

Sex-Specific Differences and the Role of Environmental Enrichment in the Expression of Hippocampal CB1 Receptors following Chronic Unpredictable Stress
 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Manipulations
2.3. Tissue Processing
2.3.1. Immunohistochemical Analysis
2.3.2. Western Blot (WB) Analysis
2.4. Statistical Analysis
3. Results
3.1. Analysis of CB1 Expression in DG, CA3 and CA1 by Immunohistochemistry
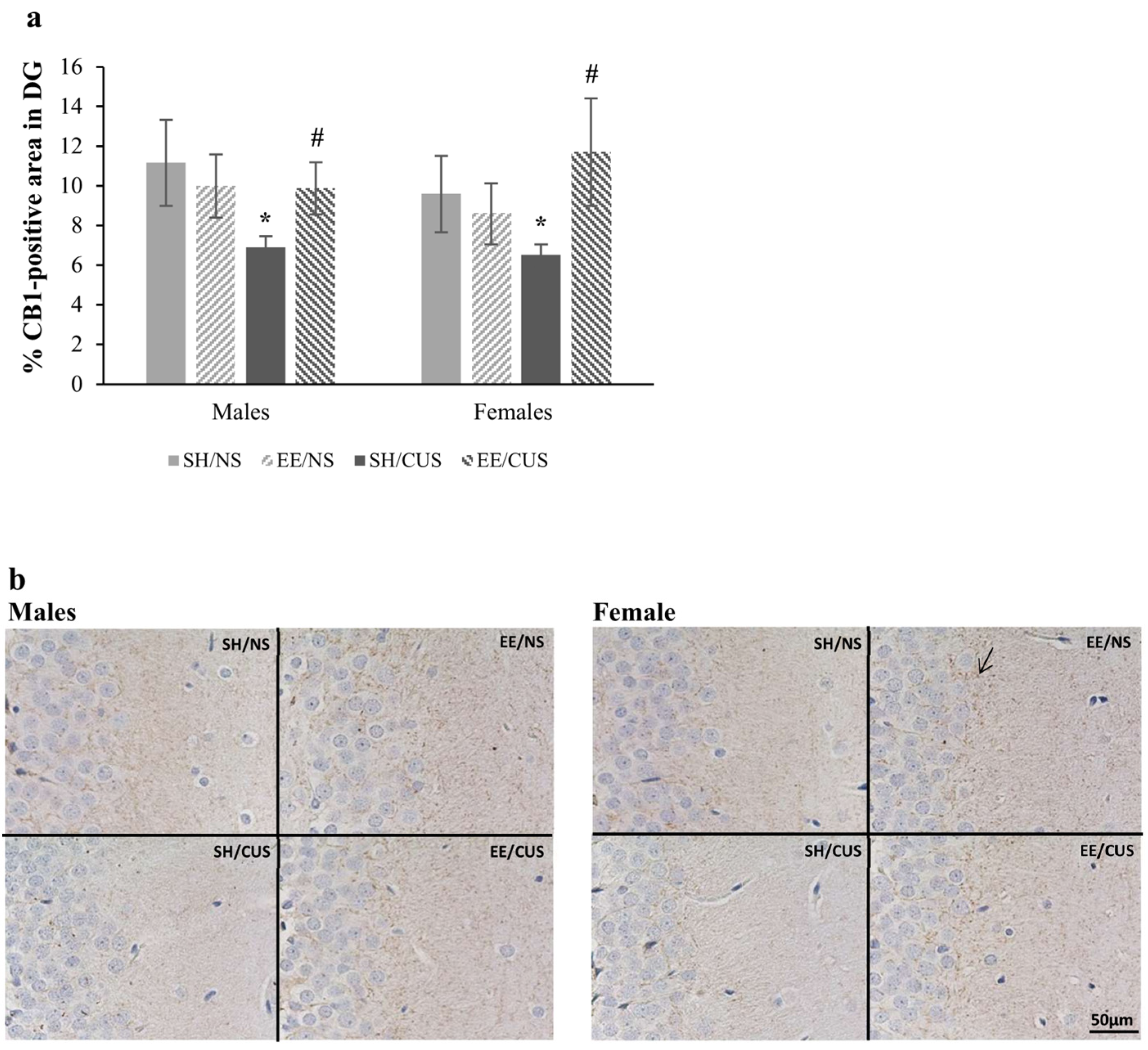
3.1.1. EE Exhibited a Protective Effect against the Stress-Induced Reductions in the Percentage of CB1-Positive Area in DG
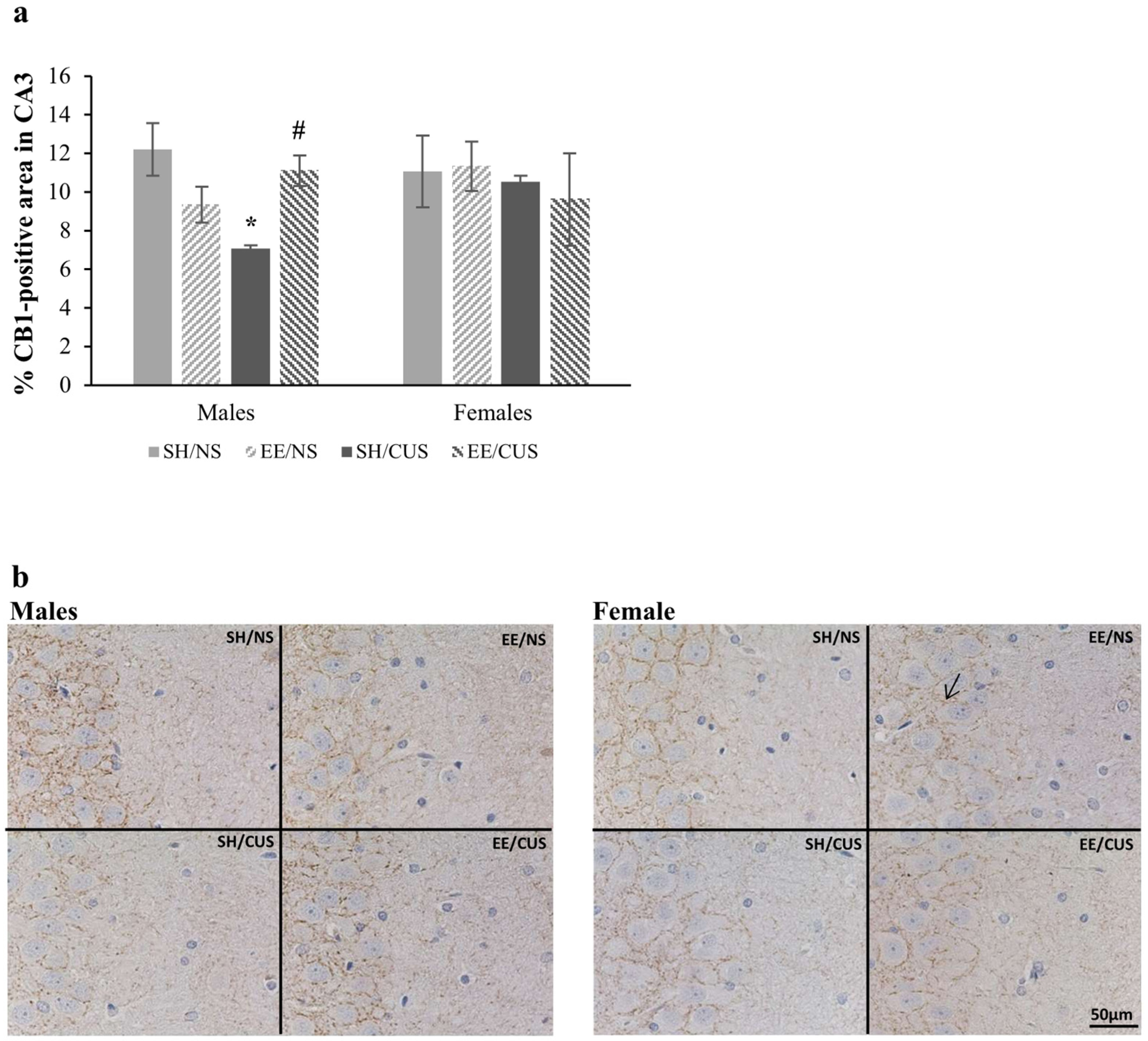
3.1.2. EE Exhibited a Protective Effect against the Stress-Induced Reductions in the Percentage of CB1-Positive Area in CA3 of Male Rats
3.1.3. EE Exhibited a Protective Effect against the Stress-Induced Reductions in the Percentage of CB1-Positive Area in CA1
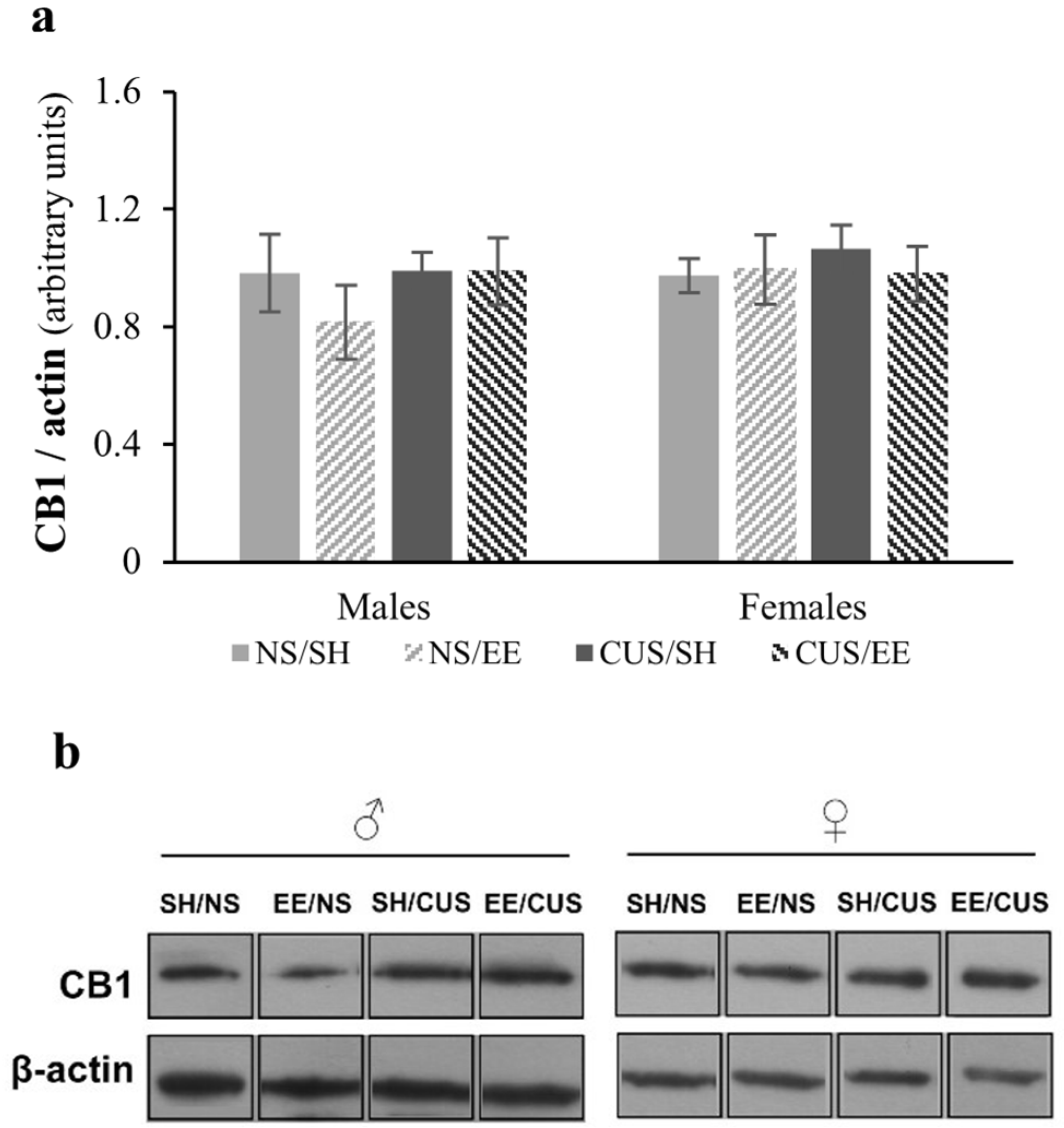
3.2. Analysis of CB1 Total Protein Expression in the Hippocampus Using Western Blotting
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Knezevic, E.; Nenic, K.; Milanovic, V.; Knezevic, N.N. The Role of Cortisol in Chronic Stress, Neurodegenerative Diseases, and Psychological Disorders. Cells 2023, 12, 2726. [Google Scholar] [CrossRef] [PubMed]
- Milligan Armstrong, A.; Porter, T.; Quek, H.; White, A.; Haynes, J.; Jackaman, C.; Villemagne, V.; Munyard, K.; Laws, S.M.; Verdile, G.; et al. Chronic Stress and Alzheimer’s Disease: The Interplay between the Hypothalamic–Pituitary–Adrenal Axis, Genetics and Microglia. Biol. Rev. 2021, 96, 2209–2228. [Google Scholar] [CrossRef] [PubMed]
- Soares, N.M.; Pereira, G.M.; Altmann, V.; de Almeida, R.M.M.; Rieder, C.R.M. Cortisol Levels, Motor, Cognitive and Behavioral Symptoms in Parkinson’s Disease: A Systematic Review. J. Neural Transm. 2019, 126, 219–232. [Google Scholar] [CrossRef] [PubMed]
- Du, X.; Pang, T.Y. Is Dysregulation of the HPA-Axis a Core Pathophysiology Mediating Co-Morbid Depression in Neurodegenerative Diseases? Front. Psychiatry 2015, 6, 123351. [Google Scholar] [CrossRef] [PubMed]
- Pietrzak, R.H.; Laws, S.M.; Lim, Y.Y.; Bender, S.J.; Porter, T.; Doecke, J.; Ames, D.; Fowler, C.; Masters, C.L.; Milicic, L.; et al. Plasma Cortisol, Brain Amyloid-β, and Cognitive Decline in Preclinical Alzheimer’s Disease: A 6-Year Prospective Cohort Study. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2017, 2, 45–52. [Google Scholar] [CrossRef]
- Willner, P. Reliability of the Chronic Mild Stress Model of Depression: A User Survey. Neurobiol. Stress 2017, 6, 68–77. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Tao, Y.; Wang, T.; Zhou, J.; Yang, Y.; Cheng, L.; Zhu, H.; Zhang, W.; Huang, F.; Wu, X. Long-Term Stability and Characteristics of Behavioral, Biochemical, and Molecular Markers of Three Different Rodent Models for Depression. Brain Behav. 2019, 10, e01508. [Google Scholar] [CrossRef] [PubMed]
- Micale, V.; Drago, F. Endocannabinoid System, Stress and HPA Axis. Eur. J. Pharmacol. 2018, 834, 230–239. [Google Scholar] [CrossRef]
- Kilaru, A.; Chapma, K.D. The Endocannabinoid System. Essays Biochem. 2020, 64, 485–499. [Google Scholar]
- Lu, H.C.; MacKie, K. An Introduction to the Endogenous Cannabinoid System. Biol. Psychiatry 2016, 79, 516–525. [Google Scholar] [CrossRef]
- Woodhams, S.G.; Chapman, V.; Finn, D.P.; Hohmann, A.G.; Neugebauer, V. The Cannabinoid System and Pain. Neuropharmacology 2017, 124, 105–120. [Google Scholar] [CrossRef]
- Hill, M.N.; McEwen, B.S. Involvement of the Endocannabinoid System in the Neurobehavioural Effects of Stress and Glucocorticoids. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2010, 34, 791–797. [Google Scholar] [CrossRef] [PubMed]
- Dhopeshwarkar, A.; Mackie, K. CB2cannabinoid Receptors as a Therapeutic Target-What Does the Future Hold? Mol. Pharmacol. 2014, 86, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Turcotte, C.; Blanchet, M.R.; Laviolette, M.; Flamand, N. The CB2 Receptor and Its Role as a Regulator of Inflammation. Cell. Mol. Life Sci. 2016, 73, 4449–4470. [Google Scholar] [CrossRef]
- Winters, B.L.; Vaughan, C.W. Mechanisms of Endocannabinoid Control of Synaptic Plasticity. Neuropharmacology 2021, 197, 108736. [Google Scholar] [CrossRef] [PubMed]
- Blessing, E.M.; Steenkamp, M.M.; Manzanares, J.; Marmar, C.R. Cannabidiol as a Potential Treatment for Anxiety Disorders. Neurotherapeutics 2015, 12, 825–836. [Google Scholar] [CrossRef] [PubMed]
- Viveros, M.P.; Marco, E.M.; File, S.E. Endocannabinoid System and Stress and Anxiety Responses. Pharmacol. Biochem. Behav. 2005, 81, 331–342. [Google Scholar] [CrossRef] [PubMed]
- Dow-Edwards, D.; Frank, A.; Wade, D.; Weedon, J.; Izenwasser, S. Sexually-Dimorphic Alterations in Cannabinoid Receptor Density Depend upon Prenatal/Early Postnatal History. Neurotoxicol. Teratol. 2016, 58, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Mikulska, J.; Juszczyk, G.; Gawrońska-Grzywacz, M.; Herbet, M. HPA Axis in the Pathomechanism of Depression and Schizophrenia: New Therapeutic Strategies Based on Its Participation. Brain Sci. 2021, 11, 1298. [Google Scholar] [CrossRef]
- Akirav, I. Cannabinoids and Glucocorticoids Modulate Emotional Memory after Stress. Neurosci. Biobehav. Rev. 2013, 37, 2554–2563. [Google Scholar] [CrossRef]
- Hillard, C.J. Stress Regulates Endocannabinoid-CB1 Receptor Signaling. Semin. Immunol. 2014, 26, 380–388. [Google Scholar] [CrossRef] [PubMed]
- Tasker, J.G.; Chen, C.; Fisher, M.O.; Fu, X.; Rainville, J.R.; Weiss, G.L. Endocannabinoid Regulation of Neuroendocrine Systems. Int. Rev. Neurobiol. 2015, 125, 163–201. [Google Scholar] [CrossRef]
- Joshi, N.; Onaivi, E.S. Psychiatric Disorders and Cannabinoid Receptors. In Advances in Experimental Medicine and Biology; Springer: Berlin/Heidelberg, Germany, 2021; Volume 1264, pp. 131–153. [Google Scholar]
- Antoniuk, S.; Bijata, M.; Ponimaskin, E.; Wlodarczyk, J. Chronic Unpredictable Mild Stress for Modeling Depression in Rodents: Meta-Analysis of Model Reliability. Neurosci. Biobehav. Rev. 2019, 99, 101–116. [Google Scholar] [CrossRef] [PubMed]
- D’Aquila, P.S.; Brain, P.; Willner, P. Effects of Chronic Mild Stress on Performance in Behavioural Tests Relevant to Anxiety and Depression. Physiol. Behav. 1994, 56, 861–867. [Google Scholar] [CrossRef]
- Hill, M.N.; Hellemans, K.G.C.; Verma, P.; Gorzalka, B.B.; Weinberg, J. Neurobiology of Chronic Mild Stress: Parallels to Major Depression. Neurosci. Biobehav. Rev. 2012, 36, 2085–2117. [Google Scholar] [CrossRef]
- Dandi, Ε.; Spandou, E.; Dalla, C.; Tata, D. Τhe Neuroprotective Role of Environmental Enrichment against Behavioral, Morphological, Neuroendocrine and Molecular Changes Following Chronic Unpredictable Mild Stress: A Systematic Review. Eur. J. Neurosci. 2023, 58, 3003–3025. [Google Scholar] [CrossRef]
- McLaughlin, R.J.; Gobbi, G. Cannabinoids and Emotionality: A Neuroanatomical Perspective. Neuroscience 2012, 204, 134–144. [Google Scholar] [CrossRef]
- Morena, M.; Patel, S.; Bains, J.S.; Hill, M.N. Neurobiological Interactions Between Stress and the Endocannabinoid System. Neuropsychopharmacology 2016, 41, 80–102. [Google Scholar] [CrossRef]
- Hill, M.N.; Patel, S.; Carrier, E.J.; Rademacher, D.J.; Ormerod, B.K.; Hillard, C.J.; Gorzalka, B.B. Downregulation of Endocannabinoid Signaling in the Hippocampus Following Chronic Unpredictable Stress. Neuropsychopharmacology 2005, 30, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Hill, M.N.; Hunter, R.G.; McEwen, B.S. Chronic Stress Differentially Regulates Cannabinoid CB1 Receptor Binding in Distinct Hippocampal Subfields. Eur. J. Pharmacol. 2009, 614, 66–69. [Google Scholar] [CrossRef] [PubMed]
- Marco, E.M.; Ballesta, J.A.; Irala, C.; Hernández, M.D.; Serrano, M.E.; Mela, V.; López-Gallardo, M.; Viveros, M.P. Sex-Dependent Influence of Chronic Mild Stress (CMS) on Voluntary Alcohol Consumption; Study of Neurobiological Consequences. Pharmacol. Biochem. Behav. 2017, 152, 68–80. [Google Scholar] [CrossRef] [PubMed]
- Reich, C.G.; Taylor, M.E.; McCarthy, M.M. Differential Effects of Chronic Unpredictable Stress on Hippocampal CB1 Receptors in Male and Female Rats. Behav. Brain Res. 2009, 203, 264–269. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.M.; Dias-Rocha, C.P.; Calviño, C.; Trevenzoli, I.H. Lipid Endocannabinoids in Energy Metabolism, Stress and Developmental Programming. Mol. Cell. Endocrinol. 2022, 542, 111522. [Google Scholar] [CrossRef] [PubMed]
- Black, N.; Stockings, E.; Campbell, G.; Tran, L.T.; Zagic, D.; Hall, W.D.; Farrell, M.; Degenhardt, L. Cannabinoids for the Treatment of Mental Disorders and Symptoms of Mental Disorders: A Systematic Review and Meta-Analysis. Lancet Psychiatry 2019, 6, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Surkin, P.N.; Gallino, S.L.; Luce, V.; Correa, F.; Solari, J.F.; De Laurentiis, A. Pharmacological Augmentation of Endocannabinoid Signaling Reduces the Neuroendocrine Response to Stress. Psychoneuroendocrinology 2018, 87, 131–140. [Google Scholar] [CrossRef]
- Jiang, S.; Zheng, C.; Wen, G.; Bu, B.; Zhao, S.; Xu, X. Down-Regulation of NR2B Receptors Contributes to the Analgesic and Antianxiety Effects of Enriched Environment Mediated by Endocannabinoid System in the Inflammatory Pain Mice. Behav. Brain Res. 2022, 435, 114062. [Google Scholar] [CrossRef] [PubMed]
- Dandi, E.; Spandou, E.; Tata, D.A. Investigating the Role of Environmental Enrichment Initiated in Adolescence against the Detrimental Effects of Chronic Unpredictable Stress in Adulthood: Sex-Specific Differences in Behavioral and Neuroendocrinological Findings. Behav. Process. 2022, 200, 104707. [Google Scholar] [CrossRef]
- Dandi, Ε.; Theotokis, P.; Petri, M.C.; Sideropoulou, V.; Spandou, E.; Tata, D.A. Environmental Enrichment Initiated in Adolescence Restores the Reduced Expression of Synaptophysin and GFAP in the Hippocampus of Chronically Stressed Rats in a Sex-Specific Manner. Dev. Psychobiol. 2023, 65, e22422. [Google Scholar] [CrossRef]
- Dandi, Ε.; Kalamari, A.; Touloumi, O.; Lagoudaki, R.; Nousiopoulou, E.; Simeonidou, C.; Spandou, E.; Tata, D.A. Beneficial Effects of Environmental Enrichment on Behavior, Stress Reactivity and Synaptophysin/BDNF Expression in Hippocampus Following Early Life Stress. Int. J. Dev. Neurosci. 2018, 67, 19–32. [Google Scholar] [CrossRef]
- Kempermann, G. Environmental Enrichment, New Neurons and the Neurobiology of Individuality. Nat. Rev. Neurosci. 2019, 20, 235–245. [Google Scholar] [CrossRef]
- Simpson, J.; Kelly, J.P. The Impact of Environmental Enrichment in Laboratory Rats--Behavioural and Neurochemical Aspects. Behav. Brain Res. 2011, 222, 246–264. [Google Scholar] [CrossRef] [PubMed]
- Vaquero-Rodríguez, A.; Ortuzar, N.; Lafuente, J.V.; Bengoetxea, H. Enriched Environment as a Nonpharmacological Neuroprotective Strategy. Exp. Biol. Med. 2023, 248, 553–560. [Google Scholar] [CrossRef] [PubMed]
- El Rawas, R.; Thiriet, N.; Nader, J.; Lardeux, V.; Jaber, M.; Solinas, M. Early Exposure to Environmental Enrichment Alters the Expression of Genes of the Endocannabinoid System. Brain Res. 2011, 1390, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Banaei-Boroujeni, G.; Rezayof, A.; Alijanpour, S.; Nazari-Serenjeh, F. Targeting Mediodorsal Thalamic CB1 Receptors to Inhibit Dextromethorphan-Induced Anxiety/Exploratory-Related Behaviors in Rats: The Post-Weaning Effect of Exercise and Enriched Environment on Adulthood Anxiety. J. Psychiatr. Res. 2023, 157, 212–222. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C. The Rat Brain in Stereotaxic Coordinates, 6th ed.; Elevier: Boston, MA, USA, 2007. [Google Scholar]
- Tannenbaum, C.; Ellis, R.P.; Eyssel, F.; Zou, J.; Schiebinger, L. Sex and Gender Analysis Improves Science and Engineering. Nature 2019, 575, 137–146. [Google Scholar] [CrossRef] [PubMed]
- Howell, G.T.; Lacroix, G.L. Decomposing Interactions Using GLM in Combination with the COMPARE, LMATRIX and MMATRIX Subcommands in SPSS. Tutor. Quant. Methods Psychol. 2012, 8, 1–22. [Google Scholar] [CrossRef]
- Coelho, A.A.; Lima-Bastos, S.; Gobira, P.H.; Lisboa, S.F. Endocannabinoid Signaling and Epigenetics Modifications in the Neurobiology of Stress-Related Disorders. Neuronal Signal. 2023, 7, NS20220034. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.W.; Zhou, C.H.; Xue, S.S.; Yu, H.; Shi, Q.Q.; Xue, F.; Chen, Y.H.; Tan, Q.R.; Wang, H.N. High-Frequency Repetitive Transcranial Magnetic Stimulation Regulates Neural Oscillations of the Hippocampus and Prefrontal Cortex in Mice by Modulating Endocannabinoid Signalling. J. Affect. Disord. 2023, 331, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Stachowicz, K. Deciphering the Mechanisms of Reciprocal Regulation or Interdependence at the Cannabinoid CB1 Receptors and Cyclooxygenase-2 Level: Effects on Mood, Cognitive Implications, and Synaptic Signaling. Neurosci. Biobehav. Rev. 2023, 155, 105439. [Google Scholar] [CrossRef]
- Christensen, R.; Kristensen, P.K.; Bartels, E.M.; Bliddal, H.; Astrup, A. Efficacy and Safety of the Weight-Loss Drug Rimonabant: A Meta-Analysis of Randomised Trials. Lancet 2007, 370, 1706–1713. [Google Scholar] [CrossRef]
- Romero-Torres, B.M.; Alvarado-Ramírez, Y.A.; Duran-Alonzo, S.R.; Ruiz-Contreras, A.E.; Herrera-Solis, A.; Amancio-Belmont, O.; Prospéro-García, O.E.; Méndez-Díaz, M. A Potential Role of Hippocampus on Impulsivity and Alcohol Consumption Though CB1R. Pharmacol. Biochem. Behav. 2023, 225, 173558. [Google Scholar] [CrossRef]
- Prospéro-García, O.; Ruiz Contreras, A.E.; Ortega Gómez, A.; Herrera-Solís, A.; Méndez-Díaz, M. Endocannabinoids as Therapeutic Targets. Arch. Med. Res. 2019, 50, 518–526. [Google Scholar] [CrossRef] [PubMed]
- Alijanpour, S.; Rezayof, A. Activation of Ventral Hippocampal CB1 Receptors Inhibits Ketamine-Induced Anxiogenic-like Behavior: Alteration of BDNF/c-Fos Levels in the Mouse Hippocampus. Brain Res. 2023, 1810, 148378. [Google Scholar] [CrossRef]
- Gabriel, P.; Mastracchio, T.A.; Bordner, K.; Jeffrey, R. Impact of Enriched Environment during Adolescence on Adult Social Behavior, Hippocampal Synaptic Density and Dopamine D2 Receptor Expression in Rats. Physiol. Behav. 2020, 226, 113133. [Google Scholar] [CrossRef]
- Joushi, S.; Esmaeilpour, K.; Masoumi-Ardakani, Y.; Esmaeili-Mahani, S.; Sheibani, V. Effects of Short Environmental Enrichment on Early-Life Adversity Induced Cognitive Alternations in Adolescent Rats. J. Neurosci. Res. 2021, 99, 3373–3391. [Google Scholar] [CrossRef] [PubMed]
- Lookfong, N.A.; Raup-Konsavage, W.M.; Silberman, Y. Potential Utility of Cannabidiol in Stress-Related Disorders. Cannabis Cannabinoid Res. 2023, 8, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Heng, L.; Beverley, J.A.; Steiner, H.; Tseng, K.Y. Differential Developmental Trajectories for CB1 Cannabinoid Receptor Expression in Limbic/Associative and Sensorimotor Cortical Areas. Synapse 2011, 65, 278–286. [Google Scholar] [CrossRef]
- Spiacci, G.B.L.; Antero, L.S.; Reis, D.G.; Lisboa, S.F.; Resstel, L.B. Dorsal Hippocampus Cannabinoid Type 1 Receptors Modulate the Expression of Contextual Fear Conditioning in Rats: Involvement of Local Glutamatergic/Nitrergic and GABAergic Neurotransmissions. Eur. Neuropsychopharmacol. 2016, 26, 1579–1589. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dandi, E.; Kesidou, E.; Simeonidou, C.; Spandou, E.; Grigoriadis, N.; Tata, D.A. Sex-Specific Differences and the Role of Environmental Enrichment in the Expression of Hippocampal CB1 Receptors following Chronic Unpredictable Stress. Brain Sci. 2024, 14, 357. https://doi.org/10.3390/brainsci14040357
Dandi E, Kesidou E, Simeonidou C, Spandou E, Grigoriadis N, Tata DA. Sex-Specific Differences and the Role of Environmental Enrichment in the Expression of Hippocampal CB1 Receptors following Chronic Unpredictable Stress. Brain Sciences. 2024; 14(4):357. https://doi.org/10.3390/brainsci14040357
Chicago/Turabian StyleDandi, Evgenia, Evangelia Kesidou, Constantina Simeonidou, Evangelia Spandou, Nikolaos Grigoriadis, and Despina A. Tata. 2024. "Sex-Specific Differences and the Role of Environmental Enrichment in the Expression of Hippocampal CB1 Receptors following Chronic Unpredictable Stress" Brain Sciences 14, no. 4: 357. https://doi.org/10.3390/brainsci14040357