

Nanoformulations with Leishmania braziliensis Antigens Triggered Controlled Parasite Burden in Vaccinated Golden Hamster (Mesocricetus auratus) against Visceral Leishmaniasis
 , ,
, ,  ,
,  ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Studied Animals and Experimental Groups
2.3. Production of Vaccine Formulations
2.4. Experimental Protocol
2.5. Assessment of Total Anti-Leishmania IgG Antibodies
2.6. Histology Analysis
2.7. Parasite Burden Evaluation
2.7.1. Limiting Dilution Assay
2.7.2. Quantitative PCR (qPCR)
2.8. Statistical Analyses
3. Results
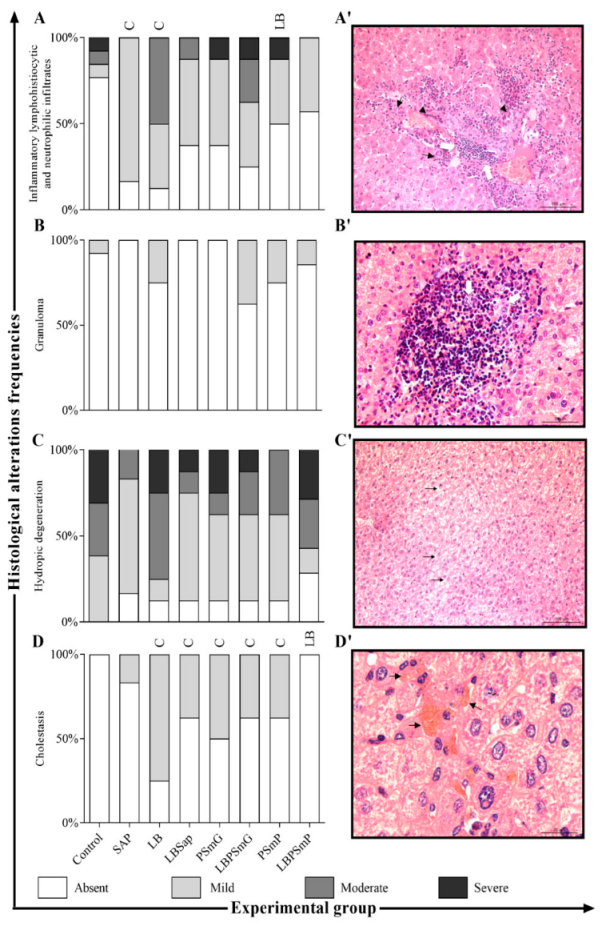
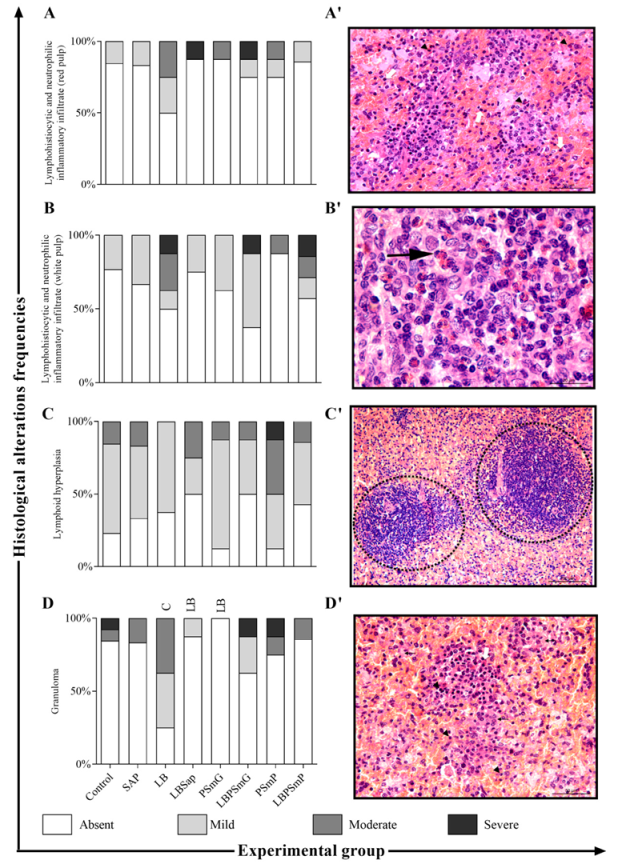
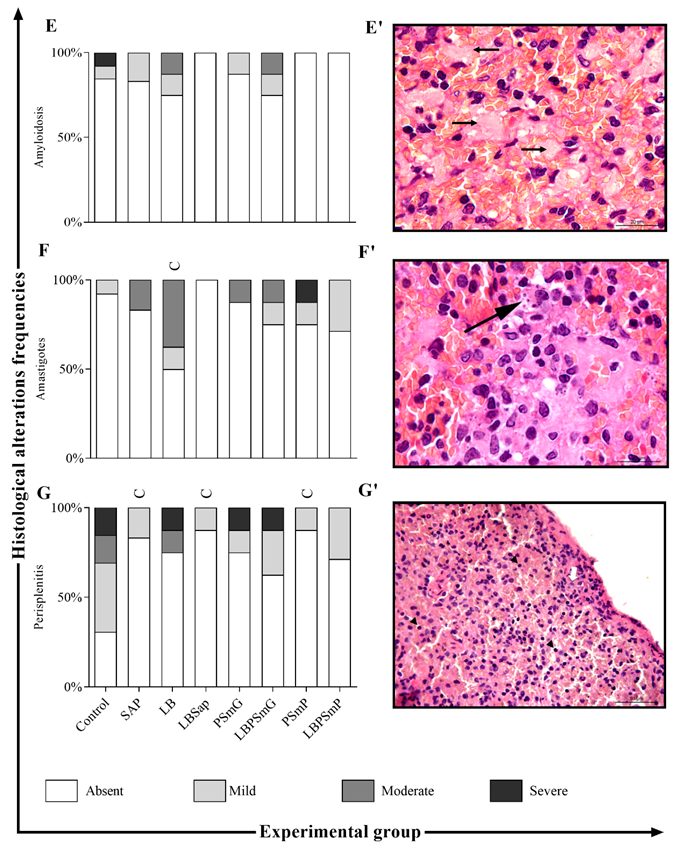
3.1. Immunobiological Safety and Harmless Analysis
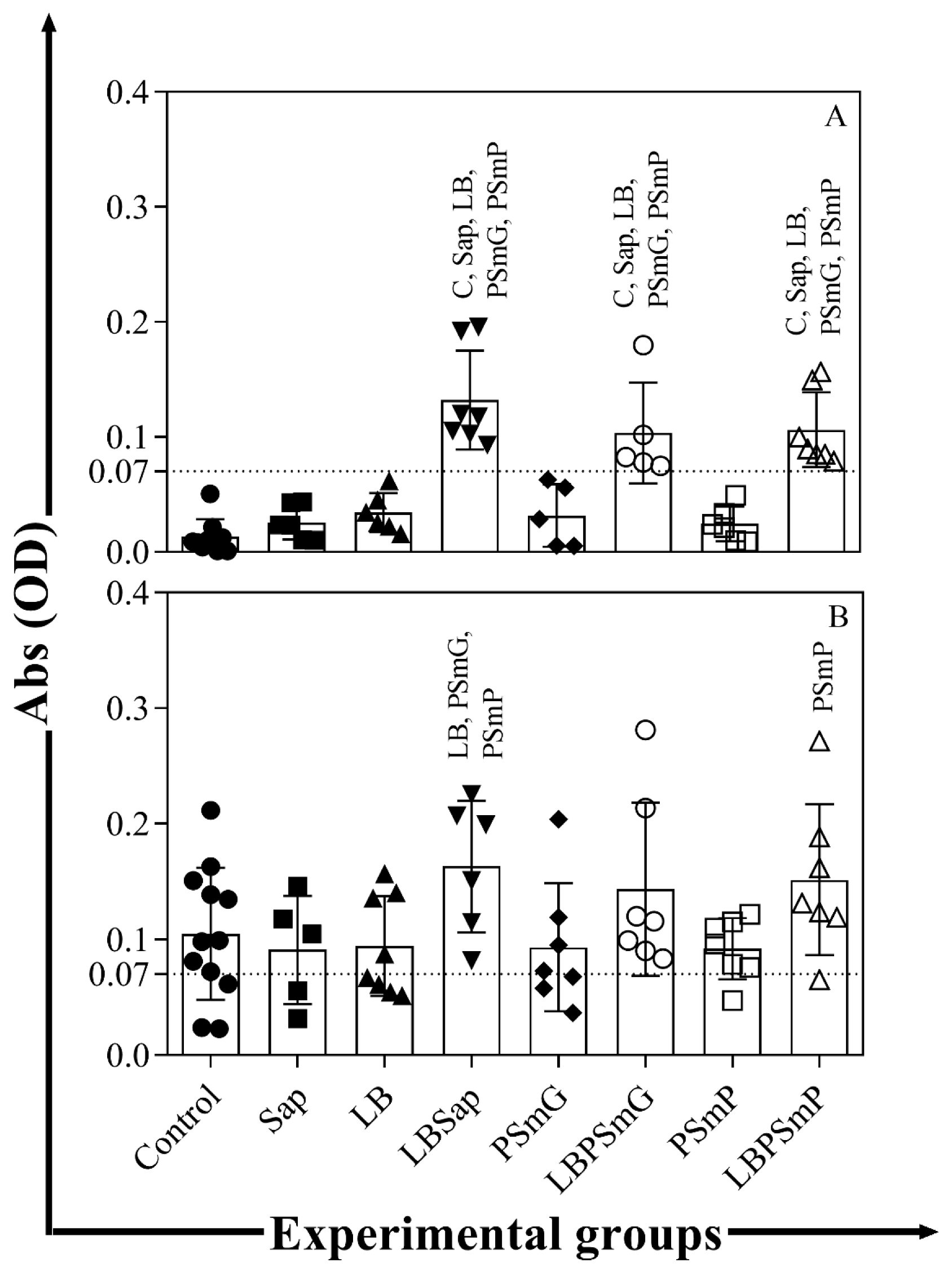
3.2. Immunogenic Potential of Nanoformulations
3.3. Evaluation of Changes in Parasite Target Tissues
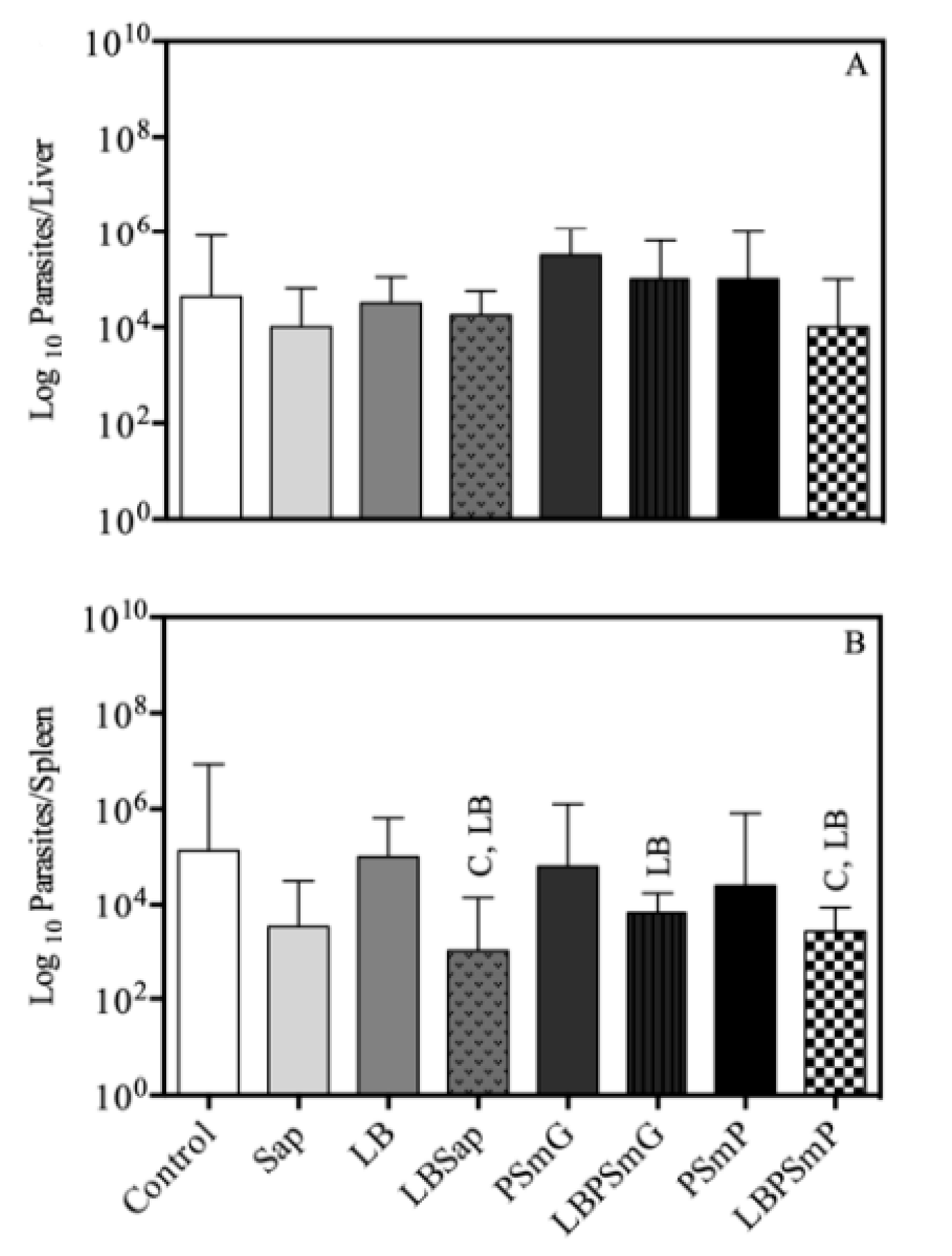
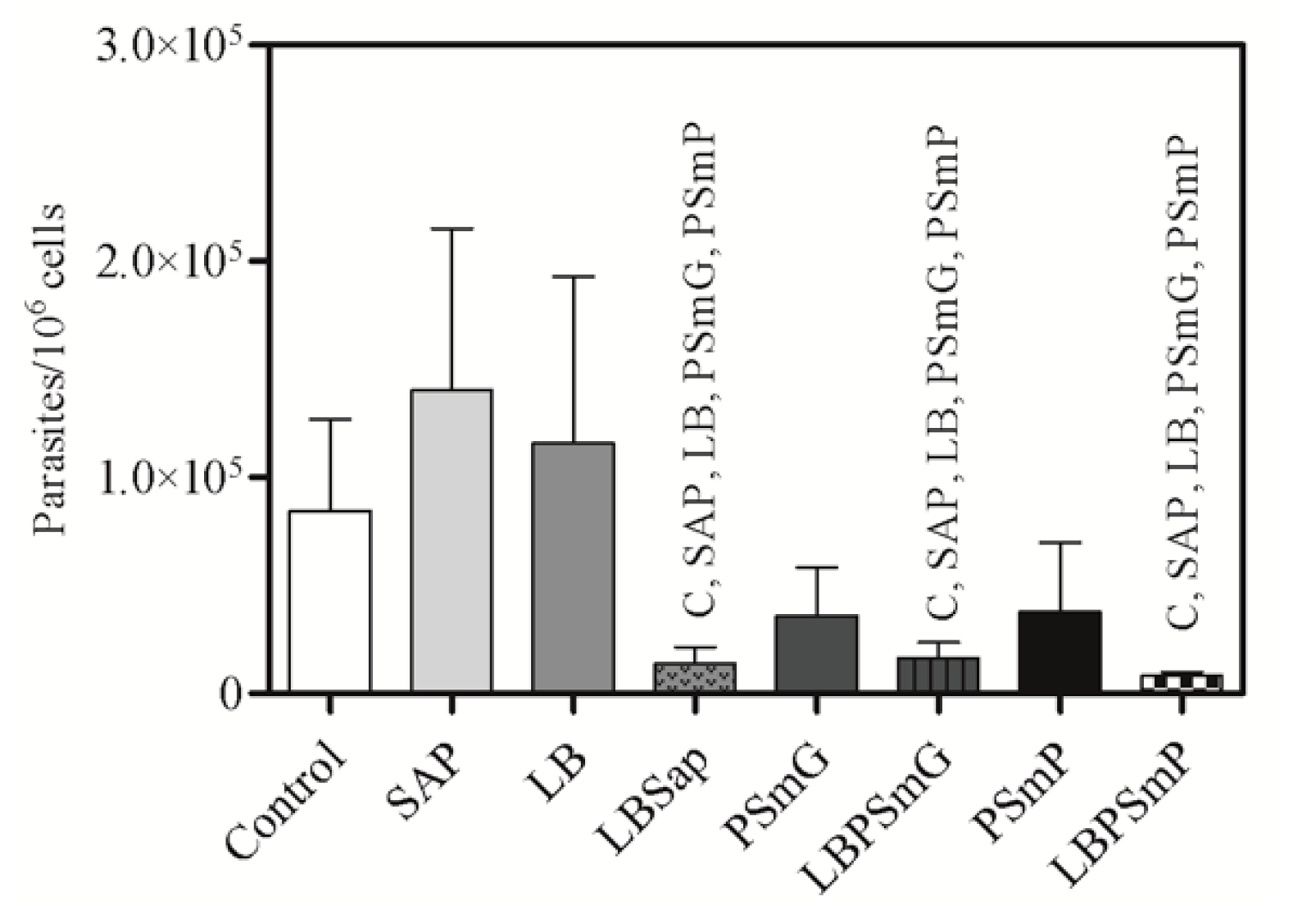
3.4. Efficacy Evaluation Based on Parasite Load
3.4.1. Limiting Dilution Assay
3.4.2. Quantitative Molecular Analysis (qPCR)
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Gutiérrez, V.; Seabra, A.B.; Reguera, R.M.; Khandare, J.; Calderón, M. New Approaches from Nanomedicine for Treating Leishmaniasis. Chem. Soc. Rev. 2015, 45, 152–168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ministério da Saúde. Guia de Vigilância em Saúde-Volume 3; Ministério da Saúde: Araraquara, Brazil, 2017; Volume 3, ISBN 9788533421790. [Google Scholar]
- Hernández, D.; Martínez, C.; Garcia, L.; Rodríguez, N.; Convit, J. Leishmania braziliensis Causing Visceral Leishmaniasis In A Patient With Human Immunodeficiency Virus Infection, Identified With The Aid Of The Polymerase Chain Reaction. Trans. R. Soc. Trop. Med. Hyg. 1993, 87, 627–628. [Google Scholar] [CrossRef]
- Kaye, P.M.; Beattie, L. Lessons from Other Diseases: Granulomatous Inflammation in Leishmaniasis. Semin. Immunopathol. 2016, 38, 249–260. [Google Scholar] [CrossRef] [Green Version]
- Tolezano, J.E.; Uliana, S.R.B.; Taniguchi, H.H.; Araújo, M.F.L.; Barbosa, J.A.R.; Barbosa, J.E.R.; Floeter-Winter, L.M.; Shaw, J.J. The First Records of Leishmania (Leishmania) amazonensis in Dogs (Canis familiaris) Diagnosed Clinically as Having Canine Visceral Leishmaniasis from Araçatuba County, São Paulo State, Brazil. Vet. Parasitol. 2007, 149, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Dantas-Torres, F. The Role of Dogs as Reservoirs of Leishmania Parasites, with Emphasis on Leishmania (Leishmania) infantum and Leishmania (Viannia) braziliensis. Vet. Parasitol. 2007, 149, 139–146. [Google Scholar] [CrossRef]
- Valdivia, H.O.; Almeida, L.V.; Roatt, B.M.; Reis-Cunha, J.L.; Pereira, A.A.S.; Gontijo, C.; Fujiwara, R.T.; Reis, A.B.; Sanders, M.J.; Cotton, J.A.; et al. Comparative Genomics of Canine-Isolated Leishmania (Leishmania) amazonensis from an Endemic Focus of Visceral Leishmaniasis in Governador Valadares, Southeastern Brazil. Sci. Rep. 2017, 7, 40804. [Google Scholar] [CrossRef] [Green Version]
- Estevam, L.G.T.M.; Veloso, L.B.; Silva, G.G.; Mori, C.C.; Franco, P.F.; Lima, A.C.V.M.R.; Ássimos, G.R.; Reis, I.A.; Andrade-Filho, J.D.; Araújo, M.S.S.; et al. Leishmania infantum Infection Rate in Dogs Housed in Open-Admission Shelters Is Higher than of Domiciled Dogs in an Endemic Area of Canine Visceral Leishmaniasis. Epidemiological Implications. Acta Trop. 2022, 232, 106492. [Google Scholar] [CrossRef]
- Marzochi, M.C.; Coutinho, S.G.; de Souza, W.J.; Toledo, L.M.D.; Grimaldi, G., Jr.; Momen, H.; Pacheco, R.d.S.; Sabroza, P.C.; de Souza, M.A.; Rangel, F.B., Jr.; et al. Canine Visceral Leishmaniasis in Rio de Janeiro, Brazil. Clinical, Parasitological, Therapeutical and Epidemiological Findings (1977–1983). Memórias do Inst. Oswaldo Cruz 1985, 80, 349–357. [Google Scholar] [CrossRef] [Green Version]
- Ministério da Saúde. Manual de Vigilância e Controle da Leishmaniose Visceral; Ministério da Saúde: Araraquara, Brazil, 2014; ISBN 9788533407428. [Google Scholar]
- Getts, D.R.; Shea, L.D.; Miller, S.D.; King, N.J.C. Harnessing Nanoparticles for Immune Modulation. Trends Immunol. 2015, 36, 419–427. [Google Scholar] [CrossRef] [Green Version]
- Doroud, D.; Rafati, S. Leishmaniasis: Focus on the Design of Nanoparticulate Vaccine Delivery Systems. Expert Rev. Vaccines 2012, 11, 69–86. [Google Scholar] [CrossRef]
- Zadeh Mehrizi, T.; Khamesipour, A.; Shafiee Ardestani, M.; Ebrahimi Shahmabadi, H.; Haji Molla Hoseini, M.; Mosaffa, N.; Ramezani, A. Comparative Analysis between Four Model Nanoformulations of Amphotericin B-Chitosan, Amphotericin B-Dendrimer, Betulinic Acid-Chitosan and Betulinic Acid-Dendrimer for Treatment of Leishmania Major: Real-Time PCR Assay Plus. Int. J. Nanomed. 2019, 14, 7593–7607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tafaghodi, M.; Khamesipour, A.; Jaafari, M.R. Immunization against Leishmaniasis by PLGA Nanospheres Encapsulated with Autoclaved Leishmania major (ALM) and CpG-ODN. Parasitol. Res. 2011, 108, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Melby, P.C.; Chandrasekar, B.; Zhao, W.; Coe, J.E. The Hamster as a Model of Human Visceral Leishmaniasis: Progressive Disease and Impaired Generation of Nitric Oxide in the Face of a Prominent Th1-like Cytokine Response. J. Immunol. 2001, 166, 1912–1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiménez-Antón, M.D.; Grau, M.; Olías-Molero, A.I.; Alunda, J.M. Syrian Hamster as an Advanced Experimental Model for Visceral Leishmaniasis; Humana Press: New York, NY, USA, 2019; pp. 303–314. [Google Scholar]
- Giunchetti, R.C.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Roatt, B.M.; de Oliveira Aguiar-Soares, R.D.; de Souza, J.V.; das Dores Moreira, N.; Malaquias, L.C.C.; Mota e Castro, L.L.; et al. Immunogenicity of a Killed Leishmania Vaccine with Saponin Adjuvant in Dogs. Vaccine 2007, 25, 7674–7686. [Google Scholar] [CrossRef]
- Vitoriano-Souza, J.; Reis, A.B.; Moreira, N.D.; Giunchetti, R.C.; Correa-Oliveira, R.; Carneiro, C.M. Kinetics of Cell Migration to the Dermis and Hypodermis in Dogs Vaccinated with Antigenic Compounds of Leishmania braziliensis plus Saponin. Vaccine 2008, 26, 3922–3931. [Google Scholar] [CrossRef]
- Moreira, N.d.D.; Giunchetti, R.C.; Carneiro, C.M.; Vitoriano-Souza, J.; Roatt, B.M.; Malaquias, L.C.C.; Corrêa-Oliveira, R.; Reis, A.B. Histological Study of Cell Migration in the Dermis of Hamsters after Immunisation with Two Different Vaccines against Visceral Leishmaniasis. Vet. Immunol. Immunopathol. 2009, 128, 418–424. [Google Scholar] [CrossRef] [Green Version]
- Fessi, H.; Puisieux, F.; Devissaguet, J.P.; Ammoury, N.; Benita, S. Nanocapsule Formation By Interfacial Polymer Deposition Following Solvent Displacement. Int. J. Pharm. 1989, 55, R1–R4. [Google Scholar] [CrossRef]
- Garcia, G.M.; Oliveira, L.T.; Pitta, I.D.R.; De Lima, M.D.C.A.; Vilela, J.M.C.; Andrade, M.S.; Abdalla, D.S.P.; Mosqueira, V.C.F. Improved Nonclinical Pharmacokinetics and Biodistribution of a New PPAR Pan-Agonist and COX Inhibitor in Nanocapsule Formulation. J. Control. Release 2015, 209, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Moreira, D.; Vitoriano-souza, J.; Roatt, B.M.; Melo, P.; Vieira, D.A.; Ker, H.G.; Mirelle, J.; Cardoso, D.O.; Giunchetti, R.C.; Carneiro, M.; et al. Parasite Burden in Hamsters Infected with Two Different Strains of Leishmania (Leishmania) infantum: “Leishman Donovan Units ” versus Real-Time PCR. PLoS ONE 2012, 7, e47907. [Google Scholar] [CrossRef] [Green Version]
- Marques-Da-Silva, E.A.; Coelho, E.A.F.; Gomes, D.C.O.; Vilela, M.C.; Masioli, C.Z.; Tavares, C.A.P.; Fernandes, A.P.; Afonso, L.C.C.; Rezende, S.A. Intramuscular Immunization with P36(LACK) DNA Vaccine Induces IFN-γ Production but Does Not Protect BALB/c Mice against Leishmania chagasi Intravenous Challenge. Parasitol. Res. 2005, 98, 67–74. [Google Scholar] [CrossRef]
- Titus, R.G.; Marchand, M.; Boon, T.; Louis, J.A. A Limiting Dilution Assay for Quantifying Leishmania Major in Tissues of Infected Mice. Parasite Immunol. 1985, 7, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, M.S.; Bento, G.A.; de Almeida, L.V.; de Castro, J.C.; Reis-Cunha, J.L.; Barbosa, V.d.A.; de Souza, C.F.; Brazil, R.P.; Valdivia, H.O.; Bartholomeu, D.C. Detection of Multiple Circulating Leishmania Species in Lutzomyia longipalpis in the City of Governador Valadares, Southeastern Brazil. PLoS ONE 2019, 14, e0211831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vakili, B.; Nezafat, N.; Negahdaripour, M.; Ghasemi, Y. A Structural Vaccinology Approach for in Silico Designing of a Potential Self-Assembled Nanovaccine against Leishmania infantum. Exp. Parasitol. 2022, 239, 108295. [Google Scholar] [CrossRef] [PubMed]
- Nafari, A.; Cheraghipour, K.; Sepahvand, M.; Shahrokhi, G.; Gabal, E.; Mahmoudvand, H. Nanoparticles: New Agents toward Treatment of Leishmaniasis. Parasite Epidemiol. Control 2020, 10, e00156. [Google Scholar] [CrossRef] [PubMed]
- Sekhon, B.S.; Saluja, V. Nanovaccines-an Overview. Int. J. Pharm. Front. Res. 2011, 1, 101–109. [Google Scholar]
- Garg, R.; Dube, A. Animal Models for Vaccine Studies for Visceral Leishmaniasis. Indian J. Med. Res. 2006, 123, 439–454. [Google Scholar]
- Hochman, B.; Ferreira, L.M.; Bôas, F.C.V.; Mariano, M. Experimental Model in Hamster (Mesocricetus auratus) to Study Heterologous Graft of Scars and Cutaneous Diseases in Plastic Surgery. Acta Cir. Bras. 2004, 19, 69–78. [Google Scholar] [CrossRef] [Green Version]
- Nieto, A.; Domínguez-Bernal, G.; Orden, J.A.; De La Fuente, R.; Madrid-Elena, N.; Carrión, J. Mechanisms of Resistance and Susceptibility to Experimental Visceral Leishmaniosis: BALB/c Mouse versus Syrian Hamster Model. Vet. Res. 2011, 42, 39. [Google Scholar] [CrossRef] [Green Version]
- Giunchetti, R.C.; Corrêa-Oliveira, R.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Roatt, B.M.; Aguiar-Soares, R.D.d.O.; Coura-Vital, W.; Abreu, R.T.d.; Malaquias, L.C.C.; Gontijo, N.F.; et al. A Killed Leishmania Vaccine with Sand Fly Saliva Extract and Saponin Adjuvant Displays Immunogenicity in Dogs. Vaccine 2008, 26, 623–638. [Google Scholar] [CrossRef] [Green Version]
- Giunchetti, R.C.; Reis, A.B.; da Silveira-Lemos, D.; Martins-Filho, O.A.; Corrêa-Oliveira, R.; Bethony, J.; Vale, A.M.; da Silva Quetz, J.; Bueno, L.L.; França-Silva, J.C.; et al. Antigenicity of a Whole Parasite Vaccine as Promising Candidate against Canine Leishmaniasis. Res. Vet. Sci. 2008, 85, 106–112. [Google Scholar] [CrossRef]
- de Mendonça, L.Z.; Resende, L.A.; Lanna, M.F.; Aguiar-Soares, R.D.d.O.; Roatt, B.M.; Castro, R.A.d.O.E.; Batista, M.A.; Silveira-Lemos, D.; Gomes, J.d.A.S.; Fujiwara, R.T.; et al. Multicomponent LBSap Vaccine Displays Immunological and Parasitological Profiles Similar to Those of Leish-Tec® and Leishmune® Vaccines against Visceral Leishmaniasis. Parasites Vectors 2016, 9, 472. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roatt, B.M.; Aguiar-soares, R.D.D.O.; Vitoriano-souza, J.; Coura, W.; Corre, R.; Martins-filho, O.A.; Leo, S.; Lana, M.D.; Gontijo, N.F.; Jose, M.; et al. Performance of LBSap Vaccine after Intradermal Challenge with L. infantum and Saliva of Lu. longipalpis: Immunogenicity and Parasitological Evaluation. PLoS ONE 2012, 7, e49780. [Google Scholar] [CrossRef] [PubMed]
- Aguiar-Soares, R.D.d.O.; Roatt, B.M.; Mathias, F.A.S.; Reis, L.E.S.; Cardoso, J.M.d.O.; Brito, R.C.F.d.; Ker, H.G.; Corrêa-Oliveira, R.; Giunchetti, R.C.; Reis, A.B. Phase I and II Clinical Trial Comparing the LBSap, Leishmune®, and Leish-Tec® Vaccines against Canine Visceral Leishmaniasis. Vaccines 2020, 8, 690. [Google Scholar] [CrossRef] [PubMed]
- Reis, A.B.; Martins-Filho, O.A.; Teixeira-Carvalho, A.; Giunchetti, R.C.; Carneiro, C.M.; Mayrink, W.; Tafuri, W.L.; Corrêa-Oliveira, R. Systemic and Compartmentalized Immune Response in Canine Visceral Leishmaniasis. Vet. Immunol. Immunopathol. 2009, 128, 87–95. [Google Scholar] [CrossRef] [Green Version]
- Moreira, N.d.D.; Vitoriano-Souza, J.; Roatt, B.M.; Vieira, P.M.d.A.; Coura-Vital, W.; Cardoso, J.M.d.O.; Rezende, M.T.; Ker, H.G.; Giunchetti, R.C.; Carneiro, C.M.; et al. Clinical, Hematological and Biochemical Alterations in Hamster (Mesocricetus auratus) Experimentally Infected with Leishmania infantum through Different Routes of Inoculation. Parasites Vectors 2016, 9, 181. [Google Scholar] [CrossRef] [Green Version]
- Costa-Pereira, C.; Moreira, M.L.; Soares, R.P.; Marteleto, B.H.; Ribeiro, V.M.; França-Dias, M.H.; Cardoso, L.M.; Viana, K.F.; Giunchetti, R.C.; Martins-Filho, O.A.; et al. One-Year Timeline Kinetics of Cytokine-Mediated Cellular Immunity in Dogs Vaccinated against Visceral Leishmaniasis. BMC Vet. Res. 2015, 11, 92. [Google Scholar] [CrossRef] [Green Version]
- Hojatizade, M.; Soleymani, M.; Tafaghodi, M.; Badiee, A.; Chavoshian, O.; Jaafari, M.R. Chitosan Nanoparticles Loaded with Whole and Soluble Leishmania Antigens, and Evaluation of Their Immunogenecity in a Mouse Model of Leishmaniasis. Iran. J. Immunol. IJI 2018, 15, 281–293. [Google Scholar] [CrossRef]
- Kolios, G.; Valatas, V.; Kouroumalis, E. Role of Kupffer Cells in the Pathogenesis of Liver Disease. World J. Gastroenterol. WJG 2006, 12, 7413–7420. [Google Scholar] [CrossRef]
- Melo, F.A.; Moura, E.P.; Ribeiro, R.R.; Alves, C.F.; Caliari, M.V.; Tafuri, W.L.; Calabrese, K.D.S.; Tafuri, W.L. Hepatic Extracellular Matrix Alterations in Dogs Naturally Infected with Leishmania (Leishmania) chagasi. Int. J. Exp. Pathol. 2009, 90, 538–548. [Google Scholar] [CrossRef]
- Vianna, V.; Takiya, C.; De Brito-Gitirana, L. Histopathologic Analysis of Hamster Hepatocytes Submitted to Experimental Infection with Leishmania donovani. Parasitol. Res. 2002, 88, 829–836. [Google Scholar] [CrossRef]
- Requena, J.M.; Soto, M.; Doria, M.D.; Alonso, C. Immune and Clinical Parameters Associated with Leishmania infantum Infection in the Golden Hamster Model. Vet. Immunol. Immunopathol. 2000, 76, 269–281. [Google Scholar] [CrossRef]
- Fiuza, J.A.; Dey, R.; Davenport, D.; Abdeladhim, M.; Meneses, C.; Oliveira, F.; Kamhawi, S.; Valenzuela, J.G.; Gannavaram, S.; Nakhasi, H.L. Intradermal Immunization of Leishmania donovani Centrin Knock-Out Parasites in Combination with Salivary Protein LJM19 from Sand Fly Vector Induces a Durable Protective Immune Response in Hamsters. PLoS Negl. Trop. Dis. 2016, 10, e0004322. [Google Scholar] [CrossRef] [PubMed]
- Riça-Capela, M.J.; Cortes, S.; Leandro, C.; Peleteiro, M.C.; Santos-Gomes, G.; Campino, L. Immunological and Histopathological Studies in a Rodent Model Infected with Leishmania infantum Promastigotes or Amastigotes. Parasitol. Res. 2003, 89, 163–169. [Google Scholar] [CrossRef] [PubMed]
- d’El-R Hermida, M.; de Melo, C.V.B.; Lima, I.d.S.; Oliveira, G.G.d.S.; Dos-Santos, W.L.C. Histological Disorganization of Spleen Compartments and Severe Visceral Leishmaniasis. Front. Cell. Infect. Microbiol. 2018, 8, 394. [Google Scholar] [CrossRef] [Green Version]
- Stanley, A.C.; Engwerda, C.R. Balancing Immunity and Pathology in Visceral Leishmaniasis. Immunol. Cell Biol. 2007, 85, 138–147. [Google Scholar] [CrossRef]
- Smelt, S.C.; Engwerda, C.R.; McCrossen, M.; Kaye, P.M. Destruction of Follicular Dendritic Cells during Chronic Visceral Leishmaniasis. J. Immunol. 1997, 158, 3813–3821. [Google Scholar]
- dos-Santos, W.L.; Pagliari, C.; Santos, L.G.; Almeida, V.A.; e Silva, T.L.; Coutinho, J.d.J.; Souza, T.; Duarte, M.I.; de Freitas, L.A.; Costa, C.H. A Case of Conventional Treatment Failure in Visceral Leishmaniasis: Leukocyte Distribution and Cytokine Expression in Splenic Compartments. BMC Infect. Dis. 2014, 14, 491. [Google Scholar] [CrossRef]
- Lima, V.M.F.d.; Fattori, K.R.; de Souza, F.; Eugênio, F.R.; Santos, P.S.P.d.; Rozza, D.B.; Machado, G.F. Apoptosis in T Lymphocytes from Spleen Tissue and Peripheral Blood of L. (L.) chagasi Naturally Infected Dogs. Vet. Parasitol. 2012, 184, 147–153. [Google Scholar] [CrossRef] [Green Version]
- Roatt, B.M.; Aguiar-Soares, R.D.d.O.; Reis, L.E.S.; Cardoso, J.M.d.O.; Mathias, F.A.S.; Brito, R.C.F.d.; da Silva, S.M.; Gontijo, N.D.F.; Ferreira, S.d.A.; Valenzuela, J.G.; et al. A Vaccine Therapy for Canine Visceral Leishmaniasis Promoted Significant Improvement of Clinical and Immune Status with Reduction in Parasite Burden. Front. Immunol. 2017, 8, 217. [Google Scholar] [CrossRef] [Green Version]
- Lima, W.G.; Oliveira, P.S.; Caliari, M.V.; Gonçalves, R.; Michalick, M.S.M.; Melo, M.N.; Tafuri, W.L.; Tafuri, W.L. Histopathological and Immunohistochemical Study of Type 3 Complement Receptors (CD11b/CD18) in Livers and Spleens of Asymptomatic and Symptomatic Dogs Naturally Infected with Leishmania (Leishmania) chagasi. Vet. Immunol. Immunopathol. 2007, 117, 129–136. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ottino, J.; Leite, J.C.; Melo-Júnior, O.A.; González, M.A.C.; de Carvalho, T.F.; Garcia, G.M.; Batista, M.A.; Silveira, P.; Cardoso, M.S.; Bueno, L.L.; et al. Nanoformulations with Leishmania braziliensis Antigens Triggered Controlled Parasite Burden in Vaccinated Golden Hamster (Mesocricetus auratus) against Visceral Leishmaniasis. Vaccines 2022, 10, 1848. https://doi.org/10.3390/vaccines10111848
Ottino J, Leite JC, Melo-Júnior OA, González MAC, de Carvalho TF, Garcia GM, Batista MA, Silveira P, Cardoso MS, Bueno LL, et al. Nanoformulations with Leishmania braziliensis Antigens Triggered Controlled Parasite Burden in Vaccinated Golden Hamster (Mesocricetus auratus) against Visceral Leishmaniasis. Vaccines. 2022; 10(11):1848. https://doi.org/10.3390/vaccines10111848
Chicago/Turabian StyleOttino, Jennifer, Jaqueline Costa Leite, Otoni Alves Melo-Júnior, Marco Antonio Cabrera González, Tatiane Furtado de Carvalho, Giani Martins Garcia, Maurício Azevedo Batista, Patrícia Silveira, Mariana Santos Cardoso, Lilian Lacerda Bueno, and et al. 2022. "Nanoformulations with Leishmania braziliensis Antigens Triggered Controlled Parasite Burden in Vaccinated Golden Hamster (Mesocricetus auratus) against Visceral Leishmaniasis" Vaccines 10, no. 11: 1848. https://doi.org/10.3390/vaccines10111848