

Post-Vaccination Neutralization Responses to Omicron Sub-Variants
 , ,
, ,
Abstract
:1. Introduction
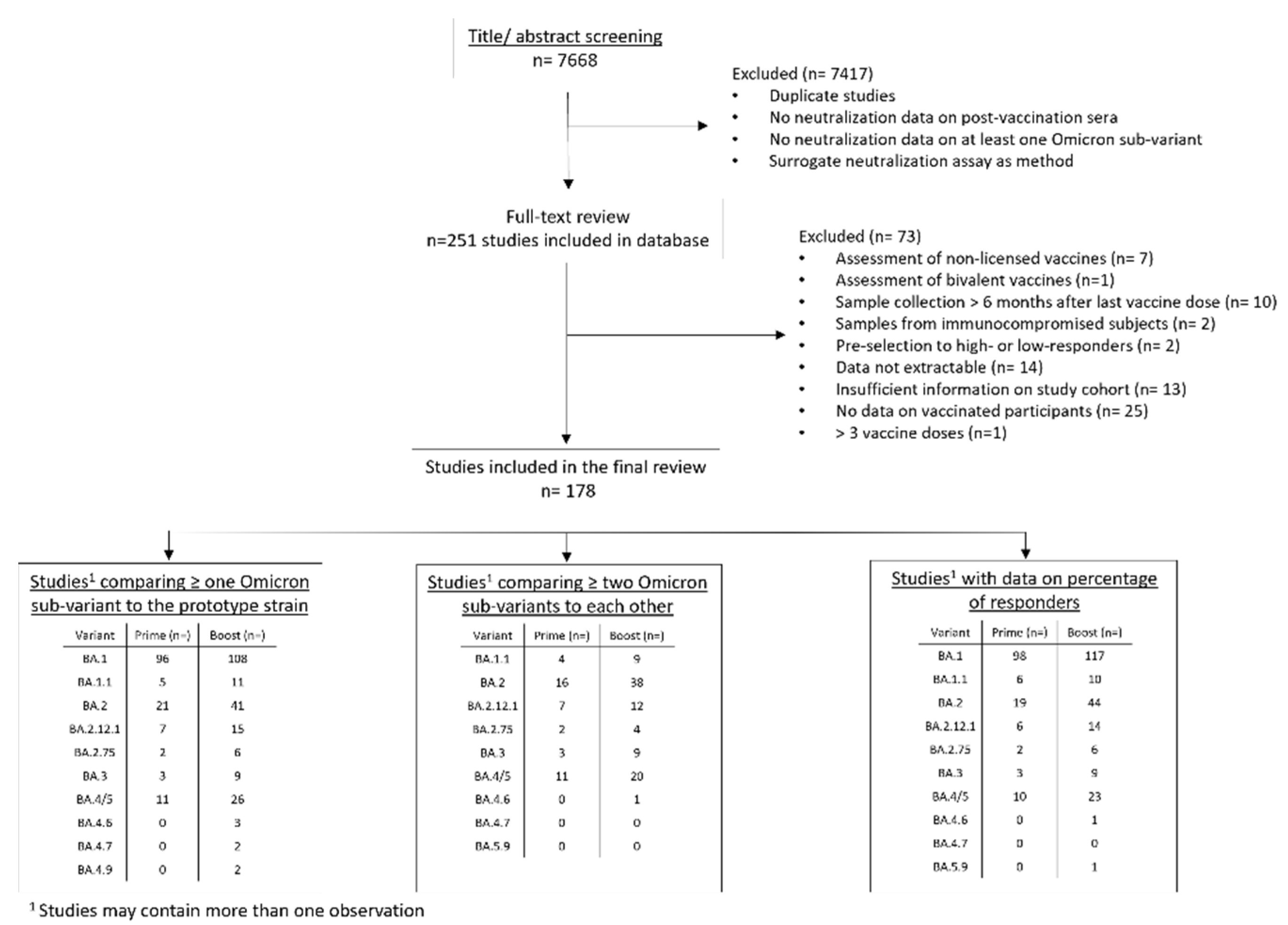
2. Methods
3. Results
3.1. Percentage of Samples with Titers above the Limit of Detection (Percentage of Responders)
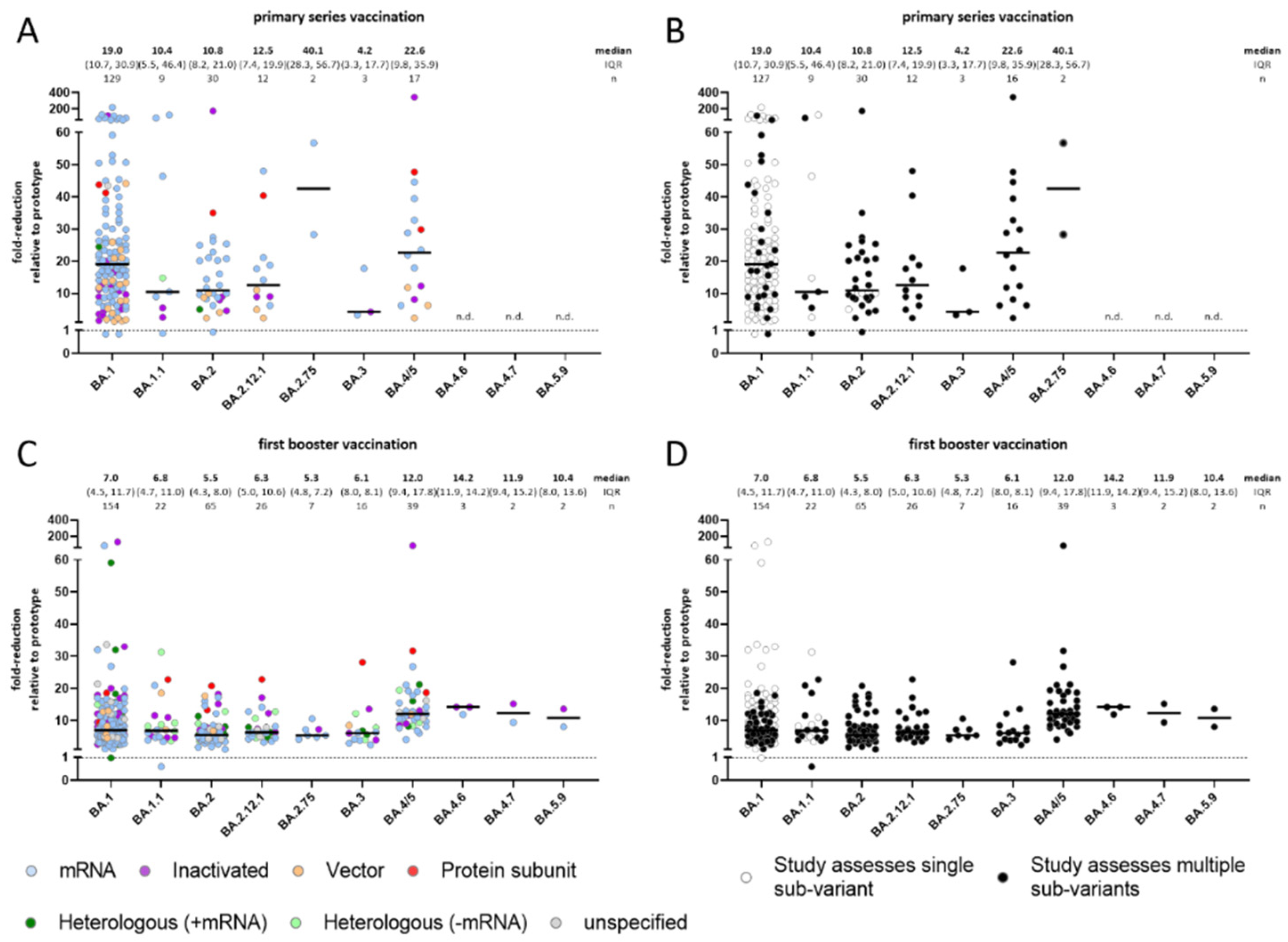
3.2. Comparison of Omicron Sub-Variants to the Prototype (WT) Strain
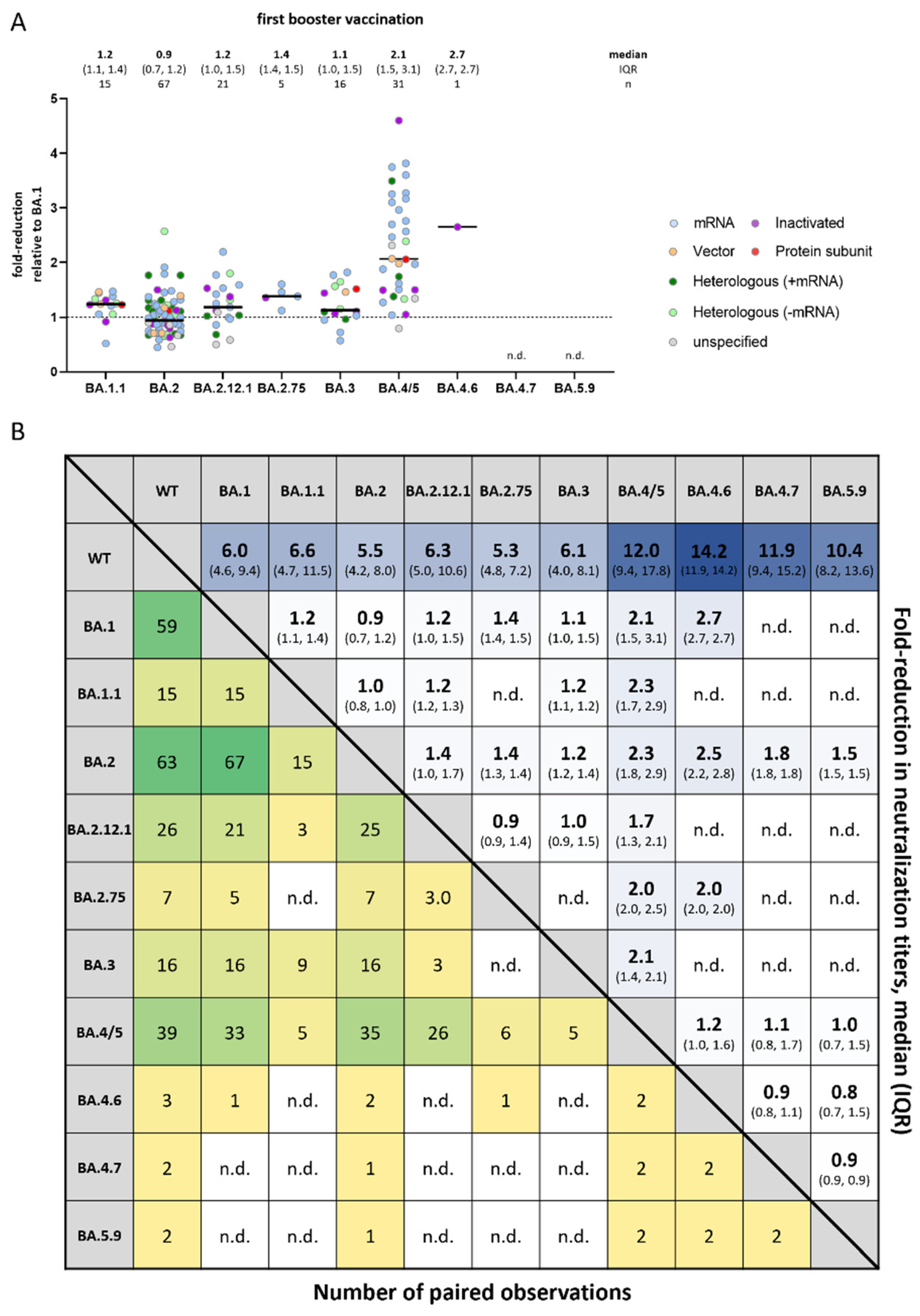
3.3. Comparison of Omicron Sub-Variants to Each Other
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Walker, A.S.; Vihta, K.-D.; Gethings, O.; Pritchard, E.; Jones, J.; House, T.; Bell, I.; Bell, J.; Newton, J.N.; Farrar, J. Tracking the Emergence of SARS-CoV-2 Alpha Variant in the United Kingdom. N. Engl. J. Med. 2021, 385, 2582–2585. [Google Scholar] [CrossRef] [PubMed]
- WHO. Weekly Epidemiological Update on COVID-19–29 June 2022. 2022. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---29-june-2022 (accessed on 1 October 2022).
- Edara, V.V.; Manning, K.E.; Ellis, M.; Lai, L.; Moore, K.M.; Foster, S.L.; Floyd, K.; Daves-Gardner, M.E.; Mantus, G.; Nyhoff, L.E. mRNA-1273 and BNT162b2 mRNA vaccines have reduced neutralizing activity against the SARS-CoV-2 omicron variant. Cell Rep. Med. 2022, 3, 100529. [Google Scholar] [CrossRef] [PubMed]
- Cameroni, E.; Bowen, J.E.; Rosen, L.E.; Saliba, C.; Zepeda, S.K.; Culap, K.; Pinto, D.; Van Blargan, L.A.; De Marco, A.; di Iulio, J. Broadly neutralizing antibodies overcome SARS-CoV-2 Omicron antigenic shift. Nature 2022, 602, 664–670. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.M.S.; Mok, C.K.P.; Leung, Y.W.Y.; Ng, S.S.; Chan, K.C.K.; Ko, F.W.; Chen, C.; Yio, K.; Lam, B.H.S.; Lau, E.H.Y. Neutralizing antibodies against the SARS-CoV-2 Omicron variant BA.1 following homologous and heterologous CoronaVac or BNT162b2 vaccination. Nat. Med. 2022, 28, 486–489. [Google Scholar] [CrossRef] [PubMed]
- Higdon, M.M.; Baidya, A.; Walter, K.K.; Patel, M.K.; Issa, H.; Espié, E.; Feikin, D.R.; Deloria Knoll, M. Duration of effectiveness of vaccination against COVID-19 caused by the omicron variant. Nat. Med. 2022, 28, 486–489. [Google Scholar] [CrossRef]
- Nordström, P.; Ballin, M.; Nordström, A. Risk of infection, hospitalisation, and death up to 9 months after a second dose of COVID-19 vaccine: A retrospective, total population cohort study in Sweden. Lancet 2022, 399, 814–823. [Google Scholar] [CrossRef]
- Goldberg, Y.; Mandel, M.; Bar-On, Y.M.; Bodenheimer, O.; Freedman, L.S.; Ash, N.; Alroy-Preis, S.; Huppert, A.; Milo, R. Protection and Waning of Natural and Hybrid Immunity to SARS-CoV-2. N. Engl. J. Med. 2022, 386, 2201–2212. [Google Scholar] [CrossRef] [PubMed]
- Planas, D.; Saunders, L.; Maes, P.; Guivel-Benhassine, F.; Planchais, C.; Buchrieser, J.; Bolland, W.-H.; Porrot, F.; Staropoli, I.; Lemoine, F. Considerable escape of SARS-CoV-2 Omicron to antibody neutralization. Nature 2022, 602, 671–675. [Google Scholar] [CrossRef]
- Andrews, N.; Stowe, J.; Kirsebom, F.; Toffa, S.; Rickeart, T.; Gallagher, E.; Gower, C.; Kall, M.; Groves, N.; O’Connel, A.-M. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N. Engl. J. Med. 2022, 386, 1532–1546. [Google Scholar] [CrossRef]
- Kurhade, C.; Zou, J.; Xia, H.; Cai, H.; Yang, Q.; Cutler, M.; Cooper, D.; Muik, A.; Jansen, K.U.; Xie, X. Neutralization of Omicron BA.1, BA.2, and BA.3 SARS-CoV-2 by 3 doses of BNT162b2 vaccine. Nat. Commun. 2022, 13, 3602. [Google Scholar] [CrossRef]
- Tegally, H.; Moir, M.; Everatt, J.; Giovanetti, M.; Scheepers, C.; Wilkinson, E.; Subramoney, K.; Moyo, S.; Amoako, D.G.; Baxter, C. Continued Emergence and Evolution of Omicron in South Africa: New BA.4 and BA.5 lineages. medRxiv 2022. [Google Scholar] [CrossRef]
- Wise, J. Covid-19: Omicron sub variants driving new wave of infections in UK. BMJ 2022, 377, o1506. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Phillips, D.J.; White, T.; Sayal, H.; Aley, P.K.; Bibi, S.; Dold, D.; Fuskova, M.; Gilbert, S.C.; Hirsch, I. Correlates of protection against symptomatic and asymptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 2032–2040. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F. A correlate of protection for SARS-CoV-2 vaccines is urgently needed. Nat. Med. 2021, 27, 1147–1148. [Google Scholar] [CrossRef]
- Khoury, D.S.; Cromer, D.; Reynaldi, A.; Schlub, T.E.; Wheatley, A.K.; Juno, J.A.; Subbarao, K.; Kent, S.J.; Triccas, J.A.; Davenport, M.P. Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 2021, 27, 1205–1211. [Google Scholar] [CrossRef]
- Evans, J.P.; Zeng, C.; Carlin, C.; Lozanski, G.; Saif, L.J.; Oltz, E.M.; Gumina, R.J.; Liu, S.-L. Neutralizing antibody responses elicited by SARS-CoV-2 mRNA vaccination wane over time and are boosted by breakthrough infection. Sci. Transl. Med. 2022, 14, eabn8057. [Google Scholar] [CrossRef]
- Rössler, A.; Riepler, L.; Batne, D.; von Laer, D.; Kimpel, J. SARS-CoV-2 Omicron Variant Neutralization in Serum from Vaccinated and Convalescent Persons. N. Engl. J. Med. 2022, 386, 698–700. [Google Scholar] [CrossRef]
- Evans, J.P.; Zeng, C.; Qu, P.; Faraone, J.; Zheng, Y.-M.; Garlin, C.; Bednash, J.S.; Zhou, T.; Lozanski, G.; Mallampalli, R. Neutralization of SARS-CoV-2 Omicron sub-lineages BA.1, BA.1.1, and BA.2. Cell Host Microbe 2022, 30, 1093–1102.e3. [Google Scholar] [CrossRef]
- Dejnirattisai, W.; Shaw, R.H.; Supasa, P.; Liu, C.; Stuart, A.S.; Pollard, A.J.; Liu, X.; Lambe, T.; Crook, D.; Stuart, D.I. Reduced neutralisation of SARS-CoV-2 omicron B.1.1.529 variant by post-immunisation serum. Lancet 2022, 399, 234–236. [Google Scholar] [CrossRef]
- Wu, M.; Wall, E.C.; Carr, E.J.; Harvey, R.; Townley, H.; Mears, H.V.; Adams, L.; Kjaer, S.; Kelly, G.; Warchal, S. Three-dose vaccination elicits neutralising antibodies against omicron. Lancet 2022, 399, 715–717. [Google Scholar] [CrossRef]
- Zhao, Y.; Zhao, X.; Zhang, R.; Ye, B.; Yuan, X.; Zhang, D.; Li, L.; Tian, J.; Guo, Y.; Gao, G.F.; et al. Heterologous BBIBP-CorV/ZF2001 vaccination augments neutralization against SARS-CoV-2 variants: A preliminary observation. Lancet Reg. Health–West. Pac. 2022, 21, 100440. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, H.; Sitaras, I.; Jurgensmeyer, M.; Mulders, M.N.; Goldblatt, D.; Feiking, D.R.; Bar-Zeev, N.; Higdon, M.M.; Deloria Knoll, M. Assessing the Reliability of SARS-CoV-2 Neutralization Studies That Use Post-Vaccination Sera. Vaccines 2022, 10, 850. [Google Scholar] [CrossRef]
- WHO. COVID-19 Weekly Epidemiological Update, Edition 76. 2022. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---25-january-2022 (accessed on 1 October 2022).
- WHO. Weekly Epidemiological Update on COVID-19–6 July 2022. 2022. Available online: https://www.who.int/publications/m/item/weekly-epidemiological-update-on-covid-19---6-july-2022 (accessed on 1 October 2022).
- Altarawneh, H.N.; Chemaitelly, H.; Ayoub, H.H.; Hasan, M.R.; Coyle, P.; Yassine, H.M.; Al-Khatib, H.A.; Benslimane, F.M.; Al-Kanaani, Z.; Al-Kuwari, E. Protection of SARS-CoV-2 natural infection against reinfection with the Omicron BA.4 or BA.5 subvariants. medRxiv 2022. [Google Scholar] [CrossRef]
- Bates, T.A.; McBridge, S.K.; Leier, H.C.; Guzman, G.; Lyski, Z.L.; Schoen, D.; Winders, B.; Lee, J.-Y.; Lee, D.X.; Messer, W.B.; et al. Vaccination before or after SARS-CoV-2 infection leads to robust humoral response and antibodies that effectively neutralize variants. Sci. Immunol. 2022, 7, eabn8014. [Google Scholar] [CrossRef] [PubMed]
- Canaday, D.H.; Qyebanji, O.A.; White, E.; Keresztesy, D.; Payne, M.; Wilk, D.; Carias, L.; Aung, H.; St. Denis, K.; Sheehan, M.L.; et al. COVID-19 vaccine booster dose needed to achieve Omicron-specific neutralisation in nursing home residents. EBioMedicine 2022, 80, 104066. [Google Scholar] [CrossRef] [PubMed]
- Chemaitelly, H.; Nagelkerke, N.; Ayoub, H.H.; Coyle, P.; Tang, P.; Yassine, H.M.; Al-Khatib, H.A.; Smatti, M.K.; Hasan, M.R.; Al-Kanaani, Z.; et al. Duration of immune protection of SARS-CoV-2 natural infection against reinfection in Qatar. medRxiv 2022. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Vaccine Platform | Omicron Sub-variant Compared to Prototype | Primary Series Vaccination | First Booster Vaccination | ||||
|---|---|---|---|---|---|---|---|
| Studies | Observations | Sera | Studies | Observations | Sera | ||
| mRNA | BA.1 | 69 | 87 | 1748 | 76 | 86 | 2368 |
| BA.1.1 | 3 | 6 | 103 | 6 | 8 | 119 | |
| BA.2 | 17 | 20 | 323 | 33 | 34 | 629 | |
| BA.2.12.1 | 5 | 6 | 124 | 12 | 12 | 255 | |
| BA.2.75 | 2 | 2 | 35 | 5 | 6 | 134 | |
| BA.3 | 2 | 2 | 20 | 6 | 6 | 97 | |
| BA.4/5 | 8 | 9 | 164 | 20 | 21 | 738 | |
| BA.4.6 | 0 | 0 | 0 | 1 | 1 | 15 | |
| BA.4.7 | 0 | 0 | 0 | 1 | 1 | 15 | |
| BA.5.9 | 0 | 0 | 0 | 1 | 1 | 15 | |
| Inactivated | BA.1 | 15 | 16 | 657 | 24 | 24 | 1118 |
| BA.1.1 | 2 | 2 | 33 | 5 | 5 | 156 | |
| BA.2 | 4 | 4 | 59 | 9 | 10 | 229 | |
| BA.2.12.1 | 2 | 2 | 37 | 4 | 4 | 101 | |
| BA.2.75 | 0 | 0 | 0 | 1 | 1 | 40 | |
| BA.3 | 1 | 1 | 10 | 3 | 3 | 76 | |
| BA.4/5 | 3 | 3 | 49 | 6 | 6 | 153 | |
| BA.4.6 | 0 | 0 | 0 | 2 | 2 | 80 | |
| BA.4.7 | 0 | 0 | 0 | 1 | 1 | 40 | |
| BA.5.9 | 0 | 0 | 0 | 1 | 1 | 40 | |
| Vector | BA.1 | 17 | 20 | 334 | 5 | 5 | 115 |
| BA.1.1 | 0 | 0 | 0 | 2 | 2 | 82 | |
| BA.2 | 3 | 4 | 47 | 2 | 2 | 82 | |
| BA.2.12.1 | 2 | 3 | 37 | 0 | 0 | 0 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 1 | 1 | 41 | |
| BA.4/5 | 2 | 3 | 37 | 1 | 1 | 41 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Protein Subunit | BA.1 | 2 | 2 | 39 | 4 | 4 | 96 |
| BA.1.1 | 0 | 0 | 0 | 1 | 1 | 20 | |
| BA.2 | 1 | 1 | 10 | 1 | 2 | 40 | |
| BA.2.12.1 | 1 | 1 | 10 | 1 | 1 | 20 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 1 | 1 | 20 | |
| BA.4/5 | 2 | 2 | 39 | 2 | 2 | 68 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Heterologous (+mRNA) | BA.1 | 2 | 3 | 114 | 14 | 18 | 1029 |
| BA.1.1 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.2 | 1 | 1 | 10 | 6 | 8 | 165 | |
| BA.2.12.1 | 0 | 0 | 0 | 2 | 3 | 56 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 1 | 2 | 30 | |
| BA.4/5 | 0 | 0 | 0 | 2 | 3 | 56 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Heterologous (-mRNA) | BA.1 | 0 | 0 | 24 | 8 | 9 | 234 |
| BA.1.1 | 1 | 1 | 24 | 5 | 6 | 186 | |
| BA.2 | 0 | 0 | 0 | 4 | 5 | 126 | |
| BA.2.12.1 | 0 | 0 | 0 | 3 | 3 | 88 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 3 | 3 | 76 | |
| BA.4/5 | 0 | 0 | 0 | 3 | 3 | 88 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Unspecified | BA.1 | 1 | 1 | 48 | 6 | 8 | 321 |
| BA.1.1 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.2 | 0 | 0 | 0 | 2 | 4 | 79 | |
| BA.2.12.1 | 0 | 0 | 0 | 1 | 3 | 35 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4/5 | 0 | 0 | 0 | 1 | 3 | 35 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total | BA.1 | 106 | 129 | 2964 | 137 | 154 | 5281 |
| BA.1.1 | 6 | 9 | 160 | 19 | 22 | 563 | |
| BA.2 | 26 | 30 | 449 | 57 | 65 | 1350 | |
| BA.2.12.1 | 10 | 12 | 208 | 23 | 26 | 555 | |
| BA.2.75 | 2 | 2 | 35 | 6 | 7 | 174 | |
| BA.3 | 3 | 3 | 30 | 15 | 16 | 340 | |
| BA.4/5 | 15 | 17 | 289 | 35 | 39 | 1179 | |
| BA.4.6 | 0 | 0 | 0 | 3 | 3 | 95 | |
| BA.4.7 | 0 | 0 | 0 | 2 | 2 | 55 | |
| BA.5.9 | 0 | 0 | 0 | 2 | 2 | 55 | |
| Vaccine Platform | Omicron Sub-variant Compared to BA.1 | Primary Series Vaccination | First Booster Vaccination | ||||
|---|---|---|---|---|---|---|---|
| Studies | Observations | Sera | Studies | Observations | Sera | ||
| mRNA | BA.1.1 | 3 | 4 | 55 | 6 | 6 | 100 |
| BA.2 | 16 | 18 | 282 | 28 | 29 | 520 | |
| BA.2.12.1 | 5 | 6 | 124 | 10 | 10 | 195 | |
| BA.2.75 | 2 | 2 | 35 | 3 | 4 | 74 | |
| BA.3 | 2 | 2 | 20 | 6 | 6 | 97 | |
| BA.4/5 | 7 | 8 | 164 | 17 | 18 | 751 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Inactivated | BA.1.1 | 1 | 1 | 10 | 3 | 3 | 76 |
| BA.2 | 4 | 4 | 59 | 9 | 9 | 213 | |
| BA.2.12.1 | 2 | 2 | 37 | 3 | 3 | 85 | |
| BA.2.75 | 0 | 0 | 0 | 1 | 1 | 40 | |
| BA.3 | 1 | 1 | 10 | 3 | 3 | 76 | |
| BA.4/5 | 3 | 3 | 49 | 4 | 4 | 97 | |
| BA.4.6 | 0 | 0 | 0 | 1 | 1 | 40 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Vector | BA.1.1 | 0 | 0 | 0 | 2 | 2 | 82 |
| BA.2 | 2 | 3 | 37 | 4 | 4 | 161 | |
| BA.2.12.1 | 2 | 3 | 37 | 0 | 0 | 0 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 1 | 1 | 41 | |
| BA.4/5 | 2 | 3 | 37 | 2 | 2 | 49 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Protein Subunit | BA.1.1 | 0 | 0 | 0 | 1 | 1 | 20 |
| BA.2 | 1 | 1 | 10 | 1 | 1 | 20 | |
| BA.2.12.1 | 1 | 1 | 10 | 0 | 0 | 0 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 1 | 1 | 20 | |
| BA.4/5 | 2 | 2 | 39 | 1 | 1 | 48 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Heterologous (+mRNA) | BA.1.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| BA.2 | 0 | 0 | 0 | 8 | 14 | 355 | |
| BA.2.12.1 | 0 | 0 | 0 | 2 | 3 | 56 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 1 | 2 | 30 | |
| BA.4/5 | 0 | 0 | 0 | 2 | 3 | 56 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Heterologous (-mRNA) | BA.1.1 | 0 | 0 | 0 | 3 | 3 | 76 |
| BA.2 | 0 | 0 | 0 | 6 | 6 | 156 | |
| BA.2.12.1 | 0 | 0 | 0 | 2 | 2 | 68 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 3 | 3 | 76 | |
| BA.4/5 | 0 | 0 | 0 | 2 | 2 | 68 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Unspecified | BA.1.1 | 0 | 0 | 0 | 0 | 0 | 0 |
| BA.2 | 0 | 0 | 0 | 2 | 4 | 79 | |
| BA.2.12.1 | 0 | 0 | 0 | 1 | 3 | 35 | |
| BA.2.75 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.3 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4/5 | 0 | 0 | 0 | 1 | 3 | 35 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.4.7 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Total | BA.1.1 | 4 | 5 | 65 | 15 | 15 | 354 |
| BA.2 | 23 | 26 | 388 | 58 | 67 | 1504 | |
| BA.2.12.1 | 10 | 12 | 208 | 18 | 21 | 439 | |
| BA.2.75 | 2 | 2 | 35 | 4 | 5 | 114 | |
| BA.3 | 3 | 3 | 30 | 15 | 16 | 340 | |
| BA.4/5 | 14 | 16 | 289 | 29 | 33 | 1104 | |
| BA.4.6 | 0 | 0 | 0 | 1 | 1 | 40 | |
| BA.4.6 | 0 | 0 | 0 | 0 | 0 | 0 | |
| BA.5.9 | 0 | 0 | 0 | 0 | 0 | 0 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jacobsen, H.; Katzmarzyk, M.; Higdon, M.M.; Jiménez, V.C.; Sitaras, I.; Bar-Zeev, N.; Knoll, M.D. Post-Vaccination Neutralization Responses to Omicron Sub-Variants. Vaccines 2022, 10, 1757. https://doi.org/10.3390/vaccines10101757
Jacobsen H, Katzmarzyk M, Higdon MM, Jiménez VC, Sitaras I, Bar-Zeev N, Knoll MD. Post-Vaccination Neutralization Responses to Omicron Sub-Variants. Vaccines. 2022; 10(10):1757. https://doi.org/10.3390/vaccines10101757
Chicago/Turabian StyleJacobsen, Henning, Maeva Katzmarzyk, Melissa M. Higdon, Viviana Cobos Jiménez, Ioannis Sitaras, Naor Bar-Zeev, and Maria Deloria Knoll. 2022. "Post-Vaccination Neutralization Responses to Omicron Sub-Variants" Vaccines 10, no. 10: 1757. https://doi.org/10.3390/vaccines10101757