Different Hemodynamic Responses of the Primary Motor Cortex Accompanying Eccentric and Concentric Movements: A Functional NIRS Study



{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
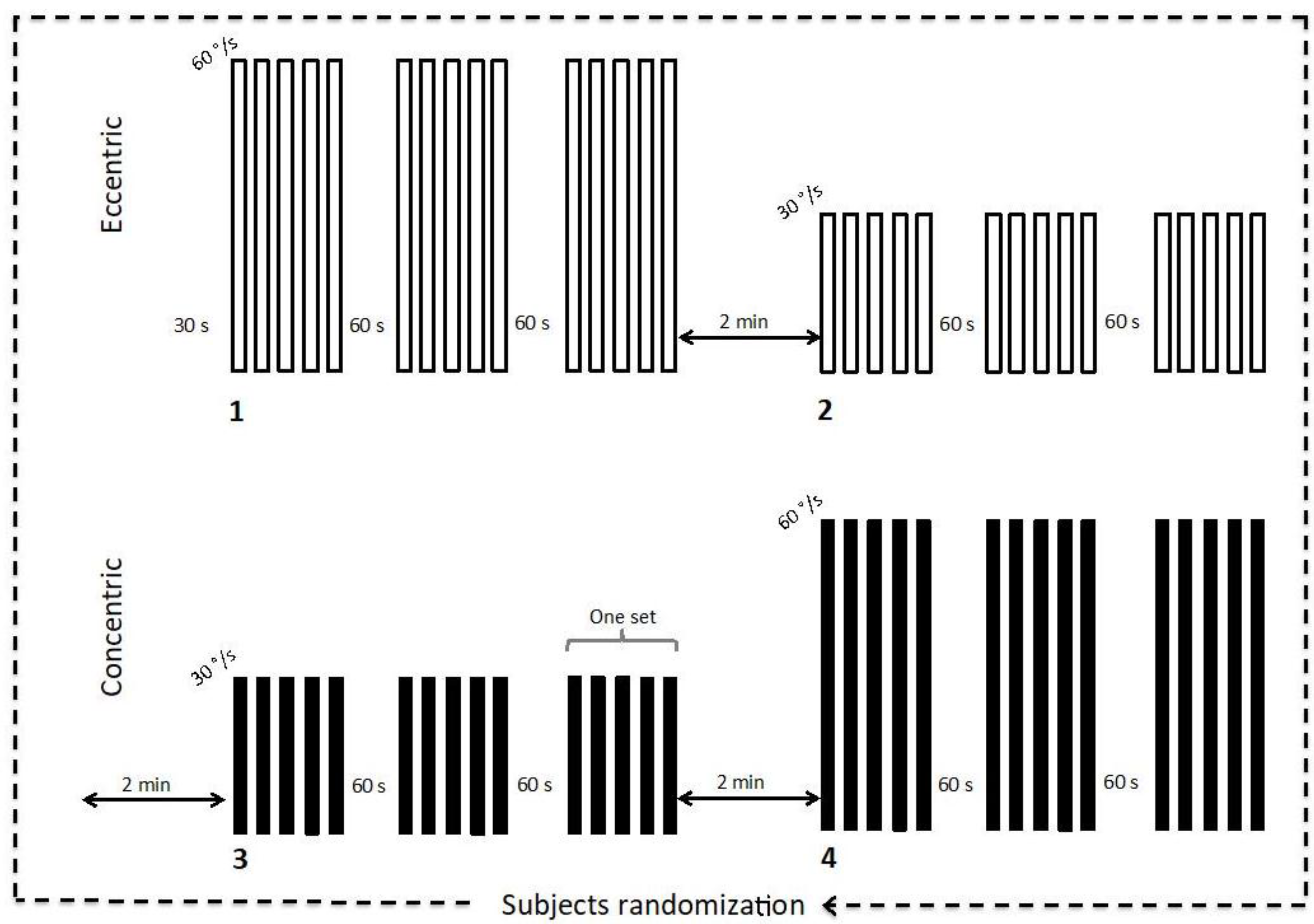
2.2. Experimental Protocol
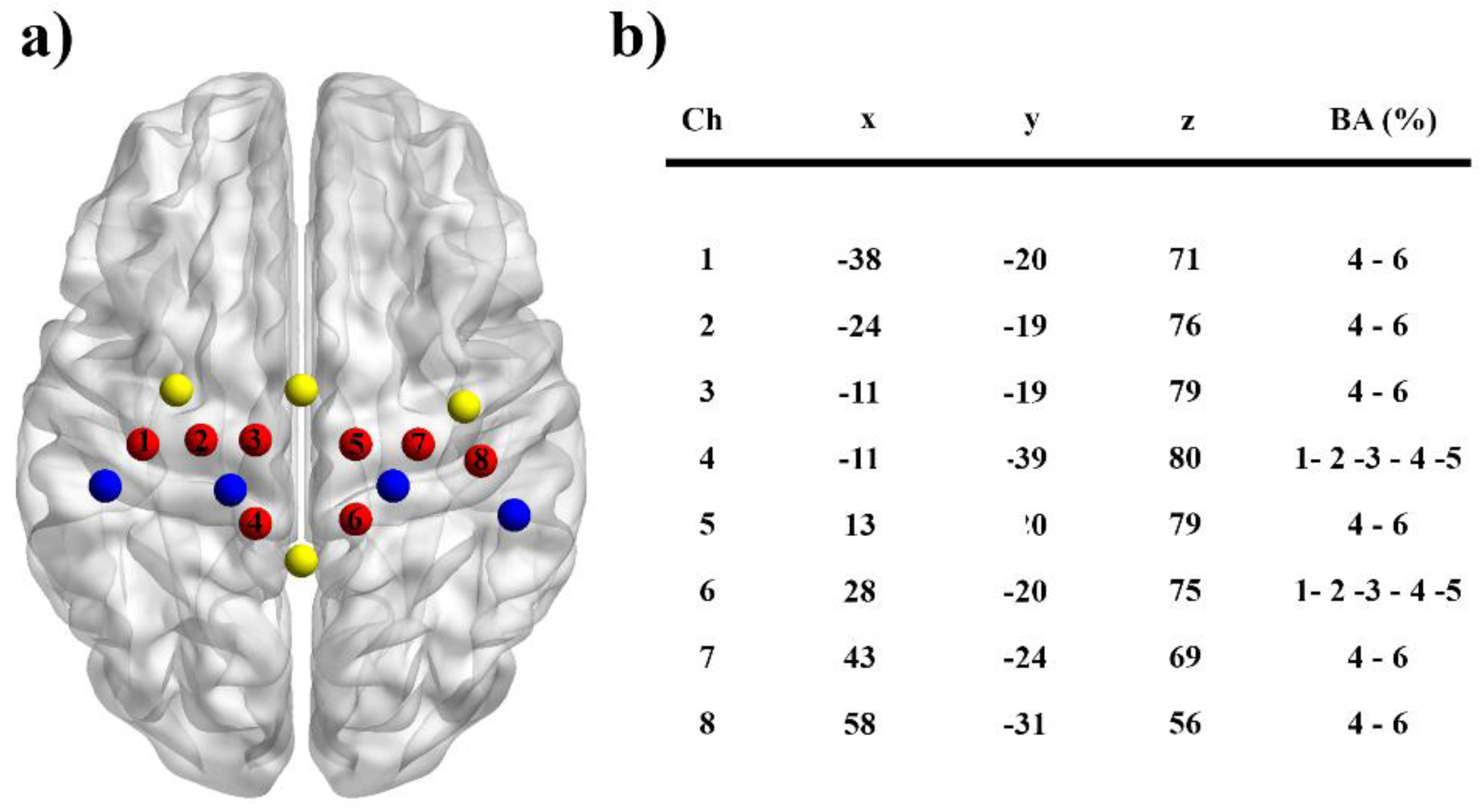
2.3. fNIRS Recordings
2.4. Data Analysis
2.4.1. fNIRS Responses
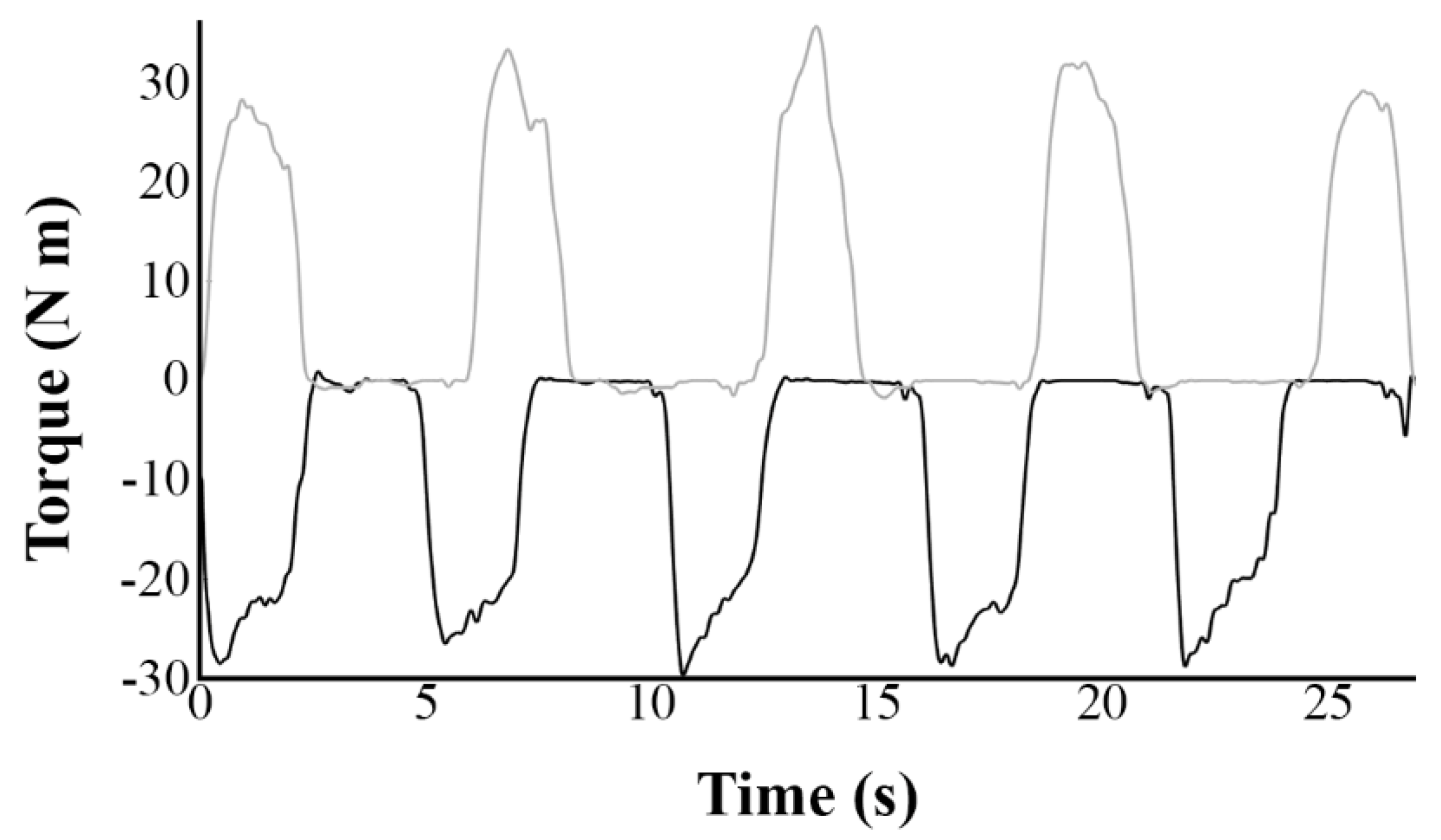
2.4.2. Torque Values
2.5. Statistical Analysis
3. Results
3.1. Torque Values
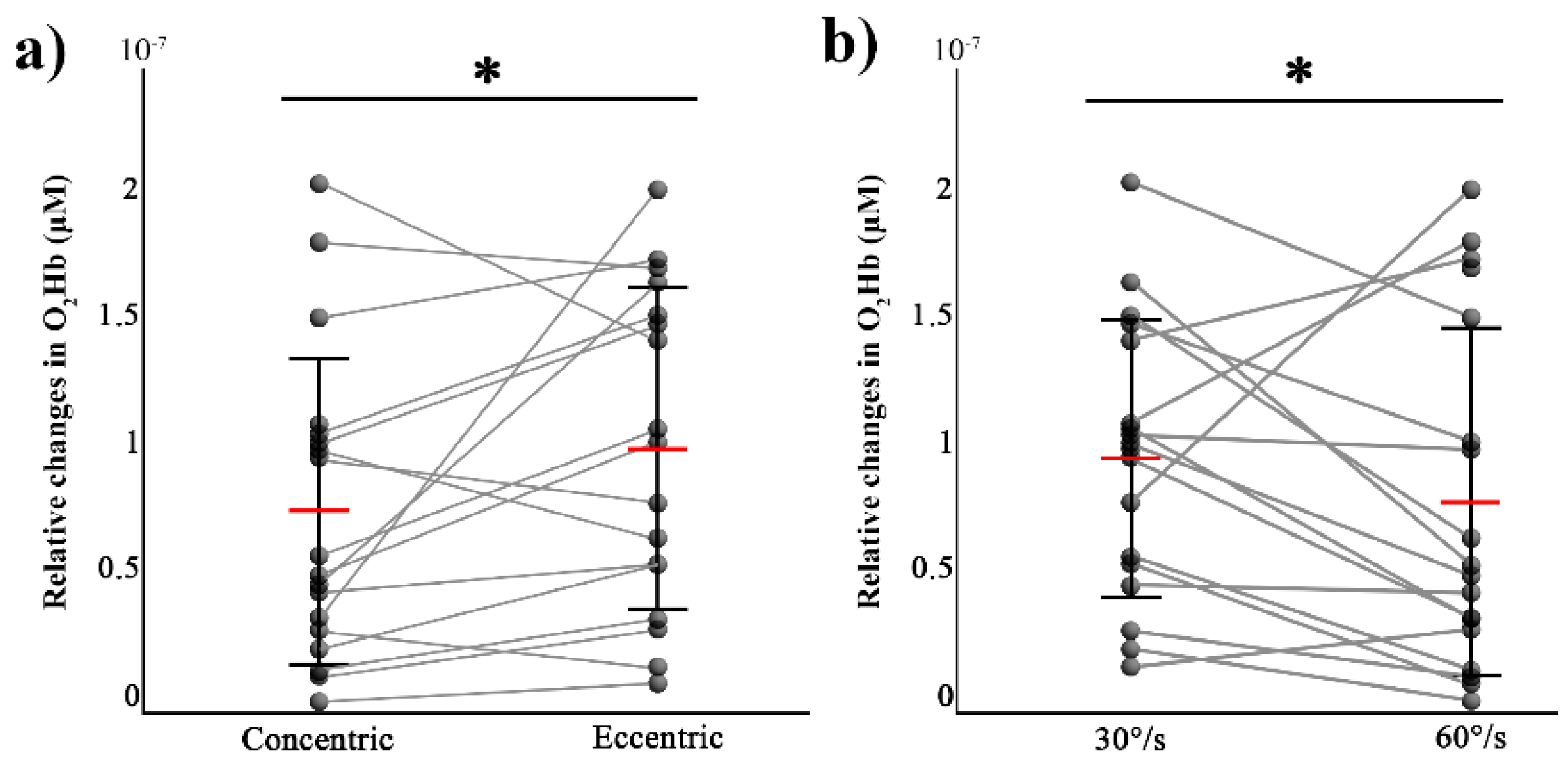
3.2. fNIRS Responses
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Perrey, S.; Betik, A.; Candau, R.; Rouillon, J.D.; Hughson, R.L. Comparison of oxygen uptake kinetics during concentric and eccentric cycle exercise. J. Appl. Physiol. 2001, 91, 2135–2142. [Google Scholar] [CrossRef] [PubMed]
- Dufour, S.P.; Doutreleau, S.; Lonsdorfer-Wolf, E.; Lampert, E.; Hirth, C.; Piquard, F.; Lonsdorfer, J.; Geny, B.; Mettauer, B.; Richard, R. Deciphering the metabolic and mechanical contributions to the exercise-induced circulatory response: Insights from eccentric cycling. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 292, R1641–R1648. [Google Scholar] [CrossRef] [PubMed]
- Peñailillo, L.; Mackay, K.; Abbiss, C.R. RPE during concentric and eccentric cycling: Are we measuring effort or exertion? Int. J. Sports Physiol. Perform. 2017, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Duchateau, J.; Enoka, R.M. Neural control of shortening and lengthening contractions: Influence of task constraints. J. Physiol. 2008, 586, 5853–5864. [Google Scholar] [CrossRef] [PubMed]
- Perrey, S. Brain activation associated with eccentric movement: A narrative review of the literature. Eur. J. Sport Sci. 2018, 18, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Kwon, Y.H.; Park, J.W. Different cortical activation patterns during voluntary eccentric and concentric muscle contractions: An fMRI study. NeuroRehabilitation 2011, 29, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Sekiguchi, H.; Kimura, T.; Yamanaka, K.; Nakazawa, K. Lower excitability of the corticospinal tract to transcranial magnetic stimulation during lengthening contractions in human elbow flexors. Neurosci. Lett. 2001, 312, 83–86. [Google Scholar] [CrossRef]
- Olsson, C.J.; Hedlund, M.; Sojka, P.; Lundström, R.; Lindström, B. Increased prefrontal activity and reduced motor cortex activity during imagined eccentric compared to concentric muscle actions. Front. Hum. Neurosci. 2012, 6, 255. [Google Scholar] [CrossRef] [PubMed]
- Yao, W.X.; Li, J.; Jiang, Z.; Gao, J.H.; Franklin, C.G.; Huang, Y.; Lancaster, J.L.; Yue, G.H. Aging interferes central control mechanism for eccentric muscle contraction. Front. Aging Neurosci. 2014, 6, 86. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Siemionow, V.; Sahgal, V.; Xiong, F.; Yue, G.H. Greater movement-related cortical potential during human eccentric versus concentric muscle contractions. J. Neurophysiol. 2001, 86, 1764–1772. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.; Siemionow, V.; Sahgal, V.; Xiong, F.; Yue, G.H. Distinct brain activation patterns for human maximal voluntary eccentric and concentric muscle actions. Brain Res. 2004, 1023, 200–212. [Google Scholar] [CrossRef] [PubMed]
- Perrey, S. Non-invasive NIR spectroscopy of human brain function during exercise. Methods 2008, 45, 289–299. [Google Scholar] [CrossRef] [PubMed]
- Obrig, H.; Villringer, A. Beyond the visible—Imaging the human brain with light. J. Cereb. Blood Flow Metab. 2003, 23, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Oldfield, R.C. The assessment and analysis of handedness: The Edinburgh inventory. Neuropsychologia 1971, 9, 97–113. [Google Scholar] [CrossRef]
- Perrin, D.H. Lsokinetic Exercise and Assessment; Human Kinetics Publishers: Champaign, IL, USA, 1993; pp. 21–89. [Google Scholar]
- Singh, A.K.; Okamoto, M.; Dan, H.; Jurcak, V.; Dan, I. Spatial registration of multichannel multi-subject fNIRS data to MNI space without MRI. Neuroimage 2005, 27, 842–851. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.C.; Tak, S.; Jang, K.E.; Jung, J.; Jang, J. NIRS-SPM: Statistical parametric mapping for near-infrared spectroscopy. Neuroimage 2009, 44, 428–447. [Google Scholar] [CrossRef] [PubMed]
- Eickhoff, S.B.; Stephan, K.E.; Mohlberg, H.; Grefkes, C.; Fink, G.R.; Amunts, K.; Zilles, K. A new SPM toolbox for combining probabilistic cytoarchitectonic maps and functional imaging data. Neuroimage 2005, 25, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Huppert, T.J.; Diamond, S.G.; Franceschini, M.A.; Boas, D.A. HomER: A review of time-series analysis methods for near-infrared spectroscopy of the brain. Appl. Opt. 2009, 48, D280–D298. [Google Scholar] [CrossRef] [PubMed]
- Scholkmann, F.; Spichtig, S.; Muehlemann, T.; Wolf, M. How to detect and reduce movement artifacts in near-infrared imaging using moving standard deviation and spline interpolation. Physiol. Meas. 2010, 31, 649. [Google Scholar] [CrossRef] [PubMed]
- Molavi, B.; Dumont, G.A. Wavelet-based motion artifact removal for functional near-infrared spectroscopy. Physiol. Meas. 2012, 33, 259. [Google Scholar] [CrossRef] [PubMed]
- Cooper, R.; Selb, J.; Gagnon, L.; Phillip, D.; Schytz, H.W.; Iversen, H.K.; Ashina, M.; Boas, D.A. A systematic comparison of motion artifact correction techniques for functional near-infrared spectroscopy. Front. Neurosci. 2012, 6, 147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duncan, A.; Meek, J.H.; Clemence, M.; Elwell, C.E.; Fallon, P.; Tyszczuk, L.; Cope, M.; Delpy, D.T. Measurement of cranial optical path length as a function of age using phase resolved near infrared spectroscopy. Pediatr. Res. 1996, 39, 889–894. [Google Scholar] [CrossRef] [PubMed]
- Scholkmann, F.; Kleiser, S.; Metz, A.J.; Zimmermann, R.; Pavia, J.M.; Wolf, U.; Wolf, M. A review on continuous wave functional near-infrared spectroscopy and imaging instrumentation and methodology. Neuroimage 2014, 85, 6–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Themelis, G.; D’Arceuil, H.E.; Diamond, S.G.; Thaker, S.; Huppert, T.J.; Boas, D.A.; Franceschini, M.A. Near-infrared spectroscopy measurement of the pulsatile component of cerebral blood flow and volume from arterial oscillations. J. Biomed. Opt. 2007, 12, 014033. [Google Scholar] [CrossRef] [PubMed]
- Lakens, D. Calculating and reporting effect sizes to facilitate cumulative science: A practical primer for t-tests and ANOVAs. Front. Psychol. 2013, 4, 863. [Google Scholar] [CrossRef] [PubMed]
- Chapman, D.; Newton, M.; Nosaka, K. Eccentric torque-velocity relationship of the elbow flexors. Isokinet. Exerc. Sci. 2005, 13, 139–145. [Google Scholar]
- De Ruiter, C.J.; De Haan, A. Similar effects of cooling and fatigue on eccentric and concentric force-velocity relationships in human muscle. J. Appl. Physiol. 2001, 90, 2109–2116. [Google Scholar] [CrossRef] [PubMed]
- Griffin, J.W. Differences in elbow flexion torque measured concentrically, eccentrically and isometrically. Phys. Ther. 1987, 67, 1205–1208. [Google Scholar] [CrossRef] [PubMed]
- Anwar, A.R.; Muthalib, M.; Perrey, S.; Galka, A.; Granert, O.; Wolff, S.; Heute, U.; Deuschl, G.; Raethjen, J.; Muthuraman, M. Effective connectivity of cortical sensorimotor networks during finger movement tasks: A simultaneous fNIRS, fMRI, EEG study. Brain Topogr. 2016, 29, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Thomson, D.A. Cardiac output during positive and negative work. Scand. J. Clin. Lab. Investig. 1971, 27, 193–200. [Google Scholar] [CrossRef]
- Moritani, T.; Muramatsu, S.; Muro, M. Activity of motor units during concentric and eccentric contractions. Am. J. Phys. Med. 1987, 66, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Nakazawa, K.; Kawakami, Y.; Fukunaga, T.; Yano, H.; Miyashita, M. Differences in activation patterns in elbow flexor muscles during isometric, concentric and eccentric contractions. Eur. J. Appl. Physiol. Occup. Physiol. 1993, 66, 214–220. [Google Scholar] [CrossRef] [PubMed]
- Kellis, E.; Baltzopoulos, V. Muscle activation differences between eccentric and concentric isokinetic exercise. Med. Sci. Sports Exerc. 1998, 30, 1616–1623. [Google Scholar] [CrossRef] [PubMed]
- Yue, G.H.; Liu, J.Z.; Siemionow, V.; Ranganathan, V.K.; Ng, T.C.; Sahgal, V. Brain activation during human finger extension and flexion movements. Brain Res. 2000, 856, 291–300. [Google Scholar] [CrossRef]
- Gruber, M.; Linnamo, V.; Strojnik, V.; Rantalainen, T.; Avela, J. Excitability at the motoneuron pool and motor cortex is specifically modulated in lengthening compared to isometric contractions. J. Neurophysiol. 2009, 101, 2030–2040. [Google Scholar] [CrossRef] [PubMed]
- Vergotte, G.; Torre, K.; Chirumamilla, V.C.; Anwar, A.R.; Groppa, S.; Perrey, S.; Muthuraman, M. Dynamics of the human brain network revealed by time-frequency effective connectivity in fNIRS. Biomed. Opt. Express 2017, 8, 5326–5341. [Google Scholar] [CrossRef] [PubMed]




© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Borot, L.; Vergotte, G.; Perrey, S. Different Hemodynamic Responses of the Primary Motor Cortex Accompanying Eccentric and Concentric Movements: A Functional NIRS Study. Brain Sci. 2018, 8, 75. https://doi.org/10.3390/brainsci8050075
Borot L, Vergotte G, Perrey S. Different Hemodynamic Responses of the Primary Motor Cortex Accompanying Eccentric and Concentric Movements: A Functional NIRS Study. Brain Sciences. 2018; 8(5):75. https://doi.org/10.3390/brainsci8050075
Chicago/Turabian StyleBorot, Lénaic, Grégoire Vergotte, and Stéphane Perrey. 2018. "Different Hemodynamic Responses of the Primary Motor Cortex Accompanying Eccentric and Concentric Movements: A Functional NIRS Study" Brain Sciences 8, no. 5: 75. https://doi.org/10.3390/brainsci8050075