1. Introduction
Huntington’s disease (HD) is an inherited neurodegenerative disorder in which behavioral signs, mainly characterized by involuntary movements and cognitive decline, typically emerge in adulthood [
1]. Ample evidence indicates that impaired communication between cortical and striatal networks is a leading indicator of the HD behavioral phenotype [
2,
3]. Neurons in striatum are especially vulnerable to mutant huntingtin (mHTT), the protein underlying HD [
4,
5]. Among the changes induced by mHTT is a deficit in protein palmitoylation by huntingtin interacting protein 14 (HIP14), a palmitoyl acyl transferase (PAT) that binds palmitic acid [
6,
7,
8]. Failure to palmitoylate key proteins can impair synaptic transmission, which may account for the altered response of striatal neurons to cortical activation in HD mouse models [
9]. Consistent with a role for HIP14 in HD, mice lacking HIP14 develop striatal neuropathology and motor deficits similar to those described for transgenic HD models [
10].
When transgenic HD mice are allowed to explore a plus-shaped maze, they are less likely than wild-type controls to turn into the left or right arm of the maze and continue moving instead into the opposite arm, a sign of motor inflexibility that also occurs in HD patients [
11,
12,
13]. We reported similar behavior in HIP14 knockouts, which was accompanied by abnormal firing patterns in individually isolated striatal neurons [
14]. At the choice point or center of the maze, these neurons, relative to those recorded from wild-type mice, change both the rate and pattern of spike activity. Firing rate increases as does the number of spikes that cluster together in bursts. Collectively, these results, which implicate HIP14 in striatal function, suggest that aberrant striatal processing may underlie the motor inflexibility seen in HIP14 knockout mice. Here, we extended this investigation to the population response of striatal neurons by analyzing local field potentials (LFPs) in conjunction with entering and exiting the choice point. A force-plate actometer monitored position in the maze and provided time-stamps for LFP activity.
2. Results and Discussion
When mice enter the center or choice point of the plus maze, they have the option of continuing straight into the opposite arm or turning into either the left or right arm [
11]. Wild-type (
n = 10) and HIP14 knockout mice (
n = 10) participated in multiple recording sessions; each mouse participated in at least two sessions for a total of 38 and 46 sessions, respectively, for each group. As
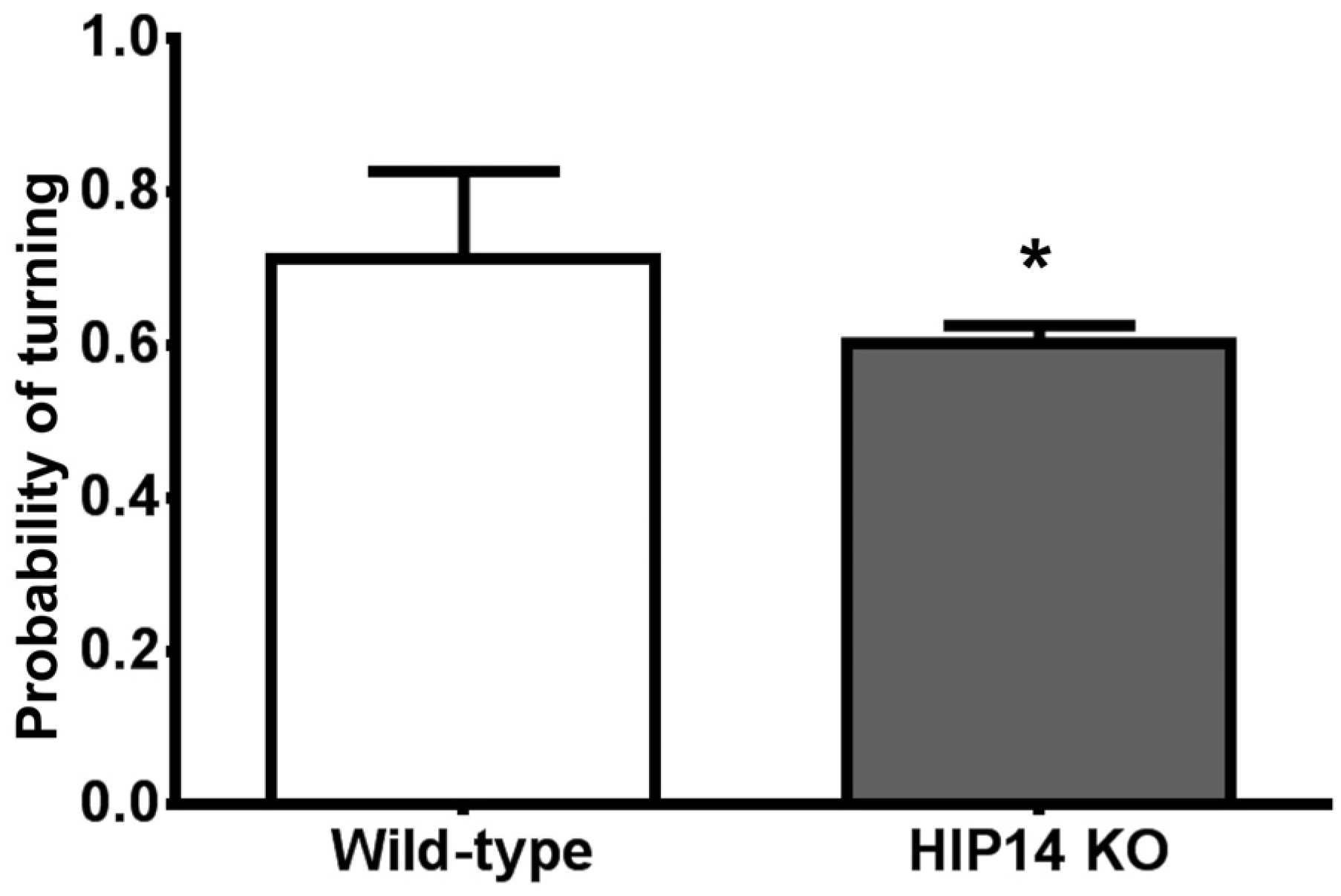
Figure 1 shows, wild-type mice turn to explore the perpendicular arms with a probability of 0.71 ± 0.01. In contrast, HIP14 knockout mice are less likely to turn (the probability of turning is 0.60 ± 0.02); instead, once they cross the choice point, they explore the opposite arm. Statistical analysis of the probability of turning revealed a significant difference between wild-type and HIP14 knockout mice (
t = 3.6, degrees of freedom (df) = 82,
p = 0.0005).
The reduced probability of turning in HIP14 knockout mice is not due to the fact that they were less active than wild-type. In fact, relative to wild-type, HIP14 knockouts were more active when exploring the plus maze, as evidenced by the total number of arm choices observed: wild-type = 69.11 ± 4.6; HIP14 knockouts = 150.9 ± 11.37 (
t = 6.19, df = 82;
p < 0.0001). Thus, although HIP14 knockouts are more active in the plus maze than wild-types, the HIP14 knockouts turn less, a common sign of motor inflexibility [
12].
Figure 1.
HIP14 knockouts (HIP14 KO) are less likely to turn left or right as they explore the plus-shaped maze. The graph shows the turning probability obtained after wild-type and HIP14 knockout mice freely explored the plus maze for 30 min. The probability was obtained from the sum of arm choices to the right or left arm divided by the total number of arm choices. Wild-type and HIP14 knockout mice participated in multiple sessions for a total of 38 and 46 sessions, respectively. Data were analyzed by the unpaired t-test. Data are expressed as the mean ± SEM. t = 3.6; df = 82; * p = 0.0001.
Figure 1.
HIP14 knockouts (HIP14 KO) are less likely to turn left or right as they explore the plus-shaped maze. The graph shows the turning probability obtained after wild-type and HIP14 knockout mice freely explored the plus maze for 30 min. The probability was obtained from the sum of arm choices to the right or left arm divided by the total number of arm choices. Wild-type and HIP14 knockout mice participated in multiple sessions for a total of 38 and 46 sessions, respectively. Data were analyzed by the unpaired t-test. Data are expressed as the mean ± SEM. t = 3.6; df = 82; * p = 0.0001.
Because evaluating patterns of arm choice provides information on behavioral flexibility, we focused our analysis of striatal LFP activity on events surrounding the choice point. We first constructed spectrograms based on total entries into the choice point obtained from five randomly selected wild-type (339 entries) and six HIP14 knockout mice (701 entries). As shown in
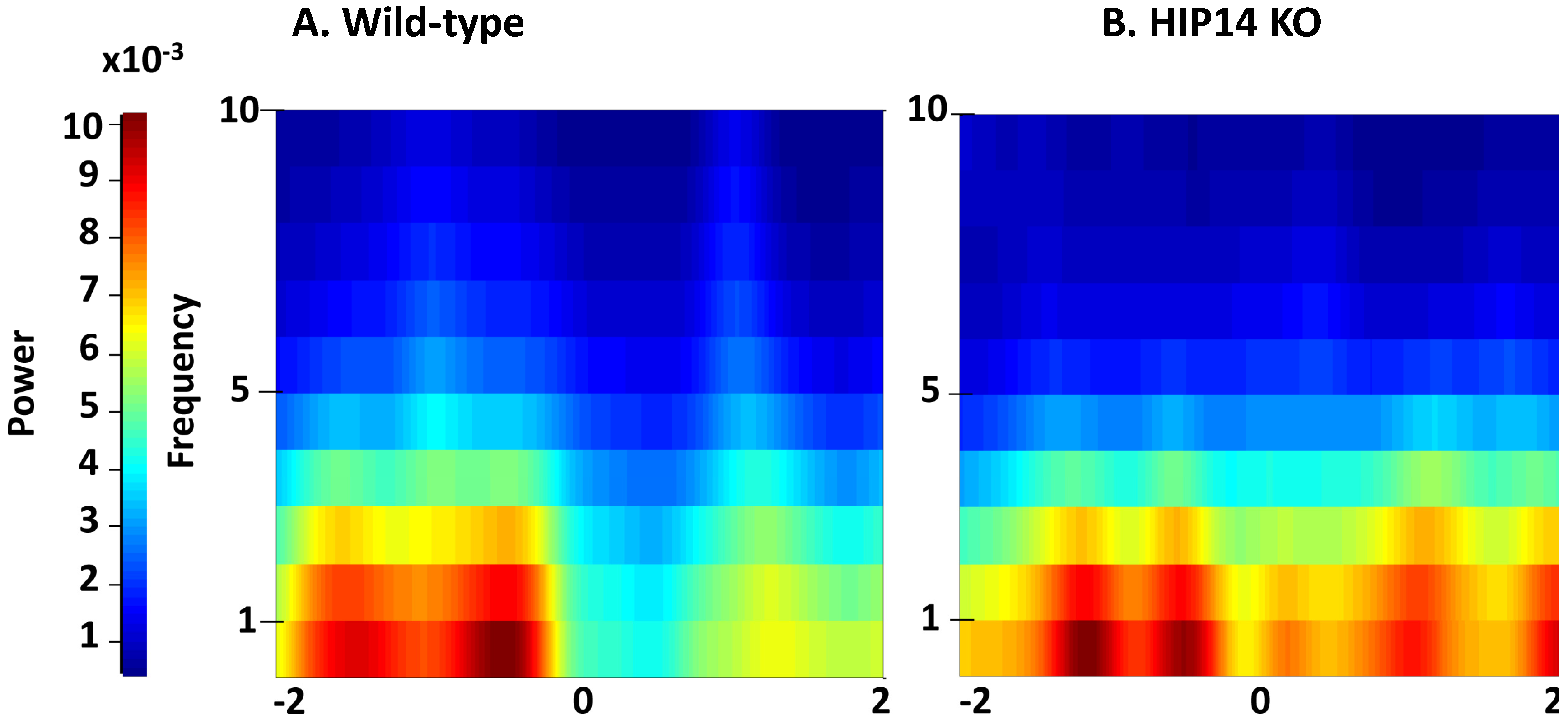
Figure 2, two seconds before entering the choice point (denoted by zero), both groups showed high power at low frequencies [delta (0.1–4 Hz), theta (4–7 Hz)]. Note, however, that when the mice cross the choice point, LFP power decreases only in wild-types; HIP14 knockouts maintain the same level of power at low frequencies.
Next, we compared choice-related LFP activity with LFP activity when the animal had subsequently moved into an arm. For this comparison, we analyzed the second immediately before choice-point entry with the second after the mouse entered an arm. In order to make the LFP comparison comparable between groups, since HIP14 knockout mice crossed the center of the maze more often than wild-type animals, we analyzed a total of 50 randomly selected choice and arm events from each mouse (regardless of the direction of the turn). Mean power spectral density (PSD) data are shown in
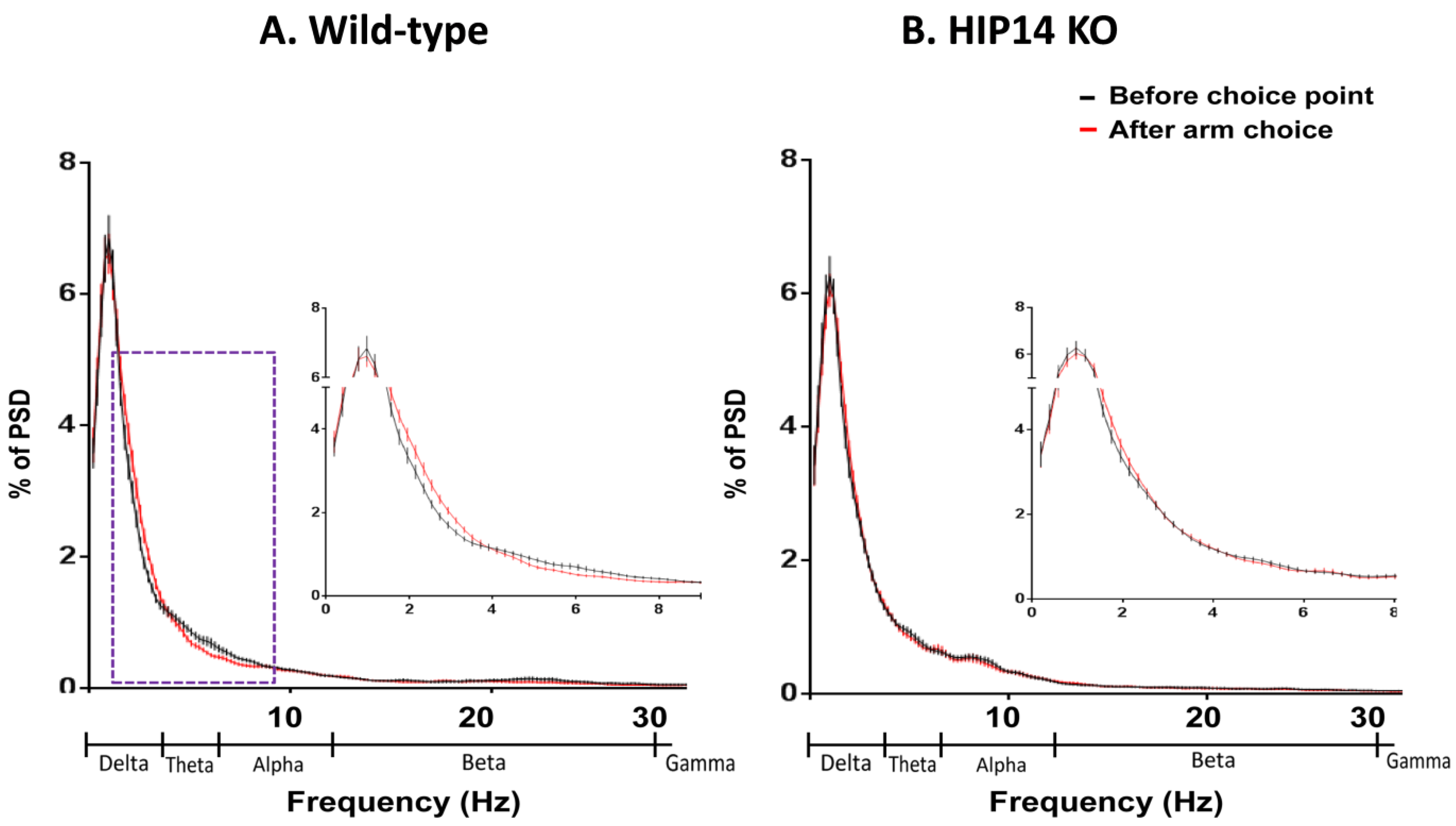
Figure 3. LFP activity in the arm was significantly decreased relative to choice-point activity in wild-type mice (
p ≤ 0.05), at delta (0.1–4 Hz), while theta (4–7 Hz) bands activity increased (
Figure 3A). Whereas HIP14 knockouts showed no change in power as they moved from choice point to arm (
Figure 3B). Thus, protein palmitoylation by HIP14 might be involved in behavioral flexibility in arm choice and for the differential responsiveness of striatal LFP activity to position in the plus maze.
Figure 2.
Striatal local field potential (LFP) activity recorded from wild-type and HIP14 knockout mice before and after entry into the choice point. Spectrogram plots were obtained 2 s before and 2 s after the wild-type (A) and HIP14 knockout (B) mice enter the center of the maze (denoted by zero). Graphs were obtained from the average of the total of crossing events obtained from 5 wild-type (339 events) and 6 HIP14 knockout mice (701 events).
Figure 2.
Striatal local field potential (LFP) activity recorded from wild-type and HIP14 knockout mice before and after entry into the choice point. Spectrogram plots were obtained 2 s before and 2 s after the wild-type (A) and HIP14 knockout (B) mice enter the center of the maze (denoted by zero). Graphs were obtained from the average of the total of crossing events obtained from 5 wild-type (339 events) and 6 HIP14 knockout mice (701 events).
Figure 3.
Mean power spectral density (PSD) plotted as a proportion of total power (% PSD) for wild-type (A) and HIP14 knockout (B) mice immediately before choice-point entry (black) and after arm entry (red). For comparison purposes, 50 random events of a one-second duration for each recording session from each mouse (43 sessions for wild-types and 44 for HIP14 knockouts) were measured. The dashed box denotes the frequencies at which the mean PSDs significantly differ. Insets provide a close up of activity at frequencies <8 Hz. PSD data were obtained from 50 random events per recording session. Data are expressed as the mean ± SEM (p ≤ 0.05).
Figure 3.
Mean power spectral density (PSD) plotted as a proportion of total power (% PSD) for wild-type (A) and HIP14 knockout (B) mice immediately before choice-point entry (black) and after arm entry (red). For comparison purposes, 50 random events of a one-second duration for each recording session from each mouse (43 sessions for wild-types and 44 for HIP14 knockouts) were measured. The dashed box denotes the frequencies at which the mean PSDs significantly differ. Insets provide a close up of activity at frequencies <8 Hz. PSD data were obtained from 50 random events per recording session. Data are expressed as the mean ± SEM (p ≤ 0.05).
HIP14 knockout mice are less likely to turn left or right when they navigate the plus maze (
Figure 1). Instead, they typically continue moving, straight into the opposite arm; and, when they eventually make a left or right turn, they again persist in exploring the opposite arm. This behavior is a sign of motor inflexibility, which refers to a decreased likelihood of alternating arm entries [
12]. Given that the striatum constitutes part of the neuronal circuitry that shapes motor control, our data suggest that a change in low frequency striatal LFP activity plays a critical role in shaping behavioral choice.
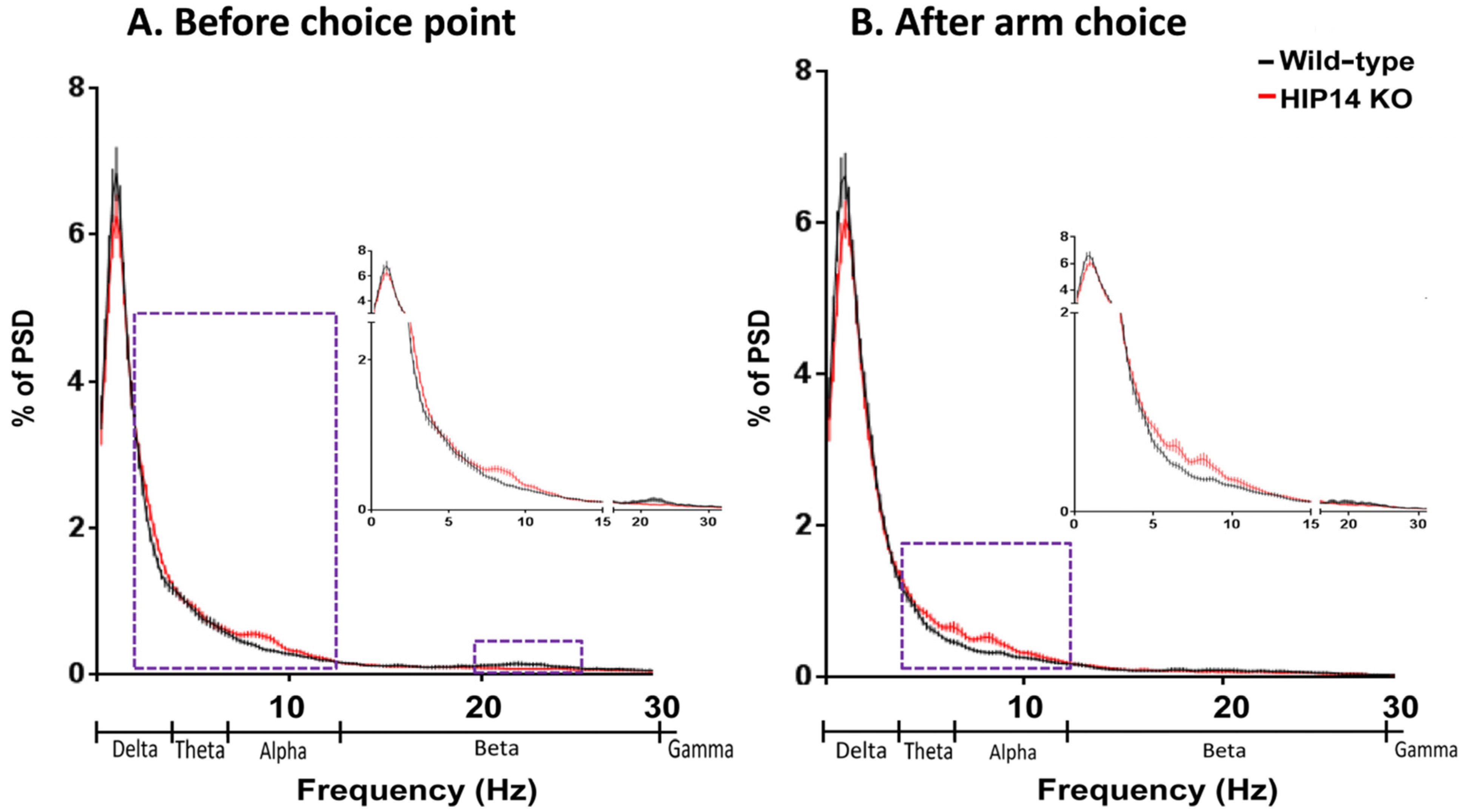
As shown in
Figure 4, we also compared choice-related and arm-related LFP activity between wild-type and HIP14 knockout mice. Striatal LFPs in HIP14 knockout mice relative to wild-type show significant increases in delta (0.1–4 Hz) and alpha (8–13 Hz) frequencies and decreases in beta (13–30 Hz) immediately before the choice point, but increases in the theta (4–7 Hz) and alpha (8–13 Hz) range after arm entry.
Figure 4.
Striatal PSD immediately before the choice point (A) and after arm entry (B) for wild-type (black) and HIP14 knockout mice (red). In each case, data are based on one second of LFP activity obtained from 50 random events per recording session (43 wild-type and 44 HIP14-knockout sessions). Dashed boxes denote the frequencies at which the mean PSD significantly differ. Data are expressed as the mean ± SEM (p ≤ 0.05). Insert: close up of frequencies at which differences were observed.
Figure 4.
Striatal PSD immediately before the choice point (A) and after arm entry (B) for wild-type (black) and HIP14 knockout mice (red). In each case, data are based on one second of LFP activity obtained from 50 random events per recording session (43 wild-type and 44 HIP14-knockout sessions). Dashed boxes denote the frequencies at which the mean PSD significantly differ. Data are expressed as the mean ± SEM (p ≤ 0.05). Insert: close up of frequencies at which differences were observed.
Overall, our analysis indicates that when HIP14 knockout mice navigate the plus maze, striatal LFPs maintain an increase in power at lower frequencies, both before and after the choice point, which may contribute to behavioral inflexibility. A summary of averaged striatal LFPs at relevant frequencies is provided in
Table 1 for both wild-type and HIP14 knockout mice.
Table 1.
Average of % of power spectral density in wild-type and HIP14 knockout mice.
Table 1.
Average of % of power spectral density in wild-type and HIP14 knockout mice.
| Frequency | Wild-type | HIP14 KO |
|---|
| Before | After | Before | After |
|---|
| Delta (0.1–4 Hz) | 143.7 ± 0.08 * | 152.8 ± 0.07 | 149.1 ± 0.07 | 151.1 ± 0.07 |
| Theta (4–7 Hz) | 33.4 ± 0.04 * | 27.8 ± 0.03 † | 35.9 ± 0.03 | 35.0 ± 0.03 |
| Alpha (8–13 Hz) | 10.8 ± 0.01 *,† | 13.3 ± 0.01 † | 14.5 ± 0.02 * | 18.4 ± 0.02 |
| Beta (13–30 Hz) | 4.1 ± 0.01 | 3.4 ± 0.009 | 3.4 ± 0.008 | 3.3 ± 0.006 |
| Gamma (30–50 Hz) | 1.0 ± 0.003 | 0.9 ± 0.003 | 0.9 ± 0.002 | 0.9 ± 0.002 |
Our results suggest that alerted striatal dynamics in HIP14 knockouts may underlie the behavioral signs of motor inflexibility observed in arm-choice selection in the plus maze. Along with altered LFP, we previously reported that striatal neurons in HIP14 knockout mice show an increased firing rate and reduced synchronized activity at the choice point of the maze [
14]. HIP14 knockouts also show deficits in nest-building behavior, a test that provides information on motor coordination and cognitive function [
10,
14,
15]. Thus, HIP14 function, which includes protein palmitoylation, plays a critical role in striatal processing and appears necessary for normal motor output. More studies are necessary to understand the potential mechanisms (e.g., kynurenine metabolites interacting with the
N-methyl-
d-aspartate NMDA receptor) underlying impaired medium spiny neuron (MSN) processing and altered neuronal dynamics in mice lacking the HIP14 protein.






{kind=link}
{kind=link}
{kind=link}
{kind=link}