
Immunoprofiling Identifies Functional B and T Cell Subsets Induced by an Attenuated Whole Parasite Malaria Vaccine as Correlates of Sterile Immunity

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Enzyme-Linked Immunosorbent Assay (ELISA)
2.3. Immunofluorescence Antibody Assay (IFA) Using SPZ
2.4. Electro-Chemiluminescence-Based Multiplex Assay (MSD) for Antibody Detection
2.5. B Cell ELISpot
2.6. Flow Cytometric Analysis
2.7. MSD for Cytokine Detection
2.8. Statistical Analysis
2.9. Univariate Analysis
2.10. Multivariate Analysis and Machine Learning
3. Results
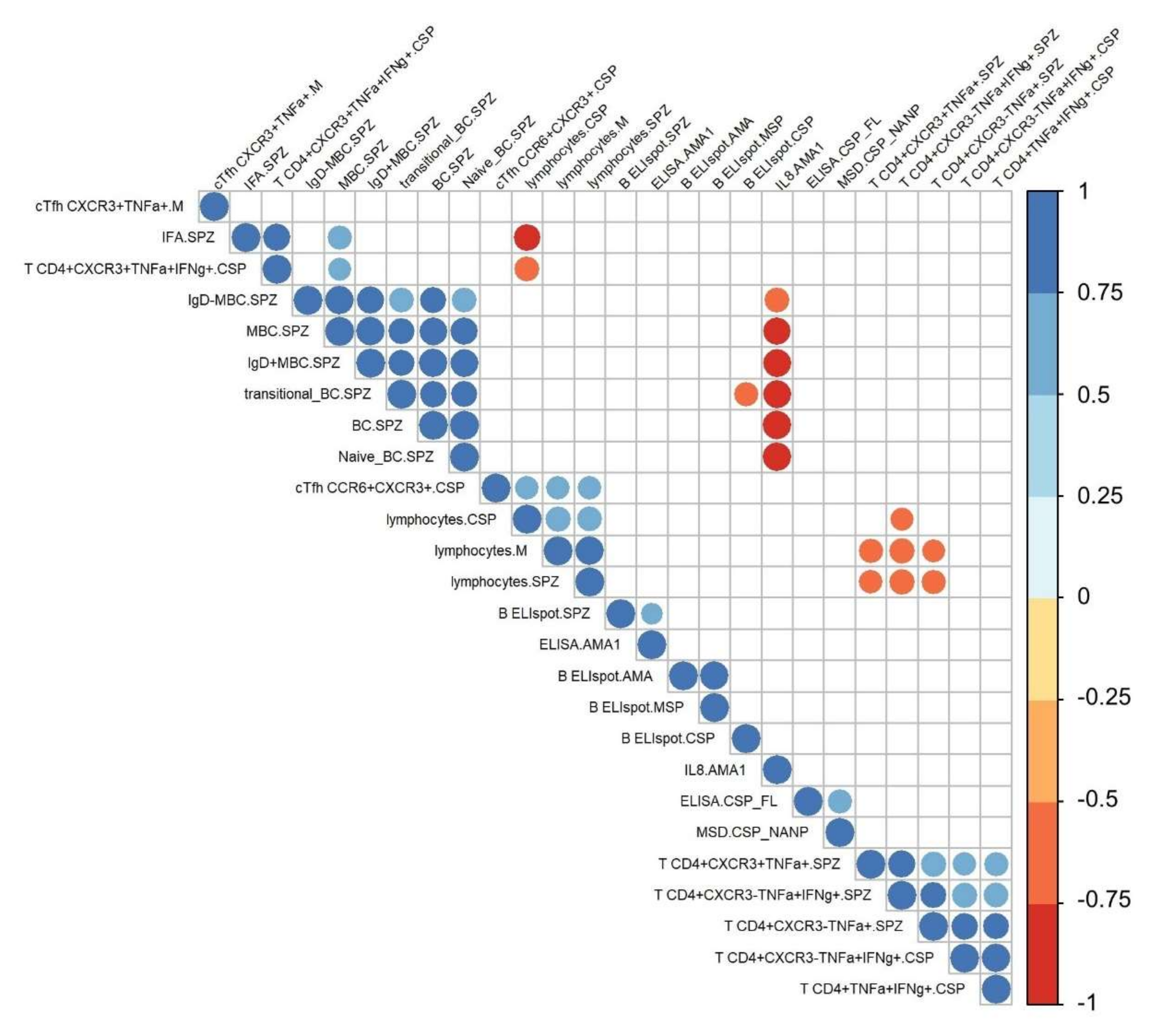
3.1. Immunoprofiling of IMRAS-Induced Humoral and Cellular Immune Response
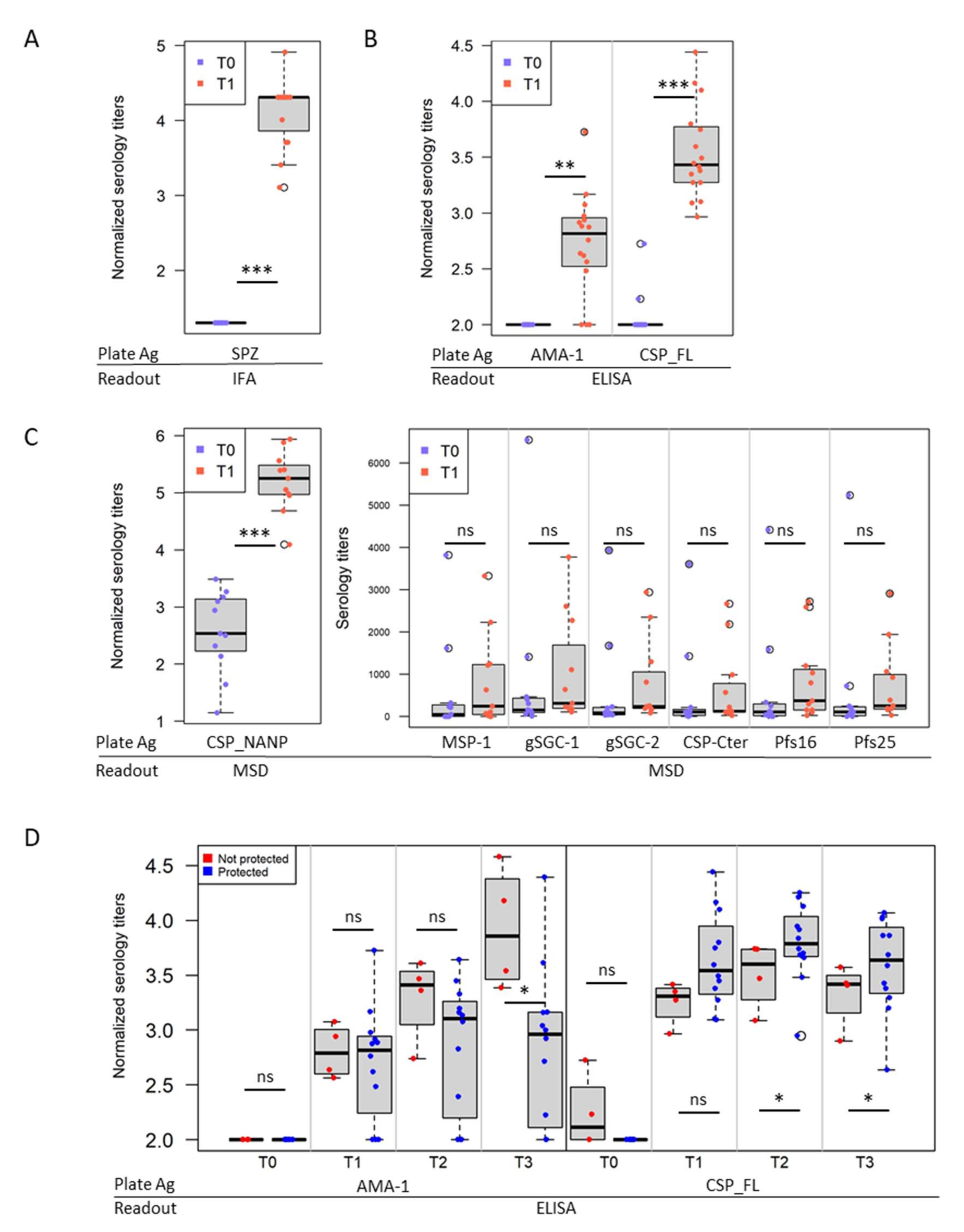
3.2. IMRAS Humoral Response Is Mainly Driven by CSP
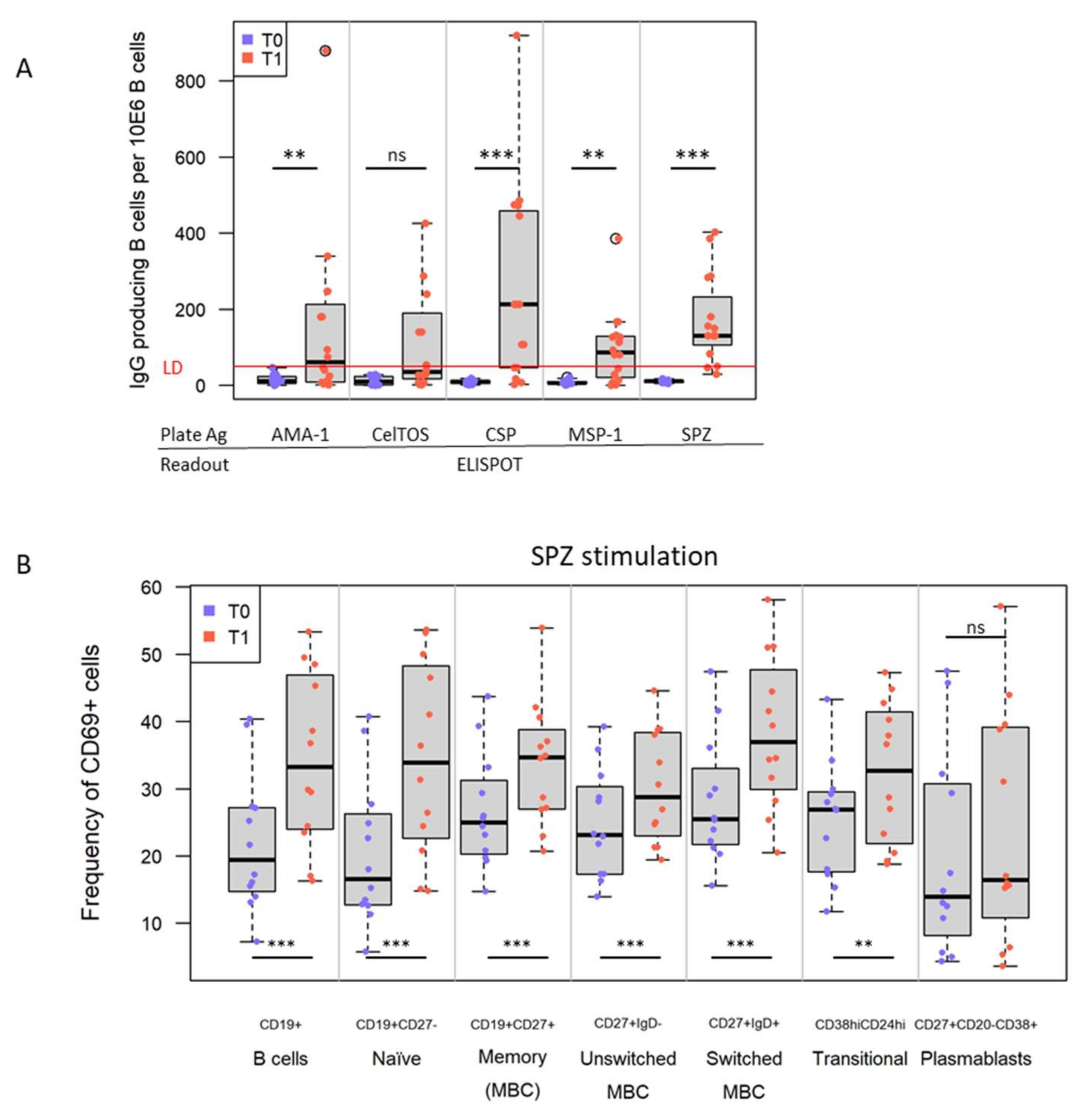
3.3. IMRAS Vaccination Leads to Significant Expansion of B Cells Specific for a Range of Plasmodial Antigens
3.4. IMRAS Induces Significant Levels of CSP-Specific Circulating Follicular T Helper Cells
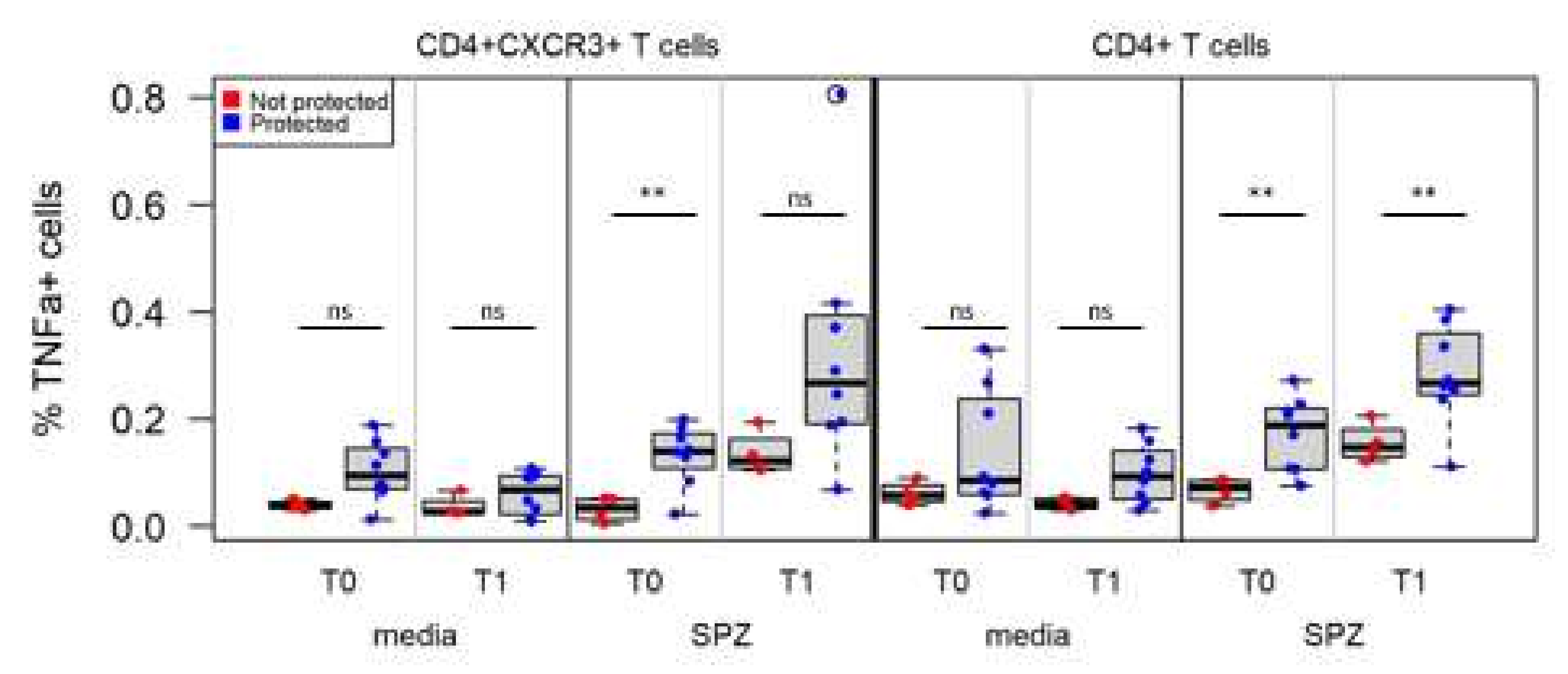
3.5. IMRAS Circulating Cellular Response Is Driven by IFNγ- and/or TNFα-Secreting CD4+ T Cells
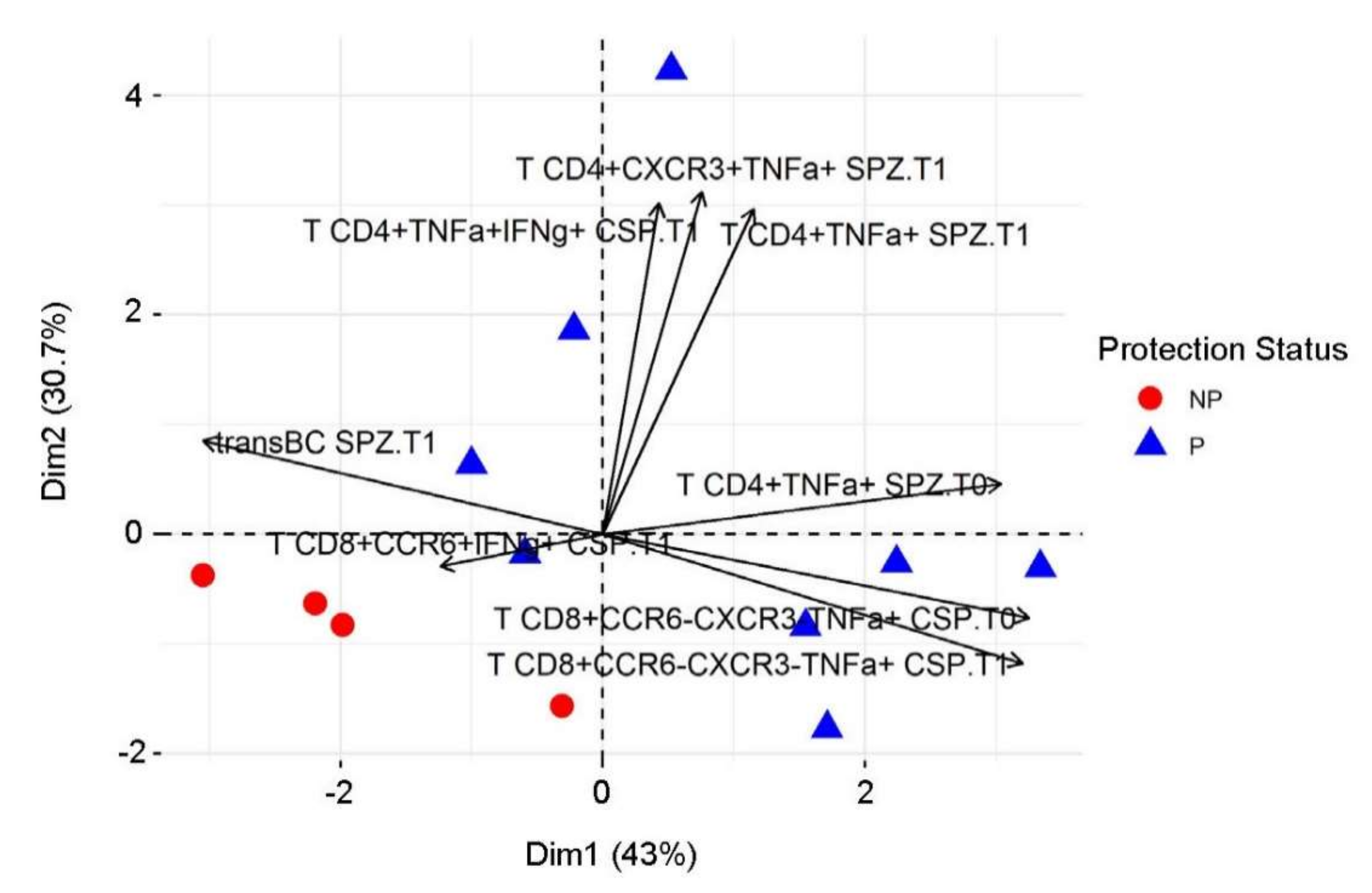
3.6. Associations between Immune Measures and Protection Using Univariate Analysis
3.7. Predictive Immune Model Using Machine Learning
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Disclaimer
Abbreviations
References
- Plotkin, S.A. Correlates of Protection Induced by Vaccination. Clin. Vaccine Immunol. 2010, 17, 1055–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farooq, F.; Bergmann-Leitner, E.S. Immune Escape Mechanisms are Plasmodium’ s Secret Weapons Foiling the Success of Potent and Persistently Efficacious Malaria Vaccines. Clin. Immunol. 2015, 161, 136–143. [Google Scholar] [CrossRef]
- World Health Organization. World Malaria Report; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- The RTS,S Clinical Trials Partnership. Efficacy and Safety of the RTS,S/AS01 Malaria Vaccine during 18 Months after Vaccination: A Phase 3 Randomized, Controlled Trial in Children and Young Infants at 11 African Sites. PLoS Med. 2014, 11, e1001685. [Google Scholar] [CrossRef] [Green Version]
- The RTS,S Clinical Trials Partnership; Agnandji, S.T.; Lell, B.; Fernandes, J.F.; Abossolo, B.P.; Methogo, B.G.; Kabwende, A.L.; Adegnika, A.; Mordmuller, B.; Issifou, S.; et al. A Phase 3 Trial of RTS,S/AS01 Malaria Vaccine in African Infants. N. Engl. J. Med. 2012, 367, 2284–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Datoo, M.S.; Natama, M.H.; Somé, A.; Traoré, O.; Rouamba, T.; Bellamy, D.; Yameogo, P.; Valia, D.; Tegneri, M.; Ouedraogo, F.; et al. Efficacy of a low-dose candidate malaria vaccine, R21 in adjuvant Matrix-M, with seasonal administration to children in Burkina Faso: A randomised controlled trial. Lancet 2021, 397, 1809–1818. [Google Scholar] [CrossRef]
- Duffy, P.E.; Gorres, J.P. Malaria vaccines since 2000: Progress, priorities, products. npj Vaccines 2020, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hoffman, S.L.; Billingsley, P.F.; James, E.; Richman, A.; Loyevsky, M.; Li, T.; Chakravarty, S.; Gunasekera, A.; Chattopadhyay, R.; Li, M.; et al. Development of a metabolically active, non-replicating sporozoite vaccine to prevent Plasmodium falciparum malaria. Hum. Vaccines 2010, 6, 97–106. [Google Scholar] [CrossRef] [Green Version]
- Lyke, K.E.; Ishizuka, A.S.; Berry, A.A.; Chakravarty, S.; DeZure, A.; Enama, M.E.; James, E.R.; Billingsley, P.F.; Gunasekera, A.; Manoj, A.; et al. Attenuated PfSPZ Vaccine induces strain-transcending T cells and durable protection against heterologous controlled human malaria infection. Proc. Natl. Acad. Sci. USA 2017, 114, 2711–2716. [Google Scholar] [CrossRef] [Green Version]
- Sissoko, M.S.; Healy, S.; Katile, A.; Omaswa, F.; Zaidi, I.; Gabriel, E.; Kamate, B.; Samake, Y.; Guindo, M.; Dolo, A.; et al. Safety and efficacy of PfSPZ Vaccine against Plasmodium falciparum via direct venous inoculation in healthy malaria-exposed adults in Mali: A randomised, double-blind phase 1 trial. Lancet Infect. Dis. 2017, 17, 498–509. [Google Scholar] [CrossRef]
- Spring, M.; Murphy, J.; Nielsen, R.; Dowler, M.; Bennett, J.W.; Zarling, S.; Williams, J.; de la Vega, P.; Ware, L.; Komisar, J.; et al. First-in-human evaluation of genetically attenuated Plasmodium falciparum sporozoites administered by bite of Anopheles mosquitoes to adult volunteers. Vaccine 2013, 31, 4975–4983. [Google Scholar] [CrossRef]
- Clyde, D.F.; McCarthy, V.C.; Miller, R.M.; Hornick, R.B. Specificity of protection of man immunized against sporozoite-induced falciparum malaria. Am. J. Med. Sci. 1973, 266, 398–404. [Google Scholar] [CrossRef]
- Nussenzweig, R.S.; Vanderberg, J.; Most, H.; Orton, C. Protective Immunity produced by the Injection of X-irradiated Sporozoites of Plasmodium berghei. Nature 1967, 216, 160–162. [Google Scholar] [CrossRef]
- Rieckmann, K.H.; Beaudoin, R.L.; Cassells, J.S.; Sell, K.W. Use of attenuated sporozoites in the immunization of human volunteers against falciparum malaria. Bull. World Health Organ. 1979, 57 (Suppl. S1), 261–265. [Google Scholar]
- Goh, Y.S.; McGuire, D.; Rénia, L. Vaccination with Sporozoites: Models and Correlates of Protection. Front. Immunol. 2019, 10, 1227. [Google Scholar] [CrossRef]
- Hoffman, S.L.; Goh, L.M.L.; Luke, T.C.; Schneider, I.; Le, T.P.; Doolan, D.; Sacci, J.; de la Vega, P.; Dowler, M.; Paul, C.; et al. Protection of Humans against Malaria by Immunization with Radiation-Attenuated Plasmodium falciparum Sporozoites. J. Infect. Dis. 2002, 185, 1155–1164. [Google Scholar] [CrossRef] [Green Version]
- Nkumama, I.N.; Osier, F.H.A. Malaria vaccine roller coaster. Nat. Microbiol. 2021, 6, 1345–1346. [Google Scholar] [CrossRef]
- Bergmann-Leitner, E.S.; Mease, R.M.; de la Vega, P.; Savranskaya, T.; Polhemus, M.; Ockenhouse, C.; Angov, E. Immunization with Pre-Erythrocytic Antigen CelTOS from Plasmodium falciparum Elicits Cross-Species Protection against Heterologous Challenge with Plasmodium berghei. PLoS ONE 2010, 5, e12294. [Google Scholar] [CrossRef] [Green Version]
- Doolan, D.; Southwood, S.; Freilich, D.A.; Sidney, J.; Graber, N.L.; Shatney, L.; Bebris, L.; Florens, L.; Dobano, C.; Witney, A.; et al. Identification of Plasmodium falciparum antigens by antigenic analysis of genomic and proteomic data. Proc. Natl. Acad. Sci. USA 2003, 100, 9952–9957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trieu, A.; Kayala, M.A.; Burk, C.; Molina, D.M.; Freilich, D.A.; Richie, T.; Baldi, P.; Felgner, P.L.; Doolan, D.L. Sterile Protective Immunity to Malaria is Associated with a Panel of Novel, P. falciparum Antigens. Mol. Cell. Proteom. 2011, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doll, K.L.; Harty, J.T. Correlates of protective immunity following whole sporozoite vaccination against malaria. Immunol. Res. 2014, 59, 166–176. [Google Scholar] [CrossRef] [PubMed]
- Atre, T.; Robinson, T.M.; Savransky, T.; Dutta, S.; Epstein, J.E.; Bergmann-Leitner, E.S. Novel sporozoite-based ELISpot assay to assess frequency of parasite-specific B cells after vaccination with irradiated sporozoites. Malar. J. 2019, 18, 186. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, E.S.; Ballou, R.W.; Krzych, U. Detection of CD4+CD45RO+T Lymphocytes Producing IL-4 in Response to Antigens on Plasmodium falciparum Erythrocytes: An in Vitro Correlate of Protective Immunity Induced with Attenuated Plasmodium falciparum Sporozoites. Cell. Immunol. 1997, 180, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Doll, K.L.; Pewe, L.L.; Kurup, S.P.; Harty, J.T. Discriminating Protective from Nonprotective Plasmodium-Specific CD8+ T Cell Responses. J. Immunol. 2016, 196, 4253–4262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Doolan, D.; Hoffman, S.L. Pre–erythrocytic–stage immune effector mechanisms in Plasmodium spp. infections. Philos. Trans. R. Soc. B Biol. Sci. 1997, 352, 1361–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hickey, B.; Teneza-Mora, N.; Lumsden, J.; Reyes, S.; Sedegah, M.; Garver, L.; Hollingdale, M.R.; Banania, J.G.; Ganeshan, H.; Dowler, M.; et al. IMRAS—A clinical trial of mosquito-bite immunization with live, radiation-attenuated P. falciparum sporozoites: Impact of immunization parameters on protective efficacy and generation of a repository of immunologic reagents. PLoS ONE 2020, 15, e0233840. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S.; Duncan, E.H.; Atre, T.; Dutta, S.; Spring, M.D.; Leitner, W.W.; Bergmann-Leitner, E.S. Combining immunoprofiling with machine learning to assess the effects of adjuvant formulation on human vaccine-induced immunity. Hum. Vaccines Immunother. 2020, 16, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, S.; Duncan, E.H.; Atre, T.; Storme, C.K.; Beck, K.; Kaba, S.A.; Lanar, D.E.; Bergmann-Leitner, E.S. Identification of Immune Signatures of Novel Adjuvant Formulations Using Machine Learning. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Pallikkuth, S.; Chaudhury, S.; Lu, P.; Pan, L.; Jongert, E.; Wille-Reece, U.; Pahwa, S. A delayed fractionated dose RTS,S AS01 vaccine regimen mediates protection via improved T follicular helper and B cell responses. Elife 2020, 9, e51889. [Google Scholar] [CrossRef]
- Ali, Z.M.; Bakli, M.; Fontaine, A.; Bakkali, N.; Hai, V.V.; Audebert, S.; Boublik, Y.; Pagès, F.; Remoué, F.; Rogier, C.; et al. Assessment of Anopheles salivary antigens as individual exposure biomarkers to species-specific malaria vector bites. Malar. J. 2012, 11, 439. [Google Scholar] [CrossRef] [Green Version]
- Bolton, J.S.; Chaudhury, S.; Dutta, S.; Gregory, S.; Locke, E.; Pierson, T.; Bergmann-Leitner, E.S. Comparison of ELISA with electro-chemiluminescence technology for the qualitative and quantitative assessment of serological responses to vaccination. Malar. J. 2020, 19, 1–13. [Google Scholar] [CrossRef]
- R Core Team. A language and environment for statistical computing. In R Foundation for Statistical Computing; R Core Team: Vienna, Austria, 2013. [Google Scholar]
- Morita, R.; Schmitt, N.; Bentebibel, S.-E.; Ranganathan, R.; Bourdery, L.; Zurawski, G.; Foucat, E.; Dullaers, M.; Oh, S.; Sabzghabaei, N.; et al. Human Blood CXCR5+CD4+ T Cells Are Counterparts of T Follicular Cells and Contain Specific Subsets that Differentially Support Antibody Secretion. J. Immun. 2011, 34, 108–121. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, A.R.; Engelhard, V. CD8 T Cells Activated in Distinct Lymphoid Organs Differentially Express Adhesion Proteins and Coexpress Multiple Chemokine Receptors. J. Immunol. 2010, 184, 4079–4086. [Google Scholar] [CrossRef] [Green Version]
- Oo, Y.H.; Weston, C.J.; Lalor, P.F.; Curbishley, S.M.; Withers, D.R.; Reynolds, G.M.; Shetty, S.; Harki, J.; Shaw, J.C.; Eksteen, B.; et al. Distinct Roles for CCR4 and CXCR3 in the Recruitment and Positioning of Regulatory T Cells in the Inflamed Human Liver. J. Immunol. 2010, 184, 2886–2898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simms, P.E.; Ellis, T.M. Utility of flow cytometric detection of CD69 expression as a rapid method for determining poly- and oligoclonal lymphocyte activation. Clin. Diagn. Lab. Immunol. 1996, 3, 301–304. [Google Scholar] [CrossRef] [Green Version]
- Tough, D.F.; Sun, S.; Sprent, J. T cell stimulation in vivo by lipopolysaccharide (LPS). J. Exp. Med. 1997, 185, 2089–2094. [Google Scholar] [CrossRef] [Green Version]
- Farooq, F.; Beck, K.; Paolino, K.M.; Phillips, R.; Waters, N.C.; Regules, J.A.; Bergmann-Leitner, E.S. Circulating follicular T helper cells and cytokine profile in humans following vaccination with the rVSV-ZEBOV Ebola vaccine. Sci. Rep. 2016, 6, 27944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmitt, N.; Bentebibel, S.E.; Ueno, H. Phenotype and functions of memory Tfh cells in human blood. Trends Immunol. 2014, 35, 436–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ueno, H. Human Circulating T Follicular Helper Cell Subsets in Health and Disease. J. Clin. Immunol. 2016, 36, 34–39. [Google Scholar] [CrossRef]
- Matsui, K.; Adelsberger, J.W.; Kemp, T.J.; Baseler, M.W.; Ledgerwood, J.E.; Pinto, L.A. Circulating CXCR5+CD4+ T Follicular-Like Helper Cell and Memory B Cell Responses to Human Papillomavirus Vaccines. PLoS ONE 2015, 10, e0137195. [Google Scholar] [CrossRef] [Green Version]
- Feng, J.; Gulati, U.; Zhang, X.; Keitel, W.A.; Thompson, D.M.; James, J.A.; Thompson, L.F.; Air, G.M. Antibody quantity versus quality after influenza vaccination. Vaccine 2009, 27, 6358–6362. [Google Scholar] [CrossRef] [Green Version]
- Röltgen, K.; Powell, A.E.; Wirz, O.F.; Stevens, B.A.; Hogan, C.A.; Najeeb, J.; Hunter, M.; Wang, H.; Sahoo, M.K.; Huang, C.; et al. Defining the features and duration of antibody responses to SARS-CoV-2 infection associated with disease severity and outcome. Sci. Immunol. 2020, 5, eabe0240. [Google Scholar] [CrossRef]
- Leitner, W.W.; Haraway, M.; Pierson, T.; Bergmann-Leitner, E.S. Role of Opsonophagocytosis in Immune Protection against Malaria. Vaccines 2020, 8, 264. [Google Scholar] [CrossRef]
- Grüner, A.C.; Mauduit, M.; Tewari, R.; Romero, J.F.; Depinay, N.; Kayibanda, M.; Lallemand, E.; Chavatte, J.M.; Crisanti, A.; Sinnis, P.; et al. Sterile protection against malaria is independent of immune responses to the circumsporozoite protein. PLoS ONE. 2007, 2, e1371. [Google Scholar] [CrossRef] [Green Version]
- Locci, M.; Havenar-Daughton, C.; Landais, E.; Wu, J.; Kroenke, M.A.; Arlehamn, C.L.; Su, L.F.; Cubas, R.; Davis, M.M.; Sette, A.; et al. Human circulating PD-1+CXCR3-CXCR5+ memory Tfh cells are highly functional and correlate with broadly neutralizing HIV antibody responses. Immunity 2013, 39, 758–769. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, M.T.; Bejon, P.; Olotu, A.; Griffin, J.T.; Bojang, K.; Lusingu, J.; Salim, N.; Abdulla, S.; Otsyula, N.; Agnandji, S.T.; et al. A combined analysis of immunogenicity, antibody kinetics and vaccine efficacy from phase 2 trials of the RTS, S malaria vaccine. BMC Med. 2014, 12, 117. [Google Scholar] [CrossRef]
- Currier, J.; Sattabongkot, J.; Good, M.F. ‘Natural’ T cells responsive to malaria: Evidence implicating immunological cross-reactivity in the maintenance of TCR alpha beta+ malaria-specific responses from non-exposed donors. Int. Immunol. 1992, 4, 985–994. [Google Scholar] [CrossRef] [PubMed]
- Obeng-Adjei, N.; Portugal, S.; Tran, T.M.; Yazew, T.B.; Skinner, J.; Li, S.; Jain, A.; Felgner, P.L.; Doumbo, O.K.; Kayentao, K.; et al. Circulating Th1-Cell-type Tfh Cells that Exhibit Impaired B Cell Help Are Preferentially Activated during Acute Malaria in Children. Cell Rep. 2015, 13, 425–439. [Google Scholar] [CrossRef] [Green Version]
- Groom, J.R.; Luster, A.D. CXCR3 in T cell function. Exp. Cell Res. 2011, 317, 620–631. [Google Scholar] [CrossRef] [PubMed]
- Kuo, P.T.; Zeng, Z.; Salim, N.; Mattarollo, S.; Wells, J.W.; Leggatt, G.R. The Role of CXCR3 and Its Chemokine Ligands in Skin Disease and Cancer. Front. Med. 2018, 5, 271. [Google Scholar] [CrossRef]
- Thomas, S.Y.; Banerji, A.; Medoff, B.D.; Lilly, C.M.; Luster, A.D. Multiple chemokine receptors, including CCR6 and CXCR3, regu-late antigen-induced T cell homing to the human asthmatic airway. J. Immunol. 2007, 179, 1901–1912. [Google Scholar] [CrossRef]
- Curbishley, S.M.; Eksteen, B.; Gladue, R.P.; Lalor, P.; Adams, D.H. CXCR 3 activation promotes lymphocyte transendothelial mi-gration across human hepatic endothelium under fluid flow. Am. J. Pathol. 2005, 167, 887–899. [Google Scholar] [CrossRef] [Green Version]
- Oo, Y.H.; Banz, V.; Kavanagh, D.; Liaskou, E.; Withers, D.; Humphreys, E.; Reynolds, G.M.; Lee-Turner, L.; Kalia, N.; Hubscher, S.G.; et al. CXCR3-dependent recruitment and CCR6-mediated positioning of Th-17 cells in the inflamed liver. J. Hepatol. 2012, 57, 1044–1051. [Google Scholar] [CrossRef] [Green Version]
- Schofield, L.; Villaquiran, J.; Ferreira, A.; Schellekens, H.; Nussenzweig, R.; Nussenzweig, V. γ Interferon, CD8+ T cells and antibodies required for immunity to malaria sporozoites. Nature 1987, 330, 664–666. [Google Scholar] [CrossRef]
- Weiss, W.R.; Jiang, C.G. Protective CD8+ T lymphocytes in Primates Immunized with Malaria Sporozoites. PLoS ONE 2012, 7, e31247. [Google Scholar] [CrossRef]
- Weiss, W.R.; Sedegah, M.; Beaudoin, R.L.; Miller, L.H.; Good, M.F. CD8+ T cells (cytotoxic/suppressors) are required for protection in mice immunized with malaria sporozoites. Proc. Natl. Acad. Sci. USA 1988, 85, 573–576. [Google Scholar] [CrossRef] [Green Version]
- Cockburn, I.A.; Amino, R.; Kelemen, R.K.; Kuo, S.C.; Tse, S.W.; Radtke, A.; Mac-Daniel, L.; Ganusov, V.V.; Zavala, F.; Ménard, R. In vivo imaging of CD8+ T cell-mediated elimina-tion of malaria liver stages. Proc. Natl. Acad. Sci. USA 2013, 110, 9090–9095. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, G.A.; Kumar, K.A.; Calvo-Calle, J.M.; Othoro, C.; Altszuler, D.; Nussenzweig, V.; Nardin, E.H. Class II-Restricted Protective Immunity Induced by Malaria Sporozoites. Infect. Immun. 2008, 76, 1200–1206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodrigues, M.; Nussenzweig, R.S.; Romero, P.; Zavala, F. The in vivo cytotoxic activity of CD8+ T cell clones correlates with their levels of expression of adhesion molecules. J. Exp. Med. 1992, 175, 895–905. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.; Nussenzweig, R.S.; Zavala, F. The relative contribution of antibodies, CD4+ and CD8+ T cells to sporozo-ite-induced protection against malaria. Immunology 1993, 80, 1–5. [Google Scholar]
- Penha-Gonçalves, C. Genetics of Malaria Inflammatory Responses: A Pathogenesis Perspective. Front. Immunol. 2019, 10, 1771. [Google Scholar] [CrossRef] [Green Version]
- Depinay, N.; Franetich, J.F.; Grüner, A.C.; Mauduit, M.; Chavatte, J.-M.; Luty, A.J.F.; van Gemert, G.-J.; Sauerwein, R.W.; Siksik, J.-M.; Hannoun, L.; et al. Inhibitory Effect of TNF-α on Malaria Pre-Erythrocytic Stage Development: Influence of Host Hepatocyte/Parasite Combinations. PLoS ONE 2011, 6, e17464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blair, P.A.; Noreña, L.Y.; Flores-Borja, F.; Rawlings, D.J.; Isenberg, D.A.; Ehrenstein, M.R.; Mauri, C. CD19(+)CD24(hi)CD38(hi) B cells ex-hibit regulatory capacity in healthy individuals but are functionally impaired in systemic Lupus Erythematosus patients. Immunity 2010, 32, 129–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chakravarty, S.; Cockburn, I.; Kuk, S.; Overstreet, M.; Sacci, J.B.; Zavala, F. CD8+ T lymphocytes protective against malaria liver stages are primed in skin-draining lymph nodes. Nat. Med. 2007, 13, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Ruiz, D.F.; Ng, W.Y.; Holz, L.; Ma, J.; Zaid, A.; Wong, Y.C.; Lau, L.S.; Mollard, V.; Cozijnsen, A.; Collins, N.; et al. Liver-Resident Memory CD8 + T Cells Form a Front-Line Defense against Malaria Liver-Stage Infection. Immunity 2016, 45, 889–902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hokeness, K.L.; Deweerd, E.S.; Munks, M.W.; Lewis, C.A.; Gladue, R.P.; Salazar-Mather, T.P. CXCR3-dependent recruitment of anti-gen-specific T lymphocytes to the liver during murine cytomegalovirus infection. J. Virol. 2007, 81, 1241–1250. [Google Scholar] [CrossRef] [Green Version]
- Geissmann, F.; Cameron, T.O.; Sidobre, S.; Manlongat, N.; Kronenberg, M.; Briskin, M.J.; Dustin, M.L.; Littman, D.R. Intravascular Immune Surveillance by CXCR6+ NKT Cells Patrolling Liver Sinusoids. PLOS Biol. 2005, 3, e113. [Google Scholar] [CrossRef] [Green Version]
- Bertolino, P.; Schrage, A.; Bowen, D.G.; Klugewitz, K.; Ghani, S.; Eulenburg, K.; Holz, L.; Hogg, N.; McCaughan, G.W.; Hamann, A. Early intrahepatic antigen-specific retention of naïve CD8+ T cells is predominantly ICAM-1/LFA-1 dependent in mice. Hepatology 2005, 42, 1063–1071. [Google Scholar] [CrossRef]
- Shimizu, Y.; Minemura, M.; Murata, H.; Hirano, K.; Nakayama, Y.; Higuchi, K.; Watanabe, A.; Yasuyama, T.; Tsukada, K. Preferential accumulation of CD103+ T cells in human livers; its association with extrathymic T cells. J. Hepatol. 2003, 39, 918–924. [Google Scholar] [CrossRef]
- Homey, B.; Dieu-Nosjean, M.C.; Wiesenborn, A.; Massacrier, C.; Pin, J.J.; Oldham, E.; Catron, D.; Buchanan, M.E.; Müller, A.; Malefyt, R.D.; et al. Up-regulation of macrophage inflamma-tory protein-3 alpha/CCL20 and CC chemokine receptor 6 in psoriasis. J. Immunol. 2000, 164, 6621–6632. [Google Scholar] [CrossRef] [Green Version]
- Cockburn, I.A.; Seder, R.A. Malaria prevention: From immunological concepts to effective vaccines and protective antibodies. Nat. Immunol. 2018, 19, 1199–1211. [Google Scholar] [CrossRef]
- Hafalla, J.C.R.; Bauza, K.; Friesen, J.; Gonzalez-Aseguinolaza, G.; Hill, A.V.S.; Matuschewski, K. Identification of Targets of CD8+ T Cell Responses to Malaria Liver Stages by Genome-wide Epitope Profiling. PLOS Pathog. 2013, 9, e1003303. [Google Scholar] [CrossRef]
- Romero, P.; Maryanski, J.L.; Corradin, G.; Nussenzweig, R.S.; Nussenzweig, V.; Zavala, F. Cloned cytotoxic T cells recognize an epitope in the circumsporozoite protein and protect against malaria. Nature 1989, 341, 323–326. [Google Scholar] [CrossRef]
- Tsang, J.S.; Schwartzberg, P.L.; Kotliarov, Y.; Biancotto, A.; Xie, Z.; Germain, R.N.; Wang, E.; Olnes, M.J.; Narayanan, M.; Golding, H.; et al. Global Analyses of Human Immune Variation Reveal Baseline Predictors of Postvaccination Responses. Cell 2014, 157, 499–513. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Immune Parameter | Technique | Measurement | Ag-Specificity | Time-Point |
|---|---|---|---|---|
| Antibodies | ELISA | IgG titre | AMA-1, CSP-FL | T0, T1, T2, T3 |
| Mesoscale | IgG titre | CSP-NANP, CSP-Cterm (Pf16), MSP-1, Pept-2, Pept-4, AMA-1, Pfs16, Pfs25 | T0, T1 | |
| Immunofluorescence assay | IFA titre | SPZ | T0, T1 | |
| B cells | Elispot | Number of Ag-spec. IgG secreting B cells/106 B cells | AMA-1, CelTOS, CSP, MSP-1, SPZ | T0, T1 |
| Flow Cytometry | Frequency of activated B cells and subtypes (naïve, memory, memory switched, transitional, plasmablast) | SPZ | T0, T1, T2, T3 | |
| cTfh | Flow Cytometry | Frequency of activated cTfh and subtypes (cTfh1, cTfh2, cTfh17) | SPZ, CSP | T0, T1, T2, T3 |
| CD4/CD8 T cells | Flow Cytometry | Frequency of activated CD4 T cells and subtypes (CXCR3+, CCR6+, Treg) | SPZ, CSP | T0, T1, T2, T3 |
| Frequency of activated CD8 T cells and subtypes (CXCR3+, CCR6+) | SPZ, CSP | T0, T1, T2, T3 | ||
| Cytokines | Flow Cytometry ICS | Frequency of TNF-α and IFN-γ secreting CD4+ and CD8+ T cells | SPZ, CSP | T0, T1, T2, T3 |
| Mesoscale | pg/ml of IFN-γ, TNF-α, IL-1β, IL-10, IL-2, IL-4, IL-6, IL-8, IL-12p70, IL-13 produced by 106 cells | AMA-1, CelTOS, CSP, MSP-1, hSPZ, lSPZ, TRAP | T0, T1 |
| Immune Measure a | Stimulation | Time Point | Relative Weights |
|---|---|---|---|
| TNFα secreting CD4+ T cells | SPZ | T1 | 100 |
| TNFα secreting CD4+ T cells | SPZ | T0 | 79.3 |
| Transitional B cells CD19+CD38hiCD24hiCD69+ | SPZ | T1 | 60.6 |
| TNFα secreting CD8+CXCR3−CCR6− T cells | CSP | T0 | 55.9 |
| TNFα secreting CD8+CXCR3−CCR6− T cells | CSP | T1 | 47.2 |
| IFNγ secreting CD8+CCR6+ T cells | CSP | T1 | 18.3 |
| TNFα and IFNγ secreting CD4+CXCR3−CCR6− T cells | CSP | T1 | 11.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mura, M.; Lu, P.; Atre, T.; Bolton, J.S.; Duncan, E.H.; Chaudhury, S.; Bergmann-Leitner, E.S. Immunoprofiling Identifies Functional B and T Cell Subsets Induced by an Attenuated Whole Parasite Malaria Vaccine as Correlates of Sterile Immunity. Vaccines 2022, 10, 124. https://doi.org/10.3390/vaccines10010124
Mura M, Lu P, Atre T, Bolton JS, Duncan EH, Chaudhury S, Bergmann-Leitner ES. Immunoprofiling Identifies Functional B and T Cell Subsets Induced by an Attenuated Whole Parasite Malaria Vaccine as Correlates of Sterile Immunity. Vaccines. 2022; 10(1):124. https://doi.org/10.3390/vaccines10010124
Chicago/Turabian StyleMura, Marie, Pinyi Lu, Tanmaya Atre, Jessica S. Bolton, Elizabeth H. Duncan, Sidhartha Chaudhury, and Elke S. Bergmann-Leitner. 2022. "Immunoprofiling Identifies Functional B and T Cell Subsets Induced by an Attenuated Whole Parasite Malaria Vaccine as Correlates of Sterile Immunity" Vaccines 10, no. 1: 124. https://doi.org/10.3390/vaccines10010124