
Overcoming Aging-Associated Poor Influenza Vaccine Responses with CpG 1018 Adjuvant


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Mice
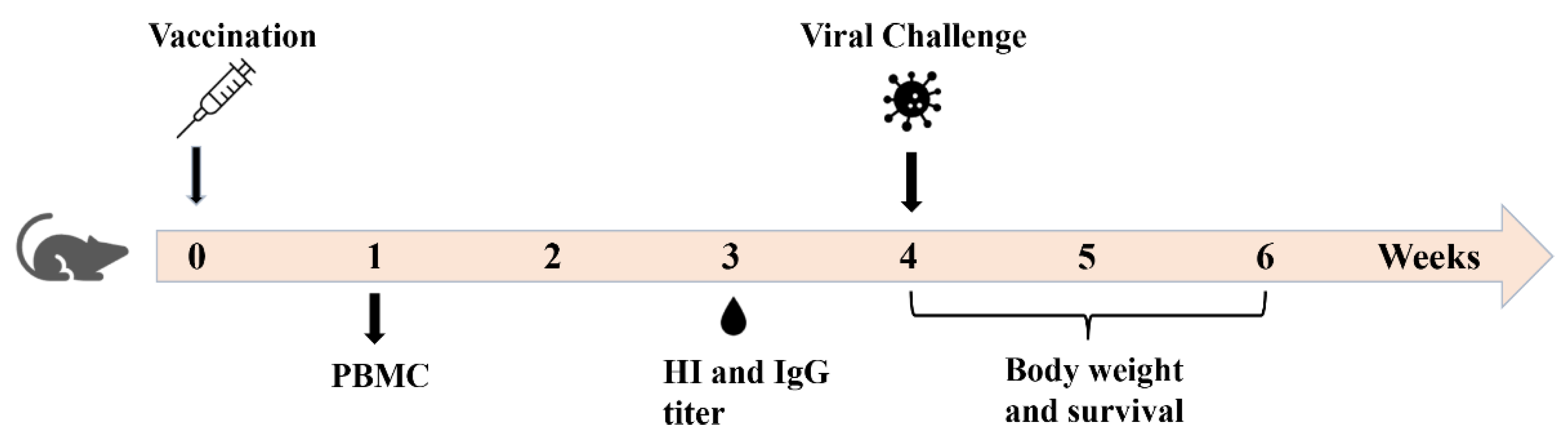
2.3. Immunization
2.4. Hemagglutination Inhibition (HI) Titer
2.5. ELISA Antibody Titer
2.6. Cell-Mediated Immune Responses
2.7. Lethal Viral Challenge
2.8. Statistics
3. Results
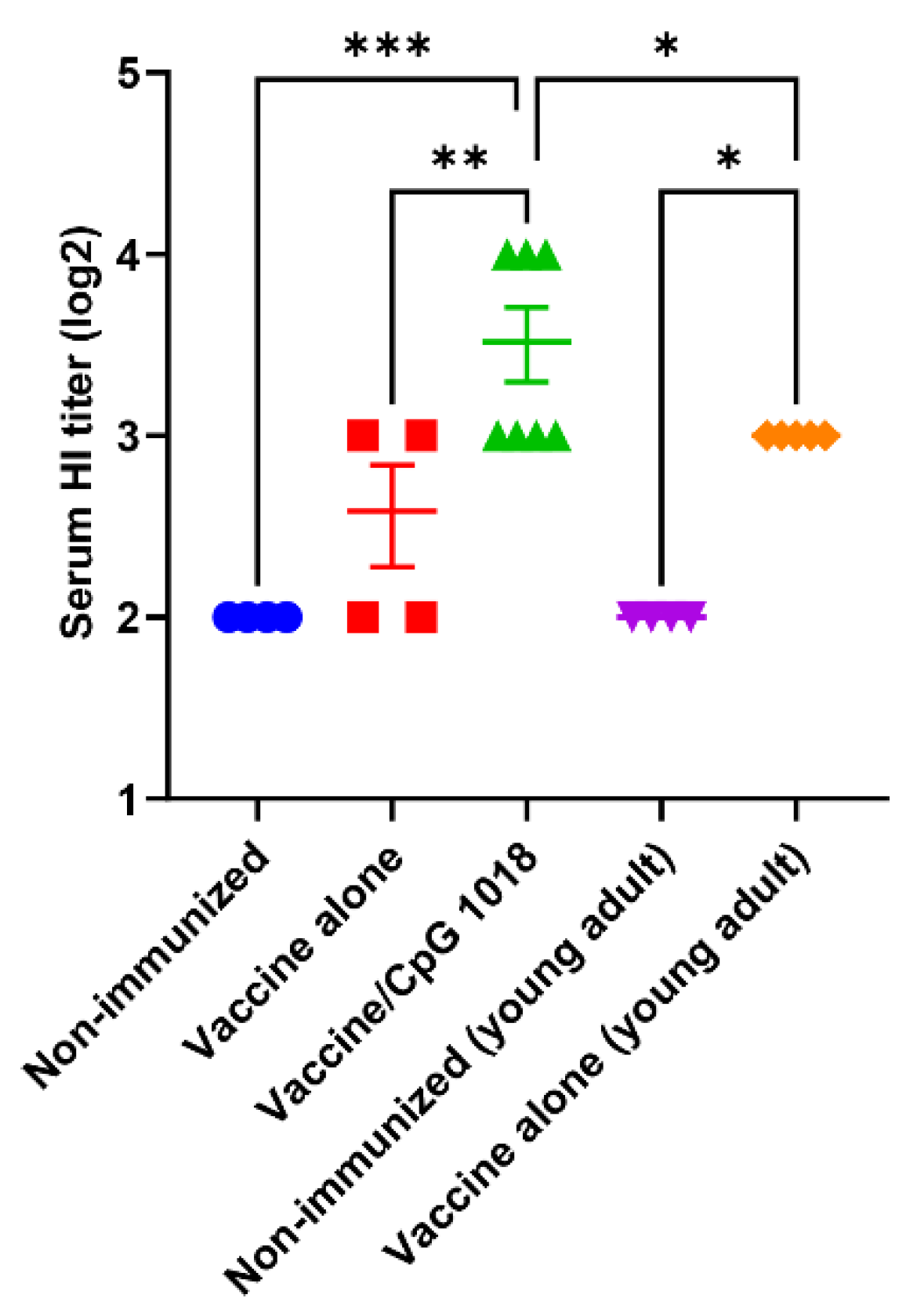
3.1. CpG 1018 Enhances HI Titer
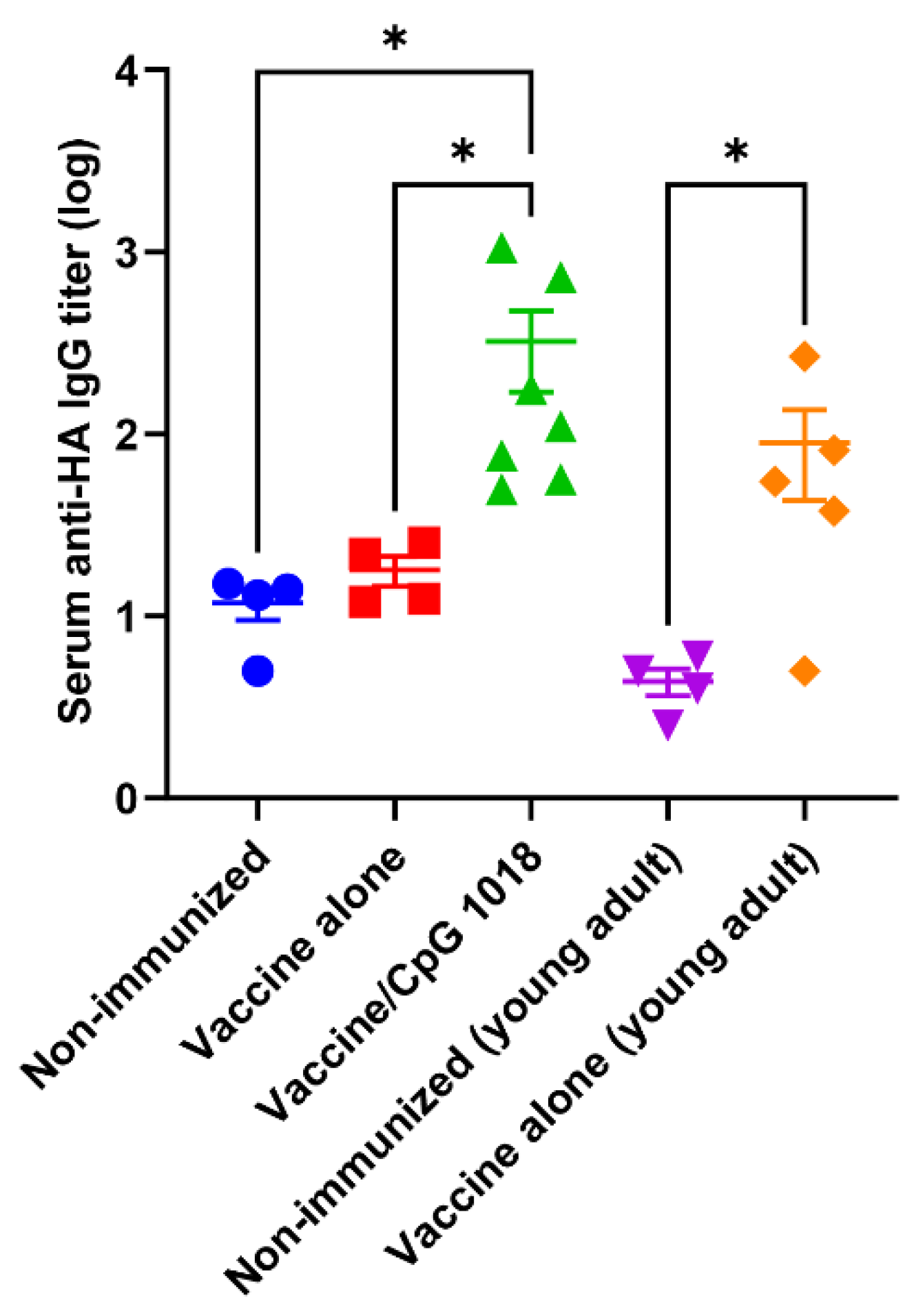
3.2. CpG 1018 Enhances ELISA Antibody Titer
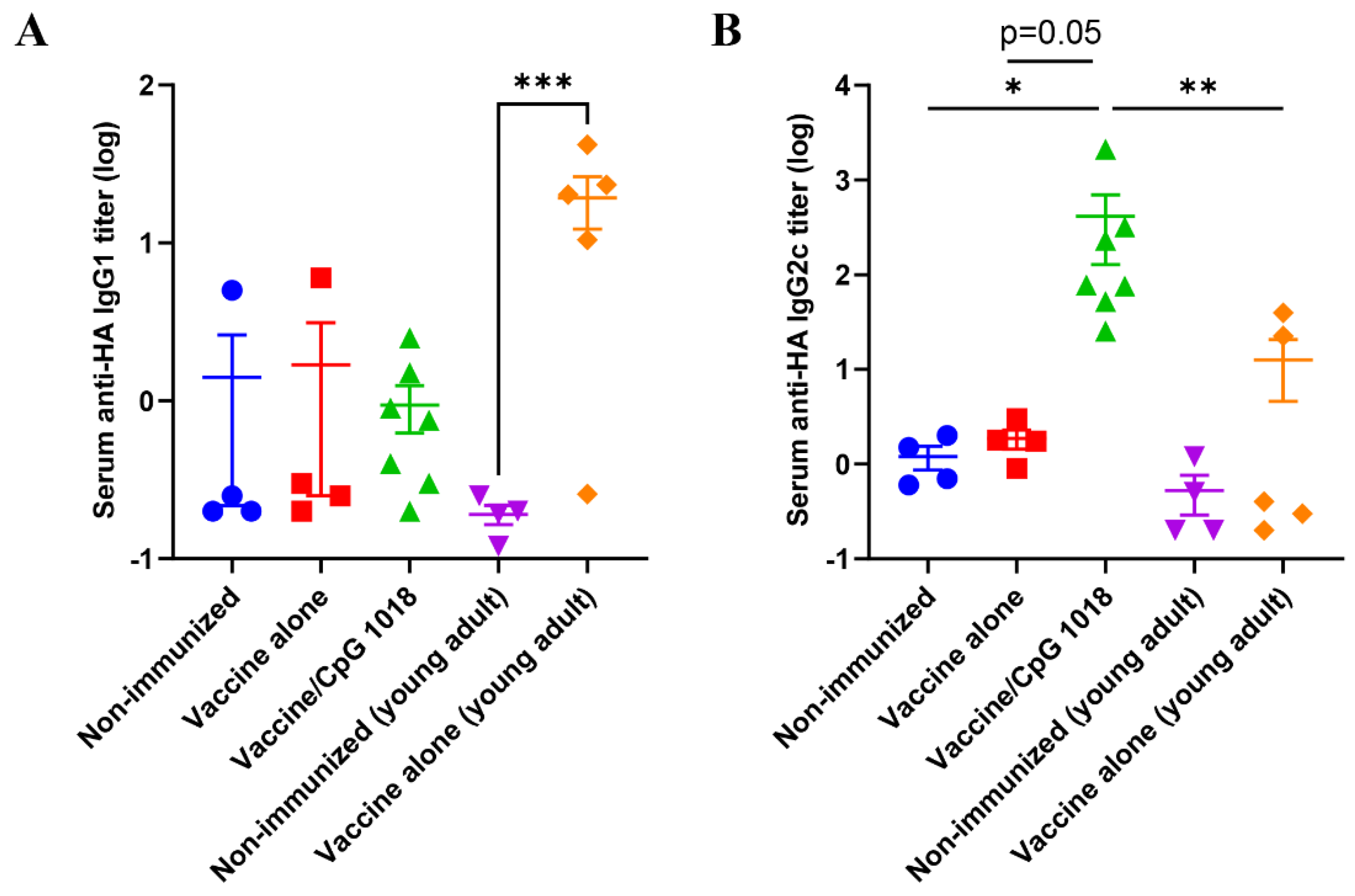
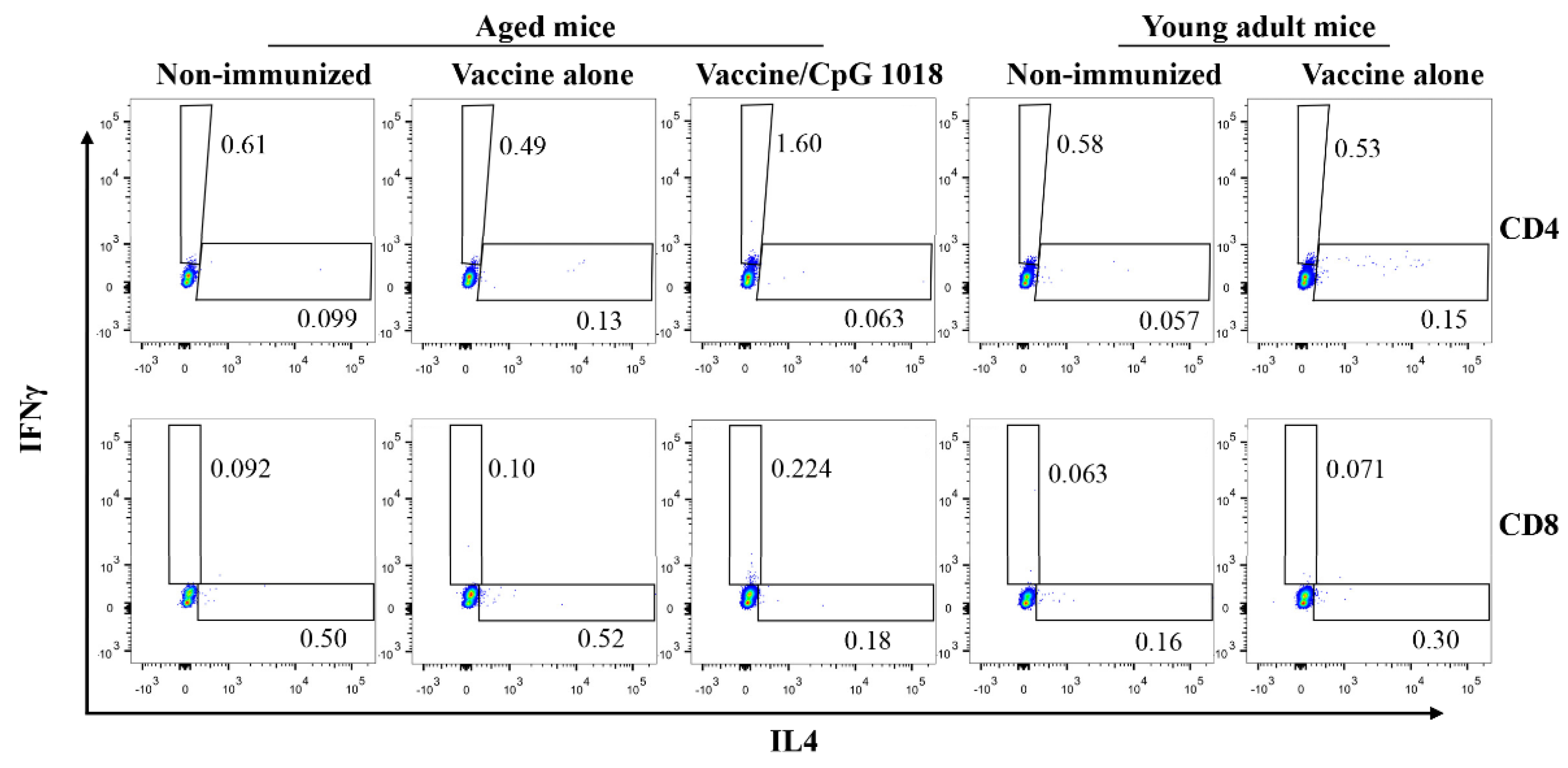
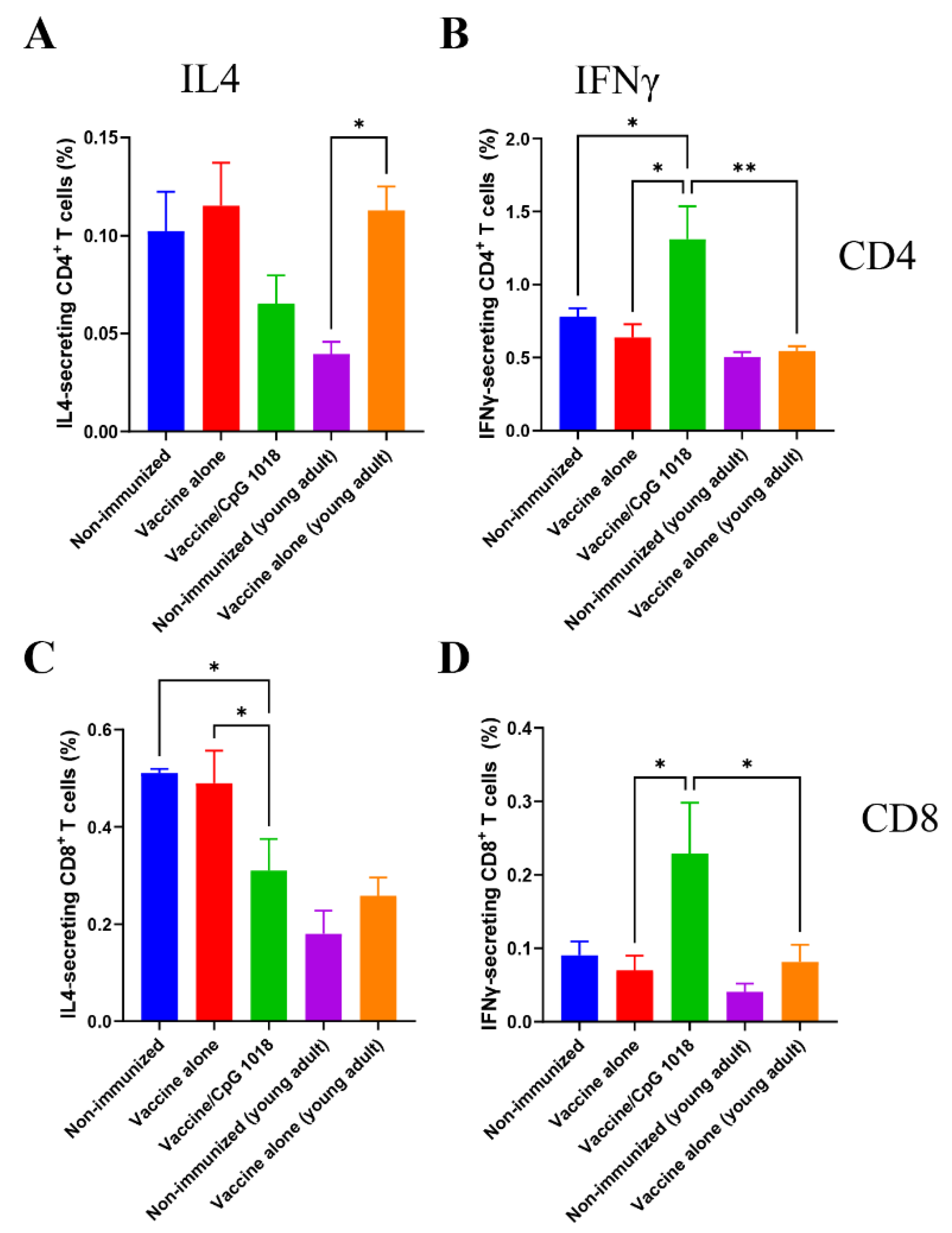
3.3. CpG 1018 Induces Th1 Cells and CTLs
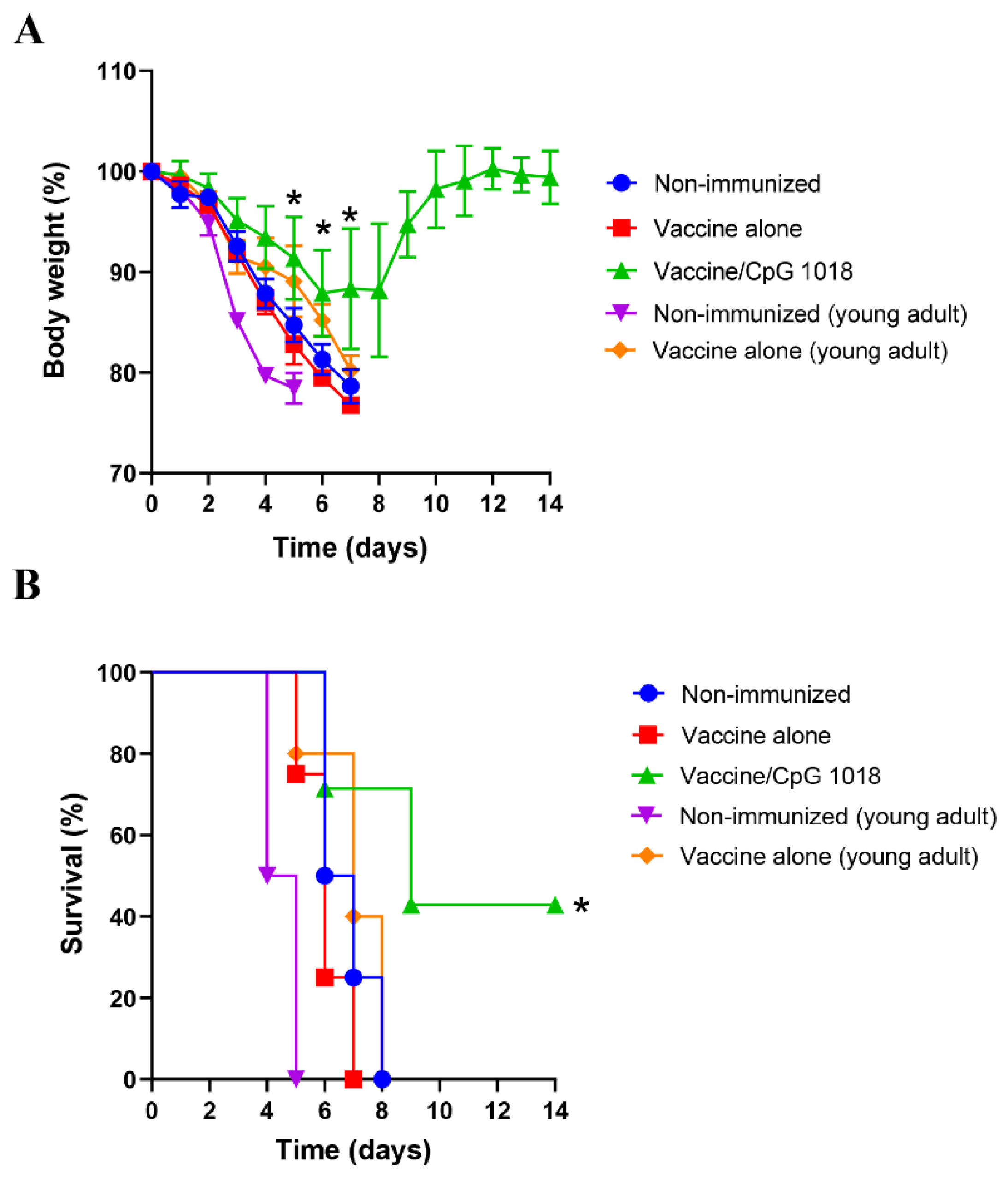
3.4. CpG 1018 Enhances Vaccine-Induced Protection
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Peteranderl, C.; Herold, S.; Schmoldt, C. Human Influenza Virus Infections. Semin. Respir. Crit. Care Med. 2016, 37, 487–500. [Google Scholar] [CrossRef] [PubMed]
- Petrova, V.N.; Russell, C.A. The evolution of seasonal influenza viruses. Nat. Rev. Microbiol. 2018, 16, 47–60. [Google Scholar] [CrossRef]
- McElhaney, J.E.; Verschoor, C.P.; Andrew, M.K.; Haynes, L.; Kuchel, G.A.; Pawelec, G. The immune response to influenza in older humans: Beyond immune senescence. Immun. Ageing 2020, 17, 10. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Vargas, E.A.; Wilk, E.; Canini, L.; Toapanta, F.R.; Binder, S.C.; Uvarovskii, A.; Ross, T.M.; Guzmán, C.A.; Perelson, A.S.; Meyer-Hermann, M. Effects of aging on influenza virus infection dynamics. J. Virol. 2014, 88, 4123–4131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palekar, R.S.; Rolfes, M.A.; Arriola, C.S.; Acosta, B.O.; Guidos, P.A.; Vargas, X.B.; Bancej, C.; Ramirez, J.B.; Baumeister, E.; Bruno, A.; et al. Burden of influenza-associated respiratory hospitalizations in the Americas, 2010–2015. PLoS ONE 2019, 14, e0221479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kulkarni, U.; Zemans, R.L.; Smith, C.A.; Wood, S.C.; Deng, J.C.; Goldstein, D.R. Excessive neutrophil levels in the lung underlie the age-associated increase in influenza mortality. Mucosal Immunol. 2019, 12, 545–554. [Google Scholar] [CrossRef] [Green Version]
- Mittelbrunn, M.; Kroemer, G. Hallmarks of T cell aging. Nat. Immunol. 2021, 22, 687–698. [Google Scholar] [CrossRef]
- Frasca, D.; Blomberg, B.B. Aging induces B cell defects and decreased antibody responses to influenza infection and vaccination. Immun. Ageing 2020, 17, 37. [Google Scholar] [CrossRef]
- Mertz, D.; Kim, T.H.; Johnstone, J.; Lam, P.-P.; Science, M.; Kuster, S.P.; Fadel, S.A.; Tran, D.; Fernandez, E.; Bhatnagar, N.; et al. Populations at risk for severe or complicated influenza illness: Systematic review and meta-analysis. BMJ 2013, 347, f5061. [Google Scholar] [CrossRef] [Green Version]
- Rondy, M.; El Omeiri, N.; Thompson, M.G.; Levêque, A.; Moren, A.; Sullivan, S.G. Effectiveness of influenza vaccines in preventing severe influenza illness among adults: A systematic review and meta-analysis of test-negative design case-control studies. J. Infect. 2017, 75, 381–394. [Google Scholar] [CrossRef]
- Machado, M.A.A.; Moura, C.S.; Abrahamowicz, M.; Ward, B.J.; Pilote, L.; Bernatsky, S. Relative effectiveness of influenza vaccines in elderly persons in the United States, 2012/2013–2017/2018 seasons. NPJ Vaccines 2021, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- Pelton, S.I.; Divino, V.; Shah, D.; Mould-Quevedo, J.; DeKoven, M.; Krishnarajah, G.; Postma, M.J. Evaluating the Relative Vaccine Effectiveness of Adjuvanted Trivalent Influenza Vaccine Compared to High-Dose Trivalent and Other Egg-Based Influenza Vaccines among Older Adults in the US during the 2017-2018 Influenza Season. Vaccines 2020, 8, 446. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Ott, G.S.; Nest, G.V.; Rappuoli, R.; Giudice, G.D. The history of MF59((R)) adjuvant: A phoenix that arose from the ashes. Expert Rev. Vaccines 2013, 12, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Thomas, P.G.; Keating, R.; Hulse-Post, D.J.; Doherty, P.C. Cell-mediated protection in influenza infection. Emerg. Infect. Dis. 2006, 12, 48–54. [Google Scholar] [CrossRef]
- Campbell, J.D. Development of the CpG Adjuvant 1018: A Case Study. Methods Mol. Biol. 2017, 1494, 15–27. [Google Scholar]
- Li, Z.; Zhao, Y.; Li, Y.; Chen, X. Adjuvantation of Influenza Vaccines to Induce Cross-Protective Immunity. Vaccines 2021, 9, 75. [Google Scholar] [CrossRef]
- Bode, C.; Zhao, G.; Steinhagen, F.; Kinjo, T.; Klinman, D.M. CpG DNA as a vaccine adjuvant. Expert Rev.Vaccines 2011, 10, 499–511. [Google Scholar] [CrossRef] [Green Version]
- Brazolot Millan, C.L.; Weeratna, R.; Krieg, A.M.; Sierist, C.-A.; Davis, H.L. CpG DNA can induce strong Th1 humoral and cell-mediated immune responses against hepatitis B surface antigen in young mice. Proc. Natl. Acad. Sci. USA 1998, 95, 15553–15558. [Google Scholar] [CrossRef] [Green Version]
- Sticchi, L.; Alberti, M.; Alicino, C.; Crovari, P. The intradermal vaccination: Past experiences and current perspectives. J. Prev. Med. Hyg. 2010, 51, 7–14. [Google Scholar]
- Li, Z.; Kim, K.H.; Bhatnagar, N.; Park, B.R.; Jeeva, S.; Jung, Y.-J.; Raha, J.; Kang, S.-M.; Chen, X. Physical radiofrequency adjuvant enhances immune responses to influenza H5N1 vaccination. FASEB J. 2022, 36, e22182. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, Z.; Voyer, J.; Li, Y.; Chen, X. Flagellin/Virus-like Particle Hybrid Platform with High Immunogenicity, Safety, and Versatility for Vaccine Development. ACS Appl. Mater. Interfaces 2022, 14, 21872–21885. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; Zhu, X.; Hossen, M.N.; Kakar, P.; Zhao, Y.; Chen, X. Augmentation of vaccine-induced humoral and cellular immunity by a physical radiofrequency adjuvant. Nat. Commun. 2018, 9, 3695. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Cao, Y.; Li, Y.; Zhao, Y.; Chen, X. Vaccine delivery alerts innate immune systems for more immunogenic vaccination. JCI Insight 2021, 6, e144627. [Google Scholar] [CrossRef]
- Ramirez, A.; Co, M.; Mathew, A. CpG Improves Influenza Vaccine Efficacy in Young Adult but Not Aged Mice. PLoS ONE 2016, 11, e0150425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.C.; Jarrahian, C.; Zehrung, D.; Mitragotri, S.; Prausnitz, M.R. Delivery systems for intradermal vaccination. Curr. Top. Microbiol. Immunol. 2012, 351, 77–112. [Google Scholar]
- Chen, X.; Kositratna, G.; Zhou, C.; Manstein, D.; Wu, M.X. Micro-fractional epidermal powder delivery for improved skin vaccination. J. Control. Release 2014, 192, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Arnou, R.; Icardi, G.; De Decker, M.; Ambrozaitis, A.; Kazek, M.-P.; Weber, F.; Van Damme, P. Intradermal influenza vaccine for older adults: A randomized controlled multicenter phase III study. Vaccine 2009, 27, 7304–7312. [Google Scholar] [CrossRef]
- Camilloni, B.; Basileo, M.; Valente, S.; Nunzi, E.; Iorio, A.M. Immunogenicity of intramuscular MF59-adjuvanted and intradermal administered influenza enhanced vaccines in subjects aged over 60: A literature review. Hum. Vaccines Immunother. 2015, 11, 553–563. [Google Scholar] [CrossRef] [Green Version]
- Nanishi, E.; Borriello, F.; O’Meara, T.R.; Mcgrath, M.E.; Saito, Y.; Haupt, R.E.; Seo, H.-S.; van Haren, S.D.; Cavazzoni, C.B.; Brook, B.; et al. An aluminum hydroxide:CpG adjuvant enhances protection elicited by a SARS-CoV-2 receptor binding domain vaccine in aged mice. Sci. Transl. Med. 2022, 14, eabj5305. [Google Scholar] [CrossRef]
- Brito, L.A.; Malyala, P.; O’Hagan, D.T. Vaccine adjuvant formulations: A pharmaceutical perspective. Semin. Immunol. 2013, 25, 130–145. [Google Scholar] [CrossRef]

 ) or in the presence of CpG 1018 adjuvant (40 µg,
) or in the presence of CpG 1018 adjuvant (40 µg,  ) or left non-immunized (
) or left non-immunized ( ). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine (
). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine ( ) or left non-immunized (
) or left non-immunized ( ). Serum HI titer was measured 3 weeks after immunization. n = 4–7. One-way ANOVA with Fisher’s LSD test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum HI titer was measured 3 weeks after immunization. n = 4–7. One-way ANOVA with Fisher’s LSD test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
). Serum HI titer was measured 3 weeks after immunization. n = 4–7. One-way ANOVA with Fisher’s LSD test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum HI titer was measured 3 weeks after immunization. n = 4–7. One-way ANOVA with Fisher’s LSD test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001. ) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti-HA IgG titer was measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti-HA IgG titer was measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti-HA IgG titer was measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti-HA IgG titer was measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05. ) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti−HA IgG1 (A) and IgG2c antibody titers (B) were measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti−HA IgG1 (A) and IgG2c antibody titers (B) were measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti−HA IgG1 (A) and IgG2c antibody titers (B) were measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
) or in the presence of CpG 1018 adjuvant (40 µg, ) or left non-immunized (). Young adult mice (6–8 weeks) were intradermally immunized with the same amount of pdm09 vaccine () or left non-immunized (). Serum anti−HA IgG1 (A) and IgG2c antibody titers (B) were measured 3 weeks after immunization by coating ELISA plate with rHA of pdm09 virus. n = 4–7. One-way ANOVA with Kruskal–Wallis multiple comparison test was used to compare differences between groups. *, p < 0.05; **, p < 0.01; ***, p < 0.001.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day 1 | Day 2 | Day 3 | Day 4 | Day 5 | |
|---|---|---|---|---|---|
| Significant level | NS | NS | NS | p < 0.001 | p < 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, X.; Li, Y.; Zhao, Y.; Chen, X. Overcoming Aging-Associated Poor Influenza Vaccine Responses with CpG 1018 Adjuvant. Vaccines 2022, 10, 1894. https://doi.org/10.3390/vaccines10111894
Kang X, Li Y, Zhao Y, Chen X. Overcoming Aging-Associated Poor Influenza Vaccine Responses with CpG 1018 Adjuvant. Vaccines. 2022; 10(11):1894. https://doi.org/10.3390/vaccines10111894
Chicago/Turabian StyleKang, Xinliang, Yibo Li, Yiwen Zhao, and Xinyuan Chen. 2022. "Overcoming Aging-Associated Poor Influenza Vaccine Responses with CpG 1018 Adjuvant" Vaccines 10, no. 11: 1894. https://doi.org/10.3390/vaccines10111894