
Neural Basis of Brain Dysfunction Produced by Early Sleep Problems

Abstract
:1. Introduction
2. Sleep Affects the Structure and Function of the Brain

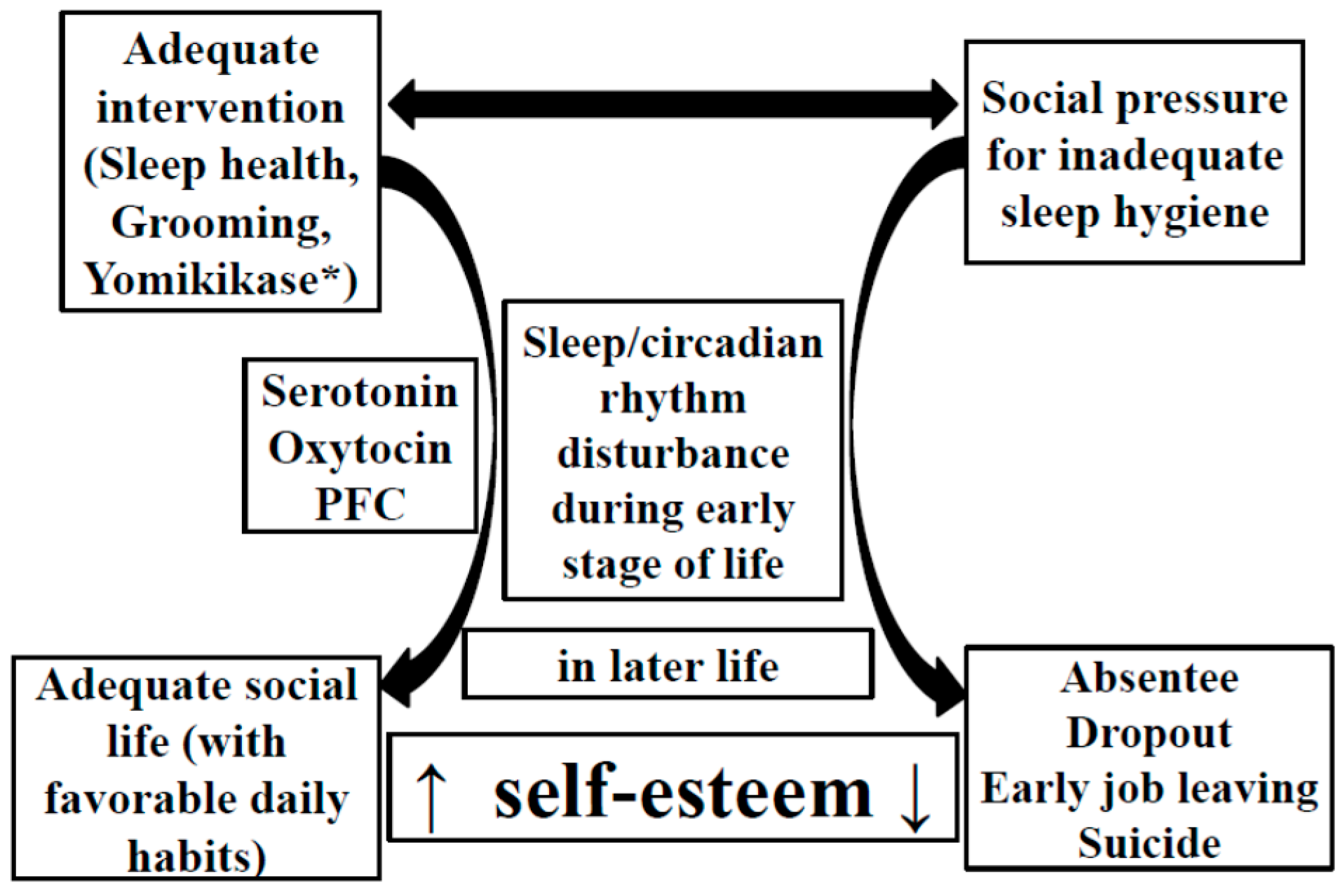
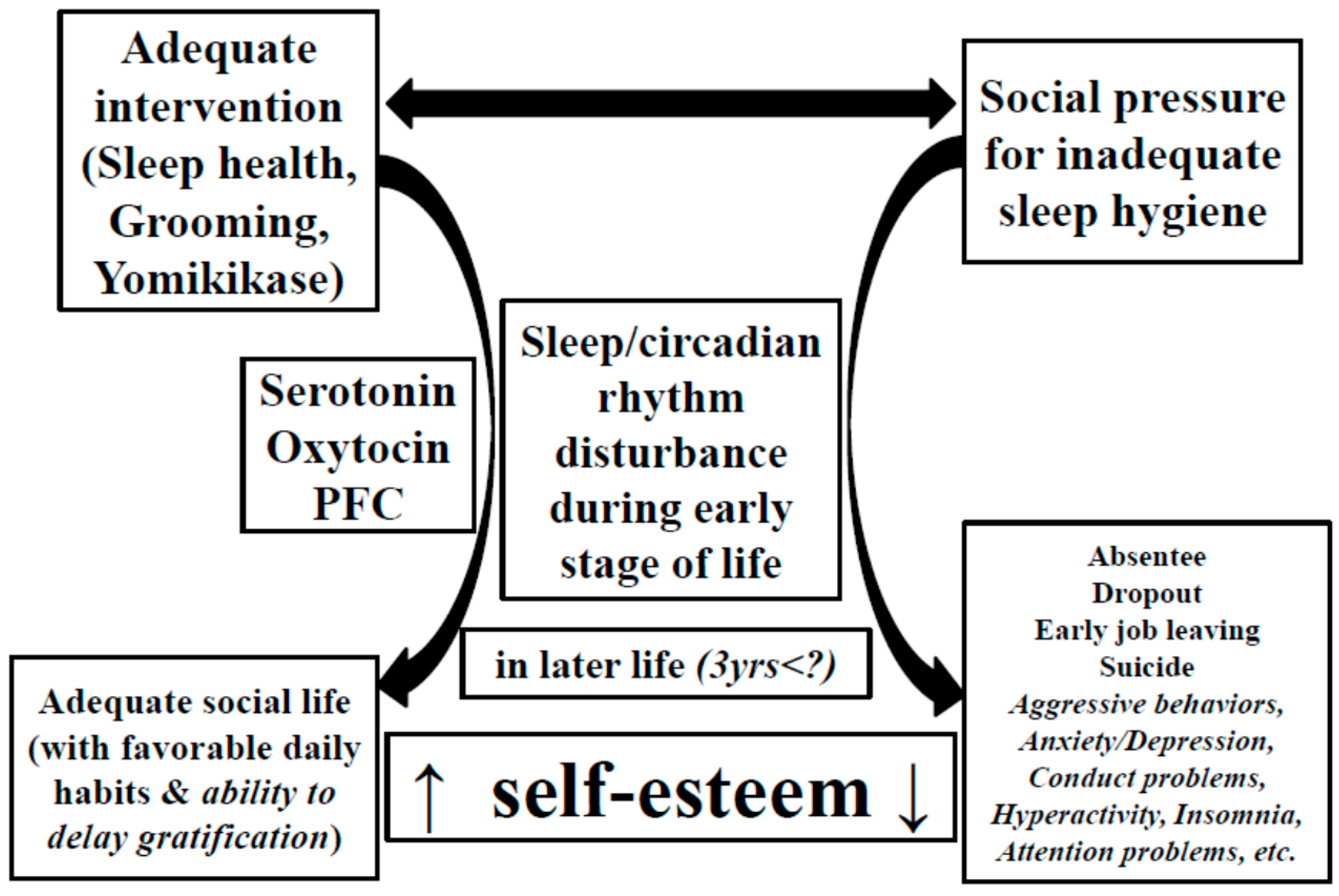
3. Sleep Deterioration during the Early Stage of Life Has a Long-Term Negative Impact on Brain Function
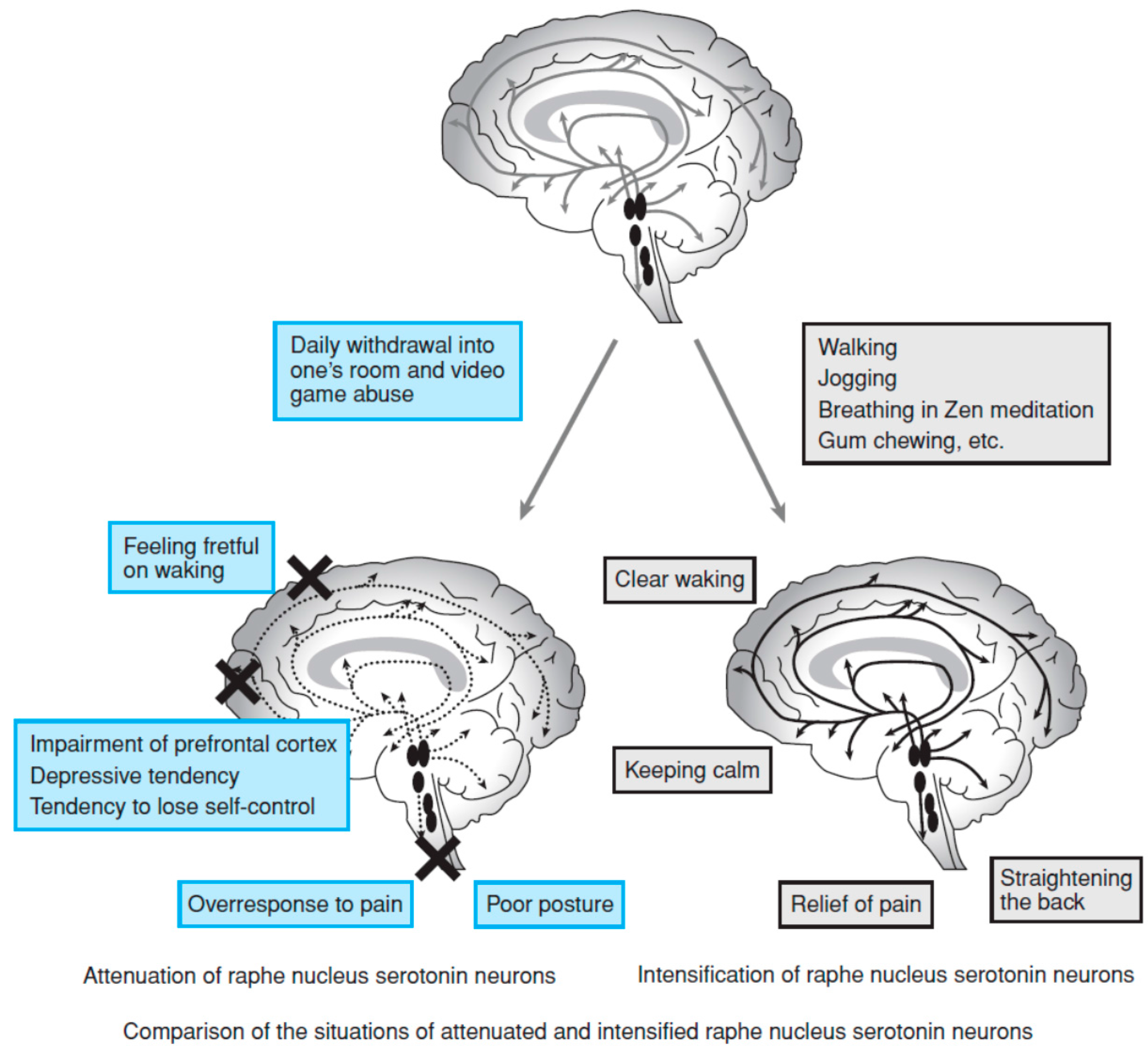
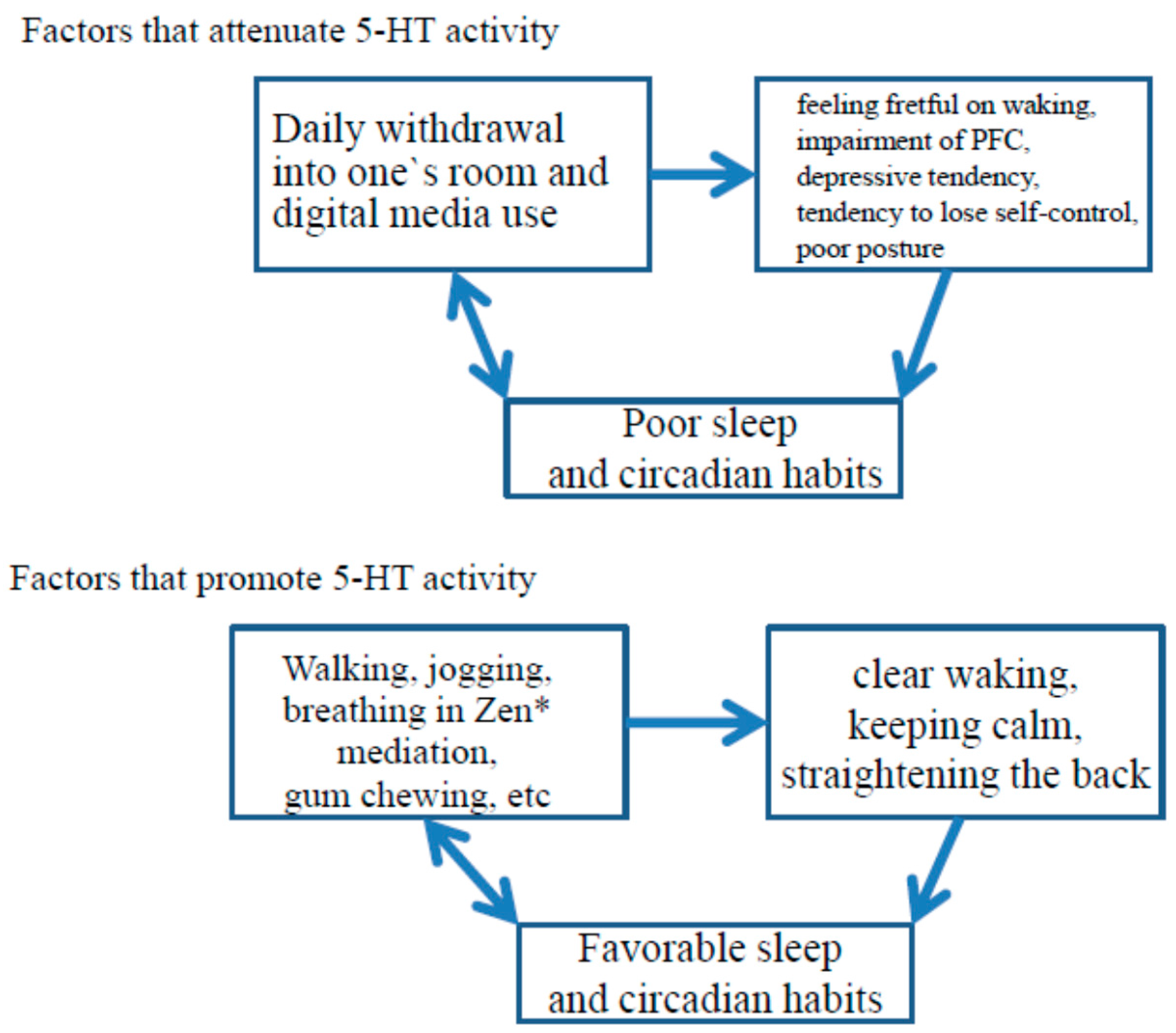
4. Theories on the Development of the 5-HT System


{kind=link}
{kind=link}
{kind=link}
{kind=link}
5. Marshmallow Test and Possible Involvement of the 5-HT System
6. 5-HT Activity, Sleep, and Development
7. Conclusions

Conflicts of Interest
References
- Mindell, J.A.; Owens, J.; Alves, R.; Bruni, O.; Goh, D.Y.; Hiscock, H.; Kohyama, J.; Sadeh, A. Give children and adolescents the gift of a good night’s sleep: A call to action. Sleep Med. 2011, 12, 203–204. [Google Scholar] [CrossRef] [PubMed]
- Yokomaku, A.; Misao, K.; Omoto, F.; Yamagishi, R.; Tanaka, K.; Takada, K.; Kohyama, J. A study of the association between sleep habits and problematic behaviors in preschool children. Chronobiol. Int. 2008, 25, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Osonoi, Y.; Mita, T.; Osonoi, T.; Saito, M.; Tamasawa, A.; Nakayama, S.; Someya, Y.; Ishida, H.; Kanazawa, A.; Gosho, M.; et al. Morningness-eveningness questionnaire score and metabolic parameters in patients with type 2 diabetes mellitus. Chronobiol. Int. 2014, 31, 1017–1023. [Google Scholar] [CrossRef] [PubMed]
- Schlarb, A.A.; Sopp, R.; Ambiel, D.; Grünwald, J. Chronotype-related differences in childhood and adolescent aggression and antisocial behavior—A review of the literature. Chronobiol. Int. 2014, 31, 1–16. [Google Scholar]
- Iemura, A.; Iwasaki, M.; Yamakawa, N.; Tomiwa, K.; Anji, Y.; Sakakihara, Y.; Kakuma, T.; Nagamitsu, S.; Matsuishi, T. Influence of sleep-onset time on the development of 18-month-old infants: Japan Children’s cohort study. Brain Dev. 2015. [Google Scholar] [CrossRef] [PubMed]
- Landhuis, C.E.; Poulton, R.; Welch, D.; Hancox, R.J. Childhood sleep time and long-term risk for obesity: A 32-year prospective birth cohort study. Pediatrics 2008, 122, 955–960. [Google Scholar] [CrossRef] [PubMed]
- Kohyama, J. The possible long-term effects of early-life circadian rhythm disturbance on social behavior. Expert Rev. Neurother. 2014, 14, 745–755. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.H.; Wiesel, T.N. The period of susceptibility to the physiological effects of unilateral eye closure in kittens. J. Physiol. 1970, 206, 419–436. [Google Scholar] [CrossRef] [PubMed]
- Frank, M.G.; Issa, N.P.; Stryker, M.P. Sleep enhances plasticity in the developing visual cortex. Neuron 2001, 30, 275–287. [Google Scholar] [CrossRef]
- Kohyama, J. Sleep, serotonin, and suicide in Japan. J. Physiol. Anthropol. 2011, 30, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Haji, T.; Tajima, N.; Matsumoto, M.; Sasaki, T.; Hachisuka, M.; Aihara, M.; Taira, M. Cortical activations of mother and child during reading to child—Yomikikase: An fNIRS study. In Proceedings of the 84th Congress of the Physiological Society of Japan, Osaka, Japan, 20–22 March 2007; The Physiological Society of Japan: Tokyo, Japan, 2007. [Google Scholar]
- Taira, M. Yomikikase ha Kokoro no Nou ni Todoku; Kumon Publisher: Tokyo, Japan, 2009. (In Japanese) [Google Scholar]
- Maret, S.; Faraguna, U.; Nelson, A.B.; Cirelli, C.; Tononi, G. Sleep and waking modulate spine turnover in the adolescent mouse cortex. Nat. Neurosci. 2011, 14, 1418–1420. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Gan, W.B. Sleep contributes to dendritic spine formationand elimination in the developing mouse somatosensory cortex. Dev. Neurobiol. 2012, 72, 1391–1398. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Lai, C.S.; Cichon, J.; Ma, L.; Li, W.; Gan, W.B. Sleep promotes branch-specific formation of dendritic spines after learning. Science 2014, 344, 1173–1178. [Google Scholar] [CrossRef] [PubMed]
- Tononi, G.; Cirelli, C. Sleep and the price of plasticity: From synaptic and cellular homeostasis to memory consolidation and integration. Neuron 2014, 81, 12–34. [Google Scholar] [CrossRef] [PubMed]
- Macey, P.M.; Kumar, R.; Woo, M.A.; Valladares, E.M.; Yan-Go, F.L.; Harper, R.M. Brain structural changes in obstructive sleep apnea. Sleep 2008, 31, 967–977. [Google Scholar] [PubMed]
- Telzer, E.H.; Goldenberg, D.; Fuligni, A.J.; Lieberman, M.D.; Gálvan, A. Sleep variability in adolescence is associated with altered brain development. Dev. Cognit. Neurosci. 2015, 14, 16–22. [Google Scholar] [CrossRef] [PubMed]
- Halbower, A.C.; Degaonkar, M.; Barker, P.B.; Earley, C.J.; Marcus, C.L.; Smith, P.L.; Prahme, M.C.; Mahone, E.M. Childhood obstructive sleep apnea associates with neuropsychological deficits and neuronal brain injury. PLoS Med. 2006, 3, e301. [Google Scholar] [CrossRef] [PubMed]
- Thunström, M. Severe sleep problems in infancy associated with subsequent development of attention deficit/hyperactivity disorder at 5.5 years of age. Acta Paediatr. 2002, 91, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.M.; O’Connor, T.G. Sleep problems in childhood: A longitudinal study of developmental change and association with behavioral problems. J. Am. Acad. Child Adolesc. Psychiatry 2002, 41, 964–971. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.M.; Van der Ende, J.; Willis, T.A.; Verhulst, F.C. Parent-reported sleep problems during development and self-reported anxiety/depression, attention problems, and aggressive behavior later in life. Arch. Pediatr. Adolesc. Med. 2008, 162, 330–335. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.M.; Eley, T.C.; O’Connor, T.G.; Plomin, R. Etiologies of associations between childhood sleep and behavioral problems in a large twin sample. J. Am. Acad. Child Adolesc. Psychiatry 2004, 43, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Yorifuji, T.; Yamakawa, M.; Oka, M.; Inoue, S.; Yoshinaga, H.; Doi, H. Poor toddler-age sleep schedules predict school-age behavioral disorders in a longitudinal survey. Brain Dev. 2015, 37, 572–578. [Google Scholar] [CrossRef] [PubMed]
- The Ministry of Education, Culture, Sports, Science and Technology. Annual report on educational problems including problematic behaviors among pupils and students in 2014 and 2015. Available online: http://www.mext.go.jp/b_menu/houdou/27/09/__icsFiles/afieldfile/2015/10/07/1362012_1_1.pdf (accessed on 28 December 2015). (In Japanese)
- Wang, H.; Sekine, M.; Chen, X.; Yamagami, T.; Kagamimori, S. Lifestyle at 3 years of age and quality of life (QOL) in first-year junior high school students in Japan: Results of the Toyama Birth Cohort Study. Qual. Life Res. 2008, 17, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Sekine, M. Nocturnal sleep and lifestyle disease. Child Health 2003, 6, 662–665. (In Japanese) [Google Scholar]
- Dunedin Multidisciplinary Health and Development Research Unit. Available online: http://dunedinstudy.otago.ac.nz/ (accessed on 28 December 2015).
- Gregory, A.M.; Caspi, A.; Eley, T.C.; Moffitt, T.E.; O’Connor, T.G.; Poulton, R. Prospective longitudinal associations between persistent sleep problems in childhood and anxiety and depression disorders in adulthood. J. Abnorm. Child Psychol. 2005, 33, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.M.; Caspi, A.; Moffitt, T.E.; Poulton, R. Sleep problems in childhood predict neuropsychological functioning in adolescence. Pediatrics 2009, 123, 1171–1176. [Google Scholar] [CrossRef] [PubMed]
- Gregory, A.M.; Caspi, A.; Moffitt, T.E.; Poulton, R. Family conflict in childhood: A predictor of later insomnia. Sleep 2006, 29, 1063–1067. [Google Scholar] [PubMed]
- Caspi, A. The child is father of the man: Personality continuities from childhood to adulthood. J. Pers. Soc. Psychol. 2000, 78, 158–172. [Google Scholar] [CrossRef] [PubMed]
- Chess, S.; Thomas, A. Origins and Evolution of Behavior Disorders: From Infancy to Early Adult Life; Harvard University Press: Cambridge, MA, USA, 1987. [Google Scholar]
- Greene, G.; Gregory, A.M.; Fone, D.; White, J. Childhood sleeping difficulties and depression in adulthood: The 1970 British Cohort Study. J. Sleep Res. 2015, 24, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Maher, M.J.; Rego, S.A.; Asnis, G.M. Sleep disturbances in patients with post-traumatic stress disorder: Epidemiology, impact and approaches to management. CNS Drugs 2006, 20, 567–590. [Google Scholar] [CrossRef] [PubMed]
- Nautiyal, K.M.; Tanaka, K.F.; Barr, M.M.; Tritschler, L.; le Dantec, Y.; David, D.J.; Gardier, A.N.; Blanco, C.; Hen, R.; Ahmari, S.E. Distinct circuits underlie the effects of 5-HT1B receptors on aggression and impulsivity. Neuron 2015, 86, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Levitt, P.; Harvey, J.A.; Friedman, E.; Simansky, K.; Murphy, E.H. New evidence for neurotransmitter influences on brain development. Trends Neurosci. 1997, 20, 269–274. [Google Scholar] [CrossRef]
- Gaspar, P.; Cases, O.; Maroteaux, L. The developmental role of serotonin: News from mouse molecular genetics. Nat. Rev. Neurosci. 2003, 4, 1002–1012. [Google Scholar] [CrossRef] [PubMed]
- Koh, T.; Nakazawa, M.; Kani, K.; Maeda, T. Investigation of origins of serotonergic projection to developing rat visual cortex: A combined retrograde tracing and immunohistochemical study. Brain Res. Bull. 1991, 27, 675–684. [Google Scholar] [CrossRef]
- Segawa, M. Epochs of development of the sleep-wake cycle reflect the modulation of the higher cortical function particular for each epoch. Sleep Biol. Rhythms 2006, 4, 4–15. [Google Scholar] [CrossRef]
- Arita, H. Brain mechanisms of poor anger management. JMAJ 2009, 52, 184–190. [Google Scholar]
- Young, S.N. How to increase serotonin in the human brain without drugs. J. Psychiatry Neurosci. 2007, 32, 394–399. [Google Scholar] [PubMed]
- Jacobs, B.L.; Azmitia, E.C. Structure and function of the brain serotonin system. Physiol. Rev. 1992, 72, 165–229. [Google Scholar] [PubMed]
- Cagampang, F.R.; Yamazaki, S.; Otori, Y.; Inouye, S.I. Serotonin in the raphe nuclei: Regulation by light and an endogenous pacemaker. Neuroreport 1993, 5, 49–52. [Google Scholar] [CrossRef] [PubMed]
- American Academy of Sleep Medicine. The International Classification of Sleep Disorder, 3rd ed.; American Academy of Sleep Medicine: Westchester, NY, USA, 2014. [Google Scholar]
- Kohyama, J.; Shiiki, T.; Ohinata-Sugimoto, J.; Hasegawa, T. Potentially harmful sleep habits of 3-year-old children in Japan. J. Dev. Behav. Pediatr. 2002, 23, 67–70. [Google Scholar] [CrossRef] [PubMed]
- Mischel, W. The Marshmallow Test; Little, Brown and Company: New York, NY, USA, 2014. [Google Scholar]
- Mischel, W. Father-absence and delay of gratification: Crosscultural comparisons. J. Abnorm. Soc. Psychol. 1961, 63, 116–124. [Google Scholar] [CrossRef] [PubMed]
- Mischel, W.; Ayduk, O.; Berman, M.G.; Casey, B.J.; Gotlib, I.H.; Jonides, J.; Kross, E.; Teslovich, T.; Wilson, N.L.; Zayas, V.; et al. “Willpower” over the life span: Decomposing self-regulation. Soc. Cogn. Affect. Neurosci. 2011, 6, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Mischel, W.; Shoda, Y.; Rodriguez, M.I. Delay of gratification in children. Science 1989, 244, 933–938. [Google Scholar] [CrossRef] [PubMed]
- Casey, B.J.; Somerville, L.H.; Gotlib, I.H.; Ayduk, O.; Franklin, N.T.; Askren, M.K.; Jonides, J.; Berman, M.G.; Wilson, N.L.; Teslovich, T.; et al. Behavioral and neural correlates of delay of gratification 40 years later. Proc. Natl. Acad. Sci. USA 2011, 108, 14998–15003. [Google Scholar] [CrossRef] [PubMed]
- Ma, N.; Dinges, D.F.; Basner, M.; Rao, H. How acute total sleep loss affects the attending brain: A meta-analysis of neuroimaging studies. Sleep 2015, 38, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Frith, C.; Frith, U. Theory of mind. Curr. Biol. 2005, 15, R644–R645. [Google Scholar] [CrossRef] [PubMed]
- Hikosaka, O.; Bromberg-Martin, E.; Hong, S.; Matsumoto, M. New insights on the subcortical representation of reward. Curr. Opin. Neurobiol. 2008, 18, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Sescousse, G.; Caldú, X.; Segura, B.; Dreher, J.C. Processing of primary and secondary rewards: A quantitative meta-analysis and review of human functional neuroimaging studies. Neurosci. Biobehav. Rev. 2013, 37, 681–696. [Google Scholar] [CrossRef] [PubMed]
- Salamone, J.D.; Pardo, M.; Yohn, S.E.; López-Cruz, L.; SanMiguel, N.; Correa, M. Mesolimbic dopamine and the regulation of motivated behavior. Curr. Top. Behav. Neurosci. 2015. [Google Scholar] [CrossRef]
- Seo, D.; Patrick, C.J.; Kennealy, P.J. Role of serotonin and dopamine system interactions in the neurobiology of impulsive aggression and its comorbidity with other clinical disorders. Aggress. Violent Behav. 2008, 13, 383–395. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.C.; Schweighofer, N.; Asahi, S.; Shishida, K.; Okamoto, Y.; Yamawaki, S.; Doya, K. Serotonin differentially regulates short- and long-term prediction of rewards in the ventral and dorsal striatum. PLoS ONE 2007, 2, e1333. [Google Scholar] [CrossRef] [PubMed]
- Crean, J.; Richards, J.B.; de Wit, H. Effect of tryptophan depletion on impulsive behavior in men with or without a family history of alcoholism. Behav. Brain Res. 2002, 136, 349–357. [Google Scholar] [CrossRef]
- Schweighofer, N.; Bertin, M.; Shishida, K.; Okamoto, Y.; Tanaka, S.C.; Yamawaki, S.; Doya, K. Low-serotonin levels increase delayed reward discounting in humans. J. Neurosci. 2008, 28, 4528–4532. [Google Scholar] [CrossRef] [PubMed]
- Bilderbeck, A.C.; Brown, G.D.; Read, J.; Woolrich, M.; Cowen, P.J.; Behrens, T.E.; Rogers, R.D. Serotonin and social norms: Tryptophan depletion impairs social comparison and leads to resource depletion in a multiplayer harvesting game. Psychol. Sci. 2014, 25, 1303–1313. [Google Scholar] [CrossRef] [PubMed]
- Chiavegatto, S.; Quadros, I.M.; Ambar, G.; Miczek, K.A. Individual vulnerability to escalated aggressive behavior by a low dose of alcohol: Decreased serotonin receptor mRNA in the prefrontal cortex of male mice. Genes Brain Behav. 2010, 9, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Kohyama, J. Sleep, Serotonin, and Suicide. J. Behav. Brain Sci. 2012, 2, 471–478. [Google Scholar] [CrossRef]
- Pigeon, W.R.; Pinquart, M.; Conner, K. Meta-analysis of sleep disturbance and suicidal thoughts and behavior. J. Clin. Psychiatry 2012, 73, e1160–e1167. [Google Scholar] [CrossRef] [PubMed]
- Larsen, B.; Luna, B. In vivo evidence of neurophysiological maturation of the human adolescent striatum. Dev. Cogn. Neurosci. 2015, 12, 74–85. [Google Scholar] [CrossRef] [PubMed]
© 2016 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kohyama, J. Neural Basis of Brain Dysfunction Produced by Early Sleep Problems. Brain Sci. 2016, 6, 5. https://doi.org/10.3390/brainsci6010005
Kohyama J. Neural Basis of Brain Dysfunction Produced by Early Sleep Problems. Brain Sciences. 2016; 6(1):5. https://doi.org/10.3390/brainsci6010005
Chicago/Turabian StyleKohyama, Jun. 2016. "Neural Basis of Brain Dysfunction Produced by Early Sleep Problems" Brain Sciences 6, no. 1: 5. https://doi.org/10.3390/brainsci6010005