Common and Distinct Functional Brain Networks for Intuitive and Deliberate Decision Making

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Experimental Design
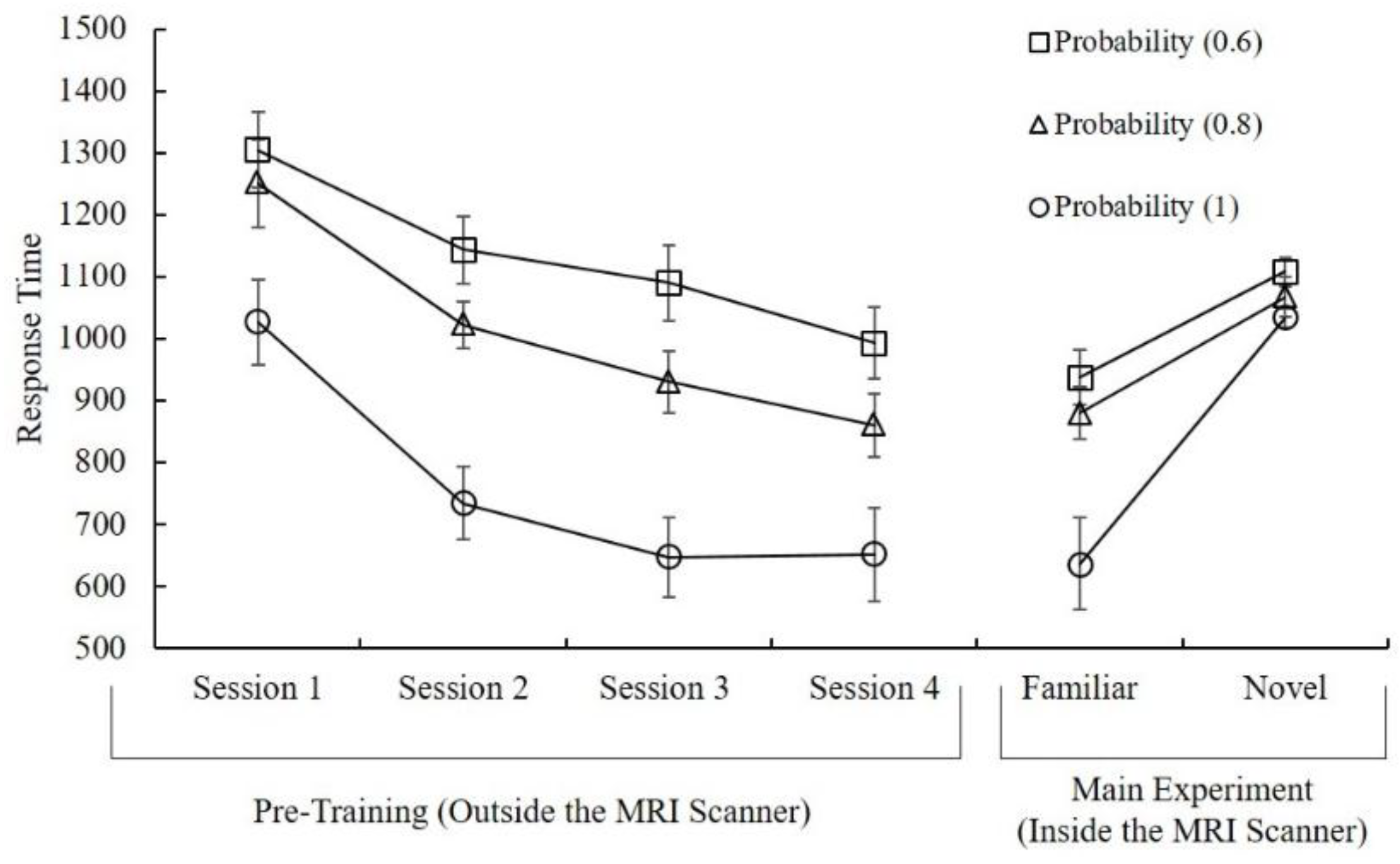
2.2.1. Pre-Training
2.2.2. Main Experiment
2.3. Functional Magnetic Resonance Imaging
2.3.1. Image Acquisition
2.3.2. Image Analysis
3. Results
3.1. Behavioral Results
3.2. Imaging Results
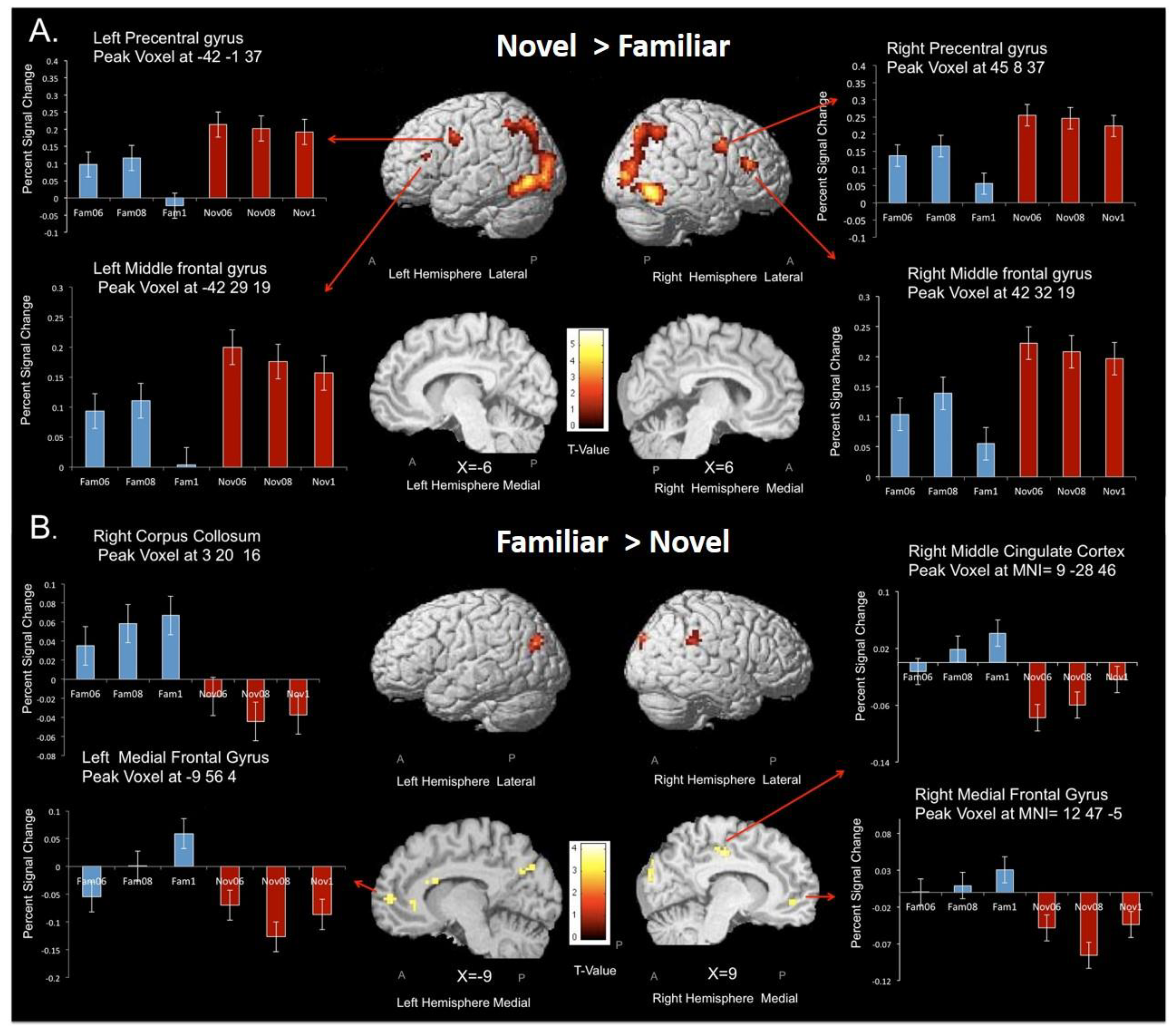
3.3. Subtraction Analyses—Novel vs. Familiar Stimuli
3.4. Psycho-Physiological Interaction Analysis
4. Discussion
5. Conclusions and Limitation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Balleine, B.W.; Delgado, M.R.; Hikosaka, O. The Role of the Dorsal Striatum in Reward and Decision-Making. J. Neurosci. 2007, 27, 8161–8165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kennerley, S.W.; Walton, M.E. Decision making and reward in frontal cortex: Complementary evidence from neurophysiological and neuropsychological studies. Behav. Neurosci. 2011, 125, 297–317. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Hairston, J.; Schrier, M.; Fan, J. Common and Distinct networks underlying reward valence and processing strategies: A meta-analysis of functional imaging studies. Neurosci. Biobehav. Rev. 2011, 35, 1219–1236. [Google Scholar] [CrossRef] [PubMed]
- Rangel, A.; Camerer, C.; Montague, P.R. A framework for studying the neurobiology of value-based decision making. Nat. Rev. Neurosci. 2008, 9, 545–556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolan, R.J.; Dayan, P. Goals and habits in the brain. Neuron. 2013, 80, 312–325. [Google Scholar] [CrossRef] [PubMed]
- Bodenhausen, G.V.; Todd, A.R. Automatic aspects of judgement and decision making. In Implicit Social Cognition: Measurement, Theory, and Applications; Gawronski, B., Keith Payne, B., Eds.; Guilford Press: Guilford, NY, USA, 2010; pp. 278–294. [Google Scholar]
- Kruglanski, A.W.; Gigerenzer, G. Intuitive and deliberate judgments are based on common principles. Psychol. Rev. 2011, 118, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Balleine, B.W.; Daw, N.D.; O’Doherty, J.P. Multiple Forms of Value Learning and the Function of Dopamine; Elsevier BV: Amsterdam, The Netherlands, 2009; pp. 367–387. [Google Scholar]
- Balleine, B.W.; O’Doherty, J.P. Human and rodent homologies in action control: Corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology 2010, 35, 48–69. [Google Scholar] [CrossRef]
- Glascher, J.; Hampton, A.N.; O’Doherty, J.P. Determining a role for ventromedial prefrontal cortex in encoding action-based value signals during reward related decision making. Cereb. Cortex 2009, 19, 483–495. [Google Scholar] [CrossRef]
- Logan, G.D. Towards and instance theory of automatization. Psych. Rev. 1988, 95, 492–527. [Google Scholar] [CrossRef]
- Johnson-Laird, P.N. Mental models and human reasoning. Proc. Natl. Acad. Sci. USA 2010, 107, 18243–18250. [Google Scholar] [CrossRef] [Green Version]
- Sutton, R.; Barto, A. Reinforcement Learning: An Introduction. IEEE Trans. Neural Netw. 1998, 9, 1054. [Google Scholar] [CrossRef]
- Dickinson, A.; Balleine, B. The role of learning in the operation of motivational systems. In Steven’s Handbook of Experimental Psychology: Learning/Motivation and Emotion, 3rd ed.; Gallistel, C.R., Ed.; Wiley: New York, NY, USA, 2000; pp. 497–534. [Google Scholar]
- Ostlund, S.B.; Balleine, B.W. Theories of Goal-directed behaviour theory. In Encylcopedia of Neuroscience; Squire, L.R., Ed.; Academic Press: Oxford, UK, 2009; pp. 943–949. [Google Scholar]
- Rangel, A.; Hare, T. Neural computations associated with goal-directed choice. Curr. Opin. Neurobiol. 2010, 20, 262–270. [Google Scholar] [CrossRef]
- Redgrave, P.; Rodriguez, M.; Smith, Y.; Rodriguez-Oroz, M.C.; Lehericy, S.; Bergman, H.; Obeso, J.A. Goal-directed and habitual control in the basal ganglia: Implications for Parkinson’s disease. Nat. Rev. Neurosci. 2010, 11, 760–772. [Google Scholar] [CrossRef]
- Wood, W.; Neal, D.T. A new look at habits and the habit-goal interface. Psychol. Rev. 2007, 114, 843–863. [Google Scholar] [CrossRef]
- Glockner, A.; Witteman, C. Beyond dual-process models: A categorization of processes underlying intuitive judgement and decision making. Think. Reason. 2010, 16, 1–25. [Google Scholar] [CrossRef]
- Kim, H.F.; Hikosaka, O. Parallel basal ganglia circuits for voluntary and automatic behaviour to reach rewards. Brain 2015, 138, 1776–1800. [Google Scholar] [CrossRef] [Green Version]
- Keren, G.; Schul, Y. Two Is Not Always Better Than One: A Critical Evaluation of Two-System Theories. Perspect. Psychol. Sci. 2009, 4, 533–550. [Google Scholar] [CrossRef]
- Saling, L.; Phillips, J.; Phillips, J. Automatic behaviour: Efficient not mindless. Brain Res. Bull. 2007, 73, 1–20. [Google Scholar] [CrossRef]
- Graybiel, A.M. Habits, Rituals, and the Evaluative Brain. Annu. Rev. Neurosci. 2008, 31, 359–387. [Google Scholar] [CrossRef] [Green Version]
- Hélie, S.; Cousineau, D. The cognitive neuroscience of automaticity: Behavioral and brain signatures. Cogn. Sci. 2011, 6, 35. [Google Scholar]
- O’Doherty, J.; Dayan, P.; Schultz, J.; Deichmann, R.; Friston, K.; Dolan, R.J. Dissociable Roles of Ventral and Dorsal Striatum in Instrumental Conditioning. Science 2004, 304, 452–454. [Google Scholar] [CrossRef] [Green Version]
- Rolls, E.T.; McCabe, C.; Redoute, J. Expected value, reward outcome, and temporal difference error representations in a probabilistic decision task. Cereb. Cortex 2008, 18, 652–663. [Google Scholar] [CrossRef]
- Schoenbaum, G.; Roesch, M. Orbitofrontal cortex, associative learning, and expectancies. Neuron. 2005, 47, 633–636. [Google Scholar] [CrossRef]
- Tobler, P.N.; O’Doherty, J.P.; Dolan, R.J.; Schultz, W. Reward Value Coding Distinct from Risk Attitude-Related Uncertainty Coding in Human Reward Systems. J. Neurophysiol. 2007, 97, 1621–1632. [Google Scholar] [CrossRef]
- Delgado, M.; Miller, M.; Inati, S.; Phelps, E. An fMRI study of reward-related probability learning. NeuroImage 2005, 24, 862–873. [Google Scholar] [CrossRef]
- Haruno, M.; Kawato, M. Heterarchical reinforcement-learning model for integration of multiple cortico-striatal loops: fMRI examination in stimulus-action-reward association learning. Neural Netw. 2006, 19, 1242–1254. [Google Scholar] [CrossRef]
- Kahnt, T.; Heinzle, J.; Park, S.Q.; Haynes, J.-D. Decoding the Formation of Reward Predictions across Learning. J. Neurosci. 2011, 31, 14624–14630. [Google Scholar] [CrossRef] [Green Version]
- Tricomi, E.; Balleine, B.W.; O’Doherty, J.P. A specific role for posterior dorsolateral striatum in human habit learning. Eur. J. Neurosci. 2009, 29, 2225–2232. [Google Scholar] [CrossRef] [Green Version]
- Wunderlich, K.; Dayan, P.; Dolan, R.J. Mapping value based planning and extensively trained choice in the human brain. Nat. Neurosci. 2012, 15, 786–791. [Google Scholar] [CrossRef] [Green Version]
- Ashby, F.G.; Crossley, M.J. Automaticity and multiple memory systems. Wiley Interdiscip. Rev. Cogn. Sci. 2012, 3, 363–376. [Google Scholar] [CrossRef]
- Isoda, M.; Hikosaka, O. Cortico-basal ganglia mechanisms for overcoming innate, habitual and motivational behaviors. Eur. J. Neurosci. 2011, 33, 2058–2069. [Google Scholar] [CrossRef]
- Ashby, F.G.; Turner, B.O.; Horvitz, J.C. Cortical and basal ganglia contributions to habit learning and automaticity. Trends Cogn. Sci. 2010, 14, 208–215. [Google Scholar] [CrossRef] [Green Version]
- Preuschoff, K.; Bossaerts, P.; Quartz, S.R. Neural Differentiation of Expected Reward and Risk in Human Subcortical Structures. Neuron 2006, 51, 381–390. [Google Scholar] [CrossRef] [Green Version]
- Daw, N.D.; Niv, Y.; Dayan, P. Uncertainty-based competition between prefrontal and dorsolateral striatal systems for behavioural control. Nat. Neurosci. 2005, 8, 1704–1711. [Google Scholar] [CrossRef]
- Thorn, C.A.; Atallah, H.; Howe, M.; Graybiel, A.M. Differential Dynamics of Activity Changes in Dorsolateral and Dorsomedial Striatal Loops During Learning. Neuron 2010, 66, 781–795. [Google Scholar] [CrossRef]
- Seger, C.A.; Spiering, B.J. A Critical Review of Habit Learning and the Basal Ganglia. Front. Syst. Neurosci. 2011, 5, 66. [Google Scholar] [CrossRef] [Green Version]
- Jansma, J.M.; Ramsey, N.F.; Slagter, H.A.; Kahn, R.S. Functional Anatomical Correlates of Controlled and Automatic Processing. J. Cogn. Neurosci. 2001, 13, 730–743. [Google Scholar] [CrossRef]
- Kelly, A.M.C.; Garavan, H. Human Functional Neuroimaging of Brain Changes Associated with Practice. Cereb. Cortex 2004, 15, 1089–1102. [Google Scholar] [CrossRef] [Green Version]
- Kastner, S.; Ungerleider, L.G. Mechanisms of visual attention in the human Cortex. Ann. Rev. Neurosci. 2000, 23, 315–341. [Google Scholar]
- McMains, S.; Kastner, S. Interactions of Top-Down and Bottom-Up Mechanisms in Human Visual Cortex. J. Neurosci. 2011, 31, 587–597. [Google Scholar] [CrossRef]
- Miall, C. 10,000 hours to perfection. Nat. Neurosci. 2013, 16, 1168–1169. [Google Scholar] [CrossRef]
- Corbetta, M.; Shulman, G.L. Control of goal-directed and stimulus-driven attention in the brain. Nat. Rev. Neurosci. 2002, 3, 201–215. [Google Scholar] [CrossRef]
- Quintana, J.; Fuster, J.M. From Perception to Action: Temporal Integrative Functions of Prefrontal and Parietal Neurons. Cereb. Cortex 1999, 9, 213–221. [Google Scholar] [CrossRef]
- Cole, M.W.; Reynolds, J.R.; Power, J.D.; Repovs, G.; Anticevic, A.; Braver, T.S. Multi-task connectivity reveals flexible hubs for adaptive task control. Nat. Neurosci. 2013, 16, 1348–1355. [Google Scholar] [CrossRef]
- Hahn, B.; Ross, T.J.; Stein, E.A. Neuroanatomical dissociation between bottom–up and top–down processes of visuospatial selective attention. NeuroImage 2006, 32, 842–853. [Google Scholar] [CrossRef]
- Fitts, P.M.; Posner, M.I. Human Performance; Oxford University Press: Oxford, UK, 1967; pp. 8–26. [Google Scholar]
- De Martino, B.; Kumaran, D.; Seymour, B.; Dolan, R.J. Frames, Biases, and Rational Decision-Making in the Human Brain. Science 2006, 313, 684–687. [Google Scholar] [CrossRef] [Green Version]
- Debener, S.; Makeig, S.; Delorme, A.; Engel, A.K. What is novel in the novelty oddball paradigm? Functional significance of the novelty P3 event-related potential as revealed by independent component analysis. Cogn. Brain Res. 2005, 22, 309–321. [Google Scholar] [CrossRef] [Green Version]
- Deichmann, R.; Gottfried, J.; Hutton, C.; Turner, R. Optimized EPI for fMRI studies of the orbitofrontal cortex. NeuroImage 2003, 19, 430–441. [Google Scholar] [CrossRef]
- Lieberman, M.D.; Cunningham, W.A. Type I and Type II error concerns in fMRI research: re-balancing the scale. Soc. Cogn. Affect. Neurosci. 2009, 4, 423–428. [Google Scholar] [CrossRef] [Green Version]
- Bush, R.R.; Mosteller, F. Stochastic Models for Learning; John Wiley & Sons, Inc.: Oxford, UK, 1955. [Google Scholar]
- Herrnstein, R.J. Formal properties of the matching law1. J. Exp. Anal. Behav. 1974, 21, 159–164. [Google Scholar] [CrossRef]
- Shulman, G.L.; Astafiev, S.V.; Franke, D.; Pope, D.L.; Snyder, A.Z.; McAvoy, M.P.; Corbetta, M. Interaction of stimulus-driven reorienting and expectation in ventral and dorsal frontoparietal and basal ganglia-cortical networks. J. Neurosci. 2009, 29, 4392–4407. [Google Scholar] [CrossRef]
- Friston, K.; Buechel, C.; Fink, G.R.; Morris, J.; Rolls, E.; Dolan, R.; Friston, K. Psychophysiological and Modulatory Interactions in Neuroimaging. NeuroImage 1997, 6, 218–229. [Google Scholar] [CrossRef] [Green Version]
- Wood, W.; Rünger, D. Psychology of habit. Annu. Rev. Psycho. 2016, 67, 289–314. [Google Scholar] [CrossRef]
- Ringo, J.L. Stimulus specific adaptation in inferior temporal and medial temporal cortex of the monkey. Behav. Brain Res. 1996, 76, 191–197. [Google Scholar] [CrossRef]
- O’Kane, G.; Insler, R.Z.; Wagner, A.D. Conceptual and perceptual novelty effects in human medial temporal cortex. Hippocampus 2005, 15, 326–332. [Google Scholar] [CrossRef]
- Tulving, E.; Markowitsch, H.J.; Craik, F.I.M.; Habib, R.; Houle, S. Novelty and Familiarity Activations in PET Studies of Memory Encoding and Retrieval. Cereb. Cortex 1996, 6, 71–79. [Google Scholar] [CrossRef] [Green Version]
- Kirwan, C.B.; Stark, C.E. Medial temporal lobe activation during encoding and retrieval of novel face-name pairs. Hippocampus 2004, 14, 919–930. [Google Scholar] [CrossRef] [Green Version]
- Habib, R.; McIntosh, A.R.; Wheeler, M.A.; Tulving, E. Memory encoding and hippocampally-based novelty/familiarity discrimination networks. Neuropsycholology 2003, 41, 271–279. [Google Scholar] [CrossRef]
- Paulus, M.P.; Hozack, N.; Zauscher, B.; McDowell, J.E.; Frank, L.; Brown, G.G.; Braff, D.L. Prefrontal, parietal, and temporal cortex networks underlie decision-making in the presence of uncertainty. Neuroimage 2001, 13, 91–100. [Google Scholar] [CrossRef]
- Knutson, B.; Taylor, J.; Kaufman, M.; Peterson, R.; Glover, G. Distributed Neural Representation of Expected Value. J. Neurosci. 2005, 25, 4806–4812. [Google Scholar] [CrossRef]
- Rushworth, M.F.; Mars, R.B.; Summerfield, C. General mechanisms for making decisions? Curr. Opin. Neurobiol. 2009, 19, 75–83. [Google Scholar] [CrossRef]
- Wunderlich, K.; Rangel, A.; O’Doherty, J.P. Economic choices can be made using only stimulus values. Proc. Natl. Acad. Sci. USA 2010, 107, 15005–15010. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, S.C.; Balleine, B.W.; O’Doherty, J.P. Calculating consequences: Brain systems that encode the causal effects of actions. J. Neurosci. 2008, 28, 6750–6755. [Google Scholar] [CrossRef]
- Hare, T.; Camerer, C.; Rangel, A. Self-control in decision-making involves modulation of the vmPFC valuation system. NeuroImage 2009, 47, S95. [Google Scholar] [CrossRef] [Green Version]
- Campbell-Meiklejohn, D.K.; Woolrich, M.W.; Passingham, R.E.; Rogers, R.D. Knowing When to Stop: The Brain Mechanisms of Chasing Losses. Boil. Psychiatry 2008, 63, 293–300. [Google Scholar] [CrossRef] [Green Version]
- Baumgartner, T.; Knoch, D.; Hotz, P.; Eisenegger, C.; Fehr, E. Dorsolateral and ventromedial prefrontal cortex orchestrate normative choice. Nat. Neurosci. 2011, 14, 1468–1474. [Google Scholar] [CrossRef] [Green Version]
- Koechlin, E.; Ody, C.; Kouneiher, F. The Architecture of Cognitive Control in the Human Prefrontal Cortex. Science 2003, 302, 1181–1185. [Google Scholar] [CrossRef] [Green Version]
- Venkatraman, V.; Payne, J.W.; Bettman, J.R.; Luce, M.F.; Huettel, S.A. Separate neural mechanisms underlie choices and strategic preferences in risky decision making. Neuron 2009, 62, 593–602. [Google Scholar] [CrossRef]
- De Martino, B.; Fleming, S.M.; Garrett, N.; Dolan, R.J. Confidence in value-based choice. Nat. Neurosci. 2013, 16, 105. [Google Scholar] [CrossRef]
- Haber, S.N.; Knutson, B. The Reward Circuit: Linking Primate Anatomy and Human Imaging. Neuropsychopharmacol. 2009, 35, 4–26. [Google Scholar] [CrossRef] [Green Version]
- Brighton, H.; Gigerenzer, G. Homo Heuristicus: Less-is-More Effects in Adaptive Cognition. Malays. J. Med Sci. 2012, 19, 6–16. [Google Scholar]
- Daw, N.D.; Gershman, S.J.; Seymour, B.; Dayan, P.; Dolan, R.J. Model-based influences on humans’ choices and striatal prediction errors. Neuron 2011, 69, 1204–1215. [Google Scholar] [CrossRef]
- Bjork, J.M.; Hommer, D.W. Anticipating instrumentally obtained and passively-received rewards: A factorial fMRI investigation. Behav. Brain Res. 2007, 177, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, J.; Scheef, L.; Hüppe, C.; Boecker, H. Distinct striatal regions for planning and executing novel and automated movement sequences. NeuroImage 2009, 44, 1369–1379. [Google Scholar] [CrossRef]
- Kable, J.W.; Glimcher, P.W. The Neurobiology of Decision: Consensus and Controversy. Neuron 2009, 63, 733–745. [Google Scholar] [CrossRef] [Green Version]
- Mink, J.W. THE BASAL GANGLIA: FOCUSED SELECTION AND INHIBITION OF COMPETING MOTOR PROGRAMS. Prog. Neurobiol. 1996, 50, 381–425. [Google Scholar] [CrossRef]
- Redgrave, P.; Prescott, T.; Gurney, K. The basal ganglia: A vertebrate solution to the selection problem? Neurosci. 1999, 89, 1009–1023. [Google Scholar] [CrossRef]
- Rangel, A.; Clithero, J.A. The computation of stimulus values in simple choice. In Neuroeconomics: Decision-Making and the Brain, 2nd ed.; Glimcher, P., Fehr, E., Eds.; Academic Press: Cambridge, MA, USA, 2013; pp. 125–149. [Google Scholar]
- Lewis, S.J.G.; Dove, A.; Robbins, T.W.; A Barker, R.; Owen, A.M. Striatal contributions to working memory: a functional magnetic resonance imaging study in humans. F1000 Post-Publ. Peer Rev. Biomed. Lit. 2004, 19, 755–760. [Google Scholar] [CrossRef]
- Goel, V. Anatomy of deductive reasoning. Trends Cogn. Sci. 2007, 11, 435–441. [Google Scholar] [CrossRef]
- Nomura, E.M.; Maddox, W.T.; Filoteo, J.V.; Ing, A.D.; Gitelman, D.R.; Parrish, T.B.; Mesulam, M.M.; Reber, P.J. Neural correlates of rule-based and information-integration visual category learning. Cereb. Cortex 2007, 17, 37–43. [Google Scholar] [CrossRef]
- Dreher, J.C.; Kohn, P.; Berman, K.F. Neural coding of distinct statistical properties of reward information in humans. Cereb. Cortex 2006, 16, 561–573. [Google Scholar] [CrossRef]
- Buch, E.R.; Brasted, P.J.; Wise, S.P. Comparison of population activity in the dorsal premotor cortex and putamen during the learning of arbitrary visuomotor mappings. Exp. Brain Res. 2006, 169, 69–84. [Google Scholar] [CrossRef]
- Antzoulatos, E.G.; Miller, E.K. Differences between Neural Activity in Prefrontal Cortex and Striatum during Learning of Novel, Abstract Categories. Neuron 2011, 71, 243–249. [Google Scholar] [CrossRef]
- Volz, K.G.; I Schubotz, R.; Von Cramon, D. Predicting events of varying probability: uncertainty investigated by fMRI. NeuroImage 2003, 19, 271–280. [Google Scholar] [CrossRef] [Green Version]
- Torta, D.; Cauda, F. Different functions in the cingulate cortex, a meta-analytic connectivity modeling study. NeuroImage 2011, 56, 2157–2172. [Google Scholar] [CrossRef]
- Mushtaq, F.; Bland, A.R.; Schaefer, A. Uncertainty and cognitive control. Front. Psychol. 2011, 2, 249. [Google Scholar] [CrossRef]
- Botvinick, M.M. Conflict monitoring and decision making: Reconciling two perspectives on anterior cingulate function. Cogn. Affect. Behav. Neurosci. 2007, 7, 356–366. [Google Scholar] [CrossRef]
- Friston, K.J.; Holmes, A.P.; Worsley, K.J. How Many Subjects Constitute a Study? NeuroImage 1999, 10, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Desmond, J.E.; Glover, G.H. Estimating sample size in functional MRI (fMRI) neuroimaging studies: Statistical power analyses. J. Neuro. Methods 2002, 118, 115–128. [Google Scholar] [CrossRef] [Green Version]
- Turner, B.O.; Paul, E.J.; Miller, M.B.; Barbey, A.K. Small sample sizes reduce the replicability of task-based fMRI studies. Commun. Boil. 2018, 1, 62. [Google Scholar] [CrossRef]
- Edwards, W. Probability learning in 1000 trials. J. Exp. Psychol. 1961, 62, 385–394. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cluster Size (Voxels) | Montreal Neurological Institute (MNI) x, y, z {mm} | F-Value (Peak) | Side | Regions of Activation |
|---|---|---|---|---|
| Frontal Cortex | ||||
| 355 | −6, 23, −14 | 26.79 | L | Medial Orbito Frontal |
| 168 | −27, 32, 52 | 22.93 | L | Middle/Superior Frontal Gyri |
| 62 | −33, −1, 58 | 22.95 | L | Middle Frontal Gyrus |
| 29 | 27, 50, −8 | 20.00 | R | Middle/Superior Frontal Gyri |
| Temporal/Temporoparietal Cortex | ||||
| 404 | −51, −73, 25 | 49.50 | L | Temporal Gyrus |
| −60, −58, 22 | 39.66 | L | Angular Gyrus | |
| 174 | 57, −64, 28 | 41.10 | R | Angular Gyrus |
| −3, 29, −2 | 14.32 | L | Anterior Cingulate | |
| 143 | −54, 5, −20 | 26.73 | L | Middle Temporal Gyrus |
| −45, 20, −29 | 18.64 | L | Superior Frontal Gyrus | |
| 146 | −63, −37, 1 | 25.92 | L | Middle Temporal Gyrus |
| 77 | 54, −1, −17 | 22.79 | R | Middle Temporal Gyrus |
| Parietal Cortex | ||||
| 15859 | 24, −67, 40 | 121.39 | R | Precuneus |
| 248 | −6, −52, 34 | 24.76 | L | Precuneus |
| 68 | 60, −13, 25 | 33.07 | R | Postcentral gyrus |
| 65 | 45, −25, 64 | 28.72 | R | Postcentral gyrus |
| 46 | −12, 62, 19 | 18.06 | L | Precuneus |
| 11 | −12, −40, 67 | 14.63 | L | Postcentral gyrus |
| Other | ||||
| 150 | 0, −28, 28 | 51.93 | Cingulate Gyrus | |
| 24 | −30, −85, −29 | 23.56 | L | Cerebellum |
| 20 | 27, −88, −29 | 16.39 | R | Cerebellum |
| Cluster Size (Voxels) | MNI x, y, z {mm} | F−Value (Peak) | Side | Regions of Activation |
|---|---|---|---|---|
| Main Effect of Uncertainty | ||||
| 158 | 3, 26, 46 | 14.22 | R | Superior Medial Gyrus (BA8) |
| 62 | 33, 23, −5 | 17.76 | R | Inferior Frontal Gyrus |
| 87 | −30, 23, 4 | 17.72 | L | Inferior Frontal Gyrus |
| Striatum | ||||
| 95 | −30, −7, −2 | 14.27 | L | Putamen |
| 47 | 27, −1, −2 | 10.77 | R | Putamen |
| 10 | 12,20,1 | 8.88 | R | Caudate |
| Temporal/Temporoparietal | ||||
| 139 | −54, −58, 22 | 11.2 | L | Middle Temporal Gyrus |
| −45, −73, 28 | 9.52 | L | Angular Gyrus | |
| 30 | −45, −55, −8 | 13.24 | L | Inferior Temporal Gyrus |
| 16 | −51, 35, 28 | 10.75 | L | Middle Temporal Gyrus |
| 14 | −15, −34, 43 | 10.46 | R | Middle Temporal Gyrus |
| Parietal | ||||
| 226 | −3, −55, 31 | 12.37 | L | Precuneus |
| −6, −43, 31 | 11.43 | L | Posterior Cingulate Cortex | |
| 13 | −45, −67, 43 | 9.37 | L | Angular Gyrus |
| 20 | 54, −58, 28 | 9.11 | R | Angular Gyrus |
| 16 | −39, −52, 52 | 9.34 | L | Inferior Parietal Lobule |
| Uncertainty × Novelty Interaction | ||||
| Frontal Cortex | ||||
| 25 | 12, 20, 52 | 9.84 | R | Superior/Medial (BA8) |
| 23 | −18, 23, 28 | 12.13 | L | Superior Frontal Gyrus |
| 16 | 42, 14, 46 | 10.6 | R | Middle Frontal Gyrus (BA8) |
| Parietal and Posterior Cingulate Cortex | ||||
| 48 | −39, −40, 31 | 19.18 | L | Inferior Parietal Lobule (BA40) |
| 16 | 39, −46, 37 | 9.19 | R | Inferior Parietal Lobule (BA40) |
| 43 | −15, −55, 34 | 11.67 | L | Precuneus |
| −18, −52, 25 | 9.7 | L | Cingulate Gyrus | |
| 22 | 12, −76, 46 | 9.46 | R | Precuneus |
| 119 | 21, −46, 28 | 12.96 | R | Cingulate Gyrus |
| 117 | 9, 38, 34 | 16.79 | R | Middle Cingulate Cortex |
| 21, −64, 40 | 9.31 | R | Right Precuneus | |
| Occipital Cortex | ||||
| 124 | −27, −91, 19 | 11.01 | L | Middle Occipital Gyrus |
| 78 | −45, −85, −2 | 12.4 | L | Middle/Inferior Occipital Gyri |
| 25 | 21, −82, 10 | 10.28 | R | Calcarine Gyrus (BA 17) |
| 41 | 33, −67, 1 | 9.85 | R | Middle Occipital Gyrus |
| Cluster Size (Voxel) | MNI x, y, z {mm} | T-Value (Peak) | Side | Regions of Activation |
|---|---|---|---|---|
| Frontal Cortex | ||||
| 75 | 42, 32, 19 | 4.65 | R | Middle Frontal Gyrus (BA46) |
| 14 | −42, 29, 19 | 3.75 | L | Middle Frontal Gyrus (BA46) |
| 70 | 45, 8, 37 | 4.61 | R | Precentral Gyrus |
| 67 | −42, −1, 37 | 4.27 | L | Precentral Gyrus |
| 28 | 24, −1, 49 | 4.21 | R | Premotor Area (BA6) |
| Temporal Cortex | ||||
| 312 | 48. −58, −11 | 5.75 | R | Inferior Temporal & Fusiform Gyri |
| −45, −58, −11 | 4.79 | L | Inferior Temporal Gyrus | |
| 48, 0.79, 0.5 | 3.44 | R | Inferior Occipital Gyrus | |
| Parieto0ccipitalCortex | ||||
| 557 | 33, −49, 52 | 4.65 | R | Inferior Parietal Lobule |
| −24, −73, 46 | 5.48 | L | Superior Parietal Lobule | |
| 27, −70, 43 | 5.93 | R | Sup/Mid/Inf Occipital Gyri | |
| 39, −79, 16 | 4.61 | R | Middle Occipital Gyrus | |
| 27, −70, 43 | 5.93 | R | Sup/Mid/Inf Occipital Gyri | |
| Insular Cortex | ||||
| 29 | −27, 23, −2 | 4.8 | L | Insula |
| Occipital Cortex | ||||
| 809 | 45, −79, −8 | 5.7 | L | Inferior Occipital Gyrus |
| Cluster Size (Voxel) | MNI x, y, z {mm} | T-Value (Peak) | Side | Regions of Activation |
|---|---|---|---|---|
| Frontal | ||||
| 13 | −12, 56, 4 | 3.56 | L | Medial Frontal Gyrus (BA10) |
| 12 | 12, 47, −5 | 3.54 | R | Medial Frontal Gyrus (BA 10/BA32) |
| Temporal | ||||
| 28 | −45, −79, 34 | 4.03 | L | Middle Temporal Gyrus |
| −51, −76, 28 | 3.86 | L | Middle Temporal Gyrus | |
| Parietal and Cingulate | ||||
| 12 | −15, −52, 37 | 3.99 | L | Precuneus |
| 19 | 66, −43, 31 | 3.52 | R | Supramarginal Gyrus |
| 19 | 18, 2, 28 | 4.14 | R | Cingulate Gyrus |
| 23 | 9, −19, 43 | 3.77 | R | Middle Cingulate Cortex |
| 30 | 12, 91, 34 | 3.7 | R | Superior Occipital Gyrus |
| 9, −91, 19 | 3.51 | R | Cuneus (BA7) | |
| 14 | −9, −76,31 | 3.4 | L | Cuneus (BA7) |
| Occipital Cortex | ||||
| 30 | 12, 91, 34 | 3.7 | R | Superior Occipital Gyrus |
| 13 | 15, 43, −5 | 3.66 | R | Lingual Gyrus |
| Other | ||||
| 44 | 3, 20, 16 | 4.25 | R | Corpus Callosum |
| 9, 11, 22 | 3.44 | L | Corpus Collosum | |
| Cluster Size (Voxel) | MNI x, y, z {mm} | T−Value (Peak) | Side | Regions of Activation |
|---|---|---|---|---|
| Striatum | ||||
| 30 | 9, 11, −2 | 4.5 | R | Caudate |
| 18, 5, −11 | 4.37 | R | Putamen | |
| 30, 2, −14 | 4.06 | R | Putamen | |
| 13 | −33, −25, −8 | 4.99 | L | Caudate Tail covering hippocampus |
| Frontal | ||||
| 293 | −3, 5, 58 | 6.69 | L | Supplementary Motor Area |
| 9, 5, 28 | 4.69 | R | Corpus Callosum | |
| −6, 20, 34 | 4.63 | L | Cingulate Gyrus | |
| 87 | 45, 8, 28 | 5.79 | R | Inferior Frontal Gyrus |
| 48 | −45,8,28 | 4.92 | L | Inferior Frontal Gyrus |
| 17 | 33, −1, 61 | 4.44 | R | Middle Frontal Gyrus |
| 10 | 39, 44, 22 | 4.18 | R | Middle Frontal Gyrus |
| 38 | −48, −19, 37 | 4.48 | L | Precentral Gyrus |
| 30 | 18, −31, 64 | 4.37 | R | Precentral Gyrus (BA 4) |
| 38 | −48, −19, 37 | 4.48 | L | Precentral Gyrus |
| Temporal | ||||
| 534 | 33, −16, 43 | 6.54 | R | Middle Temporal Gyrus |
| 48, −67, 4 | 5.36 | R | BA 37 | |
| 13 | 36, 2, −26 | 5.89 | R | Superior Temporal/R Amygdala |
| Insula | ||||
| 189 | −27, 17, 22 | 5.62 | L | Insula |
| 22 | 30, 20, 4 | 5.02 | R | Insula |
| Parietal | ||||
| 65 | 21, −61, 58 | 5.84 | R | Superior Parietal Lobule/BA 7 |
| 13 | −30, −34, 43 | 4.22 | L | Parietal Lobe |
| 67 | −24, −91, 19 | 4.76 | L | Cuneus |
| −6, −97, 16 | 4.28 | L | BA 18 (Occipital) | |
| Occipital/Temporal | ||||
| 314 | −42, 52, −5 | 7.57 | L | Occipital Lobe |
| −42, 64, 4 | 5.65 | L | Middle Temporal Gyrus | |
| −36, −40, 7 | 552 | L | Superior Temporal Gyrus | |
| 162 | −15, −88, −11 | 6.8 | L | Occipital Lobe |
| −24, −88, −11 | 6.34 | L | Lingual Gyrus | |
| 60 | 33, −76, −11 | 5.69 | R | Occipital Lobe |
| Limbic | ||||
| 20 | −12, −7, 34 | 4.71 | L | Cingulate Gyrus |
| 20 | −12, −34, 34 | 4.5 | L | Cingulate Gyrus |
| Other | ||||
| 23 | −48, −58, −23 | 7.39 | L | Cerebellum Posterior Lobe |
| 16 | −21, −64, −44 | 4.84 | L | Cerebellum Posterior Lobe |
| 14 | 30, −64, −47 | 5.05 | R | Cerebellum Posterior Lobe |
| 44 | −18, −61, −14 | 4.73 | L | Cerebellum |
| 10 | 21, −13, −2 | 4.36 | R | Thalamus |
| Cluster Size (Voxel) | MNI x, y, z {mm} | T-Value (Peak) | Side | Regions of Activation |
|---|---|---|---|---|
| Striatum | ||||
| 88 | −36, −4, 7 | 8.31 | L | Putamen |
| 31 | 30, 17, 4 | 6.33 | R | Putamen |
| Temporal Cortex | ||||
| 10 | 51, −58, 4 | 5.35 | R | Middle Temporal Gyrus |
| Parietal Cortex | ||||
| 23 | 30, −61, 43 | 7.03 | R | Precuneus |
| 20 | −33, 0.52, 46 | 6.69 | L | Parietal Lobe |
| 24 | 33, −52, 55 | 6.44 | R | Inferior Parietal Lobule |
| 38 | 51, −25, 52 | 6.06 | R | Postcentral Gyrus |
| Occipital Cortex | ||||
| 78 | 49, 48, 4 | 7.03 | L | Occipital Lobe/Inf Occ Gyrus |
| 16 | 36, 48, 13 | 5.35 | R | Occipital Lobe |
| Limbic | ||||
| 31 | −3, 14, 40 | 6.64 | L | Cingulate Gyrus (BA32) |
| 6, 14, 43 | 5.07 | R | Cingulate Gyrus | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Erdeniz, B.; Done, J. Common and Distinct Functional Brain Networks for Intuitive and Deliberate Decision Making. Brain Sci. 2019, 9, 174. https://doi.org/10.3390/brainsci9070174
Erdeniz B, Done J. Common and Distinct Functional Brain Networks for Intuitive and Deliberate Decision Making. Brain Sciences. 2019; 9(7):174. https://doi.org/10.3390/brainsci9070174
Chicago/Turabian StyleErdeniz, Burak, and John Done. 2019. "Common and Distinct Functional Brain Networks for Intuitive and Deliberate Decision Making" Brain Sciences 9, no. 7: 174. https://doi.org/10.3390/brainsci9070174