

Effector-Memory B-Lymphocytes and Follicular Helper T-Lymphocytes as Central Players in the Immune Response in Vaccinated and Nonvaccinated Populations against SARS-CoV-2

 ,
,  ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
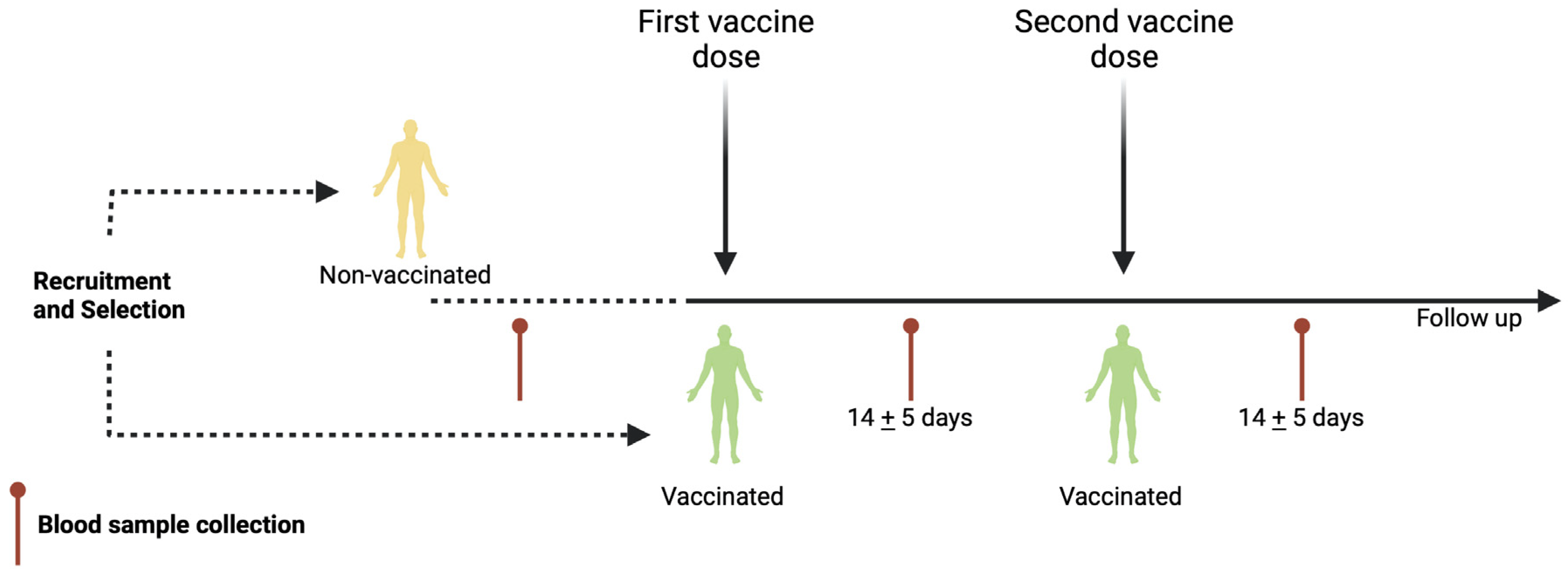
2.1. Population Studied
2.2. Blood Sample Collection
2.3. Detection of IgGs Related to SARS-CoV-2 by Enzyme-Linked Immunosorbent Assay (ELISA)
2.4. Total Count of Neutrophils, Lymphocytes, and Neutrophil–Lymphocyte Ratio (NLR)
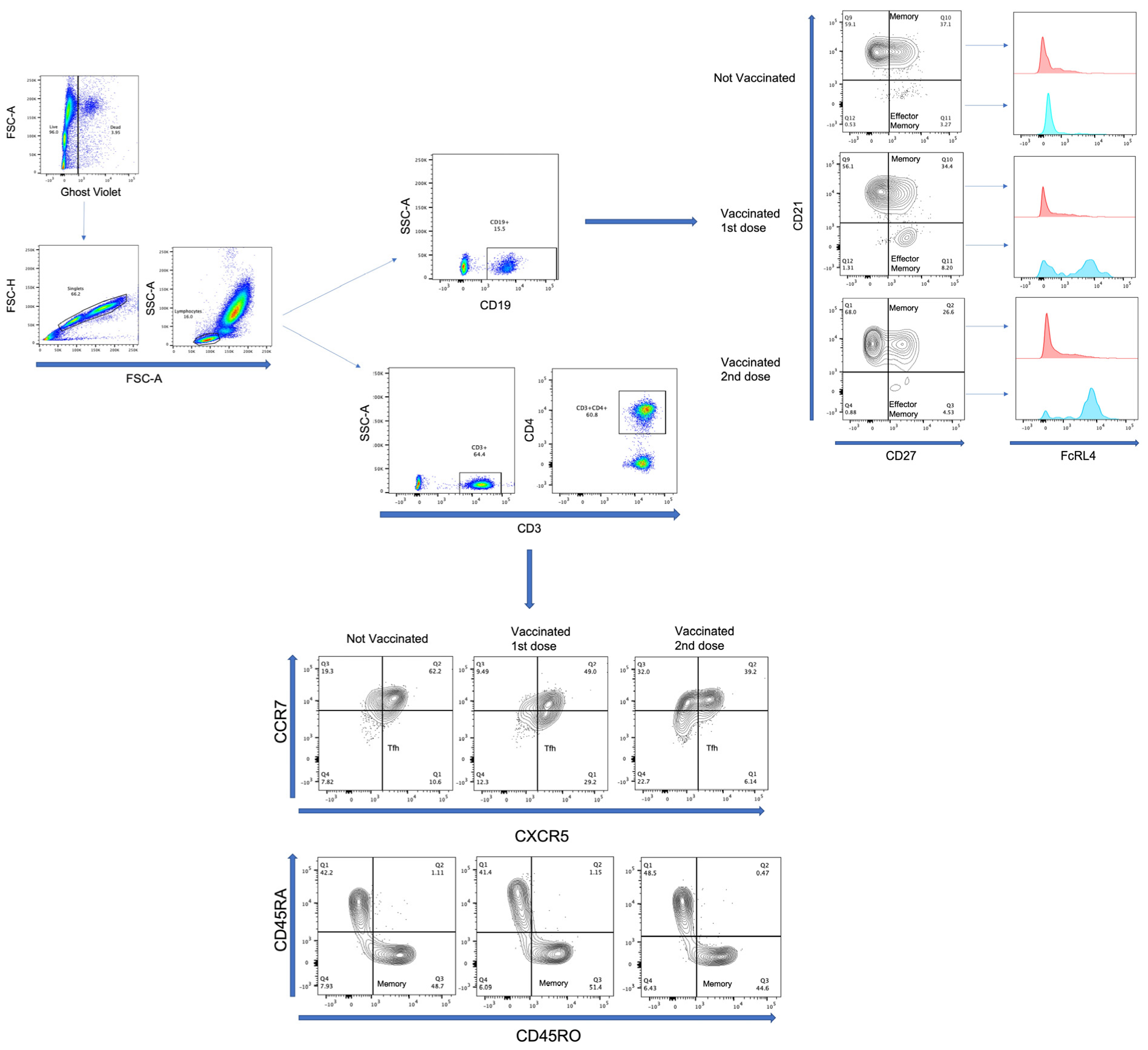
2.5. Multiparametric Flow Cytometry Analysis
2.6. Statistical Analysis
3. Results
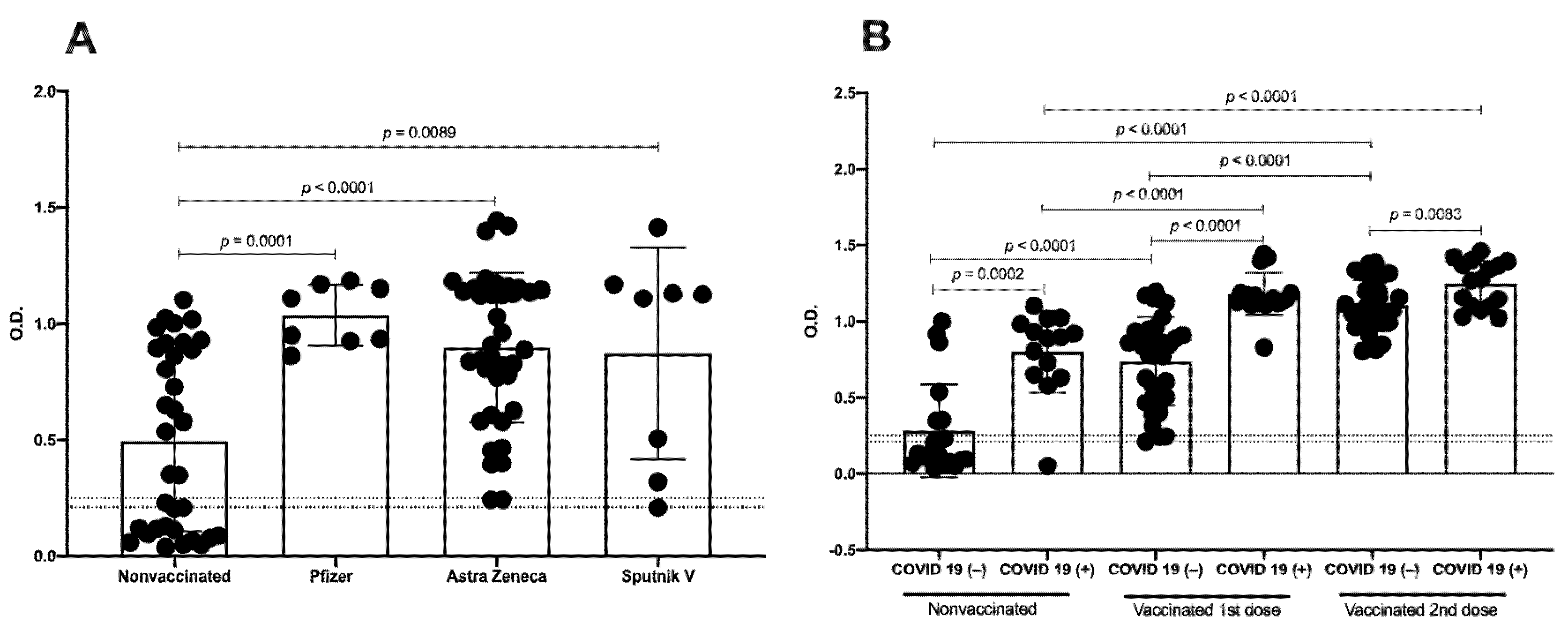
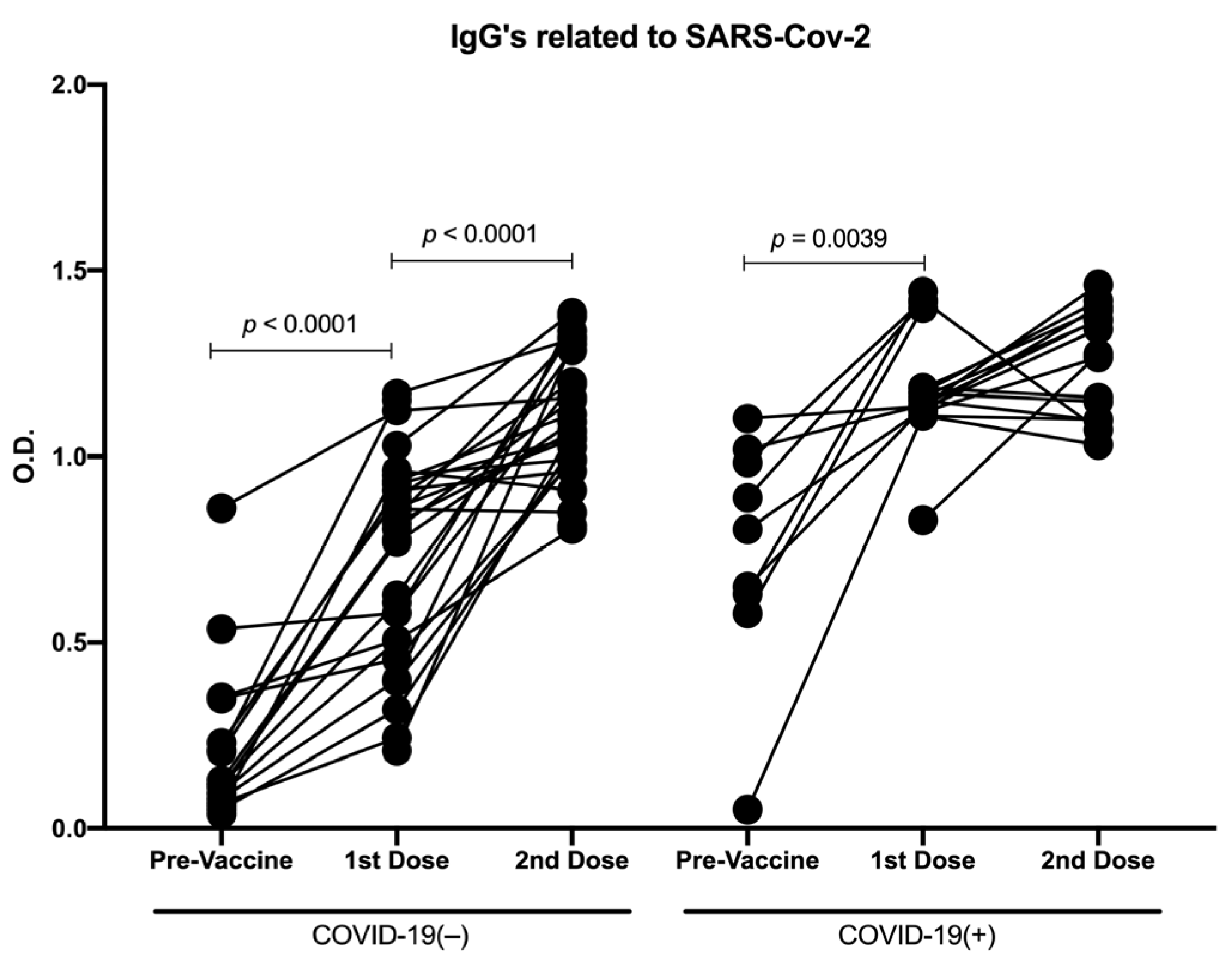
3.1. Distinct Vaccine Platforms Induced Similar Production of Antibodies against SARS-CoV-2
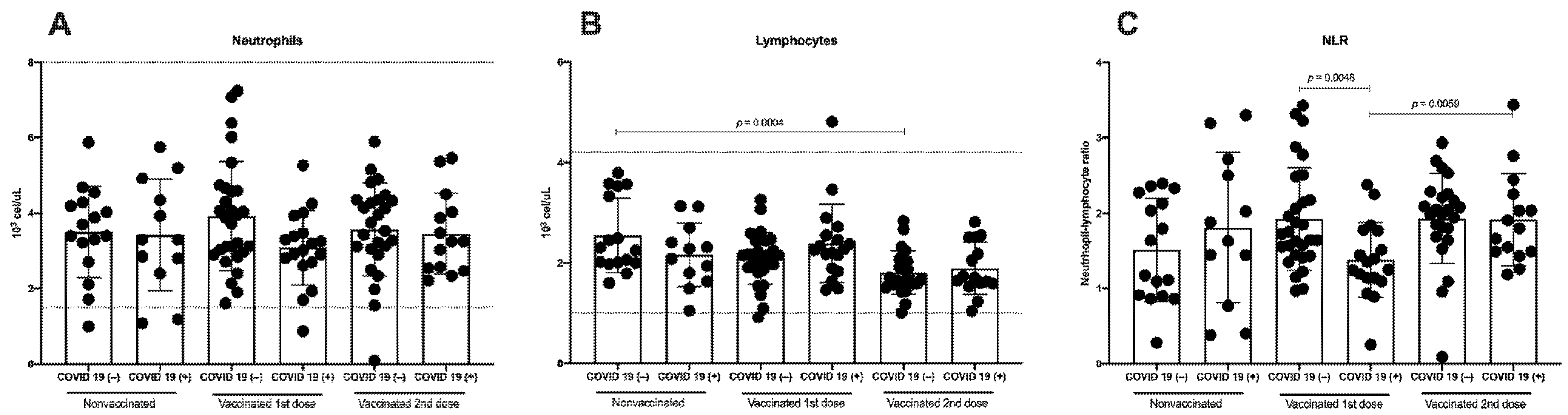
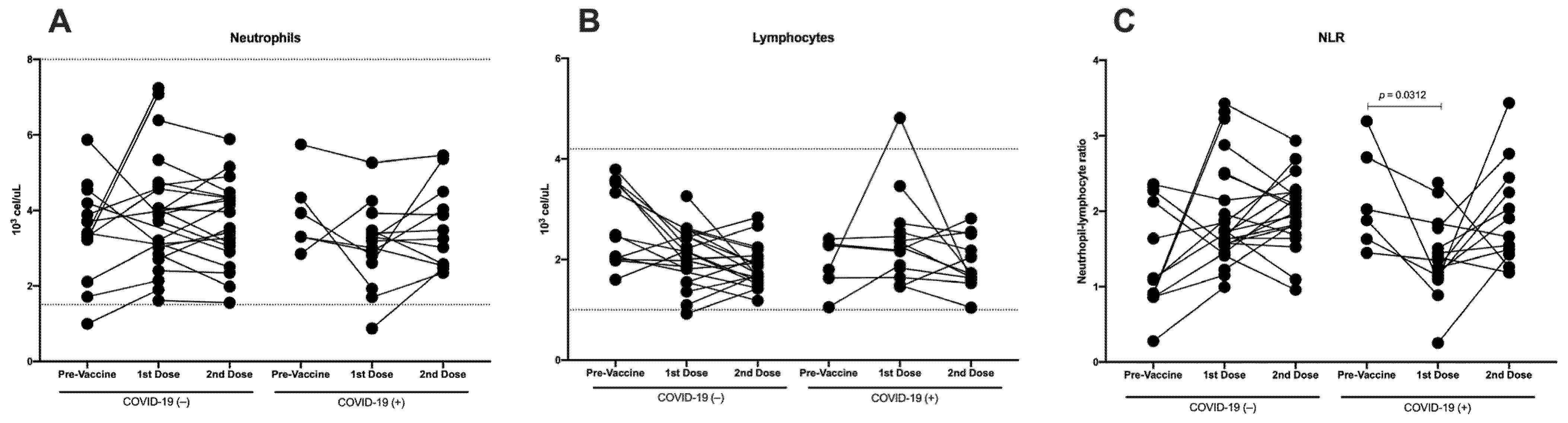
3.2. Immunization against SARS-CoV-2 Does Not Induce Differences in Total Neutrophils, Although a Decrease in Lymphocytes Is Detected
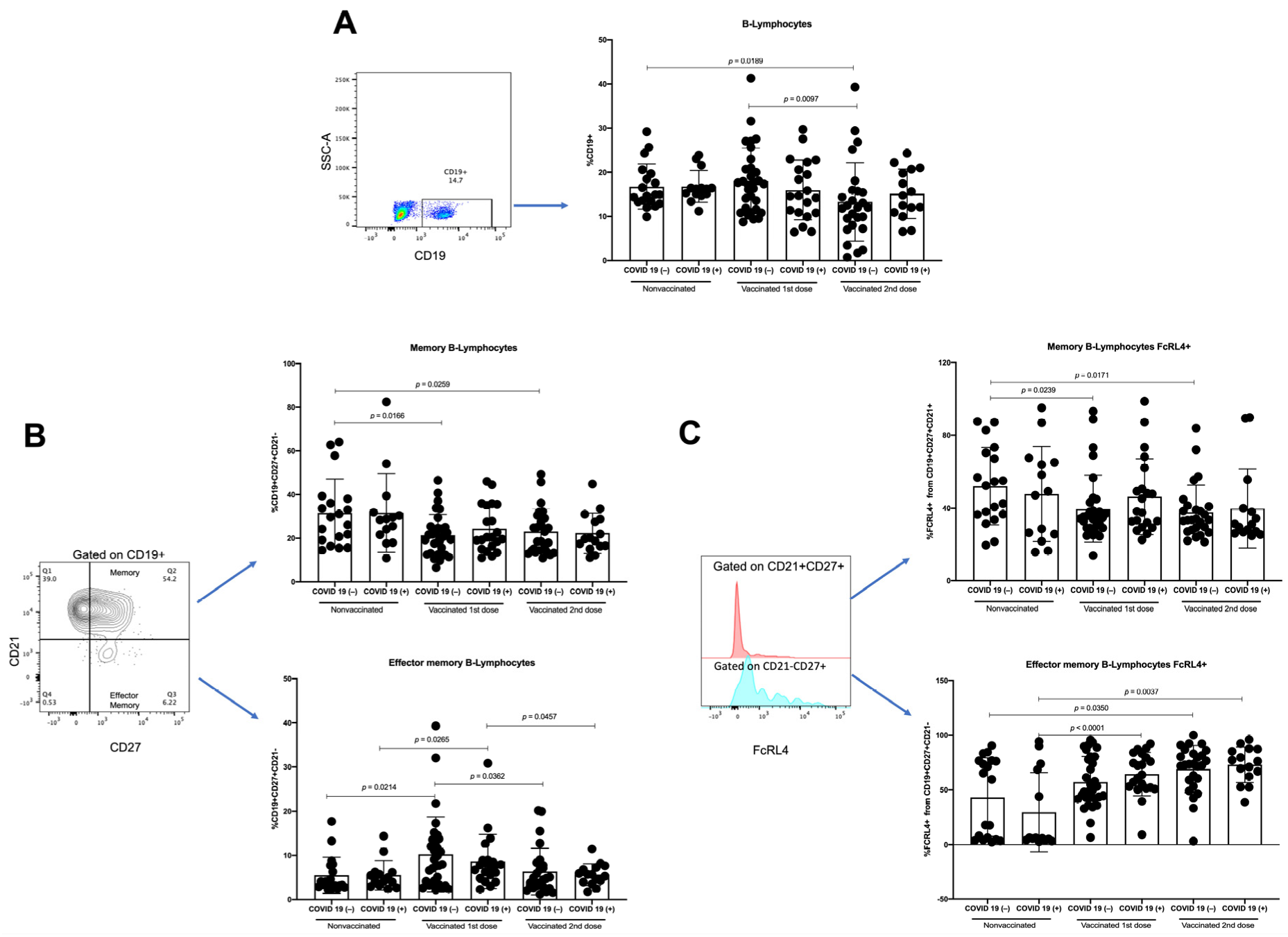
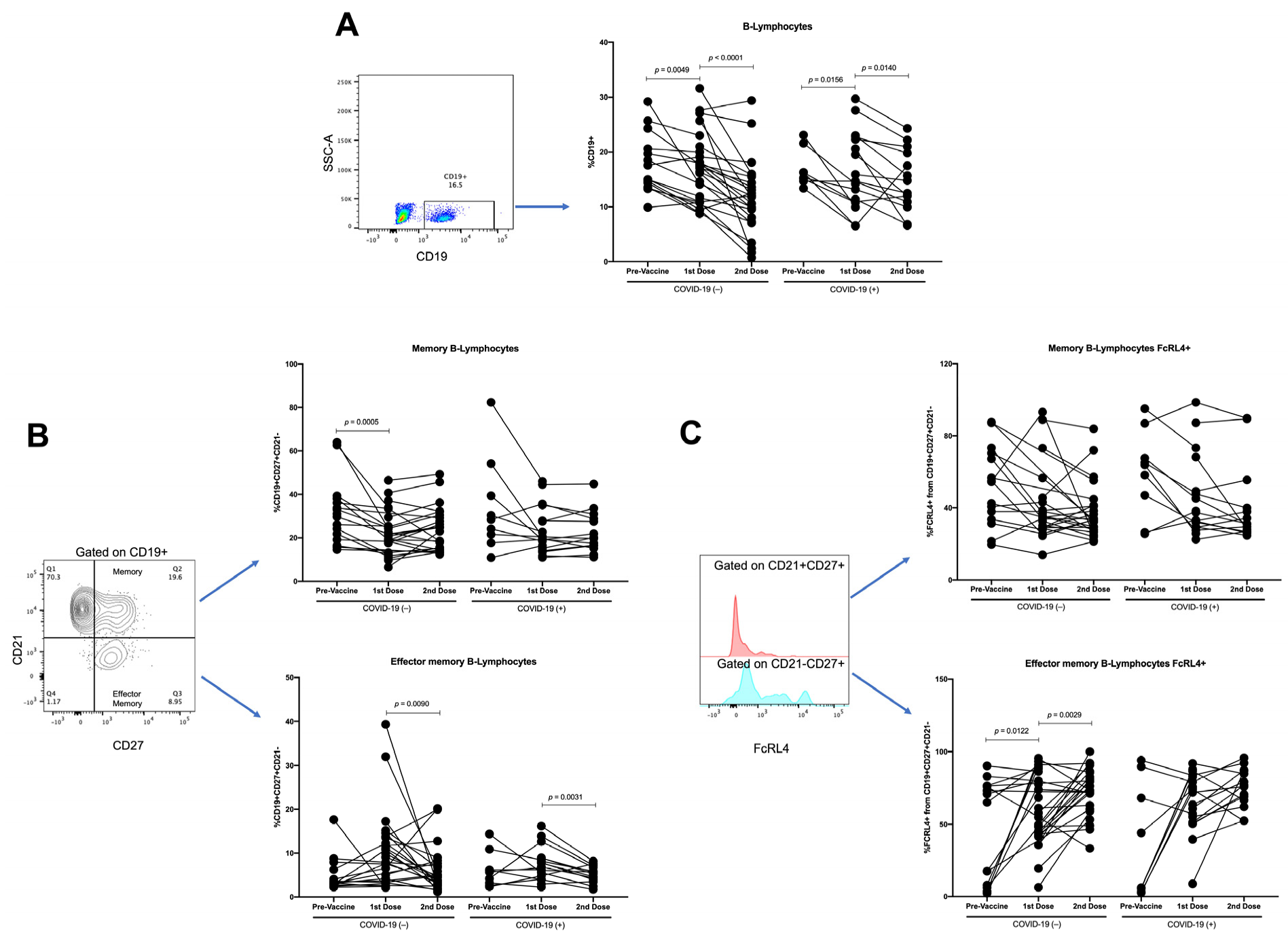
3.3. Effector-Memory B-Lymphocytes Increase with First Vaccine Dose, but There Are No Differences with Second Vaccine Dose
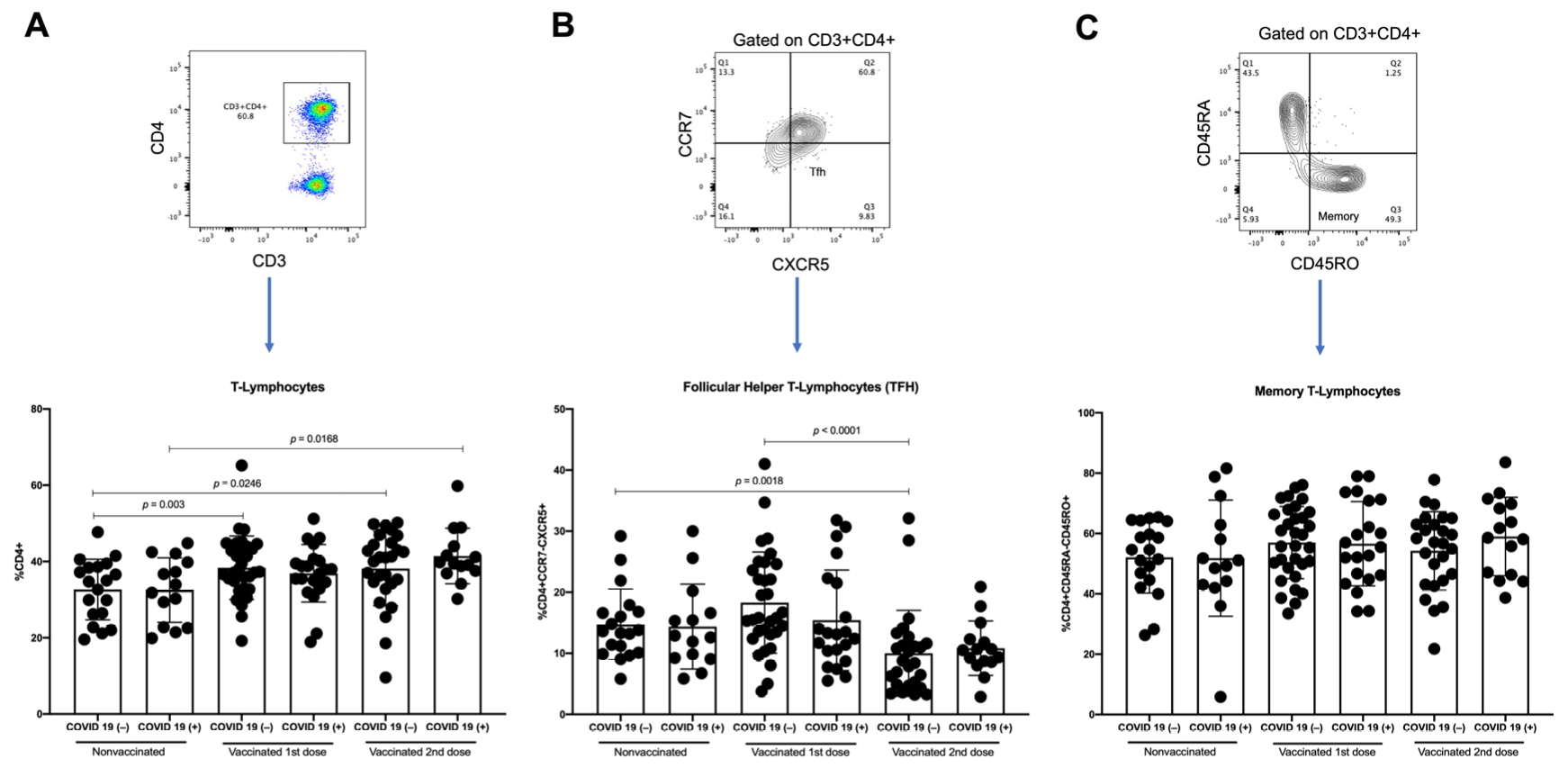
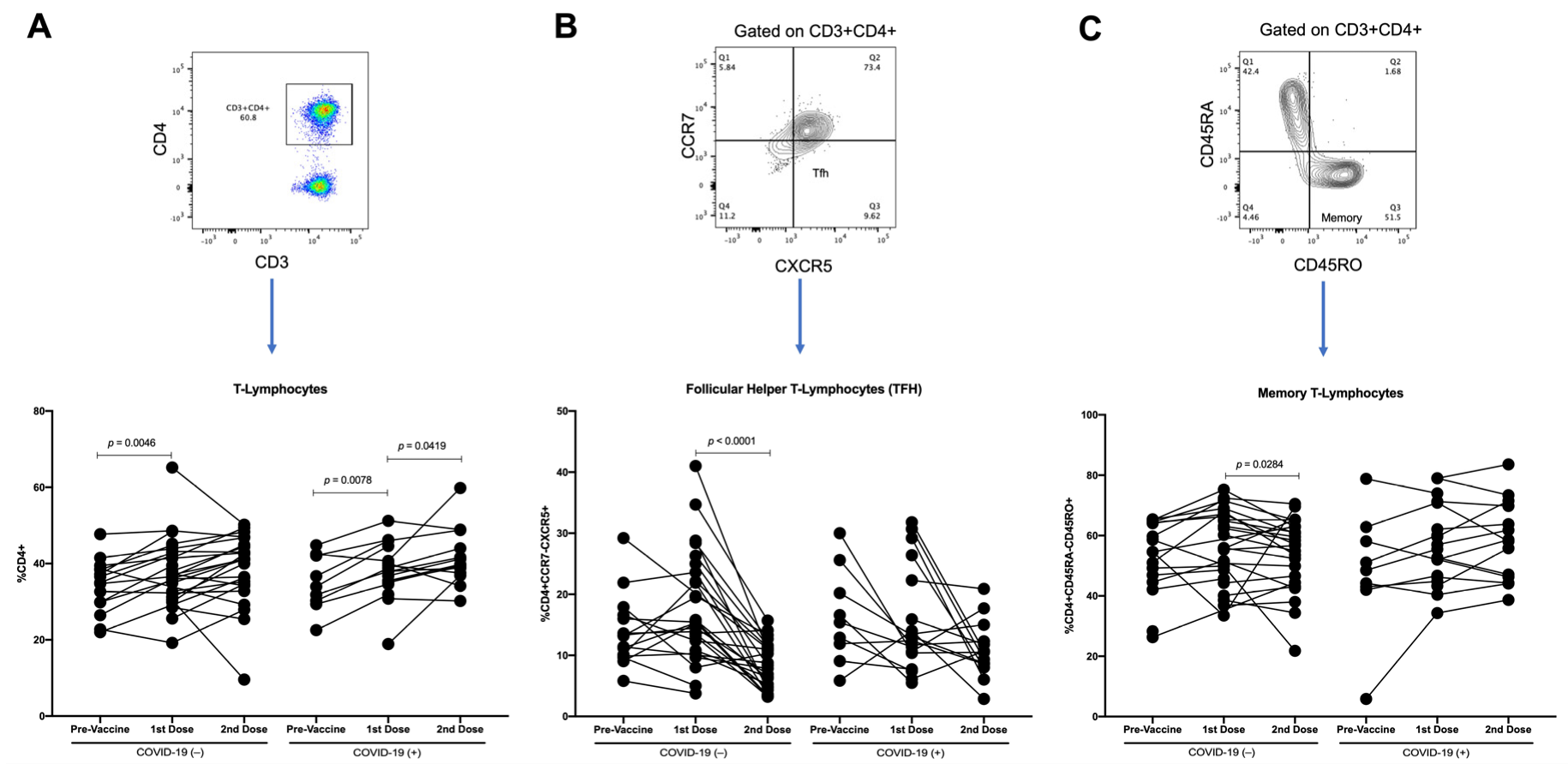
3.4. T-Lymphocytes and Follicular Helper T-Lymphocytes (TFH) Increase with First Dose of Vaccine against SARS-CoV-2 in COVID-19(−) Participants, and TFH Decreases with Second Dose
3.5. Changes in Parameters by Longitudinal Analysis throughout Follow-Up of Vaccination Schedule
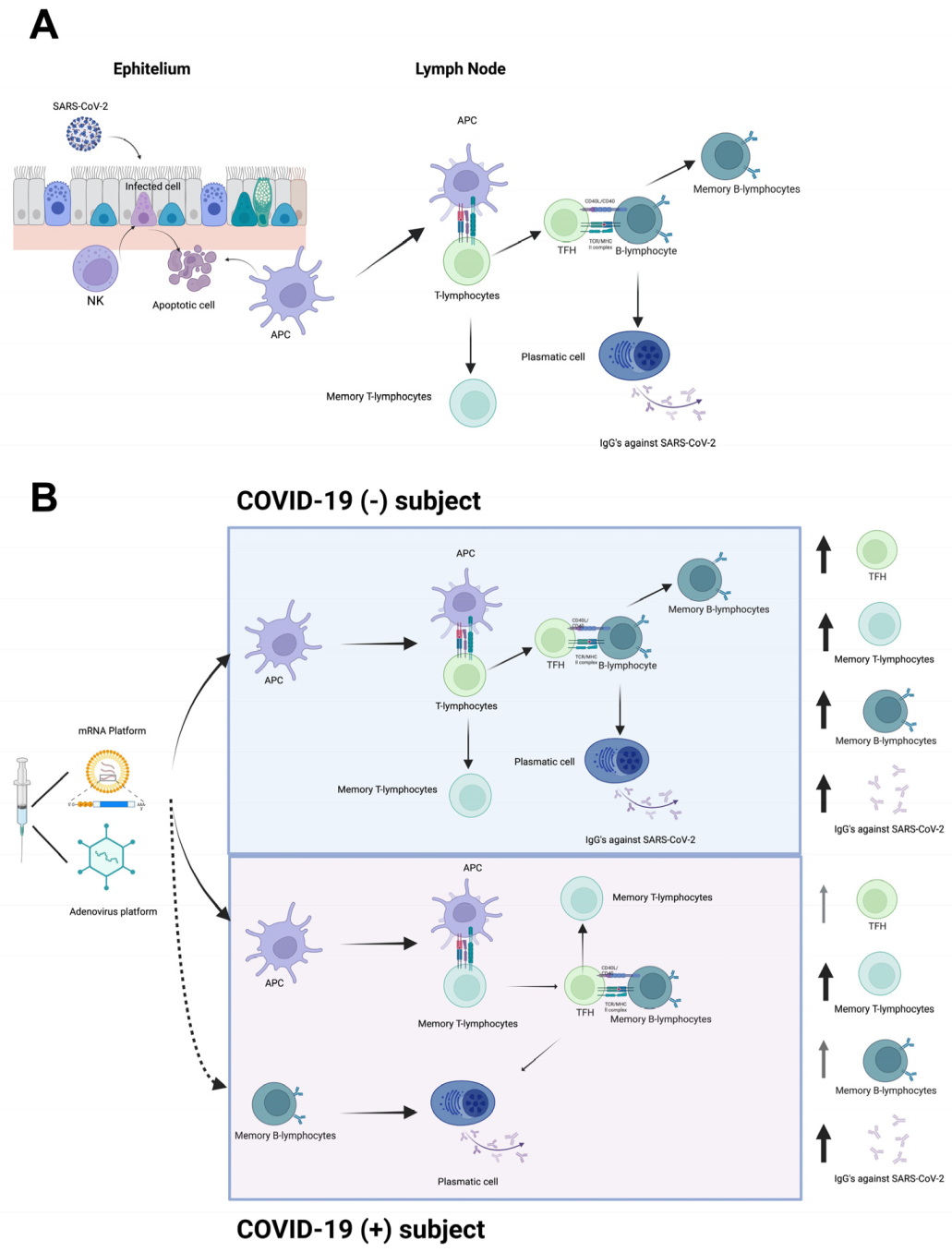
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, C.; Wang, Y.; Li, X.; Ren, L.; Zhao, J.; Hu, Y.; Zhang, L.; Fan, G.; Xu, J.; Gu, X.; et al. Clinical Features of Patients Infected with 2019 Novel Coronavirus in Wuhan, China. Lancet 2020, 395, 497–506. [Google Scholar] [CrossRef] [Green Version]
- Dan, J.M.; Mateus, J.; Kato, Y.; Hastie, K.M.; Yu, E.D.; Faliti, C.E.; Grifoni, A.; Ramirez, S.I.; Haupt, S.; Frazier, A.; et al. Immunological Memory to SARS-CoV-2 Assessed for up to 8 Months after Infection. Science 2021, 371, eabf4063. [Google Scholar] [CrossRef] [PubMed]
- Shurin, M.R.; Morris, A.; Wells, A.; Wheeler, S.E. Assessing Immune Response to SARS-CoV-2 Infection. Immunotargets Ther. 2020, 9, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of Spike Glycoprotein of SARS-CoV-2 on Virus Entry and Its Immune Cross-Reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ni, L.; Ye, F.; Cheng, M.-L.; Feng, Y.; Deng, Y.-Q.; Zhao, H.; Wei, P.; Ge, J.; Gou, M.; Li, X.; et al. Detection of SARS-CoV-2-Specific Humoral and Cellular Immunity in COVID-19 Convalescent Individuals. Immunity 2020, 52, 971–977.e3. [Google Scholar] [CrossRef] [PubMed]
- Grifoni, A.; Weiskopf, D.; Ramirez, S.I.; Mateus, J.; Dan, J.M.; Moderbacher, C.R.; Rawlings, S.A.; Sutherland, A.; Premkumar, L.; Jadi, R.S.; et al. Targets of T Cell Responses to SARS-CoV-2 Coronavirus in Humans with COVID-19 Disease and Unexposed Individuals. Cell 2020, 181, 1489–1501.e15. [Google Scholar] [CrossRef] [PubMed]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and Efficacy of the BNT162b2 MRNA COVID-19 Vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Baden, L.R.; El Sahly, H.M.; Essink, B.; Kotloff, K.; Frey, S.; Novak, R.; Diemert, D.; Spector, S.A.; Rouphael, N.; Creech, C.B.; et al. Efficacy and Safety of the MRNA-1273 SARS-CoV-2 Vaccine. N. Engl. J. Med. 2021, 384, 403–416. [Google Scholar] [CrossRef]
- Ewer, K.J.; Barrett, J.R.; Belij-Rammerstorfer, S.; Sharpe, H.; Makinson, R.; Morter, R.; Flaxman, A.; Wright, D.; Bellamy, D.; Bittaye, M.; et al. T Cell and Antibody Responses Induced by a Single Dose of ChAdOx1 NCoV-19 (AZD1222) Vaccine in a Phase 1/2 Clinical Trial. Nat. Med. 2021, 27, 270–278. [Google Scholar] [CrossRef]
- Zhu, F.-C.; Guan, X.-H.; Li, Y.-H.; Huang, J.-Y.; Jiang, T.; Hou, L.-H.; Li, J.-X.; Yang, B.-F.; Wang, L.; Wang, W.-J.; et al. Immunogenicity and Safety of a Recombinant Adenovirus Type-5-Vectored COVID-19 Vaccine in Healthy Adults Aged 18 Years or Older: A Randomised, Double-Blind, Placebo-Controlled, Phase 2 Trial. Lancet 2020, 396, 479–488. [Google Scholar] [CrossRef]
- Jarjour, N.N.; Masopust, D.; Jameson, S.C. T Cell Memory: Understanding COVID-19. Immunity 2021, 54, 14–18. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, S.E.; Shurin, G.V.; Yost, M.; Anderson, A.; Pinto, L.; Wells, A.; Shurin, M.R. Differential Antibody Response to MRNA COVID-19 Vaccines in Healthy Subjects. Microbiol. Spectr. 2021, 9, e0034121. [Google Scholar] [CrossRef] [PubMed]
- Camacho-Sandoval, R.; Nieto-Patlán, A.; Carballo-Uicab, G.; Montes-Luna, A.; Jiménez-Martínez, M.C.; Vallejo-Castillo, L.; González-González, E.; Arrieta-Oliva, H.I.; Gómez-Castellano, K.; Guzmán-Bringas, O.U.; et al. Development and Evaluation of a Set of Spike and Receptor Binding Domain-Based Enzyme-Linked Immunosorbent Assays for SARS-CoV-2 Serological Testing. Diagnostics 2021, 11, 1506. [Google Scholar] [CrossRef] [PubMed]
- Seyit, M.; Avci, E.; Nar, R.; Senol, H.; Yilmaz, A.; Ozen, M.; Oskay, A.; Aybek, H. Neutrophil to Lymphocyte Ratio, Lymphocyte to Monocyte Ratio and Platelet to Lymphocyte Ratio to Predict the Severity of COVID-19. Am. J. Emerg. Med. 2021, 40, 110–114. [Google Scholar] [CrossRef]
- Yang, A.-P.; Liu, J.-P.; Tao, W.-Q.; Li, H.-M. The Diagnostic and Predictive Role of NLR, d-NLR and PLR in COVID-19 Patients. Int. Immunopharmacol. 2020, 84, 106504. [Google Scholar] [CrossRef]
- Chams, N.; Chams, S.; Badran, R.; Shams, A.; Araji, A.; Raad, M.; Mukhopadhyay, S.; Stroberg, E.; Duval, E.J.; Barton, L.M.; et al. COVID-19: A Multidisciplinary Review. Front. Public Health 2020, 8, 383. [Google Scholar] [CrossRef]
- He, Q.; Mao, Q.; Zhang, J.; Bian, L.; Gao, F.; Wang, J.; Xu, M.; Liang, Z. COVID-19 Vaccines: Current Understanding on Immunogenicity, Safety, and Further Considerations. Front. Immunol. 2021, 12, 669339. [Google Scholar] [CrossRef]
- Fiolet, T.; Kherabi, Y.; MacDonald, C.-J.; Ghosn, J.; Peiffer-Smadja, N. Comparing COVID-19 Vaccines for Their Characteristics, Efficacy and Effectiveness against SARS-CoV-2 and Variants of Concern: A Narrative Review. Clin. Microbiol. Infect. 2022, 28, 202–221. [Google Scholar] [CrossRef]
- Gong, F.; Dai, Y.; Zheng, T.; Cheng, L.; Zhao, D.; Wang, H.; Liu, M.; Pei, H.; Jin, T.; Yu, D.; et al. Peripheral CD4+ T Cell Subsets and Antibody Response in COVID-19 Convalescent Individuals. J. Clin. Investig. 2020, 130, 6588–6599. [Google Scholar] [CrossRef]
- Rogliani, P.; Chetta, A.; Cazzola, M.; Calzetta, L. SARS-CoV-2 Neutralizing Antibodies: A Network Meta-Analysis across Vaccines. Vaccines 2021, 9, 227. [Google Scholar] [CrossRef]
- Kermali, M.; Khalsa, R.K.; Pillai, K.; Ismail, Z.; Harky, A. The Role of Biomarkers in Diagnosis of COVID-19–A Systematic Review. Life Sci. 2020, 254, 117788. [Google Scholar] [CrossRef] [PubMed]
- Huang, W.; Berube, J.; McNamara, M.; Saksena, S.; Hartman, M.; Arshad, T.; Bornheimer, S.J.; O’Gorman, M. Lymphocyte Subset Counts in COVID-19 Patients: A Meta-Analysis. Cytom. A 2020, 97, 772–776. [Google Scholar] [CrossRef] [PubMed]
- Lagunas-Rangel, F.A. Neutrophil-to-lymphocyte Ratio and Lymphocyte-to-C-reactive Protein Ratio in Patients with Severe Coronavirus Disease 2019 (COVID-19): A Meta-analysis. J. Med. Virol. 2020, 92, 1733–1734. [Google Scholar] [CrossRef] [Green Version]
- Montiel-Cervantes, L.A.; Medina, G.; Pilar Cruz-Domínguez, M.; Pérez-Tapia, S.-M.; Jiménez-Martínez, M.C.; Arrieta-Oliva, H.-I.; Carballo-Uicab, G.; López-Pelcastre, L.; Camacho-Sandoval, R. Poor Survival in COVID-19 Associated with Lymphopenia and Higher Neutrophile-Lymphocyte Ratio. Isr. Med. Assoc. J. 2021, 23, 153–159. [Google Scholar] [PubMed]
- Fathi, N.; Rezaei, N. Lymphopenia in COVID-19: Therapeutic Opportunities. Cell Biol. Int. 2020, 44, 1792–1797. [Google Scholar] [CrossRef]
- Sette, A.; Crotty, S. Adaptive Immunity to SARS-CoV-2 and COVID-19. Cell 2021, 184, 861–880. [Google Scholar] [CrossRef]
- Painter, M.M.; Mathew, D.; Goel, R.R.; Apostolidis, S.A.; Pattekar, A.; Kuthuru, O.; Baxter, A.E.; Herati, R.S.; Oldridge, D.A.; Gouma, S.; et al. Rapid Induction of Antigen-Specific CD4+ T Cells Is Associated with Coordinated Humoral and Cellular Immunity to SARS-CoV-2 MRNA Vaccination. Immunity 2021, 54, 2133–2142.e3. [Google Scholar] [CrossRef]
- Song, W.; Craft, J. T Follicular Helper Cell Heterogeneity: Time, Space, and Function. Immunol. Rev. 2019, 288, 85–96. [Google Scholar] [CrossRef]
- Sandberg, J.T.; Varnaitė, R.; Christ, W.; Chen, P.; Muvva, J.R.; Maleki, K.T.; García, M.; Dzidic, M.; Folkesson, E.; Skagerberg, M.; et al. SARS-CoV-2-specific Humoral and Cellular Immunity Persists through 9 Months Irrespective of COVID-19 Severity at Hospitalisation. Clin. Transl. Immunol. 2021, 10, e1306. [Google Scholar] [CrossRef]
- Acet Öztürk, N.A.; Ursavaş, A.; Görek DïLektaşli, A.; DemïRdöğen, E.; Coşkun, N.F.; EdïGer, D.; Uzaslan, A.E.; Yöyen-Ermïş, D.; Karaca, M.; Terzï, O.E.; et al. Interleukin-21: A Potential Biomarker for Diagnosis and Predicting Prognosis in COVID-19 Patients. Turk. J. Med. Sci. 2021, 51, 2274–2284. [Google Scholar] [CrossRef]
- Röltgen, K.; Boyd, S.D. Antibody and B Cell Responses to SARS-CoV-2 Infection and Vaccination. Cell Host Microbe 2021, 29, 1063–1075. [Google Scholar] [CrossRef] [PubMed]
- Quast, I.; Tarlinton, D. B Cell Memory: Understanding COVID-19. Immunity 2021, 54, 205–210. [Google Scholar] [CrossRef] [PubMed]
- Sohn, H.W.; Krueger, P.D.; Davis, R.S.; Pierce, S.K. FcRL4 Acts as an Adaptive to Innate Molecular Switch Dampening BCR Signaling and Enhancing TLR Signaling. Blood 2011, 118, 6332–6341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siewe, B.; Nipper, A.J.; Sohn, H.; Stapleton, J.T.; Landay, A. FcRL4 Expression Identifies a Pro-Inflammatory B Cell Subset in Viremic HIV-Infected Subjects. Front. Immunol. 2017, 8, 1339. [Google Scholar] [CrossRef] [Green Version]
- Yeo, L.; Lom, H.; Juarez, M.; Snow, M.; Buckley, C.D.; Filer, A.; Raza, K.; Scheel-Toellner, D. Expression of FcRL4 Defines a Pro-Inflammatory, RANKL-Producing B Cell Subset in Rheumatoid Arthritis. Ann. Rheum. Dis. 2015, 74, 928–935. [Google Scholar] [CrossRef] [Green Version]
- Amara, K.; Clay, E.; Yeo, L.; Ramsköld, D.; Spengler, J.; Sippl, N.; Cameron, J.A.; Israelsson, L.; Titcombe, P.J.; Grönwall, C.; et al. B Cells Expressing the IgA Receptor FcRL4 Participate in the Autoimmune Response in Patients with Rheumatoid Arthritis. J. Autoimmun. 2017, 81, 34–43. [Google Scholar] [CrossRef]
- Amanna, I.J.; Carlson, N.E.; Slifka, M.K. Duration of Humoral Immunity to Common Viral and Vaccine Antigens. N. Engl. J. Med. 2007, 357, 1903–1915. [Google Scholar] [CrossRef] [Green Version]
- Netea, M.G.; Schlitzer, A.; Placek, K.; Joosten, L.A.B.; Schultze, J.L. Innate and Adaptive Immune Memory: An Evolutionary Continuum in the Host’s Response to Pathogens. Cell Host Microbe 2019, 25, 13–26. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Not Vaccinated n = 34 | Vaccinated 1st Dose n = 54 | Vaccinated 2nd Dose n = 43 | |
|---|---|---|---|
| Age (years) Mean + range | 35 (20–49) | 37 (20–62) | 39 (23–70) |
| Men | 14 (41%) | 18 (33%) | 15 (34%) |
| Age (years) Mean + range | 35 (20–49) | 36 (20–51) | 36 (24–51) |
| Women | 20 (59%) | 36 (67%) | 28 (66%) |
| Age (years) Mean + range | 35 (20–49) | 37 (23–62) | 39 (23–60) |
| COVID-19 positive | 14 (41%) | 21 (38%) | 15 (36%) |
| COVID-19 negative | 20 (59%) | 33 (62%) | 28 (64%) |
| Pfizer-BioNTech (BNT162b2) | - | 8 (15%) | 11 (25%) |
| Astra Zeneca (AZD1222 Covishield) | - | 38 (70%) | 27 (63%) |
| Sputnik V (Gam-COVID-Vac) | - | 8 (15%) | 5 (12%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islas-Vazquez, L.; Cruz-Aguilar, M.; Velazquez-Soto, H.; Jiménez-Corona, A.; Pérez-Tapia, S.M.; Jimenez-Martinez, M.C. Effector-Memory B-Lymphocytes and Follicular Helper T-Lymphocytes as Central Players in the Immune Response in Vaccinated and Nonvaccinated Populations against SARS-CoV-2. Vaccines 2022, 10, 1761. https://doi.org/10.3390/vaccines10101761
Islas-Vazquez L, Cruz-Aguilar M, Velazquez-Soto H, Jiménez-Corona A, Pérez-Tapia SM, Jimenez-Martinez MC. Effector-Memory B-Lymphocytes and Follicular Helper T-Lymphocytes as Central Players in the Immune Response in Vaccinated and Nonvaccinated Populations against SARS-CoV-2. Vaccines. 2022; 10(10):1761. https://doi.org/10.3390/vaccines10101761
Chicago/Turabian StyleIslas-Vazquez, Lorenzo, Marisa Cruz-Aguilar, Henry Velazquez-Soto, Aida Jiménez-Corona, Sonia Mayra Pérez-Tapia, and Maria C. Jimenez-Martinez. 2022. "Effector-Memory B-Lymphocytes and Follicular Helper T-Lymphocytes as Central Players in the Immune Response in Vaccinated and Nonvaccinated Populations against SARS-CoV-2" Vaccines 10, no. 10: 1761. https://doi.org/10.3390/vaccines10101761