur J. Immunol. 2019, 49, 709–723. [Google Scholar] [CrossRef]
- Paul, S.; Lal, G. The Molecular Mechanism of Natural Killer Cells Function and Its Importance in Cancer Immunotherapy. Front. Immunol. 2017, 8, 1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chan, L.N.; Müschen, M. B-Cell Identity as a Metabolic Barrier against Malignant Transformation. Exp. Hematol. 2017, 53, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Muri, J.; Kopf, M. Redox Regulation of Immunometabolism. Nat. Rev. Immunol. 2020, 21, 363–381. [Google Scholar] [CrossRef]
- de Preter, G.; Neveu, M.A.; Danhier, P.; Brisson, L.; Payen, V.L.; Porporato, P.E.; Jordan, B.F.; Sonveaux, P.; Gallez, B. Inhibition of the Pentose Phosphate Pathway by Dichloroacetate Unravels a Missing Link between Aerobic Glycolysis and Cancer Cell Proliferation. Oncotarget 2016, 7, 2910. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patwardhan, R.S.; Sharma, D.; Sandur, S.K. Thioredoxin Reductase: An Emerging Pharmacologic Target for Radiosensitization of Cancer. Transl. Oncol. 2022, 17, 101341. [Google Scholar] [CrossRef]
- Muri, J.; Thut, H.; Kopf, M. The Thioredoxin-1 Inhibitor Txnip Restrains Effector T-Cell and Germinal Center B-Cell Expansion. Eur. J. Immunol. 2021, 51, 115–124. [Google Scholar] [CrossRef]
- Parihar, A.; Eubank, T.D.; Doseff, A.I. Monocytes and Macrophages Regulate Immunity through Dynamic Networks of Survival and Cell Death. J. Innate Immun. 2010, 2, 204. [Google Scholar] [CrossRef] [Green Version]
- Yan, X.; Orentas, R.J.; Johnson, B.D. Tumor-Derived Macrophage Migration Inhibitory Factor (MIF) Inhibits T Lymphocyte Activation. Cytokine 2006, 33, 188. [Google Scholar] [CrossRef] [Green Version]
- Guda, M.R.; Rashid, M.A.; Asuthkar, S.; Jalasutram, A.; Caniglia, J.L.; Tsung, A.J.; Velpula, K.K. Pleiotropic Role of Macrophage Migration Inhibitory Factor in Cancer. Am. J. Cancer Res. 2019, 9, 2760. [Google Scholar]
- Kim, M.J.; Kim, W.S.; Kim, D.O.; Byun, J.E.; Huy, H.; Lee, S.Y.; Song, H.Y.; Park, Y.J.; Kim, T.D.; Yoon, S.R.; et al. Macrophage Migration Inhibitory Factor Interacts with Thioredoxin-Interacting Protein and Induces NF-ΚB Activity. Cell Signal 2017, 34, 110–120. [Google Scholar] [CrossRef]
- Samadi, A.K.; Bilsland, A.; Georgakilas, A.G.; Amedei, A.; Amin, A.; Bishayee, A.; Azmi, A.S.; Lokeshwar, B.L.; Grue, B.; Panis, C.; et al. A Multi-Targeted Approach to Suppress Tumor-Promoting Inflammation. Semin Cancer Biol. 2015, 35, 151–184. [Google Scholar] [CrossRef] [PubMed]
- Soubannier, V.; Stifani, S. NF-ΚB Signalling in Glioblastoma. Biomedicines 2017, 5, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trivedi-Parmar, V.; Jorgensen, W.L. Advances and Insights for Small Molecule Inhibition of Macrophage Migration Inhibitory Factor. J. Med. Chem. 2018, 61, 8104. [Google Scholar] [CrossRef] [PubMed]
- Missiroli, S.; Perrone, M.; Boncompagni, C.; Borghi, C.; Campagnaro, A.; Marchetti, F.; Anania, G.; Greco, P.; Fiorica, F.; Pinton, P.; et al. Targeting the NLRP3 Inflammasome as a New Therapeutic Option for Overcoming Cancer. Cancers 2021, 13, 2297. [Google Scholar] [CrossRef]
- Mohamed, I.N.; Li, L.; Ismael, S.; Ishrat, T.; El-Remessy, A.B. Thioredoxin Interacting Protein, a Key Molecular Switch between Oxidative Stress and Sterile Inflammation in Cellular Response. World J. Diabetes 2021, 12, 1979. [Google Scholar] [CrossRef]
- Hamarsheh, S.; Zeiser, R. NLRP3 Inflammasome Activation in Cancer: A Double-Edged Sword. Front. Immunol. 2020, 11, 1444. [Google Scholar] [CrossRef]
- Wang, A.; Gong, Y.; Pei, Z.; Jiang, L.; Xia, L.; Wu, Y. Paeoniflorin Ameliorates Diabetic Liver Injury by Targeting the TXNIP-Mediated NLRP3 Inflammasome in Db/Db Mice. Int. Immunopharmacol. 2022, 109, 108792. [Google Scholar] [CrossRef]
- Jo, E.K.; Kim, J.K.; Shin, D.M.; Sasakawa, C. Molecular Mechanisms Regulating NLRP3 Inflammasome Activation. Cell. Mol. Immunol. 2015, 13, 148–159. [Google Scholar] [CrossRef] [Green Version]
- Prabhu, N.; Dai, L.; Nordlund, P. CETSA in Integrated Proteomics Studies of Cellular Processes. Curr. Opin. Chem. Biol. 2020, 54, 54–62. [Google Scholar] [CrossRef]
- Ma, Z.; Chu, L.; Liu, H.; Wang, W.; Li, J.; Yao, W.; Yi, J.; Gao, Y. Beneficial Effects of Paeoniflorin on Non-Alcoholic Fatty Liver Disease Induced by High-Fat Diet in Rats. Sci. Rep. 2017, 7, 44819. [Google Scholar] [CrossRef] [Green Version]
- Xu, Z.; Wu, Y.; Wang, F.; Li, X.; Wang, P.; Li, Y.; Wu, J.; Li, Y.; Jiang, T.; Pan, X.; et al. Fibroblast Growth Factor 1 Ameliorates Diabetes-Induced Liver Injury by Reducing Cellular Stress and Restoring Autophagy. Front. Pharmacol. 2020, 11, 52. [Google Scholar] [CrossRef] [PubMed]
- Molla, M.D.; Akalu, Y.; Geto, Z.; Dagnew, B.; Ayelign, B.; Shibabaw, T. Role of Caspase-1 in the Pathogenesis of Inflammatory-Associated Chronic Noncommunicable Diseases. J. Inflamm. Res. 2020, 13, 749. [Google Scholar] [CrossRef] [PubMed]
- Lam, Y.T.; Tan, R.P.; Michael, P.; Yang, N.; Dunn, L.L.; Cooke, J.P.; Celermajer, D.S.; Wise, S.G.; Ng, M.K.C. Endothelial Thioredoxin Interacting Protein (TXNIP) Modulates Endothelium-Dependent Vasorelaxation in Hyperglycemia. Microvasc. Res. 2022, 143, 104396. [Google Scholar] [CrossRef]
- Zhan, Y.; Xu, D.; Tian, Y.; Qu, X.; Sheng, M.; Lin, Y.; Ke, M.; Jiang, L.; Xia, Q.; Kaldas, F.M.; et al. Novel Role of Macrophage TXNIP-Mediated CYLD–NRF2–OASL1 Axis in Stress-Induced Liver Inflammation and Cell Death. JHEP Rep. 2022, 4, 100532. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, R.; Nakamura, H.; Masutani, H.; Yodoi, J. Anti-Oxidative, Anti-Cancer and Anti-Inflammatory Actions by Thioredoxin 1 and Thioredoxin-Binding Protein-2. Pharmacol. Ther. 2010, 127, 261–270. [Google Scholar] [CrossRef] [PubMed]
- Zhou, R.; Tardivel, A.; Thorens, B.; Choi, I.; Tschopp, J. Thioredoxin-Interacting Protein Links Oxidative Stress to Inflammasome Activation. Nat. Immun. 2009, 11, 136–140. [Google Scholar] [CrossRef] [PubMed]
- Loveday, C.; Garrett, A.; Law, P.; Hanks, S.; Poyastro-Pearson, E.; Adlard, J.W.; Barwell, J.; Berg, J.; Brady, A.F.; Brewer, C.; et al. Analysis of Rare Disruptive Germline Mutations in 2,135 Enriched BRCA-Negative Breast Cancers Excludes Additional High-Impact Susceptibility Genes. Annals Oncol. 2022. [Google Scholar] [CrossRef]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer Statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Bower, J.E.; Kuhlman, K.R.; Ganz, P.A.; Irwin, M.R.; Crespi, C.M.; Cole, S.W. Childhood Maltreatment and Monocyte Gene Expression among Women with Breast Cancer. Brain Behav. Immun. 2020, 88, 396–402. [Google Scholar] [CrossRef]
- Karmi, O.; Sohn, Y.S.; Zandalinas, S.I.; Rowland, L.; King, S.D.; Nechushtai, R.; Mittler, R. Disrupting CISD2 Function in Cancer Cells Primarily Impacts Mitochondrial Labile Iron Levels and Triggers TXNIP Expression. Free Radic Biol. Med. 2021, 176, 92–104. [Google Scholar] [CrossRef]
- Stoltzman, C.A.; Kaadige, M.R.; Peterson, C.W.; Ayer, D.E. MondoA Senses Non-Glucose Sugars: Regulation of Thioredoxin-Interacting Protein (TXNIP) and the Hexose Transport Curb. J. Biol. Chem. 2011, 286, 38027–38034. [Google Scholar] [CrossRef] [PubMed]
- Alhawiti, N.M.; al Mahri, S.; Aziz, M.A.; Malik, S.S.; Mohammad, S. TXNIP in Metabolic Regulation: Physiological Role and Therapeutic Outlook. Curr. Drug Targets 2017, 18, 1095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, L.; O’Shea, J.M.; Kaadige, M.R.; Cunha, S.; Wilde, B.R.; Cohen, A.L.; Welm, A.L.; Ayer, D.E. Metabolic Reprogramming in Triple-Negative Breast Cancer through Myc Suppression of TXNIP. Proc. Natl. Acad. Sci. USA 2015, 112, 5425–5430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, X.; Sun, J.; Zhang, Y.; Li, J.; Hu, J.; Li, K.; Gao, L.; Shen, L. C-Myc-Driven Glycolysis via TXNIP Suppression Is Dependent on Glutaminase-MondoA Axis in Prostate Cancer. Biochem. Biophys Res. Commun. 2018, 504, 415–421. [Google Scholar] [CrossRef]
- Miller, K.D.; Nogueira, L.; Devasia, T.; Mariotto, A.B.; Yabroff, K.R.; Jemal, A.; Kramer, J.; Siegel, R.L. Cancer Treatment and Survivorship Statistics, 2022. CA Cancer J. Clin. 2022, 72, 409–436. [Google Scholar] [CrossRef]
- Kopantzev, E.P.; Monastyrskaya, G.S.; Vinogradova, T.V.; Zinovyeva, M.V.; Kostina, M.B.; Filyukova, O.B.; Tonevitsky, A.G.; Sukhikh, G.T.; Sverdlov, E.D. Differences in Gene Expression Levels between Early and Later Stages of Human Lung Development Are Opposite to Those between Normal Lung Tissue and Non-Small Lung Cell Carcinoma. Lung Cancer 2008, 62, 23–34. [Google Scholar] [CrossRef]
- Jin, X.; Wu, N.; Dai, J.; Li, Q.; Xiao, X.Q. TXNIP Mediates the Differential Responses of A549 Cells to Sodium Butyrate and Sodium 4-phenylbutyrate Treatment. Cancer Med. 2017, 6, 424. [Google Scholar] [CrossRef]
- Xiao, X.; Xu, Y.; Chen, H. Sodium Butyrate-Activated TRAF6-TXNIP Pathway Affects A549 Cells Proliferation and Migration. Cancer Med. 2020, 9, 3477–3488. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, A.M.; Martel, F. Targeting Glucose Transporters for Breast Cancer Therapy: The Effect of Natural and Synthetic Compounds. Cancers 2020, 12, 154. [Google Scholar] [CrossRef] [Green Version]
- Alabi, B.R.; Liu, S.; Stoyanova, T. Current and Emerging Therapies for Neuroendocrine Prostate Cancer. Pharmacol. Ther. 2022, 238, 108255. [Google Scholar] [CrossRef]
- Wei, M.; Jiao, D.; Han, D.; Wu, J.; Wei, F.; Zheng, G.; Guo, Z.; Xi, W.; Yang, F.; Xie, P.; et al. Knockdown of RNF2 Induces Cell Cycle Arrest and Apoptosis in Prostate Cancer Cells through the Upregulation of TXNIP. Oncotarget 2017, 8, 5323. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, A.D.; Andersen, J.G.; Gyawali, B.; Shrestha, A.; Shrestha, S.; Neupane, D.; Ghimire, S.; Campbell, C.; Kallestrup, P. Cervical Cancer Screening Utilization, and Associated Factors, in Nepal: A Systematic Review and Meta-Analysis. Public Health 2022, 210, 16–25. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Lee, H.J.; Choi, M.Y.; Kang, S.S.; Kim, Y.S.; Shin, J.K.; Choi, W.S. UHRF1 Induces Methylation of the TXNIP Promoter and Down-Regulates Gene Expression in Cervical Cancer. Mol. Cells 2021, 44, 146. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Tian, X.; Yin, H.; Xiao, S.; Yi, S.; Zhang, Y.; Zeng, F. TXNIP Induced by MondoA, Rather than ChREBP, Suppresses Cervical Cancer Cell Proliferation, Migration and Invasion. J. Biochem. 2020, 167, 371–377. [Google Scholar] [CrossRef]
- Zhang, Z.; Wang, S.; Yang, F.; Meng, Z.; Liu, Y. LncRNA ROR1-AS1 High Expression and Its Prognostic Significance in Liver Cancer. Oncol. Rep. 2020, 43, 55–74. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Li, W.; Xin, X.; Li, X.; Geng, J.; Sun, Y. Exosomes Secreted by M2 Macrophages Promote Cancer Stemness of Hepatocellular Carcinoma via the MiR-27a-3p/TXNIP Pathways. Int. Immunopharmacol. 2021, 101, 107585. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, Q.; Gong, L.; Xu, H.; Liu, B.; Fang, X.; Yu, D.; Li, L.; Wei, T.; Wang, Y.; et al. C-Terminal Truncated HBx Initiates Hepatocarcinogenesis by Downregulating TXNIP and Reprogramming Glucose Metabolism. Oncogene 2021, 40, 1147. [Google Scholar] [CrossRef]
- Wu, J.; Xiao, Y.; Sun, J.; Sun, H.; Chen, H.; Zhu, Y.; Fu, H.; Yu, C.; Lai, S.; Ma, L.; et al. A Single-Cell Survey of Cellular Hierarchy in Acute Myeloid Leukemia. J. Hematol. Oncol. 2020, 13, 1–19. [Google Scholar] [CrossRef]
- Noura, M.; Matsuo, H.; Koyama, A.; Adachi, S.; Masutani, H. TXNIP Induces Growth Arrest and Enhances ABT263-induced Apoptosis in Mixed-lineage Leukemia-rearranged Acute Myeloid Leukemia Cells. FEBS Open Bio 2020, 10, 1532. [Google Scholar] [CrossRef]
- Dinesh, P.; Rasool, M. Multifaceted Role of IL-21 in Rheumatoid Arthritis: Current Understanding and Future Perspectives. J. Cell Physiol. 2018, 233, 3918–3928. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.; Kershaw, N.J.; Babon, J.J. The Molecular Details of Cytokine Signaling via the JAK/STAT Pathway. Protein Sci. 2018, 27, 1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Y.; Li, H.; Deng, Y.; Tai, Y.; Zeng, K.; Zhang, Y.; Liu, W.; Zhang, Q.; Yang, Y. Cancer-Associated Fibroblasts Induce PDL1+ Neutrophils through the IL6-STAT3 Pathway That Foster Immune Suppression in Hepatocellular Carcinoma. Cell Death Disease 2018, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; di Trapani, G.; Tonissen, K.F. Expanding the Armory for Treating Lymphoma: Targeting Redox Cellular Status through Thioredoxin Reductase Inhibition. Pharmacol. Res. 2022, 177, 106134. [Google Scholar] [CrossRef]
- Pan, Y.R.; Chen, C.C.; Chan, Y.T.; Wang, H.J.; Chien, F.T.; Chen, Y.L.; Liu, J.L.; Yang, M.H. STAT3-Coordinated Migration Facilitates the Dissemination of Diffuse Large B-Cell Lymphomas. Nat. Commun. 2018, 9, 3696. [Google Scholar] [CrossRef] [Green Version]
- Busker, S.; Qian, W.; Haraldsson, M.; Espinosa, B.; Johansson, L.; Attarha, S.; Kolosenko, I.; Liu, J.; Dagnell, M.; Grandér, D.; et al. Irreversible TrxR1 Inhibitors Block STAT3 Activity and Induce Cancer Cell Death. Sci. Adv. 2020, 6, eaax7945. [Google Scholar] [CrossRef] [Green Version]
- Nakaya, A.; Sagawa, M.; Muto, A.; Uchida, H.; Ikeda, Y.; Kizaki, M. The Gold Compound Auranofin Induces Apoptosis of Human Multiple Myeloma Cells through Both Down-Regulation of STAT3 and Inhibition of NF-ΚB Activity. Leuk. Res. 2011, 35, 243–249. [Google Scholar] [CrossRef]
- Hasan, A.A.; Kalinina, E.; Tatarskiy, V.; Shtil, A. The Thioredoxin System of Mammalian Cells and Its Modulators. Biomedicines 2022, 10, 1757. [Google Scholar] [CrossRef]
- Sagwal, S.; Prasad, R.; Kaur, J.; Singh, M. Cytokine Signaling Pathway in Cystic Fibrosis: Expression of SOCS and STATs Genes in Different Clinical Phenotypes of the Disease. Mol. Cell. Biochem. 2021, 476, 2869–2876. [Google Scholar] [CrossRef]
- Chiarugi, P.; Cirri, P. Redox Regulation of Protein Tyrosine Phosphatases during Receptor Tyrosine Kinase Signal Transduction. Trends Biochem. Sci. 2003, 28, 509–514. [Google Scholar] [CrossRef]
- Sivaganesh, V.; Sivaganesh, V.; Scanlon, C.; Iskander, A.; Maher, S.; Lê, T.; Peethambaran, B. Protein Tyrosine Phosphatases: Mechanisms in Cancer. Int. J. Mol. Sci. 2021, 22, 12865. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, C.; Sims, M.M.; Sacher, J.R.; Raje, M.; Deokar, H.; Yue, P.; Turkson, J.; Buolamwini, J.K.; Pfeffer, L.M. SS-4 Is a Highly Selective Small Molecule Inhibitor of STAT3 Tyrosine Phosphorylation That Potently Inhibits GBM Tumorigenesis in Vitro and in Vivo. Cancer Lett. 2022, 533, 215614. [Google Scholar] [CrossRef] [PubMed]
- Park, J.W.; Lee, S.H.; Woo, G.H.; Kwon, H.J.; Kim, D.Y. Downregulation of TXNIP Leads to High Proliferative Activity and Estrogen-Dependent Cell Growth in Breast Cancer. Biochem. Biophys. Res. Commun. 2018, 498, 566–572. [Google Scholar] [CrossRef] [PubMed]
- de Padua, M.C.; Delodi, G.; Vucetic, M.; Durivault, J.; Vial, V.; Bayer, P.; Noleto, G.R.; Mazure, N.M.; Ždralevic, M.; Pouysségur, J. Disrupting Glucose-6-Phosphate Isomerase Fully Suppresses the “Warburg Effect” and Activates OXPHOS with Minimal Impact on Tumor Growth except in Hypoxia. Oncotarget 2017, 8, 87623–87637. [Google Scholar] [CrossRef] [Green Version]
- Neumann, D.; Woods, A.; Carling, D.; Wallimann, T.; Schlattner, U. Mammalian AMP-Activated Protein Kinase: Functional, Heterotrimeric Complexes by Co-Expression of Subunits in Escherichia Coli. Protein Expr. Purif. 2003, 30, 230–237. [Google Scholar] [CrossRef]
- Marcelo, K.L.; Means, A.R.; York, B. The Ca2+/Calmodulin/CaMKK2 Axis: Nature’s Metabolic CaMshaft. Trends Endocrinol. Metab. 2016, 27, 706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abu el Maaty, M.A.; Wölfl, S. Vitamin D as a Novel Regulator of Tumor Metabolism: Insights on Potential Mechanisms and Implications for Anti-Cancer Therapy. Int. J. Mol. Sci. 2017, 18, 2184. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Wang, H.; Geng, X.; Zhang, D.; Zhu, Z.; Zhang, G.; Hou, J. Metformin Exerts Anti-AR-Negative Prostate Cancer Activity via AMPK/Autophagy Signaling Pathway. Cancer Cell Int. 2021, 21, 1–12. [Google Scholar] [CrossRef]
- Khan, A.S.; Frigo, D.E. Regulation, Role and Therapeutic Targeting of AMPK in Prostate Cancer. Nat. Rev. Urol. 2017, 14, 164. [Google Scholar] [CrossRef] [Green Version]
- Sadria, M.; Seo, D.; Layton, A.T. The Mixed Blessing of AMPK Signaling in Cancer Treatments. BMC Cancer 2022, 22, 105. [Google Scholar] [CrossRef]
- Garza-Lombó, C.; Schroder, A.; Reyes-Reyes, E.M.; Franco, R. MTOR/AMPK Signaling in the Brain: Cell Metabolism, Proteostasis and Survival. Curr. Opin. Toxicol. 2018, 8, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Tian, T.; Li, X.; Zhang, J. MTOR Signaling in Cancer and MTOR Inhibitors in Solid Tumor Targeting Therapy. Int. J. Mol. Sci. 2019, 20, 755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almouhanna, F.; Blagojevic, B.; Can, S.; Ghanem, A.; Wölfl, S. Pharmacological Activation of Pyruvate Kinase M2 Reprograms Glycolysis Leading to TXNIP Depletion and AMPK Activation in Breast Cancer Cells. Cancer Metab. 2021, 9, 1–17. [Google Scholar] [CrossRef]
- Abu el Maaty, M.A.; Alborzinia, H.; Khan, S.J.; Büttner, M.; Wölfl, S. 1,25(OH)2D3 Disrupts Glucose Metabolism in Prostate Cancer Cells Leading to a Truncation of the TCA Cycle and Inhibition of TXNIP Expression. Biochim. Biophys. Acta BBA Mol. Cell Res. 2017, 1864, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Qayyum, N.; Haseeb, M.; Kim, M.S.; Choi, S. Role of Thioredoxin-Interacting Protein in Diseases and Its Therapeutic Outlook. Int. J. Mol. Sci. 2021, 22, 2754. [Google Scholar] [CrossRef]
- Kaadige, M.R.; Yang, J.; Wilde, B.R.; Ayer, D.E. MondoA-Mlx Transcriptional Activity Is Limited by MTOR-MondoA Interaction. Mol. Cell Biol. 2015, 35, 101–110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal. Trans. Target. Ther. 2017, 2, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Koundouros, N.; Poulogiannis, G. Phosphoinositide 3-Kinase/Akt Signaling and Redox Metabolism in Cancer. Front. Oncol. 2018, 8, 160. [Google Scholar] [CrossRef]
- Akins, N.S.; Nielson, T.C.; Le, H.V. Inhibition of Glycolysis and Glutaminolysis: An Emerging Drug Discovery Approach to Combat Cancer. Curr. Top. Med. Chem. 2018, 18, 494. [Google Scholar] [CrossRef]
- Waldhart, A.N.; Dykstra, H.; Peck, A.S.; Boguslawski, E.A.; Madaj, Z.B.; Wen, J.; Veldkamp, K.; Hollowell, M.; Zheng, B.; Cantley, L.C.; et al. Phosphorylation of TXNIP by AKT Mediates Acute Influx of Glucose in Response to Insulin. Cell Rep. 2017, 19, 2005. [Google Scholar] [CrossRef] [Green Version]
- Hoxhaj, G.; Manning, B.D. The PI3K-AKT Network at the Interface of Oncogenic Signalling and Cancer Metabolism. Nat. Rev. Cancer 2020, 20, 74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Fu, T.; He, Q.; Gao, X.; Luo, Y. Glucose-6-Phosphate Upregulates Txnip Expression by Interacting With MondoA. Front. Mol. Biosci. 2020, 6, 147. [Google Scholar] [CrossRef] [PubMed]
- Ye, Z.; Ayer, D.E. Ras Suppresses TXNIP Expression by Restricting Ribosome Translocation. Mol. Cell Biol. 2018, 38, e00178-18. [Google Scholar] [CrossRef] [Green Version]
- Nishizawa, K.; Nishiyama, H.; Matsui, Y.; Kobayashi, T.; Saito, R.; Kotani, H.; Masutani, H.; Oishi, S.; Toda, Y.; Fujii, N.; et al. Thioredoxin-Interacting Protein Suppresses Bladder Carcinogenesis. Carcinogenesis 2011, 32, 1459–1466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hong, S.Y.; Yu, F.X.; Luo, Y.; Hagen, T. Oncogenic Activation of the PI3K/Akt Pathway Promotes Cellular Glucose Uptake by Downregulating the Expression of Thioredoxin-Interacting Protein. Cell Signal. 2016, 28, 377–383. [Google Scholar] [CrossRef]
- Liu, R.X.; Tang, W.; Zheng, B.Y.; Yang, Y.; Li, Z.Y.; Gui, T.; Zhang, H.T.; Liu, N.; Zha, Z.G.; Li, J.X. YAP/MiR-524-5p Axis Negatively Regulates TXNIP Expression to Promote Chondrosarcoma Cell Growth. Biochem. Biophys. Res. Commun. 2022, 590, 20–26. [Google Scholar] [CrossRef] [PubMed]
- He, G.; Chen, K.; Wang, H.; Li, X.; Li, W.; liu, L.; Chen, J.; Yang, D.; Hu, J.; Xu, D.; et al. Fudosteine Attenuates Acute Lung Injury in Septic Mice by Inhibiting Pyroptosis via the TXNIP/NLRP3/GSDMD Pathway. Eur J. Pharmacol. 2022, 926, 175047. [Google Scholar] [CrossRef]
- Pi, M.; Kuang, H.; Yue, C.; Yang, Q.; Wu, A.; Li, Y.; Assaraf, Y.G.; Yang, D.H.; Wu, S. Targeting Metabolism to Overcome Cancer Drug Resistance: A Promising Therapeutic Strategy for Diffuse Large B Cell Lymphoma. Drug Resistance Updates 2022, 61, 100822. [Google Scholar] [CrossRef]
- Spagnol, V.; Oliveira, C.A.B.; Randle, S.J.; Passos, P.M.S.; Correia, C.R.S.T.B.; Simaroli, N.B.; Oliveira, J.S.; Mevissen, T.E.T.; Medeiros, A.C.; Gomes, M.D.; et al. The E3 Ubiquitin Ligase SCF(Fbxo7) Mediates Proteasomal Degradation of UXT Isoform 2 (UXT-V2) to Inhibit the NF-ΚB Signaling Pathway. Biochim. Biophys. BBA General Subjects 2021, 1865, 129754. [Google Scholar] [CrossRef]
- Chen, Z.; Lai, X.; Ding, H.; Zhang, A.; Sun, Y.; Ling, J.; Chiao, P.J.; Chen, Z.; Xia, X. ATF4/TXNIP/REDD1/MTOR Signaling Mediates the Antitumor Activities of Liver X Receptor in Pancreatic Cancers. Cancer Innovat. 2022, 1, 55–69. [Google Scholar] [CrossRef]
- Cheng, Z.; Lu, C.; Wang, H.; Wang, N.; Cui, S.; Yu, C.; Wang, C.; Zuo, Q.; Wang, S.; Lv, Y.; et al. Long Noncoding RNA LHFPL3-AS2 Suppresses Metastasis of Non-Small Cell Lung Cancer by Interacting with SFPQ to Regulate TXNIP Expression. Cancer Lett. 2022, 531, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.; Lin, L.; Lin, H.; Chen, W.; Chen, L.; Chen, X.; Chen, S.; Lin, Q.; Xu, Y.; Zeng, Y. KCNK3 Inhibits Proliferation and Glucose Metabolism of Lung Adenocarcinoma via Activation of AMPK-TXNIP Pathway. Cell Death Discov. 2022, 8, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Kim, G.T.; Kim, E.Y.; Shin, S.H.; Lee, H.; Lee, S.H.; Sohn, K.Y.; Kim, J.W. Suppression of Tumor Progression by Thioredoxin-Interacting Protein-Dependent Adenosine 2B Receptor Degradation in a PLAG-Treated Lewis Lung Carcinoma-1 Model of Non–Small Cell Lung Cancer. Neoplasia 2022, 31, 100815. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Pan, J.; Liu, Y.; Luo, X.; Yang, C.; Xiao, W.; Li, Q.; Yang, L.; Zhang, X. 3-Bromopyruvic Acid Regulates Glucose Metabolism by Targeting the C-Myc/TXNIP Axis and Induces Mitochondria-mediated Apoptosis in TNBC Cells. Exp. Ther. Med. 2022, 24, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wan, Y.; He, T. Circ_SAR1A Regulates the Malignant Behavior of Lung Cancer Cells via the MiR-21-5p/TXNIP Axis. J. Clin. Lab. Anal. 2022, 36, e24366. [Google Scholar] [CrossRef]
- Wang, X.; Yin, Y.; Qian, W.; Peng, C.; Shen, S.; Wang, T.; Zhao, S. Citric Acid of Ovarian Cancer Metabolite Induces Pyroptosis via the Caspase-4/TXNIP-NLRP3-GSDMD Pathway in Ovarian Cancer. FASEB J. 2022, 36, e22362. [Google Scholar] [CrossRef]
- Yuan, Y.; Liu, Q.; Wu, Z.; Zhong, W.; Lin, Z.; Luo, W. TXNIP Inhibits the Progression of Osteosarcoma through DDIT4-Mediated MTORC1 Suppression. Am. J. Cancer Res. 2022, 12, 3760. [Google Scholar]
- Chung, Y.J.; Salvi, A.; Kalailingam, P.; Alnawaz, M.; Tan, S.H.; Pan, J.Y.; Tan, N.S.; Thanabalu, T. N-WASP Attenuates Cell Proliferation and Migration through ERK2-Dependent Enhanced Expression of TXNIP. Biology 2022, 11, 582. [Google Scholar] [CrossRef]
- Jing, Y.J.; Lin, L.C.; Chen, L.L.; Huang, Z.E.; Qin, H.C.; Li, S.B.; Chen, Z.H. WT1 Inhibits Human Renal Carcinoma Cell Proliferation and Induces G2/M Arrest by Upregulating IL-24 Expression. Biomed. Res. Int 2022, 2022, 1093945. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, H.; Chen, B.; Mao, Q.; Xia, W.; Zhang, T.; Song, X.; Zhang, Z.; Xu, L.; Dong, G.; et al. CircDCUN1D4 Suppresses Tumor Metastasis and Glycolysis in Lung Adenocarcinoma by Stabilizing TXNIP Expression. Mol. Ther. Nucl. Acids 2021, 23, 355–368. [Google Scholar] [CrossRef]
- Gong, J.; Ma, L.; Peng, C.; Liu, J. LncRNA MAGI2-AS3 Acts as a Tumor Suppressor That Attenuates Non-Small Cell Lung Cancer Progression by Targeting the MiR-629-5p/TXNIP Axis. Ann. Transl. Med. 2021, 9, 1793. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, M.A.; Chattopadhyay, S.; Siddiqui, F.A.; Ur Rehman, A.; Siddiqui, S.; Prakasam, G.; Khan, A.; Sultana, S.; Bamezai, R.N.K. Silibinin Induces Metabolic Crisis in Triple-Negative Breast Cancer Cells by Modulating EGFR-MYC-TXNIP Axis: Potential Therapeutic Implications. FEBS J. 2021, 288, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Jin, Y.; Liu, F.; Jiang, J.; Cao, J.; Lu, Y.; Yang, J. Ceramide Induces the Apoptosis of Non-Small Cell Lung Cancer Cells through the Txnip/Trx1 Complex. Int J. Mol. Med. 2021, 47, 85. [Google Scholar] [CrossRef] [PubMed]
- Zhu, J.; Han, S. Histone Deacetylase 10 Exerts Antitumor Effects on Cervical Cancer via a Novel MicroRNA-223/TXNIP/Wnt/β-Catenin Pathway. IUBMB Life 2021, 73, 690–704. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Liu, T.; Bao, Y.; Zhao, T.; Wang, J.; Wang, H.; Wang, A.; Gan, X.; Wu, Z.; Wang, L. CircRNA CRAPGEF5 Inhibits the Growth and Metastasis of Renal Cell Carcinoma via the MiR-27a-3p/TXNIP Pathway. Cancer Lett. 2020, 469, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.H.; Chen, Y.H.; Shen, W.H. Amikacin Suppresses Human Breast Cancer Cell MDA-MB-231 Migration and Invasion. Toxics 2020, 8, 108. [Google Scholar] [CrossRef]
- Kim, Y.; Kim, Y.S.; Kim, M.; Kim, J.M.; Lee, H.H.; Kim, T.H. Thioredoxin-Interacting Protein (TXNIP) Mediates Thioredoxin-Dependent Antioxidant Mechanism in Endometrial Cancer Cells Treated With 1α,25-Dihydroxyvitamin D3. Anticancer Res. 2019, 39, 4795–4803. [Google Scholar] [CrossRef]
- Wang, X.; Nachliely, M.; Harrison, J.S.; Danilenko, M.; Studzinski, G.P. Participation of vitamin d-upregulated protein 1 (txnip)-ask1-jnk1 signalosome in the enhancement of aml cell death by a post-cytotoxic differentiation regimen. J. Steroid Biochem. Mol. Biol. 2019, 187, 166. [Google Scholar] [CrossRef]
- Hoshikawa, H.; Kamitori, K.; Indo, K.; Mori, T.; Kamata, M.; Takahashi, T.; Tokuda, M. Combined Treatment with D-Allose, Docetaxel and Radiation Inhibits the Tumor Growth in an in Vivo Model of Head and Neck Cancer. Oncol. Lett. 2018, 15, 3422. [Google Scholar] [CrossRef] [Green Version]
- Sherbet, G.V. Tumour Suppression and Inhibition of Angiogenesis by TXNIP (Thioredoxin Interacting Protein). Mol. Approach Cancer Manag. 2017, 101–103. [Google Scholar] [CrossRef]
- Yin, H.; Yu, S.; Xie, Y.; Dai, X.; Dong, M.; Sheng, C.; Hu, J. Cancer-Associated Fibroblasts-Derived Exosomes Upregulate MicroRNA-135b-5p to Promote Colorectal Cancer Cell Growth and Angiogenesis by Inhibiting Thioredoxin-Interacting Protein. Cell Signal. 2021, 84, 110029. [Google Scholar] [CrossRef] [PubMed]
- Meszaros, M.; Yusenko, M.; Domonkos, L.; Peterfi, L.; Kovacs, G.; Banyai, D. Expression of TXNIP Is Associated with Angiogenesis and Postoperative Relapse of Conventional Renal Cell Carcinoma. Sci. Rep. 2021, 11, 17200. [Google Scholar] [CrossRef] [PubMed]
- Kuljaca, S.; Liu, T.; Dwarte, T.; Kavallaris, M.; Haber, M.; Norris, M.D.; Martin-Caballero, J.; Marshall, G.M. The Cyclin-Dependent Kinase Inhibitor, P21(WAF1), Promotes Angiogenesis by Repressing Gene Transcription of Thioredoxin-Binding Protein 2 in Cancer Cells. Carcinogenesis 2009, 30, 1865–1871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- le Jan, S.; le Meur, N.; Cazes, A.; Philippe, J.; le Cunff, M.; Léger, J.; Corvol, P.; Germain, S. Characterization of the Expression of the Hypoxia-Induced Genes Neuritin, TXNIP and IGFBP3 in Cancer. FEBS Lett. 2006, 580, 3395–3400. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| S.no | Compound Name | Compound Nature | Type of Cancer | Target/Mechanism | Model | Reference |
|---|---|---|---|---|---|---|
| 1. | GW3965 | LXR agonist | Pancreatic cancer | ATF4/TXNIP/REDD1/mTOR signaling | Cell lines MIA PaCa-2, BXPC3, hTERT-HPNE | [121] |
| 2. | LHFPL3-AS2 | Long non-coding RNA | Non-small cell Lung cancer | SFPQ-mediated transcriptional repression | BALB/c-nude mice | [122] |
| 3. | KCNK3 | Protein Coding gene | Lung adenocarcinoma | AMPK-TXNIP pathway | LUAD cell lines | [123] |
| 4. | PLAG | Chemical synthesis | LLC1 | TXNIP expression degrading A2BR | C57BL/6J mice LLC1-implanted TXNIP-KO | [124] |
| 5. | 3-BrPA | Organic compound | TNBC | c-Myc/TXNIP axis | TNBC (HCC1143) cell lines | [125] |
| 6. | circ_SAR1A | Gene | Lung cancer | miR-21-5p/TXNIP axis | Lung cancer cell lines H1650, H1581, H460, H1299, and A549 | [126] |
| 7. | Citric acid | Organic compound | Ovarian cancer | CASP4/TXNIP-NLRP3-Gesdermin-d (GSDMD) pathway | Ovarian cancer cells | [127] |
| 8. | DDIT4 | Gene | Osteosarcoma | TXNIP/DDIT4/mTORC1 axis | Cell lines 143B, HOS, MG63, U2OS, and hFOB1 | [128] |
| 9. | N-WASP | Protein | squamous cell carcinoma | N-WASP-ERK2-FOXO1-TXNIP pathway | Cell line HSC-5 | [129] |
| 10. | WT1 | Protein | Renal cell carcinoma | WT1/IL-24/TXNIP axis | Cell line A498 | [130] |
| 11. | circDCUN1D4 | Gene | Lung adenocarcinoma | circDCUN1D4/HuR/TXNIP RNA-protein ternary complex | Cell lines HBE, A549, H1299, H1975, PC-9, and SPCA-1 | [131] |
| 12. | MAGI2-AS3 | Long non-coding RNA | Non-small cell Lung cancer | miR-629-5p/TXNIP axis | Cell lines Beas-2B, A549, and H1299 | [132] |
| 13. | Silibinin | flavonoid | TNBC | EGFR-MYC-TXNIP axis | Cell line MDA-MB-231, MDA-MB-468, BT549, BT474, and SKBR3 | [133] |
| 14. | Ceramide | Lipid molecules | Non-small-cell lung cancer | Txnip/Trx1 complex | Cell lines A549 and PC9 | [134] |
| 15. | HDAC10 | Enzyme | Cervical cancer | microRNA-223/TXNIP/Wnt/β-catenin pathway | CC cells C33A, MS751, HeLa, SiHa, and End1/E6E7 | [135] |
| 16. | cRAPGEF5 | Circular RNA | Renal cell carcinoma | miR-27a-3p/TXNIP pathway | Cell lines 769-P, Caki-1, OSRC-2, and 786-O | [136] |
| 17. | Amikacin | Antibiotic | Breast Cancer | TXNIP | cell line MDA-MB-231 | [137] |
| 18. | Vitamin D3 | Fat-soluble vitamin | Endometrial cancer | TXNIP | Human endometrial cancer cell line | [138] |
| 19. | cytarabine | Nucleoside analog | Acute myeloid leukemia | TXNIP-ASK1-JNK1 axis | Cell lines, HL60 and U937 | [139] |
| 20. | D-allose + docetaxel | Sugar + semi-synthetic drug | Head and neck cancer | TXNIP | Cell line HSC3 | [140] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Katturajan, R.; Nithiyanandam, S.; Parthasarathy, M.; Valsala Gopalakrishnan, A.; Sathiyamoorthi, E.; Lee, J.; Ramesh, T.; Iyer, M.; Prince, S.E.; Ganesan, R. Immunomodulatory Role of Thioredoxin Interacting Protein in Cancer’s Impediments: Current Understanding and Therapeutic Implications. Vaccines 2022, 10, 1902. https://doi.org/10.3390/vaccines10111902
Katturajan R, Nithiyanandam S, Parthasarathy M, Valsala Gopalakrishnan A, Sathiyamoorthi E, Lee J, Ramesh T, Iyer M, Prince SE, Ganesan R. Immunomodulatory Role of Thioredoxin Interacting Protein in Cancer’s Impediments: Current Understanding and Therapeutic Implications. Vaccines. 2022; 10(11):1902. https://doi.org/10.3390/vaccines10111902
Chicago/Turabian StyleKatturajan, Ramkumar, Sangeetha Nithiyanandam, Manisha Parthasarathy, Abilash Valsala Gopalakrishnan, Ezhaveni Sathiyamoorthi, Jintae Lee, Thiyagarajan Ramesh, Mahalaxmi Iyer, Sabina Evan Prince, and Raja Ganesan. 2022. "Immunomodulatory Role of Thioredoxin Interacting Protein in Cancer’s Impediments: Current Understanding and Therapeutic Implications" Vaccines 10, no. 11: 1902. https://doi.org/10.3390/vaccines10111902